Abstract
Resighting of adult Sooty Terns Onychoprion fuscatus that had been ringed on Bird Island, Seychelles, as chicks in 2002 indicated that birds of higher body mass at fledging had higher post-fledging survival than lighter birds. Resighting of birds that had been ringed at the same time, but at different developmental stages, suggested that birds that fledged from eggs laid late had lower post-fledging survival than chicks that fledged from eggs laid at the peak of the nesting season. These findings are discussed in relation to factors that influence chick survival within the colony and post-fledging survival in the marine environment, along with their implications for the commercial harvest of Sooty Tern eggs in Seychelles.
In Seychelles, egg laying by Sooty Terns Onychoprion fuscatus is highly synchronous (Feare Citation1976a, Jacquemet et al Citation2007), as evidenced by 75% of the estimated 395,000 eggs on Bird Island being laid within a nine-day period in the 1973 breeding season (Feare Citation1976a), the only year that the pattern of laying has been closely followed. A corollary of synchrony is that pairs that laid during peak egg-laying reared more chicks to fledging than those that laid early or late in the season (Feare Citation1976a). Three factors could account for this: social behaviour within the colony, involving aggression of adults and larger young to smaller chicks that enter their territories (Feare Citation1976a), a social component in the location of feeding opportunities that are unpredictable in space and time (Ashmole Citation1963), and a seasonal effect of food availability, which Jacquemet et al Citation(2007) demonstrated from remote sensing of sea-surface temperatures and chlorophyll concentrations in surface waters around Seychelles.
Young Sooty Terns undergo a long period of deferred maturity (Schreiber et al Citation2002) during which they have not been recorded in the Bird Island colony. On Bird Island we have found one three-year-old bird incubating, but most do not return to breed until five years old; all appear to do so by six years old (Feare & Doherty Citation2011). During this period they are suspected to travel widely in the Indian Ocean but we have only two ring recoveries, one in southern India and the other in northern Australia. Sooty Terns ringed as chicks on the point of fledging on Bird Island in 1972 and 1973, and recaptured as breeding adults between 1994 and 2002, permitted examination of pre-breeding survival in relation to body mass at fledging and time of fledging. That analysis suggested that survival into adulthood was not related to body mass at fledging, but showed that a greater proportion of chicks survived from birds that fledged from eggs laid in the middle of the breeding season compared with eggs laid earlier or later (Feare Citation2002). However, the sample of weighed fledglings that were subsequently recaptured, when over 20 years old, was small (n = 20), highlighting the need for more data. Further ringing of chicks since 1993 has proved logistically difficult in the Bird Island colony but in 2002 it was possible to ring a large number and weigh those chicks on the point of fledging.
In Seychelles, Sooty Tern eggs are regarded as a much-desired delicacy and are harvested commercially from some colonies under regulation by the Environment Department (Feare Citation1976b, Feare & Gill Citation1997). Commercial harvest involves the daily removal of eggs from specified areas of a colony in order to guarantee the freshness of the eggs. A pair of adults that loses its egg returns to lay a replacement c 14 days later, although the proportion of birds that will return to lay replacement eggs declines as the season progresses (Feare Citation1976a).
In 2002, all eggs laid in the Bird Island colony were harvested up to 17 June. This prevented birds from progressing from laying to incubation until two or three weeks after what would normally have been the peak of laying, had the colony been undisturbed. Most of the eggs laid after the cessation of the harvest were likely to have been replacement eggs although late-arriving pairs may also have laid at this time. At the termination of egg collection many birds would have been ready to lay or re-lay but birds that hatched from eggs laid this late in the season, in relation to the onset of laying, in 1973 experienced poor survival within the colony (Feare Citation1976a) and post fledging (Feare Citation2002). The artificial delay of the nesting period in 2002 provided an opportunity to investigate the effect of enforced later nesting on the survival of young to breeding status, and thus potentially to explore the relative importance of environmental seasonality (late breeding) and the birds' social behaviour (opportunities for local enhancement of feeding when large numbers of fledglings leave the colony simultaneously or greater protection from predators – at sea this is most likely to be from predatory fish when the birds are feeding; Feare Citation1976a) on the subsequent survival of young birds. If seasonality were the critical factor we would predict that late nesting in 2002 would result in poorer post-fledging survival than in 1973 birds and a progressive decline towards failure in the later fledging 2002 birds. If social behaviour were important, through more synchronised departure from the colony at fledging, we would predict that fledging success should remain high throughout the period when large numbers were fledging and to decline thereafter. This paper reports the findings from the 2002 cohort.
The overall aim of this long-term study of Seychelles Sooty Terns has been to provide advice on ensuring sustainability of the annual egg harvest. The results of this part of the investigation will be briefly discussed in this light.
METHODS
Sooty Tern chicks were ringed on Bird Island (3°43'S 55°12'E), where the colony size in 2002 exceeded 300,000 pairs (CF, pers obs). All chicks were ringed in central parts of the colony, where Feare Citation(1976a) had found breeding success to be higher than on the periphery and thus where ringed birds were more likely to fledge. Between 27 August and 4 September 2002, 1,550 Sooty Tern chicks, covering the range of plumage stages that were present in the colony at the time, were given numbered alloy British Trust for Ornithology rings. Plumage stage provides an indication of the age of the chicks after hatching and each chick was assigned to one of the developmental stages described by Feare (Citation1976a, Figs 3 & 13): Small Wings (SW, c 10–20 days old), Head Moulting (HM, 19–34), Down on Face (DF, 29–40), Tail Cocked (TC, 37–46), Short Tail (ST, 40–55) and Tail Forked (TF, 46+).
Sooty Tern chicks are fed by their parents up to and after their departure from the colony (Feare Citation1975) and thus do not lose body mass before fledging. Asymptotic mass can be difficult to establish because a chick's body mass will depend upon the size of and time elapsed since its last meal, which can account for up to 10% of the body mass of ST and TF chicks (Feare Citation1976a). In 1973, however, chicks reached their maximum body mass at 40–50 days old, at which age they attained plumage class ST (Feare Citation1976a). In 2002 chicks classed as ST and TF which had likely attained their asymptotic body mass, were weighed to the nearest 5 g using a 300-g Pesola spring balance and we assumed that, with reasonable samples of recaptures when birds returned to breed, the variance of mass due to these factors would be the same for birds relocated as breeders in the colony and those not recaptured. Smaller chicks (SW, HM, DF, TC) were not weighed as their masses are highly variable, depending on the time of weighing in relation to the time of their last feed, as meal mass can be higher in relation to body mass (Feare Citation1976a). At the time of ringing most chicks in the colony were HM, DF and TC (), covering an age range of three to four weeks and suggesting that the main exodus of fledglings from the colony would have spanned approximately this period. Fledging occurs 60–70 days after hatching (Feare Citation1976a) and these birds would be expected to have left the colony in large numbers from late September and through October 2002 (cf late August and through September in 1973).
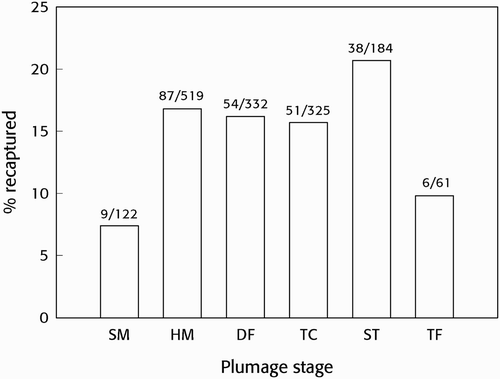
Figure 1. The proportion of Sooty Tern chicks at different plumage stages (see text) when ringed that were subsequently recaptured as breeding adults in the Bird Island colony. Figures above the columns are numbers recaptured and ringed.
Searches for ringed birds were made during incubation, in June, annually from 1994 to 2012, as described by Feare & Doherty (Citation2004, Citation2011). Search effort has varied annually due to unpredictable variations in the time of egg-laying and to logistical constraints, leading to annual variations in recapture probability, which have ranged from 6% to 25%; the highest recapture probabilities were achieved during 2003–06 (Feare & Doherty Citation2011). All ringed birds seen were caught using small hand nets and their ring numbers were recorded.
Statistical analyses were undertaken using R version 2.15.1 (R Core Team Citation2012). Data distributions and variances were checked prior to analyses; data were normally distributed (W > 9.97, P > 0.22) with equal variances (F = 1.02, P = 0.89) confirming the assumption made above relating to variances of samples of birds that were and were not recaptured as breeding adults. We used a Pearson's χ2 test to determine whether there were differences in the proportions of recruits in the different ringing stages and then we used ‘prop.test’ in R to see which stages had significantly different proportions of chicks resighted as adults.
RESULTS
Forty-two of 234 Sooty Tern chicks ringed at plumage stages ST and TF in 2002 were recaptured between 2005 (three years old) and 2012 (ten years old). The body mass at fledging of the recaptured birds (196.0 ± 2.7 g, mean ± se) was significantly greater than the body mass of chicks that were not recaptured (188.8 ± 1.2 g; two-sample t-test, t 232 = 2.47, P = 0.014).
To date, 245 of the 1,543 chicks ringed in 2002 (15.9 %) have been recaptured as breeding adults in the Bird Island colony. The distribution of individuals recaptured () showed significant differences between their plumage categories at ringing (Pearson's χ 2 test, χ 2 10 = 47.05, P < 0.001). Pairwise comparisons among the plumage categories showed that fewer chicks from the youngest group at the time of ringing (SW) were recaptured than from all of the other categories (prop.test χ 2, P < 0.01) other than TF, and that significantly more ST chicks were recaptured than TC (prop.test χ 2 = 5.12, P = 0.02) (). The proportion of the oldest and most developed group of chicks at ringing (TF) recaptured was not significantly different from those of other categories (but approached significance with ST: P = 0.054). This result might be due to the small samples of TF chicks ringed and recaptured. This analysis thus suggests that fewer very young chicks (SW) returned to breed than other categories and that, while it appears () that fewer of the oldest (TF) birds returned, this requires larger sample sizes for confirmation.
DISCUSSION
Data obtained from the chicks ringed in 1972–73 were suggestive of a positive relationship between body mass at fledging and survival to return as breeding adults but, with small sample sizes, did not attain statistical significance (Feare Citation2002). Data from the cohort of chicks ringed in 2002 and subsequently recruited to the Bird Island breeding population indicate that body mass at fledging did influence survival to recruitment in these birds. The greater strength of the statistical evidence with the birds ringed in 2002 could be simply a consequence of the much larger sample size of recaptured birds for analysis than had been available for the 1972–73 cohorts. However, an additional factor could have been the enforced delay (by egg collection) in the 2002 breeding season; it is possible that body mass at fledging could be more important to post-fledging survival in years when breeding is delayed due to egg collection, than in years when the colony has not been disturbed. A further caveat is that failure to return to the colony includes an element of emigration in addition to mortality. While we know that many returning recruits to Seychelles colonies are philopatric, two young Sooty Terns from Bird Island have been found nesting in nearby colonies (pers obs). The extent of natal dispersal from Bird Island is unknown, however, due to the lack of searches for ringed birds in other colonies, but there is no reason to believe that the amount of emigration was greater in the early 2000s than it had been in the 1970s. It is also possible that lighter young at fledging may be more prone to move to other colonies (Selonen & Hanski Citation2010) or to commence breeding later, but much more ringing, and in particular much more searching of other colonies, will be required to test this possibility of condition-dependent natal dispersal and deferred maturity in Seychelles Sooty Terns. Natal dispersal might also be stimulated by disturbance associated with egg harvesting (Feare & Lesperance Citation2002) but this has yet to be demonstrated.
Here we used plumage stage to estimate the ages of the chicks we ringed, whereas Feare Citation(2002) reported birds that had been ringed at the point of fledging. Most chick mortality in the colony occurs within the first 10 days after hatching, with deaths due largely to attacks by neighbouring adults and larger chicks (Ashmole Citation1963, Feare Citation1976a). Mortality from this source is most intense when chicks are small and unable to run quickly through neighbouring nest territories following disturbances. Consequently, this source of mortality declines later in the season as older chicks fledge and the parents of older chicks spend less time in the colony. Among the 2002 chicks ringed in this study, the smaller SW, HM and DF chicks would have spent roughly three to five weeks in the colony after ringing but, with ST and TF chicks leaving the colony when fledging, and the density of birds remaining in the colony declining, the pressure on the younger birds from older chicks and their parents would have been diminishing. Although many of the 2002 chicks were ringed some weeks before fledging, a search of the colony on 3 November 2002 revealed 177 dead and weak chicks; six of these had been ringed (Marinette Assary, pers comm), all at SW and HM plumage stages. This suggests that larger chicks had fledged successfully and that the pattern of recruitment () reflected variations mainly in post-fledging survival and had not been due to pre-fledging mortality of later chicks in the colony. In the present analysis, fewer SW chicks (7.3%) returned to breed than other plumage categories (16.0–20.7%, excluding TF), indicating that chicks from eggs laid late in the 2002 season fared less well than earlier chicks. However, although a greater proportion of ST chicks returned than TC, there was no evidence of a steady increase in survival with earliness of fledging, suggesting that the delay in the adults' ability to nest successfully in 2002 had little effect on post-fledging survival of most young.
Within the breeding colony, the high degree of synchrony of laying appears to benefit chicks that hatch from eggs laid at the peak of laying, which survive to fledging better than birds that hatch later (in 1973 eggs that were laid early were all predated before hatching). This enhanced survival was due, at least in part, to chicks at the same developmental stage as neighbours experiencing less aggression than chicks, especially smaller chicks, that were out of phase with neighbours; predation is unlikely to have been involved as, apart from man, no predators take significant numbers of eggs or chicks in the Bird Island colony (Feare Citation1976a). In 1973, the availability of food for chicks approaching fledging became more variable (probably weather-related in this year; Feare Citation1976a), leading later chicks to take longer to fledge and to fledge at a lower body mass than earlier chicks. This suggested that nesting early provided benefits additional to those conferred by nesting synchronously. Little is known of the post-fledging life of juveniles (Schreiber et al Citation2002, Jacquemet Citation2010) but once young fledge they will be freed from geographical restriction to food supplies around the breeding colony and able to move to more remote locations of food abundance, as long as they can locate such food sources. In this, social behaviour, in the form of local enhancement and/or information exchange (Shaeler Citation2002), will facilitate feeding opportunities. Fledging and dispersal from the colony in large numbers may maximise the efficiency of these social behaviours in food procurement, at a time when young are learning to feed independently of their parents, and enhance the survival of young at peak fledging time. In so doing they can escape seasonal constraints that affect food supplies close to the breeding colony. This would promote the pattern of post-fledging survival observed in the 1972–73 cohorts and could explain the apparent lack of an age-related decline in post-fledging survival in most plumage categories in the 2002 cohort of Sooty Terns reported here. However, this can be confirmed only by much more intensive ringing and searching for ringed birds.
The relevance of these findings to the design of the egg-harvesting strategy in Seychelles must not be overestimated. Only two groups of ringed chicks, 1972–73 and 2002, have been available for analysis. The failure to detect a strong seasonal decline in the post-fledging survival of chicks in both cohorts is insufficient to conclude that post-fledging survival is independent of the time of year during which birds are able to breed in harvested colonies. Crucially, the findings of this study do not provide evidence to support a return to the scheme adopted in the 1960s, where all eggs were allowed to be taken up to 15 July (Ridley & Percy Citation1958), in the expectation that eggs laid after this date would produce sufficient young to recruit to ensure population stability (Feare Citation1976b). To ensure sustainability of the annual egg harvest, further study is needed. This should include further experimental investigation of the effects of harvesting early eggs on the success of later breeding attempts, and studies of natal dispersal in both protected and harvested colonies. Investigations should also be extended to include the lifetime reproductive success of birds that are recruited to breeding colonies, and whether this is related to seasonal events within the breeding colony or to repeated egg harvesting. Whether such an intensive long-term study is feasible, logistically and financially, remains to be seen.
Acknowledgements
We are grateful to the owners of Bird Island, Guy and Marie France Savy, and to managers Georges and Margaret Norah and Serge Robert for their approval of the work and for logistical support, and to Bird Island for funding CF's second visit to ring chicks in 2002. The process of catching and ringing chicks was valuably helped by Simon Feare, James Millett and Luciana Vega. Subsequent searches for the ringed birds have been assisted by Seychelles Environment Department staff Wilna Accouche, Majella Athenase, Perley Constance, Ronley Fanchette, Davidson Jaques, Josianna Rose, Terrence Valentine, Marie France Savy, Robbie Bresson, Elsa Richard, and by volunteers Katrina Cook, Naomi Doak, Adam and Nikki Feare, Simon Feare, Georgia French, Christoph Kueffer, Lilian Kronauer, Christine Larose, Matthieu Le Corre, Eva Schumacher and Jon Taylor. The Ringing Office of the British Trust for Ornithology kindly allowed us to use BTO rings for this study. Manuscript revision was helped by the comments of two referees.
References
- Ashmole , N. P. 1963 . The biology of the Wideawake or Sooty Tern Sterna fuscata on Ascension Island . Ibis , 103b : 297 – 364 . (doi:10.1111/j.1474-919X.1963.tb06757.x)
- Feare , C. J. 1975 . Post-fledging parental care in Crested and Sooty Terns . Condor , 77 : 368 – 370 . (doi:10.2307/1366255)
- Feare , C. J. 1976a . The breeding of the Sooty Tern Sterna fuscata in the Seychelles and the effects of experimental removal of its eggs. Journal of Zoology, London . 179 : 317 – 360 .
- Feare , C. J. 1976b . The exploitation of Sooty Tern eggs in the Seychelles . Biological Conservation , 10 : 169 – 181 . (doi:10.1016/0006-3207(76)90033-1)
- Feare , C. J. 2002 . Influence on date and body mass at fledging on long-term survival of Sooty Terns Sterna fuscata . Marine Ornithology , 30 : 46 – 47 .
- Feare , C. J. and Doherty , P. F. 2004 . Survival estimates of adult Sooty Terns Sterna fuscata from Bird Island, Seychelles . Ibis , 146 : 175 – 180 . (doi:10.1111/j.1474-919x.2004.00288.x)
- Feare , C. J. and Doherty , P. F. 2011 . Age at first breeding and pre-breeding survival in Seychelles Sooty Terns Onychoprion fuscatus . Marine Ornithology , 39 : 221 – 226 .
- Feare , C. J. and Gill , E. L. 1997 . The sustainable exploitation of Sooty Tern eggs in the Seychelles 5th annual report to the Seychelles Division of Environment. 18 pp
- Feare , C. J. and Lesperance , C. 2002 . Inter- and intra-colony movements of breeding adult sooty terns . Waterbirds , 25 : 52 – 55 . (doi:10.1675/1524-4695(2002)025[0052:IAIMOB]2.0.CO;2)
- Jacquemet , S. 2010 . Postfledging parental care and unexpected foraging behaviour of Sooty Terns Onychoprion fuscata in the southern Mozambique Channel . Ostrich , 81 : 271 – 272 . (doi:10.2989/00306525.2010.519862)
- Jacquemet , S. , Le Corre , M. and Quartly , G. D. 2007 . Ocean control of the breeding regime of the sooty tern in the southwest Indian Ocean . Deep Sea Research , 54 : 130 – 142 . (doi:10.1016/j.dsr.2006.10.003)
- R Core Team . 2012 . R: a language and environment for statistical computing , Vienna, Austria : R Foundation for Statistical Computing . http://www.R-project.org/
- Ridley , M. W. and Percy , R. 1958 . The exploitation of seabirds in Seychelles . Colonial Research Studies , 25 : 1 – 78 .
- Schreiber , E. A. , Feare , C. J. , Harrington , B. , Murray , B. , Robertson , W. B. , Robertson , B. and Woolfenden , G. E. 2002 . “ Sooty Tern Sterna fuscata ” . In The birds of North America , Edited by: A. , Poole and F. , Gill . 665 Philadelphia, PA : The Birds of North America, Inc .
- Selonen , V. and Hanski , L. K. 2010 . Condition-dependent, phenotype-dependent and genetic-dependent factors in the natal dispersal of a solitary rodent . Journal of Animal Ecology , 79 : 1093 – 1100 . (doi:10.1111/j.1365-2656.2010.01714.x)
- Shaeler , D. A. 2002 . “ Foraging behaviour and food of seabirds ” . In Biology of marine birds , Edited by: E.A. , Schreiber and J. , Burger . 137 – 177 . Boca Raton , Florida : CRC Press .