
ABSTRACT
The remains of a unique fossil bony fish were discovered in late Eocene (39.1–36.7 Ma: NZ Kaiatan stage) mudstone at Burnside near Dunedin, New Zealand in the 1930s and subsequently named and described by Frederick Chapman. He interpreted the type specimen as being a large-scaled relative of the modern Thyrsites of the Gempylidae (Scombroidei: Trichiuroidea), known to be swift, large oceanic predators. However, Chapman is unlikely to have seen all of the fossil, and did not discuss the skull and caudal skeleton. Additional material now allows these to be included in the expanded description herein, including key morphologic features of the fish such as the presence of premaxillary fangs. This study describes the rather complex history of the specimen and re-examines this significant fossil fish in the University of Otago collections, giving a more complete understanding of Eothyrsites morphology, paleoecology and relationships. In summary, we suspect Eothyrsites represents an ancestral form of gempylid, closely related to the gemfish group, an important Southern Hemisphere macrofossil record from the Eocene seas around Zealandia.
Introduction
English born and later Melbourne-based Frederick Chapman (13 February 1864–10 December 1943) was the inaugural Australian Commonwealth Palaeontologist, known especially for his work on Foraminifera, and his books on Australasian fossils. In 1902 Chapman was appointed by the Victorian government to the newly created position of palaeontologist to the National Museum, Melbourne. While at the museum in 1914 Chapman was sent fish remains from New Zealand, his research later published in 1918 (as NZGS Paleontological Bulletin 7). Chapman’s descriptions covered 43 species of fish, 37 of which are chondrichthyian. His teleosts included: scales of an elopid identified as Thrissopater, from the Cretaceous of North Canterbury; Scombroclupea, a supposed clupeid from the Weka Pass area, Canterbury (presumably Oligocene); ‘Diplomystus’ from the Cretaceous of Marlborough; teeth of a sea-bream Sargus, from the Oligocene of Canterbury; and distinctive pharyngeal toothplates from two species of the wrasse Labrodon, found in mid-Cenozoic sediments in Canterbury (Chapman Citation1918).
From 1932 to 1943 Chapman was awarded for his efforts, being made an Honorary Fellow of the Royal Society of New Zealand (Benson Citation1944).
In 1933 Chapman was visited in Melbourne by William Noel Benson, University of Otago geologist, who provided Chapman the opportunity to study two fossil fish brought over from Otago collections (see note by Chapman Citation1934). The first fish was from Abbotsford, Dunedin, an impressive Early Eocene (Heretaungan NZ stage) skull that Chapman assigned to the now abandoned Cretaceous genus ‘Portheus’; Errol White (BMNH) suggested to Benson in 1945 that this specimen may be closer to the scombrid genus Scombrinus sp. (as noted in King et al. Citation2009 after Rust Citation2000).
The second fossil fish was also discovered near Dunedin, a partial skeleton from Burnside.
After first writing a popular newspaper article based on the specimen, which he named Eothyrsites or ‘dawn barracoota’ (see Chapman Citation1934), Chapman (Citation1935) formally described this specimen based on three mudstone slabs containing some skull material, pectoral fins and trunk. He interpreted the specimen as a relative of Thyrsites atun (the snake mackerel or snoek, sometimes called ‘New Zealand barracouta’) of the family Gempylidae (Nakamura and Parin Citation1993; Froese and Pauly Citation2022). This and the other 10 species of modern gemfishes found in New Zealand waters are swift, large oceanic predators, often characterised by elongate bodies and fang-like teeth (Stewart Citation2015).
Soon Chapman was working on other fossils, publishing on Australian cidaroid echinoderms in 1934. He had retired in 1935 when the article on the two New Zealand fossil fish was published by the Royal Society. In 1936 he returned to the National Museum of Australia as honorary palaeontologist. According to his assistant, micropaleontologist Irene Crespin (Citation1979), he was well liked: ‘outstanding among Chapman’s qualities were his quiet gentlemanly manner, his patience and his willingness to pass on his experience and guidance to all-comers’. Chapman was a prolific writer, publishing five books and some 500 scientific papers, some in collaboration, on geology, palaeontology and zoology. He died at Kew, Australia in 1943.
In the 1990s further material was located in the Otago University collection which appears to belong to Eothyrsites – additional fragments which contain more of the skull and the caudal fin. These have undergone examination and preparation (Rust Citation2000). It seems unlikely these pieces were ever examined by Chapman.
However, a series of letters between Benson and several paleontologists/geologists concerning a second fossil fish specimen from Burnside, dated between 1955 and 1956, are held in archives of the Hocken Library, Dunedin. Duplicates of this correspondence and a single additional letter from A. W. B. Powell (conchologist at Auckland Museum) to Benson about the fish are held by Auckland Museum.
Briefly summarising these letters: Benson contacted Powell asking if he would delay in studying the ‘new’ fossil fish material from Burnside that had been received by Professor B. J. Marples for the Otago University Museum, then packed in a box and sent to Auckland Museum (Benson Citation1955). Powell was overseas at the time and Auckland Museum director Gilbert Archey replied to Benson that the material had not been located (Archey Citation1956). Upon his return Powell wrote that the specimen had not been unpacked and would be held until further instruction was received from Benson (Powell Citation1956). Benson also wrote to fossil fish specialist Errol White at the BMNH, who had been an earlier acquaintance (Benson Citation1956), asking for assistance in revising Chapman’s description, and mentioned the material (still in Auckland) requiring further preparation. White (Citation1956) later replied declining to study the fish himself but suggested that Benson contact Belgian paleoichthyologist Edgard Casier. However Benson was busy completing his geological map of the Dunedin area and passed away not long after, in 1957. At this stage we cannot say if Casier was ever contacted about the fossil, or when the material was returned, probably by Powell, to the University of Otago collection.
In any case, Benson felt strongly that the original description of this unique New Zealand fossil genus and species warranted further examination and expansion. Even today the fossil record of marine teleost material from the New Zealand Cenozoic consists mostly of isolated bones or incomplete specimens (see review in King et al. Citation2009), so this relatively complete example is especially noteworthy.
Materials and methods
According to Chapman (Citation1935) the type series consisted of three syntypes – associated pieces of mudstone matrix containing semi-articulated fish remains; although the skeleton is crushed, elements are generally well preserved with only minimal distortion. His original description included OU 6329 a, a piece with vertebrae and scales (Chapman’s Figs 2 and 5), together with OU 6329 b, a small piece with numerous scales (a detached part of the above); OU 6332, a piece containing pectoral fin (Chapman’s Fig. 4); and OU 6854 d (once labelled 5982), a piece containing mandible (Chapman’s Fig. 3).
In addition to these we note pieces OU 6854 a, b, c plus three smaller fragmentary pieces, as yet unnumbered. This is thought to be the material sent away and discussed by Benson in his letters (Citation1955, Citation1956), but all originating from the same locality and horizon as Chapman’s specimen, the Burnside Mudstone, a unit from which macrofossils are rare.
It seems likely the material was collected at two different times, possibly decades apart. Chapman described part of the material in 1935. One of Benson’s letters to Powell (in Citation1955) stated: ‘ … in the Otago University museum a recently received fossil fish, from the Upper Eocene Burnside mudstone, which I fancied might be either Eothyrisites holosquamatus, Chapman or something related to it’. Benson clearly believed the ‘recently received’ material was a separate specimen.
The possibility exists that this assemblage constitutes parts of two separate individuals, however it seems more likely that they represent counterparts of a single skeleton, and they are treated as such herein. The remains all exhibit similar preservation, are all parts of a fish skeleton of consistent size, and there are no duplicated elements. Many pieces bear equivalent characteristic scales (hence Chapman’s species name holosquamatus), strongly suggesting all the fragments are from the same original specimen, separated in space and time during recovery and subsequent storage.
Taphonomic processes post mortem but before burial could explain aspects of the preservation of Eothyrsites, in particular the partial disintegration of the skeleton. In quiet oceanic depositional settings some fish carcasses have been shown to refloat, or partially refloat, in the days after death due to gases developing in the abdominal cavity. Often the head and tail may droop or may even detach before the carcass later re-sinks to the seafloor as a jumbled heap (Schäfer Citation1972). Hence we find in this specimen, the skeleton of Eothyrsites with overlapping elements, semi-articulated sections of trunk, a skull with the lower jaw detached from the neurocranium and upper jaw, and partially isolated fins and scales.
The fossil material is housed in the paleontology collection in the Department of Geology, University of Otago, Dunedin, New Zealand (OU). NMNZ refers to material held at the Museum of New Zealand Te Papa Tongarewa; BMNH refers to the British Museum of Natural History, now Natural History Museum (NHMUK).
Some of the additional pieces recovered required additional matrix removal before examination, to allow investigations of their skeletal structures in as much detail as possible. They were prepared using thin needles and studied using a stereomicroscope. Measurements were taken using callipers to the nearest 0.1 mm. Comparative information was derived mainly from the literature, although images of modern osteological material of Thyrsites atun (NMNZ P.002334, 002659) and Rexea solandri (NMNZ P.002339) were provided by A. Stewart from the national fish collection at Te Papa, Wellington.
Locality
The initial fossil material was collected, presumably by quarry workers during 1929–33, at the Burnside Marl Pit, formerly the quarry of the Milburn Lime and Cement Co. Ltd., also known as the Burnside Cementworks, Dunedin, New Zealand: 45.8961°S, 170.4423°E ().
Figure 1. Map of the fossil locality at Burnside, Dunedin, South Island, New Zealand. Map produced with S. Read (OU Geology).
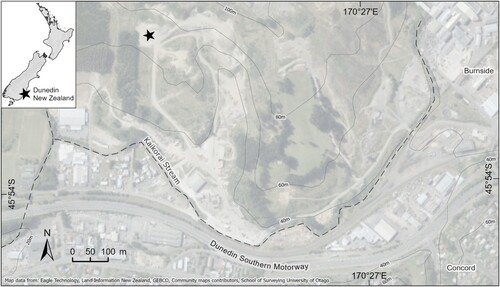
The marl or Burnside Mudstone (Benson Citation1968) is late Eocene in age (39.1–36.7 Ma: NZ Kaiatan stage, equivalent to Bartonian –Priabonian age). Details of the locality are recorded in the New Zealand Fossil Record File database (www.fred.org.nz), with the registration number I44/f0176; the map reference is NZMS 260 I44 115 757.
Geological setting and paleoenvironment
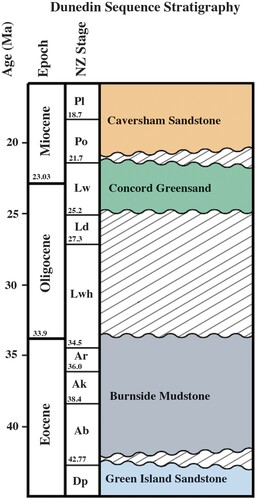
The regional distribution and lithology of the Burnside Mudstone has been discussed by Benson (Citation1968) and Bishop and Turnbull (Citation1996). The formation reaches up to 45 metres thick at its type locality from which the fossil fish was recovered (), being a fine-grained, massive mudstone, largely light grey in colour, soft semi-plastic, calcareous (up to 20% carbonate), with only occasional macrofossils. These include thin-shelled bivalves, a sea-pen (Hamilton Citation1958), and rare penguin bones (OU collections). Microfossil analyses suggest continuous deposition of the sediment probably occurred in quiet deeper waters (perhaps 200 m depth) of the outer shelf (see note by Hornibrook in Hamilton Citation1958). This is consistent with the preservational taphonomy of the fish skeleton discussed above; in addition there is little evidence of physical disturbance or scavenging of the remains before burial in the muddy sediment.
Figure 2. Stratigraphic column of locality. Boundaries are based on McMillan and Wilson (Citation1997).
Systematic paleontology
Infraclass teleostei mueller 1845
Series Pelagiaria Betancur-R et al. Citation2017.
Order Scombriformes Betancur-R et al. Citation2017.
Suborder Scombroidei (sensu Carpenter et al. Citation1995)
Trichiuroidea (sensu Nakamura & Parin Citation1993)
Family Gempylidae Gill, 1862.
Genus Eothyrsites Chapman Citation1935.
Type and only species: Eothyrsites holosquamatus Chapman Citation1935.
Diagnosis
Danilchenko (Citation1960, Citation1967) listed some key morphological features of fossil members of the family Gempylidae applicable here, notably the presence of an elongate, laterally compressed body, large head with lower jaw protruding and jaws bearing anterior large canine teeth (fangs). Gill cover smooth, without spines. Vertebrae count typically 30–55, with ribs on parapophyses. Anterior dorsal fin long, composed of comparatively thin hard rays, united with the short second (soft) dorsal fin. Anal fin opposite the second dorsal. Accessory finlets usually present. Pelvic fins below or slightly behind the pectorals, sometimes reduced to a pair of spines. Caudal fin moderately developed, deeply forked; caudal peduncle slender. However, advances in the study of molecular and morphologic (particularly skull) characters suggest the Gempylidae is actually paraphyletic and can better be termed a grade within the Trichiuroidea (summarised in Beckett et al. Citation2018, see Discussion section).
Type material: Consists of seven labelled syntypes (from 10 associated pieces of mudstone matrix containing semi-articulated fish remains from what we consider a single individual) currently catalogued as OU 6329, OU 6332 a, OU 6332 b; plus OU 6854 a, b, c and d (see ).
Figure 3. Eothyrsites holosquamatus Chapman, composite photograph of syntypes and associated material (OU 6329 a, b, OU 6332, OU 6854 a, b, c, d). Key to image: A, partial skull, OU 6854 c; B, piece containing lower jaw and opercular elements, part of tooth-bearing mandible labelled by Chapman, OU 6854 d; C, reverse side of OU 6854 d with preopercular labelled; D, close-up of maxillary teeth, OU 6854 a; E, abdominal region including vertebrae, rays, scales and small anal? fin, OU 6329 a; F, pectoral fin, OU 6854 a; G, partial pectoral fin labelled by Chapman, OU 6332; H, partial? fin, reverse side of OU 6332; I, caudal endoskeleton, OU 6854 b; J, trunk portion with scales labelled by Chapman, OU 6329 b. Note B, C, E, G, H and J were examined by Chapman (Citation1935). All scale bars 20 mm unless indicated otherwise.

The original three syntypes described by Chapman (Citation1935) were OU 6329 a + b, OU 6332 and OU 6854 d. Note that the OU 6854 d was at some stage marked 5982, and catalogued with reference to the scales.
Description of type specimen
Overall body shape: From the available material we suspect the type of Eothyrsites holosquamatus originally had a total body length in the order of 700 mm. The body was probably somewhat elongate, laterally compressed and slender, as for gemfish (Roberts and Stewart Citation1997), but perhaps not to the degree seen in most other modern gempylids (e.g. Matsubara and Iwai Citation1952; Nakamura and Parin Citation1993; Stewart Citation2015).
Skull: Mudstone slab OU 6854 c (A, ) contains much of the cranium and upper jaw, while the lower jaw is detached and fragmented, exposed along with opercular elements on piece OU 6854 d (B). We estimate the large skull of Eothyrsites was elongate and narrow, measuring approximately 190 mm in length anteroposteriorly (the posterior portion of the cranium, beyond the lateral commissure at the posterior margin of the orbit, including exoccipitals and basioccipital, is missing in the specimen), and 85 mm across transversely in the orbital region. This is however slightly larger and broader than the modern skull material of an adult Thyrsites atun (NMNZ P.002334, 002659) and Rexea solandri (NMNZ P.002339), as well as the early gempylid crania described in Beckett et al. (Citation2018). Although incomplete, an estimated skull length of approximately 30% body length is similar to modern gemfish such as Rexea spp. (e.g. Roberts and Stewart Citation1997).
Figure 4. Eothyrsites holosquamatus Chapman, photograph of skull OU 6854 c, ventral view.

When viewing the skull as exposed from the underside (), a long and narrow (approximately 100 × 20 mm) parasphenoid can be seen centrally. This compares well with those seen in modern skulls of Thyrsites and Rexea, and an English fossil specimen of a gempylid NHMUK PV OR 41318, shown in Figs 7 and 8 in Beckett et al. (Citation2018). The parasphenoid, which meets the basisphenoid posteriorly, extends anteriorly to the posterior margin of the vomer and meets this near the lateral ethmoids. The anterior tip of the vomer is missing and unfortunately it is not possible to determine if teeth were present, an important morphologic character (see Beckett et al. Citation2018). The skull roof (comprised largely of the paired frontal bones), the occipital crest and other features of the dorsal surface of the Eothyrsites skull are obscured by matrix. Towards the posterior of the cranium it is difficult to determine the sutures and shape of elements in the otic and occipital regions, including the pterotic and prootic, due to poor preservation.
Figure 5. Eothyrsites holosquamatus Chapman, line drawing of skull OU 6854 c, ventral view. Abbreviations: af, anterior premaxillary fang; boc, basioccipital; eth, ethmoid; fr, frontal; mx, maxilla; os, orbitosphenoid; pas, parasphenoid; pl, palatine; pmx, premaxilla; pro, prootic; pto, pterotic; spo, sphenotic; su, supraorbital, vo, vomer.
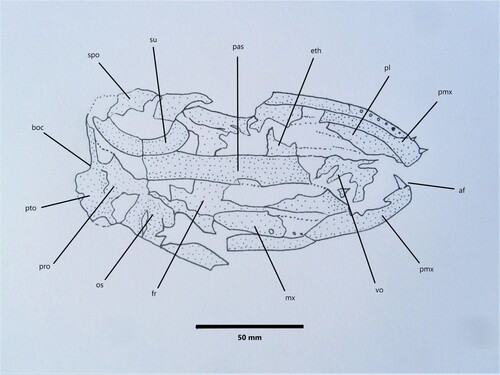
Eothyrsites probably had relatively large eyes. An estimated orbital length (horizontal across sclerotic) is 50 mm, which would constitute about 7% total body length of 700 mm, consistent within the range of modern gemfish such as Rexea (Roberts and Stewart Citation1997); or 26% skull length, slightly larger than those of modern Thyrsites atun (Stewart Citation2015), but similar to other fossil gempylids (Danilchenko Citation1960). The orbits were just posterior of the midlength of the head and behind the mouth, with the dorsal orbital margin probably coincident with the dorsal profile of the skull as for modern gemfish. Part of the curved circumorbital series (jugal + supraorbital?) or possibly a partial, slightly crushed sclerotic ring can be seen in the right orbital region of the skull (). While Carnvale (Citation2006) noted a report of the presence of ossified sclerotics in the gempylids Gempylus and Nesiarchus, he also pointed out that these ossifications are absent from other gempylid genera and the trichiurids. Sclerotic bones are apparently absent in the extinct genus Hemithyrsites (Danilchenko Citation1960).
Jaw elements: The mouth of Eothyrsites was large, with the upper jaws strengthened by the premaxilla being fixed and hence non-protrusible, typical of gempylids and other members of the Scombroidei. In the key study of scombroid phylogeny, Johnson (Citation1986, p. 21) also noted: ‘an inner row of two or three enlarged, fang-like teeth near the premaxillary symphysis characterises the outgroup taxa, Sphyraena and the gempylids. Absence of these specialised teeth is a synapomorphy of scombrids and billfishes’. Although many of the teeth of Eothyrsites are difficult to see in the fossil specimen as the skull is crushed, the presence of at least one slightly curved fang-like tooth, 11 mm long, at the anterior of the upper jaw ( and ), suggests Eothyrsites had at least one pair of symphysial teeth. We follow Bemis et al. (Citation2019) here in use of the term ‘fang’. There is no obvious barb on the Eothyrsites premaxillary fang, such as those seen in the cutlassfish Trichiurus (Bemis et al. Citation2019), and it appears to be more conical than compressed in section. This compares well with a 12 mm symphysial fang seen in a modern Thyrsites specimen (NMNZ P.002334) and a 15 mm fang in Rexea solandri (NMNZ P.002339).
In Eothyrsites each premaxilla also supported a number (at least nine) of smaller inwardly curved, conical (but not lanceolate) lateral teeth. A modern Thyrsites specimen (NMNZ P.002334) premaxilla bears 20 smaller (4–5 mm length) conical teeth; Rexea solandri (NMNZ P.002339) has a similar number of slightly larger (up to 7 mm) teeth. The lateral premaxillary teeth of Trichiurus lepturus and the snake mackerel Gempylus serpens are reduced in number but larger than those of Eothyrsites (see Bemis et al. Citation2019). Another gempylid, the oilfish Ruvettus, has similar teeth but Ruvettus (and Lepidocybium) don’t have the enlarged fangs at the anterior (A. Stewart pers. comm.).
There was probably a row of slightly smaller teeth in the lower jaw of Eothyrsites. Chapman noted and figured five ‘mandibular’ teeth in series on OU 6854 d, each 4 mm long, laterally compressed yet somewhat more slender than those of the premaxilla and slightly inwardly curved (B). We suspect these actually represent dentary teeth, as immediately adjacent to the oral fragment of the dentary this piece of the specimen also contains part of the posterior lower jaw – including at least a broad triangular 80 × 50 mm portion of the posterior end of the articular and angular. This exhibits a notable facet for the articular condyle of the quadrate and resembles in shape that seen in Thyrsites atun (NMNH P.002659). Unfortunately the anterior portion of the dentary is not obvious in the specimen, so we are unable to assess whether Eothyrsites also exhibited enlarged dentary fangs (cf. Promethicthyes) or had a protruding lower jaw (as for Thyrsites and Rexea).
Opercular series: A number of partly overlapping broad, flat elements that presumably include the opercular bones are present in the specimen on OU 6854 d (C). In fact this piece also contains part of several incomplete suspensorium and lower jaw elements (possibly the quadrate as well as the angulo-articular) compressed together under the gill cover. Chapman noted the preopercular, which is probably in fact the lower rounded portion of the interopercular, which occur paired with the subopercular and flattened, each side measuring approximately 60 × 30 mm. Typical for the family, the gill cover elements are smooth, without any spines or significant ornament (Danilchenko Citation1960). Part of several branchiostegal rays may just be visible on the edge of the specimen.
Postcranial skeleton and fins
Vertebral column: At least eight articulated amphicoelus trunk centra are present in the specimen piece OU 6329 (E, ), of length 20 mm anteroposteriorly and with a height of 18 mm. Toward the anterior and posterior of the body some are damaged and there appear to be vertebrae missing. Portions of at least three centra of similar size are present on the smaller unnumbered fragments. We would expect a total vertebral count of at least 30 as in modern gempylids (Potthoff et al. Citation1986; Nakamura and Parin Citation1993), with gemfish such as Rexea species typically having 34 (Roberts and Stewart Citation1997). Chapman noted the abdominal vertebral centra of Eothyrsites as being of similar dimensions to those in Thyrsites, but more robust.
Figure 6. Photograph of part of abdominal region of Eothyrsites holosquamatus Chapman, syntype OU 6329, lateral view.

Ribs are not well preserved, those present are moderately long and slender (∼4 mm wide) and angled caudally (seen in OU 6329).
Squamation: The body of Eothyrsites holosquamatus appears to have been covered in reasonably large (on average 4 mm but up to 7 mm diameter), robust cycloid scales. One piece of the specimen, OU 6329 b, seems to reveal a 50 × 50 mm portion of trunk covered by overlapping scales (J). These ovoid-shape scales have smooth margins, appear to lack prominent radii, and due to preservation the circular growth rings are only just visible. Chapman (Citation1935) noted that modern Thyrsites atun only has similar scales along its lateral line, the majority of its scales being very much reduced in size.
Pectoral fins: Eothyrsites had relatively large and elongate pectoral fins, which are still articulated in the specimen. According to Chapman these are narrow, slender, and only slightly curved, 70 mm in length, 21 mm at widest, as in the left pectoral figured as OU 6332. However, in addition, fragment OU 6854 a bears a second (right) fin, somewhat more complete distally (F). It is comprised of at least 10 rays, the longest being approximately 140 mm in length and reaching some 40 mm in width. The modern gemfish Rexea typically have 14 pectoral fin rays (Roberts and Stewart Citation1997). With the pectoral fins at 20% total body length, this is slightly longer than the pectoral fins of modern Thyrsites atun, and in this respect resembles those seen in the longfin escolar Scombrolabrax (Nakamura and Parin Citation2001). Unfortunately the pieces of Eothyrsites containing pectoral fin are broken away in the region of the pectoral girdle, although a small part (40 × 30 mm) of the coracoid or postcleithrum may be present on OU 6332.
Dorsal fin: Gempylids typically exhibit a notable spinous dorsal fin. We are uncertain if this is preserved in the specimen. Note that Johnson (Citation1986) considered the absence of supraneural bones as a synapomorphy of the Gempylidae (including Trichiuridae), Scombridae and billfishes (Istiophoridae and Xiphiidae). Gempylidae are characterised by having a spinous dorsal fin develop prior to the soft dorsal fin, seen in young larvae of the group (Johnson Citation1986). The dorsal-fin spines of the other gempylid genera, except Thyrsitops, are recorded as bearing serrations at least on their posterolateral keels (Carnvale Citation2006).
We note the presence of a small fin comprising ∼10 rays measuring up to 40 mm in length, seen in piece OU 6329 (). This may represent the anal or pelvic fin, or alternatively, it could represent a dorsal or anal ‘finlet’.
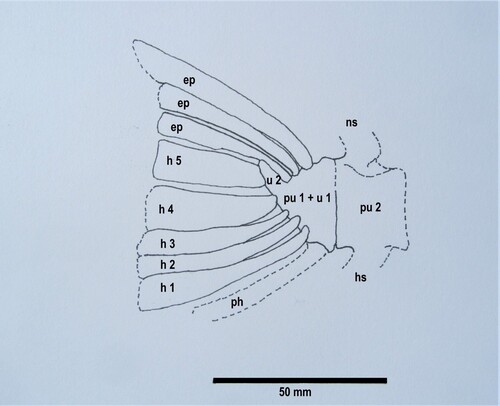
Caudal skeleton: Eothyrsites has a rather small forked caudal fin, the preserved endoskeleton (OU 6854 b) measuring approximately 80 mm from tip to tip of the lobes, and is comprised of articulated elements that become broader distally and once supported the principal rays (I). These distal rays and the procurrent rays are not present in the specimen. Despite being difficult to define due to preservation in Eothyrsites, three vertebrae once supported the fin rays. The preural and ural centra (urostyle = PU + U1) appear fused to the second ural centrum (U2) to form the terminal caudal compound centrum (). Three epurals articulate to the upper side of this, while the ventral margin supports at least three hypurals (H1–3). The specimen also suggests two slightly broader central hypurals (H4 + H5) that once supported the principal rays from their distal margin. The parhypural is not obvious and was perhaps preserved on the counterpart of the specimen. Overall a similar caudal arrangement to the specimen is seen (although not exclusively) in modern fishes of the Gempylidae. These exhibit a caudal skeleton where the overlying procurrent fin rays are usually supported by the neural and haemal elements of the three terminal centra (Potthoff et al. Citation1986).
Figure 7. Restoration of caudal skeleton of Eothyrsites holosquamatus Chapman, based on syntype OU 6854 b, lateral view.
Discussion
Some anatomical features of Eothyrsites appear similar to the extant barracuda (family Sphyraenidae), suggesting a relationship within the broader suborder Scombroidei. However, upon further study, the suspicion that Eothyrsites may have actually belonged to the Sphyraenidae (Rust Citation2000; King et al. Citation2009) is inaccurate. For example, the caudal skeleton does not suggest inclusion in Sphyraenidae. The modern barracuda Sphyraena has a caudal skeleton with fused hypurals, urostyle and uroneurals (Hollister Citation1937). Eothyrsites () does not exhibit this degree of caudal fusion into a reduced number of solid hypural plates, and in this respect also differs from the higher Scombridae and billfish (Istiophoridae and Xiphiidae).
In addition, the anterior and posterior large fang-like premaxillary symphysial teeth of the barracuda Sphyraena are somewhat compressed, bilaterally symmetrical and typically exhibit a coloured enamell on the cutting edges, or at a notable commissural barb found near the posterior apical portion of the tooth (Ballen Citation2020). This is in contrast with the smooth margins on the teeth of Eothyrsites. Also, besides having large fang-like but asymmetrical dentary symphysial teeth, the smaller dentary teeth of Sphyraena are notably lanceolate in profile and bear fine serrations (Ballen Citation2020), features not seen in Eothyrsites, although the teeth of the lower jaw are poorly preserved in our specimen.
We believe Chapman was correct in assigning the specimen to the Gempylidae within the Trichiuroidea. Obviously, there is difficulty in identifying some morphological characters in our specimen of Eothyrsites due to preservation, but it generally appears consistent with the family level diagnosis of Danilchenko (see diagnosis section). Danilchenko (Citation1960, Citation1967), Monsch and Bannikov (Citation2011) and Bannikov (Citation2017) also discuss several extinct genera of gempylids from the Oligocene of Europe, including Thyrsitocephalus (vom Rath Citation1859), which Chapman (Citation1934, Citation1935) compared to the Dunedin fossil. Thyrsitocephalus alpinus from Canton Glarus, Switzerland, was a much smaller fish (length 16 cm), the head being a fifth of the body length with rather small orbits and 12 recurved teeth (none mentioned as being enlarged as fangs) in each jaw. It had a large anterior dorsal fin with more than 20 spines, small paired fins and many finlets. Like Eothyrsites it had an elongate body (54 vertebrae) and a forked caudal fin, but scales are not known (vom Rath Citation1859). Another of these fossil gempylids, Hemithyrsites, is also a smaller fish but regarded as coeval with Promethicthyes (see discussion in Monsch and Bannikov Citation2011), although most of the fossil specimens are small/sub-adult/juvenile? Its vertebrae are typically twice as long as high, hence more elongate than those of Eothyrsites. Hemithyrsites has 17 caudal vertebrae, unfused hypurals, and a complete covering of cycloid scales (Danilchenko Citation1960, Citation1967; Monsch and Bannikov Citation2011).
The prominent cycloid scales are a distinctive character of the Burnside fossil. Chapman himself noted the contrast in scales present between Eothyrsites and modern Thyrsites, those in the latter being very much reduced (apart from those in the lateral line region). He expressed some of his thoughts on the loss of large scales as an example of a ‘degenerative process’ in Chapman (Citation1934). However, the bodies of adult specimens of modern gemfish, Promethicthyes prometheus and Rexea solandri in particular, are also entirely covered by small cycloid scales (Nakamura and Parin Citation1993; Roberts and Stewart Citation1997; Stewart Citation2015).
The fishes commonly named gemfishes are characterised (Roberts and Stewart Citation1997) by a slightly elongated body, a moderate number of vertebrae, the presence of supernumerary finlets on the caudal peduncle, a reduction of pelvic fins and the presence of small fangs on the lower jaw. The gemfish are also united in having two pairs of teeth on the vomer (Beckett et al. Citation2018). Certainly the skull and jaws (with anterior fangs and smaller posterior teeth) of Eothyrsites compare well with modern gemfishes. Like Thyrsites, large adult specimens of modern Rexea reach up to 1–1.5 m in length in New Zealand waters (Nakamura and Parin Citation1993; Stewart Citation2015). All have forked caudal fins.
Cranial characters for gempylids are taxonomically important but often difficult to distinguish in fossil specimens where the crania are often crushed (Beckett et al. Citation2018). Use of technology such as x-ray computed tomography is now aiding studies of early gempylid skull specimens from the Eocene London Clay for example (Beckett et al. Citation2018). Beckett et al. (Citation2018) also provide several detailed phylogenetic analyses of the Gempylidae (which they designate as a ‘grade’) and the closely related Trichiuridae, within the grouping Trichiuroidea. They do note a fossil skull from the London Clay, NHMUK PV OR 41318, as being similar to Rexea in exhibiting a narrowing of the frontals above the orbit and the pterotic ending before the posterior margin of the neurocranium. Unfortunately a lack of readily identifiable cranial characters in our specimen due to compression and thus far being only exposed from the underside, makes comparison and placement of Eothyrsites in their phylogenetic framework somewhat difficult at this stage. Further preparation and study of the skull in the future using novel techniques such as employed by Beckett et al. (Citation2018) should aid understanding of the species relationships.
In conclusion, Chapman’s establishment of Eothyrsites holosquamatus as a unique genus and species appears justified, however we suspect Eothyrsites represents an ancestral form of gempylid, perhaps more closely related to gemfish such as Promethicthyes or Rexea than Thyrsites.
Paleoecology
Deposition of the Burnside Mudstone at outer shelf depths suggests that like living gempylids, Eothyrsites was probably an epipelagic predatory species frequenting the upper 200 m of open waters (see Miya et al. Citation2013). Most modern gemfish frequent waters of 80–800 m depth (Roberts and Stewart Citation1997; Stewart Citation2015), and the larvae and eggs are pelagic. According to Bray and Schultz (Citation2020) Thyrsites atun in Australasian waters appears to prefer water temperatures between 13 and 18 degrees Celsius. It forms large schools, which feed, migrate and spawn together. A fast-growing species early in its life, it reaches almost 30 cm in length after the first year and 58 cm after three years. Details of the biology of Thyrsites, Rexea and other modern gempylids in New Zealand waters are discussed by Stewart (Citation2015). The strong upper jaw of modern gempylids allows them to feed on fairly large prey including fish and squid (Nakamura and Parin Citation1993; Stewart Citation2015).
The Paleogene represents an important time of increased radiation and diversification of teleost fishes. This is reflected in the phylogenetic tree of life data produced by comprehensive genetic studies, which has led to a more detailed understanding of fish classification (see Betancur-R et al. Citation2013, Citation2017). We see the first fossil record of many teleost groups in the early-middle Eocene deposits of Monte Bolca, Italy, and the London Clay (see reviews by Monsch Citation2005; Friedman et al. Citation2016; Beckett et al. Citation2018; Friedman and Carnevale Citation2018). This includes a number of epipelagic fish families, members of series Pelagiaria (Betancur-R et al. Citation2017), sometimes grouped together as a monophyletic clade under the classification ‘Pelagia’ (Miya et al. Citation2013). Some of this group perhaps originated from deep water ancestors to fill ecological niches available post KT extinction (Miya et al. Citation2013). They include fish belonging to a number of scombroid families including the Gempylidae which are recorded from the London Clay (Monsch Citation2005; Beckett et al. Citation2018). These pelagic fishes, along with other marine organisms, were able to spread rapidly across the globe via the Tethys seaway which remained open until the Miocene (Hou and Li Citation2018; Schwarzhans Citation2019).
The diverse fossil record of fish earbones (otoliths) provides information about many aspects of the New Zealand Cenozoic fish fauna where other remains are patchy: see Schwarzhans (Citation2019), Schwarzhans et al. (Citation2017). Eocene otoliths are recorded from North Otago and South Canterbury, including deeper water settings (∼200 m) similar to those at Burnside, the Bortonian age sediments exposed at Hampden Beach for example. Analysis of the otolith record also suggests linkages in the fish fauna with those of the Tethyan bioprovince during the Paleogene (Schwarzhans Citation2019). Later, increased circulation associated with the opening of the Tasman and Drake seaways and establishment of the Antarctic circumpolar current led to increased endemism in the southern ocean fauna (Schwarzhans Citation2019). Note however that the Eocene otolith record is relatively poor compared to the extensive and diverse collections made from post-Whaingaroan (Early Oligocene) strata.
We have been unable, as yet, to identify any otoliths associated with the remains of Eothyrsites. In the comprehensive review by Schwarzhans (Citation2019) only six representatives of the Scombroidei are recorded from the Latest Oligocene–Early Miocene of New Zealand, and these are not yet identified to family level. This is in contrast with the skeletal record where bony remains of large scombrids are seemingly over-represented in strata of this age, perhaps due to the presence of robust vertebrae and fused caudal elements (see Rust Citation2000).
It is possible that Gempylidae were only a minor component of the ancient fish fauna locally; only limited inferences can be made from the somewhat sparse fossil record. The presence of Eothyrsites importantly suggests the family had spread to the South Pacific and seas around Zealandia by the late Eocene. More gemfish remains, including otoliths, will hopefully be found in Neogene formations. Certainly the gemfishes, snake mackerels and cutlassfishes constitute a noteworthy and distinctive element present in the Australasian marine fish fauna today (Nakamura and Parin Citation1993; Stewart Citation2015).
The presence of predatory fish such as Eothyrsites suggest smaller fish and other prey were available locally in the Eocene, yet due largely to the predominant deposition of rather massive muddy sediment in deeper waters, fossil fish remains in general from the Eocene are relatively rare in New Zealand (Rust Citation2000). Given the relative paucity of teleost skeletal material recorded from the Cenozoic of Australia (J. Long pers. comm.), the specimens in the collections at the University of Otago Geology Museum, as previously noted by Chapman and Benson during the 1930s and 1940s, have the potential to contribute much to the study of fossil fishes from the Southern Hemisphere oceans.
Acknowledgements
Many years of fossil research, collection and curation at the University of Otago by Ewan Fordyce in particular have provided inspiration and information about the ancient life of the New Zealand region. The authors gratefully acknowledge Ewan Fordyce, Daphne Lee (OU) and Andrew Stewart (Te Papa) for their encouragement and assistance in this project. The reviewers are thanked for their constructive comments. Andrew Stewart also kindly reviewed an early stage of the manuscript. Stephen Read (OU) assisted with production of the locality map. Staff at the Hocken Library and Auckland Museum provided access to personal correspondence of key figures in Zealandian natural history. We also acknowledge the broad contributions Frederick Chapman and William Noel Benson made to Australasian paleontology.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Archey G. 1956. Personal letter correspondence with W. N. Benson, 22 February 1956. Auckland War Memorial Museum – Tāmaki Paenga Hira collection MUS-2008-22.
- Ballen GA. 2020. New records of the genus Sphyraena (Teleostei: Sphyraenidae) from the Caribbean with comments on dental characters in the genus. Journal of Vertebrate Paleontology. 40:11. doi:10.1080/02724634.2020.1849246
- Bannikov AF. 2017. New data on the morphology of the snake mackerel Abadzekhia Tarletskovi Bannikov (Perciformes, Gempylidae) from the Lower Oligocene of the Northern Caucasus. Paleontological Journal. 51:542–546. doi:10.1134/S0031030117050033.
- Beckett HT, Giles S, Johanson Z, Friedman M. 2018. Morphology and phylogenetic relationships of fossil snake mackerels and cutlassfishes (Trichiuroidea) from the Eocene (Ypresian) London Clay Formation. Papers in Palaeontology. 4(4):577–603. doi:10.1002/spp2.1221.
- Bemis KE, Burke SM, St John CA, Hilton EJ, Bemis WE. 2019. Tooth development and replacement in the Atlantic Cutlassfish, Trichiurus lepturus, with comparisons to other Scombroidei. Journal of Morphology. 280(1):78–94. doi:10.1002/jmor.20919.
- Benson WN. 1944. Obituary. Frederick Chapman, Hon. F.R.S.N.Z. Transactions and Proceedings of the Royal Society of New Zealand. 74:302–303.
- Benson WN. 1955. Personal letter correspondence with A. W. B. Powell, 19th December, 1955, Hocken library collection, University of Otago, Dunedin.
- Benson WN. 1956. Personal letter correspondence with E. I. White, 14 Nov 1956. Natural History Museum: Papers of the Department of Palaeontology Archive (as filmed by the AJCP) [microform]: [M2878-M2887] 1844–1986./Series DF100/File 100/82/Item. Correspondence with Professor W. N. Benson (Dunedin), [accessed November 2022]. TROVE. https/nla.gov.au.
- Benson WN. 1968. The W.N. Benson geological map of Dunedin district. Wellington: Department of Scientific and Industrial Research. Miscellaneous Series map / New Zealand Geological Survey 1. 1 folded map + 1 booklet, scale 1:50,000 Note: See p. 10 in compilation by Wood.
- Betancur RR, Broughton R, Wiley E, Carpenter K, López J, Li C, Holcroft N, Arcila D, Sanciangco M, Cureton J II, et al. 2013. The tree of life and a new classification of bony fishes. PLoS Currents. doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288.
- Betancur RR, Wiley EO, Arratia G, Acero A, Bailly N, Miya M, Lecointre G, Ortí G. 2017. Phylogenetic classification of bony fishes. BMC Evolutionary Biology. 17(1):162. doi:10.1186/s12862-017-0958-3. PMID: 28683774; PMCID: PMC5501477.
- Bishop DG, Turnbull IM. (compilers) 1996. Geology of the Dunedin area. Lower Hutt: Institute of Geological & Nuclear Sciences. Institute of Geological & Nuclear Sciences 1:250,000 geological map 21. 1 folded map + 52 p.
- Bray DJ, Schultz S. 2020. Thyrsites atun in fishes of Australia. [accessed 2022 Sep 22]. https://fishesofaustralia.net.au/Home/species/711.
- Carpenter KE, Collette BB, Russo JL. 1995. Unstable and stable classifications of scombroid fishes. Bulletin of Marine Science. 56(2):379–405.
- Carnvale G. 2006. A new snake mackerel from the Miocene of Algeria. Palaeontology. 49(Part 2):391–403. doi:10.1111/j.1475-4983.2006.00541.x.
- Chapman F. 1918. Descriptions and revisions of the cretaceous and tertiary fish-remains of New Zealand. New Zealand Geological Survey Paleontological Bulletin. 7:45.
- Chapman F. 1934. A common foodfish and its ancestors (1934, April 28). The Argus (Melbourne, Victoria: 1848–1957), p. 12; [accessed 15 November 2022]. http://nla.gov.au/nla.news-article10931892 reprinted in The Gisborne Times, Volume LXXX, Issue 12263, 26 May 1934, Page 7. https://paperspast.natlib.govt.nz/newspapers/GIST19340526.2.54.
- Chapman F. 1935* sometimes listed as 1934. Descriptions of fossil fish from New Zealand. Transactions and Proceedings of the Royal Society of New Zealand. 64:117–121.
- Crespin I. 1979. Chapman, Frederick (1864–1943), palaeontologist. In: Nairn B, Serle G, editors. Australian dictionary of biography. vol. 7. Melbourne: Melbourne University Press; p. 612. http://www.adb.online.anu.edu.au/biogs/A070621b.htm.
- Danilchenko PG. 1960. Bony fishes of the Maikop deposits of the Caucasus. Trudy Paleontol. Inst. Acad. Sci. USSR. 78:1–208
- Danilchenko PG. 1967. Bony fishes of the Maikop deposits of the Caucasus. Jerusalem: Israel Program for Scientific Translations. 247 p.
- Friedman M, Beckett HT, Close RA, Johanson Z. 2016. The English chalk and London clay: two remarkable British bony fish Lagerstätten. In: Johanson Z, Barrett PM, Richer M, Smith M, editors. Arthur Smith Woodward: his life and influence on modern vertebrate palaeontology. Geological Society, London: Special Publications, 430; pp. 165–200. doi:10.1144/SP430.18
- Friedman M, Carnevale G. 2018. The Bolca Lagerstätten: shallow marine life in the Eocene. Journal of the Geological Society. 175:569–579. doi:10.1144/jgs2017-164.
- Froese R, Pauly D, editors. 2022. Fishbase. World Wide Web Electronic Publication. www.fishbase.org, version (02/2022).
- Hamilton D. 1958. An Eocene sea-pen from Dunedin, New Zealand. Paleontology. 1(3):226–230.
- Hollister G. 1937. Caudal skeleton of Bermuda shallow water fishes. II. Order percomorphi, suborder percesoces: atherinidae, mugilidae, sphyraenidae. Zoologica: Scientific Contributions of the New York Zoological Society. 22(17):265–279. doi:10.5962/p.203682.
- Hou Z, Li S. 2018. Tethyan changes shaped aquatic diversification. Biological Reviews. 93(2):874–896. doi:10.1111/brv.12376.
- Johnson GD. 1986. Scombroid phylogeny: an alternative hypothesis. Bulletin of Marine Science. 39(1):1–41.
- King CM, Roberts CD, Bell BD, Fordyce RE, Nicoll RS, Worthy TH, Paulin CD, Hitchmough RA, Keyes IW, Baker AN, et al. 2009. Chapter 24: phylum chordata. In: Gordon DP, editor. New Zealand inventory of biodiversity, volume 1: Kingdom Animalia. Radiata,Lophotrochozoa, Deuterostomia. Christchurch, New Zealand: Canterbury University Press; pp. 431–555.
- Matsubara K, Iwai T. 1952. Studies on some Japanese fishes of the family Gempylidae. Pacific Science. 6(3):193–212.
- McMillan SG, Wilson GJ. 1997. Allostratigraphy of coastal south and east Otago: a stratigraphic framework for interpretation of the Great South Basin, New Zealand. New Zealand Journal of Geology and Geophysics. 40(1):91–107. doi:10.1080/00288306.1997.9514743.
- Miya M, Friedman M, Satoh TP, Takeshima H, Sado T. 2013. Evolutionary origin of the scombridae (tunas and mackerels): members of a paleogene adaptive radiation with 14 other pelagic fish families. PLoS ONE. 8(9):e73535. doi:10.1371/journal.pone.0073535.
- Monsch KA. 2005. Revision of the scombroid fishes from the Cenozoic of England Transactions of the Royal Society of Edinburgh. Earth Sciences. 95:445–489. doi:10.1017/S0263593300001164.
- Monsch KA, Bannikov AF. 2011. New taxonomic synopses and revision of the scombroid fishes (Scombroidei, Perciformes), including billfishes, from the Cenozoic of territories of the former USSR. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 102:253–300. doi:10.1017/S1755691011010085.
- Nakamura I, Parin NV. 1993. FAO species catalogue. Vol. 15. Snake mackerels and cutlassfishes of the world (families Gempylidae and Trichiuridae). An annotated and illustrated catalogue of the snake mackerels, snoeks, escolars, gemfishes, sackfishes, domine, oilfish, cutlassfishes,. scabbardfishes, hairtails, and frostfishes known to date. FAO Fish. Synop. 125(15):136.
- Nakamura I, Parin NV. 2001. Scombrolabracidae. In: Carpenter KE, Niem TH, editors. The living marine resources of the western central Pacific. FAO species identification guide for fisheries purposes. Rome: FAO; Vol. 6, p. 3684, pp. 3381–4218.
- Potthoff T, Kelley S, Javechi JC. 1986. Cartilage and bone development in Scombroid Fishes. Fishery Bulletin. 84(3):647–678.
- Powell AWB. 1956. Personal letter correspondence with W. N. Benson, 17 April, 1956. Auckland War Memorial Museum – Tāmaki Paenga Hira collection MUS-2008-22.
- Roberts CD, Stewart AL. 1997. Gemfishes (Scombroidei, Gempylidae, Rexea) of New Caledonia, southwest Pacific Ocean, with description of a new species. In: SfiRET B, editor. Resultatsdes campagnes MUSORSTOM, volume 17. Paris: Mem.Mus.natn.Hist,nat.; 174, pp. 125–141, ISBN2-85653-500-3.
- Rust S. 2000. Fossil bony fish (Teleostei) from Oligocene-Miocene marine sediments on southern New Zealand [MSc thesis]. University of Otago. 148p.
- Schäfer W. 1972. Ecology and paleoecology of marine environments. Chicago: University of Chicago Press. 568p.
- Schwarzhans W. 2019. Reconstruction of the fossil marine bony fish fauna (Teleostei) from the Eocene to Pleistocene of New Zealand by means of otoliths. Memoir of Museo di Storia Naturale di Milano 329p.
- Schwarzhans W, Lee DE, Gard HJL. 2017. Otoliths reveal diverse fish communities in Late Oligocene estuarine to deep-water paleoenvironments in southern Zealandia. New Zealand Journal of Geology and Geophysics. 60(4):433–464. doi:10.1080/00288306.2017.1365734.
- Stewart AL. 2015. Family gempylidae. Gemfishes. In: Roberts CD, Stewart AL, Struthers CD, editors. The fishes of New Zealand. Wellington: Te Papa Press, 4 volumes, 2002 p; pp. 1602–1615. ISBN: 978-0-9941041-6-8
- vom Rath G. 1859. Beitrag zur Kenntniss der fossilen Fische des Plattenberges im Canton Glarus. Zeitschrift der Deutschen Geologischen Gesellschaft. 11:108–132.
- White EI. 1956. Personal letter correspondence to W. N. Benson 24 November 1956. Natural History Museum: Papers of the Department of Palaeontology Archive (as filmed by the AJCP) [microform]: [M2878-M2887] 1844-1986./Series DF100/File 100/82/Item. Correspondence with Professor W.N. Benson (Dunedin)/Correspondence with Professor W.N. Benson (Dunedin). TROVE. https/nla.govt.au.