Abstract
The diversity of Coleoptera communities in tussock grassland at two sites in Otago, one in Canterbury and one in the central North Island, was compared. The impact of agricultural disturbance on the communities was compared between native tussock, tussock oversown with exotic pasture species and cultivated sown pasture. Coleoptera were heat-extracted from turf samples taken in summer in two consecutive years. In native and oversown treatments, Staphylinidae and Curculionidae predominated in abundance at three of the four sites, and in species richness at all sites and treatments. Carabidae were more species-rich than Curculionidae in cultivated treatments at most sites. The mean density of Coleoptera in native tussock treatments ranged between 654/m2 and 97/m2. Carnivores were the predominant trophic group followed by herbivores. Species diversity was higher in the native or oversown treatments and the Otago sites were the most diverse overall, and the least disturbed. A total of 19 exotic species were found but there was no relationship with treatment, although their density was higher in cultivated treatments. There was no evidence to suggest that modified vegetation provides a source of exotic Coleoptera species to invade native tussock.
Introduction
New Zealand is globally recognized as a ‘hot spot’ of indigenous biodiversity (Myers et al. Citation2000) and invertebrates are a major component of this. Nevertheless, the community composition and ecology of the invertebrate fauna of indigenous ecosystems are in general poorly known, including those of tussock grasslands.Footnote Some exceptions are tussock grassland sites in Otago, Canterbury and the Central North Island that have been well studied recently. For example, Barratt et al. (Citation2005) analysed data for the macro-invertebrate groups at order level or above; Bell et al. (Citation2005) recorded the diversity and trophic structure of nematodes; Sarathchandra et al. (Citation2005) and di Menna et al. (Citation2007) investigated the composition, abundance and functional diversity of soil micro-organisms; Espie & Barratt (Citation2006) investigated plant biodiversity in relation to aspects of disturbance by agricultural development and fire.
Since European colonization of New Zealand, native tussock grasslands have been modified by grazing, burning and introduction of exotic pasture species (Floate Citation1992). In the mid-altitude range (up to about 1000 m a.s.l) exotic grasses and clover have been introduced into native grasslands, often with superphosphate fertilizer either by aerial oversowing to increase stock-carrying capacity, or by cultivation to replace the existing vegetation with sown exotic pasture species (Douglas & Allan Citation1992). Fire has also been used extensively to produce palatable regrowth from tussock species, or in preparation for exotic pasture species introduction (Lowther & Douglas Citation1992). To investigate the impacts of such anthropogenic disturbances on natural grassland communities a basic understanding of their taxonomic and functional characteristics is required. Such information will improve our understanding of the effect of habitat disturbance on invertebrate communities, and the attributes that confer resilience of indigenous systems to invasion by exotic species.
The risk to native ecosystems from invasive alien species is poorly understood, especially for invertebrates (Brockerhoff et al. Citation2010). It has been noted that in comparison with other countries, the impact of exotic invertebrates on New Zealand's natural ecosystems appears to be relatively minor, especially in comparison with impacts on productive systems (Lowe Citation1973; Atkinson & Cameron Citation1993; Ridley et al. Citation2000). However, it is acknowledged that natural ecosystems in New Zealand have received relatively little attention in this regard. Brockerhoff et al. (2010) concluded that there was some support for the hypothesis that some New Zealand native plants exhibit resistance to exotic herbivorous invertebrates and that some predatory exotic invertebrates have colonized natural ecosystems and readily prey on native species. They found that closed forest ecosystems are less susceptible to exotic invertebrates than open habitats such as grassland and that habitat disturbance increases the level of invasibility.
In this study we have collected quantitative data to provide an indication of relative abundance of invertebrate groups, and hence information on the structure of communities and trophic relationships. Coleoptera are acknowledged as a good surrogate for the invertebrate fauna because they are represented by all major trophic groups; it is one of the most species-rich orders and is reasonably well known taxonomically (Hutcheson Citation1990; New Citation2007). Our knowledge of the coleopteran fauna of tussock grasslands is by no means comprehensive, but there have been some significant contributions in recent years (e.g. Barratt & Patrick Citation1987; Barratt & Kuschel Citation1996; Dickinson et al. Citation1998; Derraik et al. Citation2001; Murray Citation2001; Rufaut Citation2002; Derraik et al. Citation2003; Melo et al. Citation2003; Murray et al. Citation2003, Citation2006a, b; Patrick Citation2004; Barratt et al. Citation2006,Citation2009).
The objectives of this study are to compare the diversity of Coleoptera communities in native tussock grassland at four sites, two in Otago, one in Canterbury and one in the North Island Central Plateau, and to determine the impact of disturbance at three levels of agricultural development on those communities. We also investigate whether exotic species invade natural ecosystems after they have established in modified environments. We examine the exotic component of the Coleoptera fauna in these native and disturbed communities to determine whether the source of exotic species is nearby disturbed environments such as oversown tussock or cultivated pasture.
Methods
Sites
A description of the native tussock grassland sites at Mount Benger and Deep Stream (Otago), Cass (Canterbury) and Tukino (central North Island) is given in Barratt et al. (2005) and summarized here in . In addition to the native tussock vegetation, oversown tussock and pasture cultivated from tussock were also selected for sampling at each site. These will be described as treatments. Although the oversown and cultivated treatments were located as near as possible to the native tussock treatment, there was often a difference in altitude () especially at Mt Benger. The greatest distance between the native and developed treatments was at Tukino, where they were 29 km apart ().
Table 1 Location, land tenure, rainfall for study sites and treatments, and sampling dates.
Sampling
All sites were sampled once between January and March in summer 2003 and 2004 (). The Otago native tussock treatments (Deep Stream and Mount Benger) were sampled by taking 20 turf samples from the inter-tussock vegetation and nine turf samples over tussock plants from each of three replicate 20×20 m plots. Sample position was selected by throwing a quadrat within the plot along four parallel 5 m transects. The replicated plots at the Otago sites were within a radius of 1 km. All samples were 0.1 m2 in area and 5 cm deep. At the other two sites and from all oversown tussock and cultivated plots a single replicate of 20 inter-tussock samples was taken along an approximately straight line transect with approximately 5 m between samples (selected by throwing a quadrat), on the dates shown in . Turf samples were individually bagged in the field and stored at 4°C until extraction. In the laboratory invertebrates were extracted from each individual turf over a 7-day period using heat extractors (Crook et al. Citation2004). After extraction from turves, the invertebrates were stored in 70% ethanol at 4°C until identification.
Samples were washed through fine muslin which retained all invertebrates but allowed fine silt to pass through. The ‘cleared’ samples were then sorted and invertebrates counted under a low-power binocular microscope.
Identification
Coleoptera were identified to species as far as possible, or to morphospecies. Coleoptera larvae were identified to family but identification beyond this level was rarely possible. In the Coleoptera species analyses, larval morphospecies were included as separate taxa even though it was likely to over-estimate species numbers. We did this because it is impossible in most cases to match larvae with adults, and allows for the fact that larvae and adults are sometimes in different trophic groups, occupying a different niche and in general performing a different function in the community. The native/exotic status is given for each species/ morphospecies using Leschen et al. (Citation2003) for guidance, although information on native/exotic status was not available at the species level. Trophic status was determined as far as possible using Waterhouse (Citation1973), relevant ‘Fauna of New Zealand’ volumes or other reliable internet sources. Categories used were carnivores, herbivores, fungivores, saprophytes and carrion feeders. For analysis, the last two were combined with fungivores since their incidence and abundance was very low.
Statistical methods
Means for taxonomic groups, densities and functional groups were calculated using replicate plots and years (n=6) for the native tussock plots at Deep Stream and Mount Benger. For the oversown and cultivated plots at Deep Stream and Mount Benger, and all plots at Cass and Tukino, they were calculated as means for the two years (n=2).
Means for each of the three treatments (native, oversown and cultivated) were calculated for the entire study period for each of the four sites and formed the basis for the following analysis in Primer v5 (Clarke & Gorley Citation2001). The Shannon Wiener, Simpson evenness (1−λ) and Margalef diversity indices were calculated to compare species diversity between sites and treatments. K-dominance curves were constructed to provide additional information on the community structure for sites and treatments.
Species similarities between sites and treatments were examined using a cluster analysis of the samples based on Bray-Curtis similarities with fourth root transformation. Transformation of the data lessens the contribution of dominant species and gives more weight to less-abundant species that are of interest in the similarity analysis.
Species accumulation curves for the replicated native grassland treatments at Deep Stream and Mount Benger were calculated using EstimateS (Colwell Citation2006). The curves were calculated with 50 randomizations without sample replacement. The programme was also used to calculate estimates of total species richness ACE (Abundance-based Coverage Estimator) and Chao1. Both estimators give minimum estimates of species richness based on the abundance of rare species with fewer than 10 individuals or one and two individuals only (Chao1).
To assess the impact of disturbance on community structure, distance from lognormal was calculated using Chi2/n as outlined by Halloy and Barratt (Citation2007). Numbers of individuals per species including larvae were ranked in classes (0.5, 1.5, 3.5, 7.5, …) bounded by zeros at both ends. The lognormal was fitted to the data using the Fortran program LOGNORM, Version 2.0 (Krebs Citation1989) following the method of Cohen (Citation1961).
Results
Taxonomic composition and species richness
Coleoptera ranked second most abundant invertebrate taxon on average (after Hymenoptera, mainly Formicidae) across the four sites in native tussock grassland (Barratt et al. Citation2005). However, as a proportion of total density Coleoptera density constituted an average of 4.4%, 12.6%, 20.9% and 2.9% at Deep Stream, Mount Benger, Cass and Tukino respectively (from in Barratt et al. Citation2005).
A summary of data characterizing the Coleoptera fauna (including larval morphospecies) at the sites and treatments is shown in . Twenty-seven Coleoptera families were recorded at the four sites, ranging from 25 at Mount Benger to 13 at Cass. The families best represented in terms of abundance were Staphylinidae and Curculionidae at all sites except at Cass, where the second most abundant family was Cantharidae (). The density of Coleoptera at Tukino was low compared with the other sites. At Mount Benger, the fourth most abundant family was Ptiliidae, a family that was only represented in samples at one other site, Tukino. Appendix 1 shows the list of Coleoptera species (excluding larval morphospecies) combined for all four sites. The number of species in each family followed a pattern similar to abundance (). Staphylinidae were the most speciose family at all sites followed by Curculionidae.
Figure 1 Total density (n/m2) of the 15 most speciose families from each site shown for each treatment in order of abundance in the native tussock treatments.
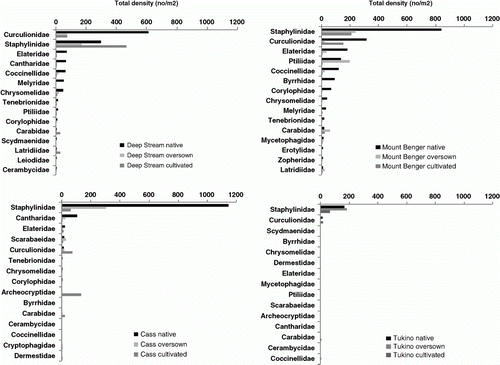
Figure 2 Number of species in the 15 most speciose families from each site shown for each treatment plotted in order of abundance in the native tussock treatments.
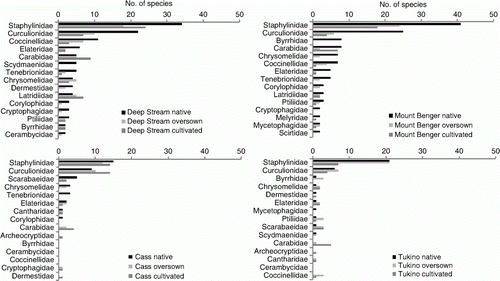
Table 2 Summary of Coleoptera families, species richness (S), density, total and exotic species found at each site and vegetation type, and percentage of species in three main trophic groups.
At both Deep Stream and Mount Benger 16 Coleoptera families were represented, with successively fewer in the oversown treatments and cultivated pastures. At Cass fewer families were recorded and the number was similar at all three vegetation treatments. At Tukino, the highest numbers of Coleoptera families was recorded at the oversown tussock treatment.
Including larvae, a total of 321 Coleoptera species/morphospecies was recorded during the study and species richness (S) ranged from 170 at Deep Stream to 77 at Cass, with 16 species common to all four sites. Within sites, S was higher in the native tussock treatments than the oversown or cultivated treatments, except at Tukino, where S in the oversown tussock treatment was highest. Between about 5% and 15% of species were found in all three vegetation types at each site. Deep Stream had the highest number of species (62) that were unique to a site ().
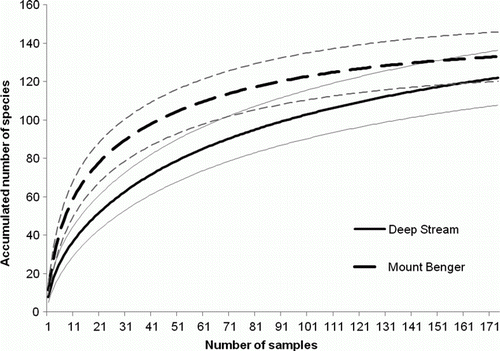
Species accumulation curves for Deep Stream and Mount Benger are shown in . Analysis of 174 samples (the total taken for both years) calculated a mean accumulated species richness of 122 for Deep Stream and 133 for Mount Benger. The estimates for total species richness predicted 149 (ACE) and 144 (Chao1) species at Deep Stream and 138 (ACE) and 136 (Chao1) species at Mount Benger. This indicates that the sampling at the Mount Benger site captured a larger proportion of predicted total number of species than at Deep Stream (97% and 82% respectively). Furthermore, to reach 50% of the predicted estimate (ACE) of species richness required 44 samples at Deep Stream but only 15 at Mount Benger. At Cass and Tukino, and all oversown and cultivated treatments, 40 samples were taken in total during the study. With reference to the Deep Stream and Mount Benger species accumulation curves, 40 samples would have resulted in about 48% and 71% of total estimated species at these sites respectively. This therefore provides an indication of the number of species that might be expected at Cass and Tukino native treatments, and oversown and cultivated treatments at all sites.
Figure 3 Coleoptera species accumulation curves for Deep Stream and Mount Benger, showing 95% confidence limits (fine solid and dashed lines).
Density and trophic composition
The mean density of Coleoptera in native tussock was highest at Cass (654/m2) and lowest at Tukino (97/m2). In oversown tussock Coleoptera densities were generally highest at Mount Benger and lowest at Tukino. In cultivated pasture Coleoptera densities were highest at Deep Stream and lowest at Tukino (). Mean density of Coleoptera per taxon (mean density/S) was below 9 for all treatments except the native tussock treatment at Cass, where it was 23.8. Here the density of Aleocharine Cass sp. 2 (Staphylinidae) was over 300/m2 in both years of the study, and Aleocharine Cass sp. 1 was over 120/m2 in each year.
At Mount Benger, Deep Stream and Tukino, carnivores were the predominant component of the Coleoptera fauna. Herbivores were the next most prominent group, followed by fungivores, which were represented by about 12%–13% of the species. At Cass, however, herbivores comprised about half of the species present and only 3%–9% of species were fungivores/saprophytes (). The carnivore component of species composition tended to be higher in cultivated pasture compared with native tussock, except at Tukino, where the reverse was true. Furthermore, at this site the proportion of species which were fungivores/saprophytes, comprising mainly Corylophidae, was considerably higher in the oversown treatment compared with the other treatments where this family was not recorded ().
Species diversity
Three commonly used indices for species diversity are shown in . The Shannon-Wiener (H′) index, which is sensitive to rare species in the community, was higher in the native and oversown treatments compared with the cultivated treatments at all sites except Cass, although the index for oversown tussock grassland was higher than native tussock at both Deep Stream and Tukino. The Simpson index of diversity (1−λ) emphasizes the dominant species in a community, where 0 equates to low diversity, and 1 denotes that all individuals belong to different species. Again at both Deep Stream and Tukino, the highest values occurred in the oversown treatments. At the native treatment at Cass, the low value suggests that the community is dominated by a few species.In this case four species of aleocharines (Staphylinidae: Aleocharinae) make up over 40% of the total abundance in the community.
Table 3 Species diversity indices for each site and treatment.
The Margalef index (d) is a simple function of species richness and abundance; this gave a range of indices which were relatively high for the native treatments at the Otago sites compared with the disturbed treatments. However, Cass values were very similar, and at Tukino, as for all the indices, the oversown treatments had the highest value because species richness was greater than at the native and cultivated treatments ().
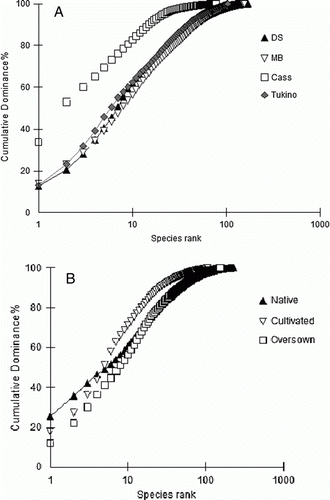
The k-dominance curves for sites (a) and treatments (b) provide a further indication of comparative species diversity. The lower the curves on the plot, the more diverse are the species assemblages. The k-dominance curves indicate that species diversity is much lower at Cass than at the other sites, and that Deep Stream and Mount Benger are the most diverse overall (a). Oversown treatments appeared to be more species diverse than native and cultivated treatments, supporting results calculated for the Shannon-Wiener and Simpson indices (b).
Figure 4 Rank-abundance curves for A, sites and B, treatments.
Similarity between sites and treatments
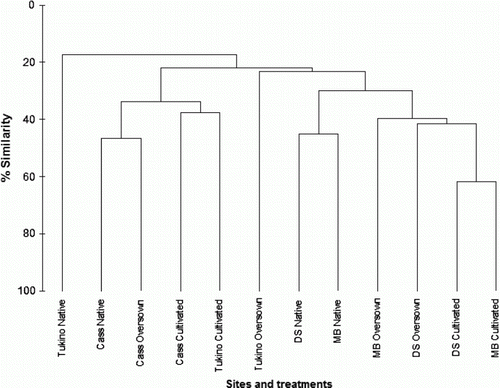
Bray-Curtis similarities plotted as a dendrogram are shown in , showing clear separation between the Cass, Tukino and Otago sites. The Tukino native vegetation stood apart from most other treatments, with only 20% similarity in species composition with any of the other treatments. Cultivated treatments at Deep Stream and Mount Benger aligned closely together, sharing about 60% of species. Similarly the native and oversown treatments at the Otago sites clustered together and shared about 40% of species. The treatments at Cass and Tukino clustered together, but separately from the treatments at the Otago sites. The three Cass treatments clustered together, with 40%–50% similarity, but the Tukino treatments showed little similarity in species composition.
Figure 5 Bray-Curtis cluster analysis for sites and treatments.
Disturbance
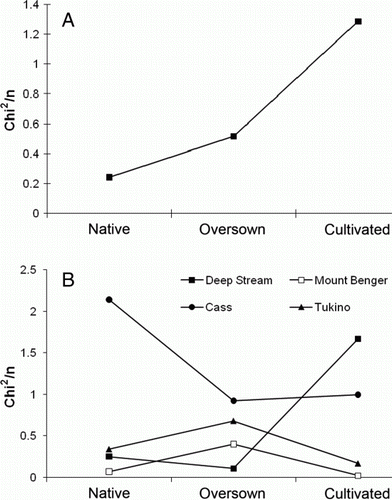
Analysis of distance from a lognormal distribution, measured as Chi2/n (Halloy & Barratt Citation2007), showed that when treatments were combined for all sites, cultivated treatments were more distant from a log-normal distribution compared with the native and oversown treatments (a). The native treatments were close to the lognormal, except for Cass (b). However, as mentioned above the previous vegetation analysis of the Cass native site suggested a history of disturbance and predominance of exotic grass species, so the analysis was repeated excluding the Cass treatments. The resulting relationship shows a strong positive relationship between distance from lognormal and increasing disturbance. At Deep Stream (b), χ2/n for the cultivated treatment was more distant from a lognormal distribution compared with the native and oversown sites, but at Mount Benger and Tukino the cultivated treatments were quite close to lognormal.
Figure 6 Values for distance from lognormal fitted for A, mean of all sites excluding Cass for each vegetation treatment; B, each site and treatment. The closer χ2/n is to zero, the better the fit to a lognormal distribution, and hence the least disturbed as indicated by the distribution of Coleoptera species abundance across species.
Exotic species
A total of 19 exotic species were found at the four sites; the highest number at Deep Stream (11), but the highest percentage at Cass, although the range for all locations was narrow (6.5%–7.8%) ().
The exotic Coleoptera species are indicated in Appendix 1, and shows the mean number and density of the species at each locality and treatment. The mean number of exotic species per sample was quite low (range 1.2–4 species), and relatively consistent between the sites and treatments, with no significant differences evident. However, density of exotic species was consistently higher in the cultivated treatments compared with the native and oversown treatments (b). At Tukino no exotic Coleoptera were identified in samples from the native treatment.
Figure 7 Mean number (±SE) A, and density (±SE) B, of exotic Coleoptera species in the three vegetation types at each locality; mean density per taxon (±SE) C, of native and exotic Coleoptera in each vegetation type for all four sites.
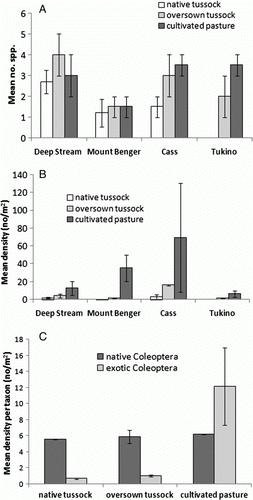
Comparing the mean density of each taxon of natives with exotics for all sites combined, the number of native taxa is quite consistent in each of the vegetation treatments, whereas the density of each exotic taxon is significantly higher in the cultivated treatments compared with the native tussock and oversown tussock treatments (c).
The only exotic species that was found at all four sites was Listronotus bonariensis (Kuschel), present at cultivated treatments at each locality, also from oversown tussock at DS and Cass, and from native tussock at Cass. For all sites combined, mean density of this species in native, oversown tussock and cultivated pasture was 0.73/m2, 1.79/m2 and 13.8/m2 respectively.
summarizes where exotic Coleoptera were found in relation to vegetation type, but this shows few patterns or consistencies with respect to family, trophic group or origin. Curculionidae were the most commonly represented family with five species.
Table 4 Distribution of exotic Coleoptera species between vegetation treatments.
Discussion
The predominant coleopteran families with the highest recorded number of species were the Staphylinidae and Curculionidae at all four sites. This was also found to be the case at 1600 m a.s.l. on the Old Man Range, close to Mount Benger (Scott Citation2007). The exception was Cass, where the density of Curculionidae was comparatively low, although the number of species was relatively high. Tukino stood out as a site where Coleoptera density was low, but the number of species in each family was only a little lower than for the other sites. Cass was completely dominated by Staphylinidae in terms of density, but the number of species was relatively low. There are few studies internationally which have employed quantitative sampling in grassland as described here; however, a comparable study of the coleopteran fauna of calcareous grasslands in the UK where turf-sampling was used found that the most abundant and species-rich family was Staphylinidae, followed by Carabidae and Curculionidae (Morris & Rispin Citation1987).
It is important for this study to take into account the different sampling effort at the sites and treatments when interpreting species richness and species diversity data. At the two Otago sites, where there was replication of the native treatments, higher values for these variables might be expected than at the Cass and Tukino sites, where a single replicate sample was taken. The species accumulation curves suggested that the sampling effort at Deep Stream and Mount Benger native treatments collected a large proportion of species present (especially at Mount Benger) at the time of year when samples were taken. The more limited sampling at Cass and Tukino, and the oversown and cultivated treatments at all sites, was estimated to have recovered between 48% and 71% of species, assuming that species accumulation curves would be similar to those calculated for Deep Stream and Mount Benger. Estimates of density and trophic structure have not been compromised by the variable sampling effort, although replication reduced variability.
At the Otago sites, both tussock and inter-tussock samples were taken from native treatments, whereas this was not the case at Cass, where Chionochloa was not a dominant grass species (Espie & Barratt Citation2006) and at Tukino. Previous work has shown that species richness and species diversity in tussock and inter-tussock samples from the Otago sites was very similar for the two years of the current study, if anything slightly lower in tussock samples at both sites (Barratt et al. Citation2009). The trophic structure for tussock and intertussock samples was very similar for numbers of taxa at both sites, but the proportion of carnivores was relatively higher than herbivores in tussock vs. inter-tussock samples at both Otago sites (Barratt et al. Citation2009).
Of the native tussock treatments, species diversity indices were highest at Mount Benger and lowest at Cass. At Mount Benger the native grasses (Chionochloa rigida and Poa colensoi) occupied 76% of ground cover (Espie & Barratt Citation2006). Cass, however, differed from the other sites in that the vegetation was dominated by the introduced grasses Agrostis capillaris and Anthoxanthum odoratum with a combined cover of 43% (Espie & Barratt Citation2006). The native grass Festuca novae-zelandiae was present but only at 5% cover. These authors suggested that this induced short tussock community at Cass might be attributable to a more frequent history of burning, and greater intensity of rabbit and stock grazing pressure than experienced at the other sites. This is supported by the greater value of ΔL(Chi2/n) for the native vegetation at Cass compared with the other three sites.
The similarity of Coleoptera community composition at the study sites and treatments indicated by the cluster analysis reached up to 60% (the two Otago cultivated treatments). The Tukino native vegetation treatment separated out from all other treatments with less than 20% similarity, possibly because of the relatively low number of species recorded, and low density of Coleoptera. This North Island site was a considerable distance (geographically) from the other South Island sites, with expected differences in faunal composition. The macro-invertebrate fauna at Tukino was heavily dominated by ants, which constituted 88% of the total density (Barratt et al. Citation2005). This may have impacted on other elements of the fauna, including Coleoptera, possibly by competition for resources (e.g. seeds) or predation. The Cass and Tukino sites separated out from the Otago sites in the cluster analysis as might be expected, except for the Tukino oversown treatment, which clustered with the Otago sites. The native tussock treatments at the Otago sites showed about 45% similarity in Coleoptera community composition. In comparison with the Coleoptera, the vegetation analysis for these treatments separated out eight groups at the 70% level (Espie & Barratt Citation2006), which is probably consistent with the lower number of species.
Many studies have shown that stable, undisturbed environments tend to have a few abundant species and many less abundant and rare species, a pattern that fits a lognormal distribution (e.g. Brown Citation1995). It follows, then, that departure from this pattern should indicate disturbance from the lognormal pattern. Halloy & Barratt (Citation2007) suggested a consistent method for measuring departure from a lognormal pattern, which is ΔL. This method of pattern analysis can be used for a number of characteristics of organisms where there is competition for resources, e.g. leaf area for plants, size of invertebrates, biomass of organisms, etc. Here we have calculated ΔL for abundance of Coleoptera species, and when combined for sites it showed an expected departure from the lognormal with increasing levels of disturbance from native tussock to cultivated treatments. When the treatments at each locality were examined separately, however, only at Deep Stream did the ΔL values closely fit this hypothesis. There are a number of reasons why at the other sites the data did not fit the expected pattern, including inadequate sampling (e.g. Cass and Tukino), and the possibility that the ‘disturbed’ treatments have actually recovered over time to a stable state even though species composition might be different, so that the fit to lognormal has re-established. So fit to a lognormal distribution should not be confused with ‘naturalness’.
The relatively few exotic Coleoptera species (19) found during this study have made it difficult to demonstrate any pattern either taxonomically, functionally or in area of origin of those species, except that all herbivores were in the family Curculionoidea. Four of the weevil species are recognized pests in pasture and forage systems, and one is an introduced biological control agent (Trichosirocalus horridus (Panzer) of Carduus nutans L.) (nodding thistle). The low numbers of exotic Coleoptera provided little opportunity to test the hypothesis that exotic Coleoptera which have established in native tussock have invaded from nearby areas of oversown or cultivated pasture. Only four of the 12 species found in native tussock were also found in cultivated pasture, and a further three were found in oversown tussock, leaving five that were only found in native tussock.
The most ‘successful’ coleopteran invader of native tussock was the graminaceous stem-boring pest, Listronotus bonariensis (Kuschel) (Argentine stem weevil), which was found at all locations and in most vegetation types. This weevil is a major pest of ryegrass in New Zealand (Prestidge et al. Citation1991) and, being ubiquitous in pasture throughout the country, has probably moved into tussock grassland from nearby populations. The extent to which this species represents a threat in native tussock is unclear, but the densities recorded in this study, unlike those found in lower-altitude pastures, would not suggest a major impact. However, it has been shown to be capable of ovipositing and feeding on Festuca novae-zelandiae and Poa cita (Lister Citation2006).
Other exotic species found during this study which have become pests in pasture and forage systems include Sitona discoideus Gyllenhal (lucerne weevil) and the more recently established species, Sitona lepidus Gyllenhal (clover root weevil). These species were only found at cultivated treatments in this study, but both are strong fliers and S. discoideus has been found in native tussock in other studies (Barratt et al. Citation2007). Establishment in native tussock would be dependent upon the presence of their host plants (Medicago sativa and Trifolium repens respectively). The latter is a common component of oversown pastures and can be transferred to native tussock with stock movement and hence S. lepidus has potential to become established in oversown and possibly native tussock. Otiorhynchus ovatus (strawberry weevil) has quite a broad plant host range, and is likely to include native plant species in its diet. T. horridus (crown weevil) was found in native tussock at Mount Benger, but establishment of this species in natural ecosystems will be dependent upon the presence of its plant host, Carduus species, to which it would be restricted. It is likely that more of the species in the taxonomically poorly known groups such as Staphylinidae are exotic but have not been recognized as such during this study.
Conclusions
The study has shown that the taxonomic structures of Coleoptera in the four native tussock grassland communities included in this study were quite similar, with Staphylinidae and Curculionidae consistently predominating in abundance and species richness. Consequentially, the trophic structure reflects this with carnivores and herbivores making up 70%–80% of species.
Habitat disturbance from agricultural activity has resulted in changes to Coleoptera community structure and diversity, particularly after soil disturbance from cultivation, and the development of simplified, exotic plant communities. With one exception (Cass), species diversity was reduced in cultivated pastures in comparison with native grassland, while at two of the four sites species diversity indices often indicated higher values for oversown tussock. The lognormal analysis, however, as another indicator of disturbance, showed, for three of the sites combined, a progressive increase in distance to lognormal from native through oversown to cultivated grassland.
The number of exotic Coleoptera species found during the study at each site was quite low, and for most species densities were also very low, although in cultivated pasture the mean density of exotic species was higher than that of native species, unlike native and oversown tussock. However, there was little evidence to support or reject the hypothesis that undisturbed habitats are resilient to exotic species given the low number of exotic Coleoptera species established for all treatments, or that modified vegetation provides a source of exotic Coleoptera species to invade native tussock.
Acknowledgements
This work was funded by New Zealand's Foundation for Research, Science & Technology through contract C02X0501, the Better Border Biosecurity (B3) programme (www.b3nz.org), and by the Department of Conservation (Science Investigation No. 3667). We thank the many AgResearch staff and students for assistance with sorting samples, especially Rory Logan, Tara Murray, Peter Tozer and Aliesha Kean. We thank Marcus Simons, Don Newman (DoC) and Kath Dickinson (University of Otago) for their continuing support for the study. We thank Al Newton (Field Museum of Natural History, Chicago) for advice on Staphylinidae. We are particularly grateful to Dr Stephan Halloy (Conservation International, Bolivia) for very helpful comments on an earlier draft of this paper.
Notes
This version has been amended. Please see Erratum (http://dx.doi.org/10.1080/03036758.2012.754325).
References
- Atkinson , IAE and Cameron , EK . 1993 . Human influence on the terrestrial biota and biotic communities of New Zealand . Trends in Ecology & Evolution , 8 : 447 – 451 . doi: 10.1016/0169-5347(93)90008-D
- Barratt , BIP and Kuschel , G . 1996 . Broad-nosed weevils (Curculionidae: Brachycerinae: Entimini) of the Lammermoor and Rock and Pillar Ranges in Otago, with descriptions of four new species of Irenimus . New Zealand Journal of Zoology , 23 : 359 – 374 . doi: 10.1080/03014223.1996.9518096
- Barratt , BIP and Patrick , BH . 1987 . Insects of snow tussock grassland on the East Otago Plateau . New Zealand Entomologist , 10 : 69 – 98 . doi: 10.1080/00779962.1987.9722513
- Barratt , BIP , Ferguson , CM , Logan , RAS , Barton , D , Bell , NL , Sarathchandra , SU and Townsend , RJ . 2005 . Biodiversity of indigenous tussock grassland sites in Otago, Canterbury and the Central North Island of New Zealand I. The macro-invertebrate fauna . Journal of the Royal Society of New Zealand , 35 : 287 – 301 . doi: 10.1080/03014223.2005.9517785
- Barratt , BIP , Tozer , P , Wiedemer , R , Ferguson , CM and Johnstone , PD . 2006 . Effect of fire on microarthropods in New Zealand indigenous grassland . Rangeland Ecology and Management , 59 : 383 – 392 . doi: 10.2111/05-190R1.1
- Barratt , BIP , Ferguson , CM , Bixley , AS , Crook , KE , Barton , DM and Johnstone , PD . 2007 . Field parasitism of non-target weevil species (Coleoptera: Curculionidae) by the introduced biological control agent Microctonus aethiopoides Loan (Hymenoptera: Braconidae) over an altitude gradient . Environmental Entomology , 36 : 826 – 839 . doi: 10.1603/0046-225X(2007)36[826:FPONWS]2.0.CO;2
- Barratt , BIP , Ferguson , CM , Barton , D and Johnstone , PD . 2009 . Impact of fire on tussock grassland invertebrate populations . Science for Conservation , 291 : 75
- Bell , NL , Davis , LT , Sarathchandra , U , Barratt , BIP , Ferguson , CM and Townsend , RJ . 2005 . Biodiversity of indigenous tussock grassland sites in Otago, Canterbury and the Central North Island of New Zealand. II Nematodes . Journal of the Royal Society of New Zealand , 35 : 303 – 319 . doi: 10.1080/03014223.2005.9517786
- Brockerhoff , EG , Barratt , BIP , Beggs , JR , Fagan , LL , Kay , MK , Phillips , CB and Vink , CJ . 2010 . Impacts of non-indigenous (exotic) invertebrates on New Zealand's indigenous species and ecosystems . New Zealand Journal of Ecology , 34 : 158 – 174 .
- Brown , JH . 1995 . Macroecology , 270 Chicago : University of Chicago Press .
- Clarke , KR and Gorley , RN . 2001 . PRIMER v5: User Manual/Tutorial , Plymouth , , UK : PRIMER-E .
- Cohen , AC . 1961 . Tables for maximum likelihood estimates: singly truncated and singly censored sample . Technometrics , 1 : 217 – 237 . doi: 10.1080/00401706.1959.10489859
- Colwell RK 2006 . EstimateS: statistical estimation of species richness and shared species from samples. Version 8.2. http://purl.oclc.org/estimates (accessed 28 August 2010) .
- Crook , KE , Ferguson , CM and Barratt , BIP . 2004 . “ Heat extraction of invertebrates from grassland turf samples ” . In Proceedings of the 8th Australasian Conference on Grassland Invertebrate Ecology Edited by: Winder , LM and Goldson , SL . 102 – 106 .
- Derraik , JGB , Barratt , BIP , Sirvid , P , Macfarlane , RP , Patrick , BH , Early , J , Eyles , AC , Johns , PM , Fraser , PM Barker , GM . 2001 . Invertebrate survey of a modified native shrubland, Brookdale Covenant, Rock and Pillar Range, Otago, New Zealand . New Zealand Journal of Zoology , 28 : 273 – 290 . doi: 10.1080/03014223.2001.9518270
- Derraik , JGB , Dickinson , KJM and Closs , GP . 2003 . Invertebrate diversity on Olearia bullata and Coprosma propinqua in a modified native shrubland, Otago, New Zealand . New Zealand Journal of Ecology , 27 : 55 – 60 .
- di Menna , ME , Sayer , ST , Barratt , BIP , Bell , NL , Ferguson , CM and Townsend , RJ . 2007 . Biodiversity of indigenous tussock grassland sites in Otago, Canterbury and the Central North Island. V. Penicillia and Aspergilli . Journal of the Royal Society of New Zealand , 37 : 131 – 137 . doi: 10.1080/03014220709510541
- Dickinson , KJM , Mark , AF , Barratt , BIP and Patrick , BH . 1998 . Rapid ecological survey, inventory and implementation: a case study from Waikaia Ecological Region, New Zealand . Journal of the Royal Society of New Zealand , 28 : 83 – 156 . doi: 10.1080/03014223.1998.9517556
- Douglas , MH and Allan , B . 1992 . “ The tussock grasslands ” . In Guide to tussock grassland farming , Edited by: Floate , MJS . 9 – 22 . Mosgiel , , New Zealand : AgResearch .
- Espie , P and Barratt , BIP . 2006 . Biodiversity of indigenous tussock grassland sites in Otago, Canterbury and the Central North Island of New Zealand IV. Vegetation and the effect of disturbance by agricultural development and fire . Journal of the Royal Society of New Zealand , 36 : 69 – 82 . doi: 10.1080/03014223.2006.9517801
- Floate , MJS . 1992 . Guide to tussock grassland farming , 127 Mosgiel , , New Zealand : AgResearch .
- Halloy , S and Barratt , BIP . 2007 . Patterns of abundance and morphology as indicators of ecosystem status: a meta-analysis . Ecological Complexity , 4 : 128 – 147 . doi: 10.1016/j.ecocom.2007.04.002
- Hutcheson , J . 1990 . Characterization of terrestrial insect communities using quantified, Malaise-trapped Coleoptera . Ecological Entomology , 15 : 143 – 151 . doi: 10.1111/j.1365-2311.1990.tb00795.x
- Krebs , CM . 1989 . Ecological Methodology , New York : Harper Collins .
- Leschen , RAB , Lawrence , JF , Kuschel , G , Thorpe , S and Wang , Q . 2003 . Coleoptera genera of New Zealand . New Zealand Entomologist , 26 : 15 – 28 . doi: 10.1080/00779962.2003.9722105
- Lister C Is the exotic weevil pest, Argentine stem weevil a threat to native grassland? 2006 10 Summer Scholarship Report, AgResearch, Mosgiel
- Lowe , AD . 1973 . “ Insects ” . In The Natural History of New Zealand – an Ecological Survey , Edited by: Williams , GR . 190 – 203 . Wellington : Reed .
- Lowther , WL and Douglas , MH . 1992 . “ Pasture establishment and development ” . In Guide to tussock grassland farming , Edited by: Floate , MJS . 55 – 60 . Mosgiel : AgResearch .
- Melo , AS , Niyogi , DK , Matthaei , CD and Townsend , CR . 2003 . Resistance, resilience, and patchiness of invertebrate assemblages in native tussock and pasture streams in New Zealand after a hydrological disturbance . Canadian Journal of Fisheries and Aquatic Sciences , 60 : 731 – 739 . doi: 10.1139/f03-061
- Morris , MG and Rispin , WE . 1987 . Abundance and diversity of the coleopterous fauna of a calcareous grassland under different cutting regimes . Journal of Applied Ecology , 24 : 451 – 465 . doi: 10.2307/2403886
- Murray , TJ . 2001 . Plant-animal associations between weevils (Coleoptera: Curculionidae) and the vegetation in two Otago tussock grasslands. Unpublished Batchelor of Science with Honours thesis , 53 Dunedin : University of Otago .
- Murray , TJ , Barratt , BIP and Dickinson , KJM . 2003 . Comparison of the weevil fauna (Coleoptera: Curculionoidea) in two tussock grassland sites in Otago, New Zealand . Journal of the Royal Society of New Zealand , 33 : 703 – 714 . doi: 10.1080/03014223.2003.9517754
- Murray , TJ , Dickinson , KJM and Barratt , BIP . 2006a . Associations between weevils (Coleoptera: Curculionidea) and plants, and conservation values in two tussock grasslands, Otago, New Zealand . Biodiversity and Conservation , 15 : 123 – 137 . doi: 10.1007/s10531-004-4246-3
- Murray , TJ , Dickinson , KJM and Barratt , BIP . 2006b . “ Associations between weevils (Coleoptera: Curculionidea) and plants, and conservation values in two tussock grasslands, Otago, New Zealand ” . In Arthropod diversity and conservation , Edited by: Hawksworth , DL and Bull , AT . 109 – 123 . Dortrecht , , The Netherlands : Springer .
- Myers , N , Mittermeier , RA , Mittermeier , CG , da Fonseca , GAB and Kent , J . 2000 . Biodiversity hotspots for conservation priorities . Nature , 403 : 853 – 858 . doi: 10.1038/35002501
- New TR . 2007 . Beetle conservation . Springer , Dortrecht , , The Netherlands . 94
- Patrick , BH . 2004 . Conservation of New Zealand's tussock grassland moth fauna . Journal of Insect Conservation , 8 : 199 – 208 .
- Prestidge , RA , Barker , GM and Pottinger , RP . 1991 . The economic cost of Argentine stem weevil in New Zealand. Proceedings of the 44th New Zealand Weed and Pest Control Conference 165 – 170 .
- Ridley GS Bain J Bulman LS Dick MA Kay MK Threats to New Zealand's indigenous forests from exotic pathogens and pests Wellington 2000 67 Science for Conservation 142 Department of Conservation
- Rufaut , CG . 2002 . Biodiversity in fragmented indigenous grasslands: Patterns of plant and invertebrate diversity in mid-altitude tall tussock grasslands (Chionochloa rigida (Raoul) Zotov), eastern Central Otago, New Zealand , 202 Dunedin : University of Otago . Unpublished PhD thesis
- Sarathchandra , SU , Burch , G , Sayer , ST , Waipara , NW , Cox , NR , Ghani , A , Bell , NL , O'Callaghan , M , Young , SD Barratt , BIP . 2005 . Biodiversity of indigenous tussock grassland sites in Otago, Canterbury and the Central North Island of New Zealand. III Soil microorganisms . Journal of the Royal Society of New Zealand , 35 : 321 – 337 . doi: 10.1080/03014223.2005.9517787
- Scott M Fine-scale ecology of alpine patterned ground, Old Man Range, Central Otago, New Zealand University of Otago Dunedin 2007 338 Unpublished PhD thesis
- Waterhouse DF . 1973 . The insects of Australia . Melbourne University Press , Melbourne . 1029
- Watt , JC . 1984 . A review of some New Zealand Scarabaeidae (Coleoptera) . New Zealand Entomologist , 8 : 4 – 24 . doi: 10.1080/00779962.1984.9722456