
Abstract
Holocene New Zealand sea lion (Phocarctos hookeri) bones collected from the north of New Zealand's South Island strengthen existing evidence for the former Holocene presence of breeding colonies of P. hookeri on mainland New Zealand. The taxonomic identity of Phocarctos bones is confirmed using both morphology and ancient DNA analysis. Five radiocarbon dates on four adult and one pup bone from Creighton's Cave near Paturau, northwest Nelson, ranged from 1290±30 yr BP to 5430±30 yr BP. Three radiocarbon dates on pup bones spanning 200 14C yr (1550±30 yr BP to 1390±30 yr BP) reveal that a prehistoric breeding rookery was present at Delaware Bay, Nelson, until shortly before the time of human arrival c. AD 1280 (670 yr BP). The Delaware Bay site in particular provides a valuable ‘snapshot’ of coastal New Zealand faunas shortly before human arrival, one that has potential to enhance our understanding of changes in the endemic coastal fauna associated with human colonisation.
Introduction
Palaeontological data can provide important insights into the composition of endemic biotas prior to human colonisation, and the subsequent ecosystem changes associated with human arrival (Holdaway Citation1989; Smith Citation2013). Such data have been applied to studies of biological change associated with human arrival in recently colonised regions of the globe such as Madagascar, New Zealand and numerous Pacific Islands (Worthy & Holdaway Citation2002; Burney et al. Citation2004; Steadman Citation2006). New Zealand's Quaternary fossil record is particularly rich and Worthy & Holdaway (Citation2002) describe it as ‘one of the best Late Quaternary (less than 50,000 years) fossil records in the world’ (p. xxx). Numerous prehistoric coastal taxa are well represented in deposits throughout coastal New Zealand (Anderson Citation1982; Worthy Citation1992, Citation1994; Smith Citation2013). Given the excellent preservation of pre-human faunal remains, it might seem surprising that relatively few studies have described and dated pre-human coastal assemblages in natural deposits (see Worthy Citation1998a).
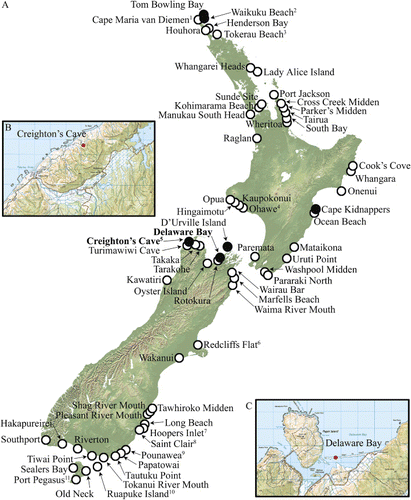
Prehistoric New Zealand sea lion Phocarctos hookeri (Gray 1844) remains have been recorded along the length of mainland New Zealand's coastline (; Appendix 1) and on offshore islands including Stewart Island, the Chatham Islands and Enderby Island (Auckland Islands) (Smith Citation1985; McFadgen Citation1994; Childerhouse & Gales Citation1998; Anderson Citation2005). These widespread remains indicate that the pre-human geographic range of this species was much broader than otherwise suggested by the predominantly high-latitude (largely sub-Antarctic) range of the species today (Smith Citation1985; Childerhouse & Gales Citation1998). The majority of P. hookeri remains from mainland sites have been excavated from archaeological middens that cover the period of early Polynesian settlement approximately 600–700 years ago (Smith Citation1985). Smaller numbers of naturally deposited (pre-human) P. hookeri specimens have also been recorded from sand dunes (Worthy Citation1994; Gill Citation1998) and caves (Worthy Citation1992) on mainland New Zealand from the late Holocene.
A lack of pup and juvenile P. hookeri bones in natural deposits and archaeological sites has, in the past, contributed to uncertainty as to whether breeding populations of P. hookeri existed on mainland New Zealand at the time of human arrival. Cawthorn (Citation1993) described the distribution of archaeological P. hookeri material as ‘consistent with the current distribution and range of Hooker's sea lion’ (p. 1), suggesting that ‘no historical evidence of breeding rookeries (i.e. pup remains) has yet been found in mainland New Zealand’. However, Worthy (Citation1994) suggested that small, porous pup bones were likely to be under-represented in natural dune deposits due to their relatively poor preservation potential. This phenomenon was discussed in relation to P. hookeri remains collected from the North Cape, where Gill (Citation1998, p. 56) noted that the pup bones ‘survived less well owing to their fragility’ in comparison with adult bones from the same location.
The northern and northwestern regions of the South Island are of particular significance in terms of understanding prehistoric P. hookeri and their pre-human breeding distribution. Worthy (Citation1994) provided the first evidence for a P. hookeri pup on mainland New Zealand from this region (Delaware Bay, Nelson), suggesting that this species was indeed breeding on the mainland during the Holocene. Worthy (Citation1994) also re-examined P. hookeri material collected from caves at Paturau (northwest Nelson), including Creighton's Cave () (Worthy Citation1992) and identified further pup bones. Holocene-aged remains of ‘young’ P. hookeri were previously identified from Hawkes Bay (Berry & King Citation1970; Weston et al. Citation1973), however it is not certain whether these bones were from pups or juveniles sufficiently young to definitively indicate breeding at this site.
Following the discovery of P. hookeri pup remains at Delaware Bay and Paturau (), additional observations of P. hookeri pup bones have been made from New Zealand's North Island. Gill (Citation1998) observed the remains of both adult and juvenile P. hookeri from Tom Bowling Bay and Waikuku Beach in Northland. These Northland pup remains were found in Holocene-aged sand dunes and are thought to predate human settlement (Gill Citation1998). A pup bone has also been recorded from a likely breeding colony on D'Urville Island (Museum of New Zealand Te Papa Tongarewa [NMNZ] collection database). Additionally, juvenile P. hookeri remains have been identified from sand dunes on the Chatham Islands (McFadgen Citation1994), providing further evidence for a formerly widespread distribution of breeding P. hookeri prior to human arrival.
Here we undertake field sampling, radiocarbon-dating and ancient DNA analysis of Holocene P. hookeri bones to shed light on the history of sea lions on mainland New Zealand. This research provides further evidence for the former presence of breeding colonies of P. hookeri on mainland New Zealand during the Holocene until just prior to human arrival using radiocarbon dating of newly excavated bones and ancient DNA analysis to confirm the taxonomic identity of these bones.
Methods
Sampling
Phocarctos hookeri bones were collected from Delaware Bay and Creighton's Cave () on 1 and 2 December 2011. Bones were collected and stored in plastic ziplock bags until required for analysis.
Delaware Bay
Delaware Bay is a sand dune site, approximately 15 km northeast of Nelson, where Worthy (Citation1994) first recorded P. hookeri pup remains on mainland New Zealand. We sampled the south-facing edge of the sand dune, described by Worthy (Citation1994), covering an area of c. 500 m2. Bones observed on the surface were collected, as well as additional bones of an individual found to be present beneath the surface (up to a depth of c. 200 mm). There is evidence of early Māori occupation near this site; umu (Māori ovens) are visible at the western end of the sand dune and middens are present in the upper levels of the nearby sand dune (Worthy Citation1994). However, the P. hookeri bones collected from Delaware Bay were all from natural deposits stratigraphically below the Polynesian middens (Worthy Citation1994). Bones of a number of different vertebrate taxa including various coastal bird species were also collected during this sampling trip, including the Waitaha penguin Megadyptes waitaha Boessenkool et al. 2009, Caspian tern Hydroprogne caspia (Pallas 1770) and fluttering shearwater Puffinus gavia (J.R. Forster 1844) (Appendix 2). These specimens are housed in the NMNZ and Canterbury Museum (CM).
Creighton's Cave
There are numerous limestone caves throughout the Paturau region of the northwest South Island, and Holocene terrestrial and coastal vertebrate assemblages have been observed in several of these sites (Worthy Citation1992). In many cases, openings in the roofs of these caves have created ‘pitfall traps’ that have facilitated accumulation of terrestrial fauna over time. Creighton's Cave is a large cave located 500 m from the present shoreline, and is currently surrounded by farmland. The cave is entered via its resurgence entrance and a little over 1 km of passages are known upstream of this point. The bones derive from gravels in the current stream bed 100–200 m upstream of the entrance in a passage that is about 10 m wide and 25 m high (Worthy et al. Citation1992). As there is one main entrance to the cave, Worthy (Citation1992) suggested it is likely that Holocene bones in Creighton's Cave have fallen through ‘sinkhole’ openings from the valley floor into the cave. Pre-European faunal remains that we collected from Creighton's Cave included extinct and locally extinct taxa such as moa (Aves: Dinornithiformes), South Island snipe Coenocorypha iredalei (Rothschild 1921) (Appendix 3) and P. hookeri. These specimens are also housed in NMNZ.
Phocarctos hookeri bones have also been recorded from nearby caves in the Paturau region (Worthy Citation1992). Sea lion bones from Wet Neck and Turimawiwi Caves have been dated to 3690–2380 yr BP and 12,036–10,882 yr BP, respectively (Worthy Citation1992).
Morphological and genetic analyses
Phocarctos hookeri remains from Delaware Bay and Creighton's Cave were identified morphologically following Worthy (Citation1992, Citation1994). Avifaunal remains were identified morphologically using collections at NMNZ and CM and their classification follows Gill et al. (Citation2010). To confirm morphological identifications of sea lion bones, a short (189 base pairs), highly variable region of the mitochondrial control region was amplified and sequenced from one individual from each of Delaware Bay and Creighton's Cave. No specimens from previous collections (Worthy Citation1992, Citation1994) were sampled for ancient DNA (aDNA) analysis. All aDNA extractions and polymerase chain reaction (PCR) set up were carried out in a purpose built ancient DNA laboratory at the University of Otago. The lab is physically isolated from any other molecular laboratories (Knapp et al. Citation2012) and we followed strict ancient DNA procedures to minimise contamination of samples with exogenous DNA (Cooper & Poinar Citation2000).
DNA was extracted from bone from one Delaware Bay pup (330890, CM Ma4105, right humerus, 204 mg) and one Creighton's Cave pup (333719, NMNZ S.45732, right humerus, 140 mg), following Rohland et al. (Citation2010). Two primer pairs (NZSLCR1f 5′ ATGTATATCGTGCATTAGTGGTTTG 3′/NZSLCR1r 5′ TTGTGGGCTAGGTGAATTA 3′ and NZSLCR2f 5′ AACTACTTCAAGCACTATAAAGTTCC 3′/NZSLCR2r 5′ AGTCTAGCTACCCCCGTT 3′) were designed, based on published whole mitochondrial genome sequences of P. hookeri (GenBank accession number AM181019.1) to amplify short, overlapping regions of the control region, resulting overall in a 189 base pair fragment (excluding primers). Each PCR was replicated twice. PCR products (from independent amplifications) were sequenced bi-directionally and blasted against existing modern P. hookeri control region sequences available on GenBank. This methodology allowed robust species identification of the bones collected.
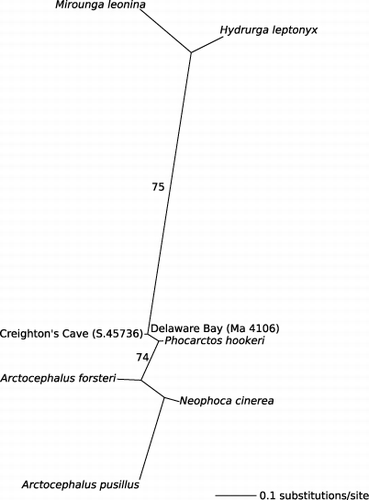
A Maximum Likelihood analysis was performed in PAUP* (Swofford Citation2003), including sequences from the Delaware Bay and Creighton's Cave individuals, modern P. hookeri (GenBank accession number AM181019.1) and pinniped outgroups (Australian sea lion Neophoca cinerea [Peron 1816] AM181020.1, Australian fur seal Arctocephalus pusillus [Schreber 1775] NC_008417, New Zealand fur seal Arctocephalus forsteri Lesson 1828 NC_004023, southern elephant seal Mirounga leonina [Linnaeus 1758] NC_008422.1, Leopard seal Hydrurga leptonyx [Blainville 1820] NC_008425.1). Sequences were aligned using the Clustal W algorithm and checked by eye. The most appropriate model of evolution, as determined by MODELTEST (Posada & Crandall Citation1998) under the Akaike information criterion, was K81uf + I. A maximum likelihood analysis was performed in PAUP* (Swofford Citation2003) using the full heuristic search option. Parameters for the K81uf + I model were then re-estimated from the data and node support was calculated with 10,000 bootstrap replicates.
Radiocarbon dating of bones
Eight P. hookeri bones were radiocarbon dated () at Beta Analytic Inc (USA). Three pup bones from Delaware Bay, and four adult and one pup bone from Creighton's Cave were included in radiocarbon dating analyses. Dates are reported as radiocarbon ages, based on Libby T1/2=5568 years, uncorrected for calendar variation, in years before present (present is AD 1950). Radiocarbon ages were calibrated using OxCal v4.2 (Bronk Ramsey Citation2009) and the Marine09 calibration curve (Reimer et al. Citation2009). A local (Ligar Bay, −35±46) ΔR value (Petchey et al. Citation2008) was applied to the calibrations of radiocarbon dates. The large geographic ranges of many pinnipeds in their adult stage can make it difficult to select appropriate ΔR values for such taxa (Yoneda et al. Citation2001; de Bruyn et al. Citation2009). To account for this issue we also applied minimum and maximum New Zealand ΔR values (Petchey et al. Citation2008) to the adult bones radiocarbon-dated. Minimum and maximum ΔR values were not necessary for the calibration of radiocarbon dates associated with pup bones, as sea lion pups do not move great distances during the early stages of their life (Wilkinson et al. Citation2003; Chilvers & Wilkinson Citation2008). As applying the minimum or maximum ΔR values did not substantially alter the calibrated ages (results not shown), the Ligar Bay ΔR value of −35±46 was used hereafter for all radiocarbon dates yielded from P. hookeri bones. Calibrated ages are reported as 95.4% confidence calibrated ages in years BP. Calibrated ages were compared with published radiocarbon dates from Wairau Bar (moa eggshell) and rat-gnawed seeds as proxies for human presence in the northern South Island and New Zealand, respectively (Higham Citation1994; Wilmshurst & Higham Citation2004; Wilmshurst et al. Citation2008). Moa eggshell and rat-gnawed seed dates were calibrated using OxCal v4.2 (Bronk Ramsey Citation2009) and the ShCal04 calibration curve (McCormac et al. Citation2004).
Results
Morphological and genetic analyses
Thirteen and nine P. hookeri individuals (minimum number of individuals, MNI, based on most common element and ontogenetic stage) were collected in 2011 from Delaware Bay and Creighton's Cave, respectively (). One pup bone from each locality was sequenced for the 189 base pair control region fragment. Both sequences exhibited phylogenetic similarity with modern P. hookeri (genetic divergence <4%) and were highly genetically distinct from all other modern pinniped taxa (genetic divergence 10.9%–23.3% ().
Table 1 Phocarctos hookeri bones collected from Delaware Bay (DEB) and Creighton's Cave (CRC). Bones were identified to element and age, following the methodology of Worthy (1992, 1994). Delaware Bay samples not consumed in analyses will be stored at the Canterbury Museum and Creighton's Cave samples will be stored at the Museum of New Zealand Te Papa Tongarewa.
Radiocarbon dating of Delaware Bay and Creighton's Cave bones
Three P. hookeri pups were radiocarbon dated from Delaware Bay. All three bones were of a similar age, covering a period of less than 200 14C yr (1264–1008 yr BP to 1122–852 yr BP; , ). Bones from four adult and one pup P. hookeri were dated from Creighton's Cave. Radiocarbon dating of these bones revealed more variation between dates, with ages ranging from 5970–5701 yr BP to 978–730 yr BP (, ). Comparison of the Delaware Bay and Creighton's Cave dates with previously published proxies for human arrival in New Zealand (Higham et al. Citation1999; Wilmshurst & Higham Citation2004; Wilmshurst et al. Citation2008) confirms the presence of breeding P. hookeri in the region prior to human colonisation ().

Table 2 Radiocarbon dates on Phocarctos hookeri remains from Delaware Bay and Creighton's Cave. Dates were calibrated using the Marine09 calibration curve (Reimer et al. Citation2009) in OxCal v4.2 (Bronk Ramsey Citation2009) and a local (Ligar Bay) ΔR of −35±46 (Petchey et al. Citation2008) was applied to all P. hookeri dates. Asterisks indicate pup bones.
Discussion
Radiocarbon dating and genetic analyses of adult and juvenile P. hookeri remains from the northern South Island strengthen existing evidence for the presence of breeding P. hookeri on mainland New Zealand prior to the time of human arrival (Worthy Citation1994).
Polynesians colonised New Zealand c. AD 1280 (Higham et al. Citation1999; Wilmshurst & Higham Citation2004; Wilmshurst et al. Citation2008). The radiocarbon dating of the Delaware Bay P. hookeri bones suggests that this site represents a ‘snapshot’ of mainland New Zealand's coastal biota shortly prior to human settlement. Specifically, all radiocarbon dated bones from Delaware Bay yielded calibrated ages from AD 744–1080 (maximum to minimum 95.4% calibrated range; , ). These data confirm that P. hookeri were breeding in the northern South Island during the Holocene and as recently as 200 years prior to human settlement. Remains of additional taxa were also recorded at Delaware Bay, including species that are now extinct or locally extirpated, such as the New Zealand coot Fulica prisca Hamilton, 1893, Fiordland crested penguin Eudyptes pachyrhynchus G.R. Gray, 1845, and Megadyptes waitaha (Appendix 2). The observation of a natural coastal site providing a temporal ‘snapshot’ of New Zealand biota, shortly prior to human arrival, has potential to enhance our understanding of human-mediated changes to the country's coastal fauna.
As expected for a cave that has been present and potentially able to accumulate fossils for thousands of years (Worthy Citation1992, Citation1994), and given the period over which the cave has been similarly placed relative to the coastline, P. hookeri bones collected from Creighton's Cave yielded a wider age range than those obtained from Delaware Bay dune strata. The oldest Creighton's Cave bone in our sample was dated to 5970–5701 yr BP (, ), whereas the youngest (978–730 yr BP; , ) was similar in age to the Delaware Bay material. These new radiocarbon-dates extend the previously estimated age range of P. hookeri in the Paturau region (from 12,053–10,894 yr BP to 3690–2380 yr BP; Worthy Citation1992), indicating the presence of P. hookeri until closer to the time of human settlement. The older of these new Creighton's Cave dates are similar in age to radiocarbon-dated sea lion material from nearby Wet Neck Cave (Worthy Citation1992). The wide range of dates obtained from Creighton's Cave suggest a continuous Holocene presence of breeding colonies of P. hookeri on mainland New Zealand. The presence of an introduced bird species (common starling Sturnus vulgaris Linnaeus 1758) in these caves (Appendix 3) shows that they have continued to act as sites of bone preservation until the present day.
The differences between faunal assemblages from Delaware Bay and Creighton's Cave (Appendix 2 and 3) reflect the different depositional environments at these sites. Delaware Bay accumulated coastal species, whereas Creighton's Cave accumulated mainly species that were trapped by pit fall (e.g. flightless or ground-dwelling taxa, such as moa and kiwi Apteryx spp.) or taxa that sometimes lived in the cave, for example, laughing owl Sceloglaux albifacies (G.R. Gray 1844). At Delaware Bay, 36 vertebrate taxa were recorded, of which 12 (33%) are locally extinct and three (8%) are globally extinct, compared with Creighton's Cave, where 18 vertebrate taxa were recorded, of which five (28%) are locally extinct and seven (39%) are globally extinct. The higher proportion of globally extinct taxa detected at Creighton's Cave reflects that flightless and ground-dwelling taxa have been disproportionately affected by human arrival (Worthy & Holdaway Citation2002).
Conclusion
The presence of a number of bones from both adult and juvenile P. hookeri from up to 5430 yr BP () across a wide geographic area () strongly suggests that prior to the time of human arrival, mainland New Zealand possessed a substantial breeding population of P. hookeri. The likely rapid human-mediated extinction mirrors those previously inferred for more than a quarter of New Zealand's terrestrial avifaunal species, including moa (Tennyson & Martinson Citation2007; Rawlence & Cooper Citation2013; Rawlence et al. Citation2012), giant geese Cnemiornis spp. and Haast's eagle Aquila moorei (Haast 1872) (Tennyson & Martinson Citation2007). We hypothesise that the local extirpation of breeding colonies of P. hookeri from the northern and northwest South Island likely occurred over a timeframe comparable to New Zealand's avian megafauna c. AD 1280–1450 (Tennyson & Martinson Citation2007).
Acknowledgements
Funding was provided by a Royal Society of New Zealand Marsden Grant (UOO1112) and the Allan Wilson Centre for Molecular Ecology and Evolution. Special thanks to Lisa Matisoo-Smith for providing access to the University of Otago ancient DNA laboratory and two anonymous reviewers who provided helpful comments on this manuscript. We thank the land owner at Paturau, Ann Sturgess and farm manager Nigel Clarke and at Delaware Bay Bruce and Carol Wilson for facilitating access to the study sites on their land.
References
- Anderson AJ 1982. The Otokia Mouth site at Brighton. New Zealand Archaeological Association Newsletter 25: 47–52.
- Anderson AJ 2005. Subpolar settlement in South Polynesia. Antiquity 79: 791–800.
- Anderson AJ, Allingham B, Smith IWG 2006. Shag river mouth: the archaeology of an early Southern Maori village. Canberra, Australian National University.
- Anderson AJ, Gillies KB, Wilson ASR, Smith IWG 1978. A site survey on the North Otago caost from Waitaki River Mouth to Warrington. Report to The New Zealand Historic Places Trust.
- Anderson AJ, Smith IWG 1992. The Papatowai site: new evidence and interpretations. Journal of the Polynesian Society 101: 245.
- Berry JE, King JE 1970. The identity of the Pliocene seal from cape kidnappers, New Zealand, previously known as Arctocephalus caninus. Tuatara 18: 13–18.
- Bone K 2012. The archaeology or early post-contact settlement: a re-evaluation of the William Cook ship building site. BA Hons dissertation (Anthropology and Archaeology). Dunedin, New Zealand, University of Otago.
- Bronk Ramsey C 2009. Bayesian analysis of radiocarbon dates. Radiocarbon 51: 337–360.
- Buist AG 1962 Archaeological evidence of the archaic phase of occupation in South Taranaki. New Zealand Archaeological Association Newsletter 5: 233–237.
- Burney DA, Burney LP, Godfrey LR, Jungers WL, Goodman SM, Wright HT, et al. 2004. A chronology for late prehistoric Madagascar. Journal of Human Evolution 47: 25–63. 10.1016/j.jhevol.2004.05.005
- Canavan T 1960. Preliminary report on excavations at the The Rangatapu site (Waingongoro River). New Zealand Archaeological Association Newsletter 3: 9–12.
- Cawthorn M 1993. Census and population estimation of Hooker's sea lion at the Auckland Islands December 1992–February 1993. Wellington, Department of Conservation Technical Series No. 2.
- Childerhouse S, Gales N 1998. Historical and modern distribution and abundance of the New Zealand sea lion Phocarctos hookeri. New Zealand Journal of Zoology 25: 1–16. 10.1080/03014223.1998.9518131
- Chilvers BL, Wilkinson IS 2008. Philopatry and site fidelity of New Zealand sea lions (Phocarctos hookeri). Wildlife Research 35: 463–470. 10.1071/WR07053
- Cooper A, Poinar HN 2000. Ancient DNA: do it right or not at all. Science 289: 1139.10.1126/science.289.5482.1139b
- Coutts PJF 1970. The Port Craig-Sand Hill Point regions of Southland: a preliminary archaeological report. Archaeology and Physical Anthropology in Oceania 5: 53–59.
- Coutts PJF 1972. The emergence of Foveax Strait Maori from prehistory: a study of culture contact. PhD thesis (Anthropology). Dunedin, New Zealand, University of Otago.
- Coutts PJF, Jurisich 1972. An archaeological survey of Ruapuke Island. Dunedin, Department of Anthropology, University of Otago.
- Davis FJ 1980. The prehistoric environment of the Dunedin area: the approach of salvage prehistory. MA thesis (Anthropology). Dunedin, New Zealand, University of Otago.
- de Bruyn M, Hall BL, Chauke LF, Baroni C, Koch PL, Hoelzel AR 2009. Rapid response of a marine mammal species to Holocene climate and habitat change. Plos Genetics 5: e1000554.10.1371/journal.pgen.1000554
- Furey L 2002. Houhora: a fourteenth century Maori village in Northland. Bulletin of the Auckland Museum No. 19. Auckland, Auckland War Memorial Museum. x + 169 p.
- Gill BJ 1998. Prehistoric breeding sites of New Zealand sea lions (Phocarctos hookeri, Carnivora: Otariidae) at North Cape. Records of the Auckland Museum 35: 55–64.
- Gill BJ, Bell BD, Chambers GK, Medway DG, Palma RL, Scofield RP, et al. 2010. Checklist of birds in New Zealand, Norfolk and Macquarie Islands, and the Ross Dependency Antarctica. Fourth Edition. Wellington, Te Papa Press in association with the Ornithological Society of New Zealand. x + 501 p.
- Hamel J 1977. Prehistoric man and his environment in the Catlins, New Zealand. PhD thesis (Anthropology). Dunedin, New Zealand, University of Otago.
- Hamel J 1980. Pounawea: the last excavation. Report to the New Zealand Historic Places Trust.
- Higham TFG 1994. Radiocarbon dating New Zealand prehistory with moa eggshell: some preliminary results. Quaternary Science Reviews 13: 163–169. 10.1016/0277-3791(94)90043-4
- Higham TFG, Anderson A, Jacomb C 1999. Dating the first New Zealanders: the chronology of Wairau Bar. Antiquity 73: 420–427.
- Holdaway R 1989. New Zealand's pre-human avifauna and its vulnerability. New Zealand Journal of Ecology 12: 11–25.
- Jacomb C, Brooks E, Mintmier M, Walter R 2010a. Final report on archaeological investigations at I44/121, St Clair, Dunedin. Southern Pacific Archaeological Research Report. 30 p.
- Jacomb C, Walter R, Brooks E 2012. Final report on salvage excavations at the Tokanui River Mouth under Authority 2011/232. Southern Pacific Archaeological Research Report Series No. 114. 51 p.
- Jacomb C, Walter R, Jennings C 2010b. Review of the archaeology of Foveaux Straight, New Zealand. Journal of the Polynesian Society 119: 25–59.
- Jeal M 1987. A Mahia Peninsula sea mammal butchery site (N127/14). New Zealand Archaeological Association Newsletter 30: 174–175.
- Knapp M, Clarke AC, Horsburgh KA, Matisoo-Smith EA 2012. Setting the stage - building and working in an ancient DNA laboratory. Annals of Anatomy - Anatomischer Anzeiger 194: 3–6. 10.1016/j.aanat.2011.03.008
- Leach HM 1976. Horticulture in prehistoric New Zealand. PhD thesis (Anthropology). Dunedin, New Zealand, University of Otago.
- Leach HM, Leach BF 1980. The Riverton site: an archaic adze manufactory in Western Southland. New Zealand Journal of Archaeology 2: 99–140.
- McCormac FG, Hogg A, Blackwell PG, Buck CE, Higham T, Reimer PJ 2004. ShCal04 Southern Hemisphere calibration, 0-11 cal kyr BP. Radiocarbon 46: 1087–1092.
- McFadgen BG 1994. Archaeology and Holocene sand dune stratigraphy on Chatham Island. Journal of the Royal Society of New Zealand 24: 17–44. 10.1080/03014223.1994.9517454
- Nichol RK 1988. Tipping the feather against a scale. PhD thesis (Anthopology). Auckland, University of Auckland.
- Petchey F, Anderson A, Hogg A, Zondervan A 2008. The marine reservoir effect in the Southern Ocean: an evaluation of extant and new ?R values and their application to archaeoogical chronologies. Journal of the Royal Society of New Zealand 38: 243–262. 10.1080/03014220809510559
- Posada D, Crandall KA 1998. MODELTEST: testing the model of DNA substitution. Bioinformatics 14: 817–818. 10.1093/bioinformatics/14.9.817
- Rawlence NJ, Cooper A 2013. Youngest reported radiocarbon age of a moa (Aves: Dinornithiformes) dated from a natural site in New Zealand. Journal of the Royal Society of New Zealand 43: 100–107.
- Rawlence NJ, Metcalf JL, Wood JR, Worthy TH, Austin JJ, Cooper A 2012. The effect of climate and environmental change on the megafaunal moa of New Zealand in the absence of humans. Quaternary Science Reviews 50: 141–153. 10.1016/j.quascirev.2012.07.004
- Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW, Blackwell PG, et al. 2009. Intcal09 and marine09 radiocarbon age calibration curves, 0–50,000 years Cal BP. Radiocarbon 51: 1111–1150.
- Robertson CJR, Hyvonen P, Fraser MJ, Pickard CR 2007. Atlas of bird distribution in New Zealand 1999–2004. Wellington, Ornithological Society of New Zealand x + 533 p.
- Rohland N, Siedel H, Hofreiter M 2010. A rapid column-based ancient DNA extraction method for increased sample throughput. Molecular Ecology Resources 10: 677–683. 10.1111/j.1755-0998.2009.02824.x
- Scarlett RK 1979. Avifauna and man. In: Anderson A. ed. Birds of a feather: osteological and archaeological papers from the South Pacific in honour of RJ Scarlett. Oxford, BAR International Series 62: 75–90.
- Scott SD 1970. Excavations at the ‘Sunde Site’, N38/24, Motutapu Island, New Zealand. Records of the Auckland Institute and Museum 7: 13–20.
- Smith IWG 1985. Sea mammal hunting and prehistoric subsistence in New Zealand. Unpublished PhD thesis. Dunedin, New Zealand, University of Otago.
- Smith IWG 1999. Settlement permanence and function at Pleasant River Mouth, east Otago, New Zealand. New Zealand Journal of Archaeology 19: 27–79.
- Smith IWG 2013. Pre-European Maori exploitation of marine resources in two New Zealand case study areas: species range and temporal change. Journal of the Royal Society of New Zealand 43: 1–37. 10.1080/03036758.2011.574709
- Spinks J n.d. MA thesis in preparation (Anthropology). Dunedin, New Zealand, University of Otago.
- Steadman DW 2006. Extinction and biogeography of tropical pacific birds. Chicago, University of Chicago Press.
- Swofford DL 2003. PAUP*. Phylogenetic analysis using parsimony (* and other methods). Version 4. Sunderland, Massachusetts, Sinauer Associates.
- Teal TF 1975. Pleasant river excavations 1959–1962. Research report, University of Otago.
- Tennyson AJD, Martinson P 2007 ( reprint). Extinct birds of New Zealand. Wellington, Te Papa Press. 180 p.
- Trotter MM 1975. Archaeological investigation at Redcliffs, Canterbury, New Zealand. Records of the Canterbury Museum 9: 189–220.
- Walter R, Jacomb C 2005. An archaeological survey of Hakapureirei (Sand Hill Point). South Pacific Archaeological Research Report. 46 p.
- Walter R, Jacomb C, Brooks E 2011a. Excavations at Cooks Cove, Tolaga Bay, New Zealand. Journal of Pacific Archaeology 2: 1–28.
- Walter R, Jacomb C, Jennings C 2011b. Final report on excavations at the Kawatiri Site (K29/8), West Coast, under NZHPT Authority (2004/172). Southern Pacific Archaeological Research Report. 51 p.
- Weston RJ, Repenning CA, Fleming CA 1973. Modern age of supposed Pliocene seal, Arctocephalus caninus Berry (= Phocarctos hookeri Gray), from New Zealand. New Zealand Journal of Science 16: 591–598.
- Wilkinson IS, Burgess J, Cawthorn MW 2003. New Zealand sea lions and squid - managing fisheries impacts on a threatened marine mammal. In Glaes N, Hindell M, Kirkwood R eds. Marine mammals: fisheries, tourism and management issues. Melbourne, CSIRO Publishing. Pp. 192–207.
- Wilmshurst JM, Anderson AJ, Higham TFG, Worthy TH 2008. Dating the late prehistoric dispersal of polynesians to New Zealand using the commensal Pacific rat. Proceedings of the National Academy of Sciences of the United States of America 105: 7676–7680. 10.1073/pnas.0801507105
- Wilmshurst JM, Higham TFG 2004. Using rat-gnawed seeds to independently date the arrival of Pacific rats and humans in New Zealand. The Holocene 14: 801–806. 10.1191/0959683604hl760ft
- Worthy TH 1992. Fossil bones of Hooker's sea lions in New Zealand caves. New Zealand Journal of Natural Sciences 19: 31–39.
- Worthy TH 1994. Holocene breeding of sea lions (Phocarctos hookeri) at Delaware Bay, Nelson, South Island, New Zealand. New Zealand Journal of Natural Sciences 21: 61–70.
- Worthy TH 1998a. Fossils indicate Pelecanoides georgicus had large colonies at Mason Bay, Stewart Island, New Zealand. Notornis 45: 229–246.
- Worthy TH 1998b. The Quaternary fossil fauna of Southland. Journal of the Royal Society of New Zealand 28: 537–89.
- Worthy TH 1998c. A remarkable fossil and archaeological avifauna from Marfells Beach, Lake Grassmere, South Island, New Zealand. Records of the Canterbury Museum 12: 79–176.
- Worthy TH, Holdaway RN 2002. The lost world of the moa: prehistoric life of New Zealand. Bloomington, Indiana University Press.
- Worthy T, Ravens J, Glasgow S 1992. Caving at Paturau, northwest Nelson, Pt 1 - Caves north of the Paturau River. New Zealand Speleological Bulletin 8: 531–549.
- Yoneda M, Hirota M, Uchida M, Uzawa K, Tanaka A, Shibata Y, et al. 2001. Marine radiocarbon reservoir effect in the western North Pacific observed in archaeological fauna. Radiocarbon 43: 465–471.
Appendix 1
Appendix 1A Archaeological sites with sea lion bones.
Appendix 1B Locations with non-archaeological sea lion bones.
Appendix 2
A list of Holocene vertebrate fossil remains found at Delaware Bay. Accession numbers are provided for specimens housed at the Museum of New Zealand Te Papa Tongarewa. Further unregistered specimens of several of these species are held in the Canterbury Museum.
+Brown teal Anas chlorotis (NMNZ S.38518)
+Fiordland crested penguin Eudyptes pachyrhynchus (NMNZ S.38566)
++ Waitaha penguin Megadyptes waitaha (NMNZ S.38489, 38513, 38325, 38530, 38564, 42156)
Little penguin Eudyptula minor (NMNZ S.38475, 38482, 38490, 38515, 38526, 385331, 38563, 38565, 42137, 42157, 45498)
+Grey-faced petrel Pterodroma cf. macroptera (NMNZ S.38560)
+Mottled petrel Pterodroma inexpectata (NMNZ S.42158)
Fairy prion Pachyptila turtur (NMNZ S.38511, 38536, 38559)
Broad-billed prion Pachyptila vittata (NMNZ S.38512, 38537, 38561, 42138)
Sooty shearwater Puffinus griseus (NMNZ S.38523)
Fluttering shearwater Puffinus gavia (NMNZ S.38479, 38509, 38524, 38548, 38558, 42159, 45499)
Common diving petrel Pelecanoides urinatrix (NMNZ S.38480, 38510)
Pied shag Phalacrocorax varius (NMNZ S.38567, 42139, 42160)
+New Zealand king shag Leucocarbo carunculatus (NMNZ S.38476, 38532)
Spotted shag Stictocarbo punctatus (NMNZ S.38477, 38483, 38491, 38517, 38527, 38568, 42161)
#Swamp harrier Circus approximans (NMNZ S.38538)
Weka Gallirallus australis (NMNZ S.38484, 38519, 38552, 38569, 42162)
+South Island takahe Porphyrio hochstetteri (NMNZ S.38570)
++New Zealand coot Fulica prisca (NMNZ S.42140)
Bar-tailed godwit Limosa lapponica (NMNZ S.38486, 38571)
Variable oystercatcher Haematopus unicolor (NMNZ S.38487, 38534, 42163)
+New Zealand dotterel Charadrius obscurus (NMNZ S.38551)
Southern black-backed gull Larus dominicanus (NMNZ S.38478, 38516, 38528, 38535, 38554, 38573)
Caspian tern Hydroprogne caspia (NMNZ S.45497)
Black-fronted tern Chlidonias albostriatus (NMNZ S.38550, 38572)
White-fronted tern Sterna striata (NMNZ S.42141)
+Kakapo Strigops habroptilus (NMNZ S.38485)
+Kaka Nestor meridionalis (NMNZ S.38574)
+Parakeet Cyanoramphus spp. (NMNZ S.38553, 38555)
Morepork Ninox novaeseelandiae (NMNZ S.38529)
++ New Zealand raven Corvus antipodum (NMNZ S.38488, 38492, 38575, 42164)
#Eurasian blackbird Turdus merula (NMNZ S.38539)
+Tuatara Sphenodon punctatus (NMNZ S.42165)
Gecko Hoplodactylus sp. (NMNZ S.42142)
#Dog Canis lupus (NMNZ S.38474, 38540, 38577)
+#Pacific rat Rattus exulans (NMNZ S.38576)
+New Zealand sea lion Phocarctos hookeri (NMNZ S.38493, 38520-3, 38546, 38578-38582, 42143-42152, 42166-42175, 45500, 45510, 45513; CM Ma 4104-4116)
++ globally extinct,+locally extinct (excludes seabird species which still commonly wash up dead on local beaches) (see Robertson et al. Citation2007), # post-human arrival
Appendix 3
A list of Holocene vertebrate fossil remains found at Creighton's Cave. These collections are housed at the Museum of New Zealand Te Papa Tongarewa.
++ Upland moa Megalapteryx didinus (NMNZ S.45711)
++ Little bush moa Anomalopteryx didiformis (NMNZ S.35128, 38682, 45715, 45716, 45727)
++ South Island Giant Moa Dinornis robustus (NMNZ S.45724)
++ Crested moa Pachyornis australis (NMNZ S.35129)
Great spotted kiwi Apteryx haastii (NMNZ S.35166)
+Brown teal Anas chlorotis (NMNZ S.35173)
Little penguin Eudyptula minor (NMNZ S.34167)
+Cook's petrel Pterodroma cookii (NMNZ S.35172)
Weka Gallirallus australis (NMNZ S.35168, 35185, 45718, 46729)
++ South Island snipe Coenocorypha iredalei (NMNZ S.45721)
Southern black-backed gull Larus dominicanus (NMNZ S.35183)
+Kakapo Strigops habroptilus (NMNZ S.35169, 35184, 38681, 45719)
Kaka Nestor meridionalis (NMNZ S.45720)
++ Laughing owl Sceloglaux albifacies (NMNZ S.35171)
++ South Island stout-legged wren Pachyplichas yaldwyni (NMNZ S.35170)
++ South Island kokako Callaeas cinerea (NMNZ S.45722)
#Common starling Sturnus vulgaris (NMNZ S.45723)
+Tuatara Sphenodon punctatus (NMNZ S.35174)
+New Zealand sea lion Phocarctos hookeri (NMNZ S.38683, 45713, 45731-45738)
#Sheep cf. Ovis (NMNZ 45714)
++globally extinct,+locally extinct, #post-human arrival