Abstract
Climate change has been shown to affect the distribution of many bird species. International Waterbird Census (IWC) data revealed that Tufted Duck Aythya fuligula and Goldeneye Bucephala clangula have shifted their European wintering distributions northeastwards as a direct response to increased winter temperatures. We tested whether it was possible to detect this shift in the wintering distribution of these species as well as Pochard Aythya ferina, using ringing and recovery data. Goldeneye conformed to our hypothesis, with its recovery distances decreasing significantly by about 400 km from the southwest to northeast from 1950 onwards, but this change could not be linked to winter temperatures. Conversely, Tufted Duck directly violated our hypothesis with its recovery distances increasing significantly by about 400 km but again this change could not be explicitly linked to winter temperatures. Pochard in contrast exhibited no change in its recovery distances. Thus, of the three species examined, ringing recoveries showed no trend in one species and of the two species where migratory short-stopping has been well described, one exhibited contraction whilst the other an apparent expansion in recovery distances. We conclude that ringing and recovery data provide little utility in detecting changes in migratory behaviour of these waterbirds.
In the European Union, considerable effort has been expended on establishing site safeguard networks to protect migratory waterbirds along their flyways, for example, through the designation of Natura 2000 sites under the EU Birds Directive (eg Stroud et al Citation1990) and the designation of Wetlands of International Importance under the Ramsar Convention. This is particularly the case for species that are subject to hunting, such as wildfowl, following concerns about their overexploitation in the 1930s (Berry Citation1941). The wintering distribution of many bird species is plastic and particularly responsive to climate (Austin & Rehfisch Citation2005, La Sorte & Thompson Citation2007). In former times, severe winter weather in northern Europe resulted in waterbirds shifting their wintering distribution south and west within their normal distributions (Ridgill & Fox Citation1990). In contrast, in recent years, wading birds have shown shifts in the centres of their winter ranges north and east during a period of milder winters (Maclean et al Citation2008). Such a trend is likely to challenge the effective maintenance of protected area networks, as birds resort to wetlands formerly unsuitable or unavailable to them in winter to the north and east of their range as these become increasingly ice-free. Concurrently, by decreasing the migration distance along flyways to wintering grounds, given a stable population size, birds will reduce or abandon the use of formerly important wetlands at the southwestern edge of their range.
During the last three decades, International Waterbird Census (IWC) data have revealed that Tufted Duck Aythya fuligula and Goldeneye Bucephala clangula shifted the centres of gravity of their wintering ranges northeastwards, correlated with a 3.8 °C increase in early winter temperatures in the northeastern part of their wintering range linked to climate change (Lehikoinen et al Citation2013). We tested whether it was possible to detect this shift in the wintering distribution of both Tufted Duck and Goldeneye, as well as Pochard Aythya ferina, using ringing and recovery data. Point-source marked individuals were used to determine if there was a reduction in the recovery distances over the period between 1950 and 2010 amongst ducks migrating south and west to their wintering grounds. The effects of climate change may not necessarily be linear with time. Thus, we also tested the hypothesis that the mean recovery distances were related to mean daily minimum winter temperatures at the wintering grounds or the difference in mean minimum temperatures between the breeding and wintering grounds each year.
METHODS
Ringing and recovery data for Pochard, Tufted Duck and Goldeneye were obtained through the data bank of the European Union for Bird Ringing (EURING) from all European countries with the exception of Russia. All the data were coded in the EURING 2000+ code (du Feu et al Citation2009). To select only individuals from populations with a known breeding origin, we included in the analysis only birds ringed during the breeding season (May, June and July) and recovered during the wintering season (defined here as November, December, January and February). It was assumed that adult ducks present at a particular location during the breeding season were breeding nearby, although it is possible that some males ringed in July may have been undergoing a moult migration after breeding at a different location. However, it was deemed that these probably represented only a small proportion of the ringing data set.
Records were discarded from the analysis to remove potential sources of bias if i) the primary identification method of an individual was not a metal ring, birds were ii) hand-reared, iii) held in captivity for more than 24 hours, iv) manipulated or transported, v) dates and locations of ringing and recovery were not recorded accurately (more than two weeks for date and >50 km for location) and vi) the distance between ringing and recovery sites was <100 km. The recovery distance was calculated according to the loxodrome (constant compass direction) method (EURING Citation2010). Recovery circumstances were described as a) shot, b) intentionally taken by man using other means, c) taken accidentally and d) taken by other methods. Recoveries were further categorised as i) direct, which were recoveries of birds ringed in the breeding season and recovered in the subsequent winter, and ii) indirect, which included recoveries of birds ringed during the breeding season and recovered in any winter after the first winter. The age of each bird at ringing was classed as either a duckling or adult, where a duckling was hatched during the breeding season in which it was ringed and an adult was at least one year old when ringed.
We selected Latvia as a source for breeding data for Pochard and Tufted Duck and Finland for Goldeneye because these species were ringed in large numbers in the second half of the 20th century in those countries. Latvia and Finland have been exclusively breeding grounds for those species, unlike countries further southwest which may act as both breeding and wintering grounds for different populations. Only records of individuals recovered from a bearing between 180° and 270° from their breeding grounds were retained for analysis, to ensure that movements were toward their wintering grounds.
The mean daily minimum winter temperature (November to February) was extracted for the centroid of the wintering grounds for each species and daily minimum temperature (May to July) was extracted for the breeding grounds. Temperature data were downloaded from the European Climate Assessment & Dataset website (http://eca.knmi.nl/dailydata/index.php) and the weather station closest to either the mean ringing location or the mean wintering location was selected for each species. For Pochard, this was Liyepaya AMSG, Latvia (56°33.00'N 21°1.12'E), on the breeding grounds and Beauvais-Tille, France (49°26.48'N 2°7.42'E), on the wintering grounds; for Tufted Duck, Liyepaya AMSG, Latvia, on the breeding grounds and Bad Lippspringe, Germany (51°47.13'N 8°50.23'E), on the wintering grounds; and for Goldeneye, Jyvaskyla Lentoasema, Finland (62°24.08'N 25°40.43'E), on the breeding grounds and Copenhagen, Landbohojskolen, Denmark (55°41.00'N 12°32.00'E), on the wintering grounds. The relative difference in the temperatures between the mean ringing and wintering location was also calculated.
Temporal trends in the mean minimum winter temperatures for the centroid of the wintering range of each species were examined using linear regressions. Change in recovery locations over time was visualised by plotting the year of each recovery and spatially interpolating missing areas using the ‘kriging’ function in the Spatial Analyst toolbox for ArcGIS 10.1 (ESRI, California, USA). Evidence of migratory short-stopping would be taken as a southwest to northeast trend progressing from old to recent recoveries indicating that the southwestern range edge was contracting and the northeastern range edge expanding, consistent with warming winter temperatures. Variation in recovery distances was examined for i) direct recoveries, and ii) all birds (direct and indirect recoveries) for each species using General Linear Models (GLMs). Recovery year was fitted as a covariate to test for change in the recovery distance and sex was fitted as a fixed factor as some species undergo differential migration of the sexes (Nichols & Haramis Citation1980, Carbone & Owen Citation1995, Wernham et al Citation2002), so females might be expected to migrate further than males. Age at ringing (duckling or adult) was also fitted as a fixed factor but only in models utilising direct recoveries because, for direct and indirect recoveries analysed together, a bird ringed as a duckling might be either a first-winter bird or an adult when recovered. Days since ringing, winter temperatures and the difference in temperature between the breeding and wintering sites were fitted as covariates. All possible model permutations were created and ranked using Akaike Information Criterion (AIC) values. The Akaike weight (ω i ) of each model was calculated within the top set of N models, where ΔAIC (Δ i ) ≤ 2 (Burnham & Anderson Citation2002). The Akaike weight of each model is the relative likelihood of that model being the best within a set of N models. To calculate the importance of each variable relative to all other variables, the Σω i of all models within the top set of models that contained the variable of interest was calculated and the variables ranked by Σω i (McAlpine et al Citation2006); the larger the value of Σω i (which varies between 0 and 1), the more important the variable. Multimodel inference and model averaging were used to determine effect sizes (β coefficient) of each variable across the top set of models (Burnham & Anderson Citation2002). Prior to model fitting, all predictor variables were tested for multicollinearity using ordinary least squares regression to ensure that all tolerance values were >0.1 and all variance inflation factor values were <10.0 (Quinn & Keough Citation2002). To allow the direct comparison of regression coefficients within models, variables were standardised to have mean x = 0 and σ = 1 prior to analysis. All statistical analyses were conducted using Genstat 15. The recovery distances of those species exhibiting significant temporal trends were plotted against time using Sigmaplot 10.
RESULTS
After the application of the selection criteria, 564 records were included in the analysis for Pochard, 230 (41%) of which were direct recoveries; 301 records were included for Tufted Duck of which 95 (32%) were direct recoveries and 181 records were included for Goldeneye of which 31 (17%) were direct recoveries. The distribution of sex and age classes among the birds ringed between May and July, 1950–2010, is given in . The majority of birds recovered had been shot ().
Table 1. Numbers and percentages for a) the recovery circumstances and b) sex and age classes for Pochard, Tufted Duck and Goldeneye ringed between May and July and recovered between November and February, 1950–2010.
In Latvia, most Pochard were ringed on Lake Engure, reflected in the mean ringing position (57°13.219'N 23°18.304'E). The mean recovery position for Pochard was in northern France (49°50.802'N 2°12.694'E) but the recoveries were dispersed throughout western Europe. Winter temperatures at this location exhibited a significant positive trend from 1950 to 2010 (F 1,59 = 10.590, P = 0.002; , but no spatiotemporal pattern was discernible in the recoveries of Pochard throughout its wintering range (. There was generally poor support for the influence of any variable in describing variation in Pochard recovery distances whilst the effects of year, winter temperatures and the difference of temperatures between the breeding and wintering grounds were inconsistent between direct and indirect recoveries (.
Figure 1. Trends in the mean daily winter (Nov–Feb) temperatures at a) Beauvais-Tille, France, b) Bad Lippspringe, Germany, and c) Copenhagen, Denmark, corresponding to the mean centroids of ring recoveries during winter for d) Pochard, e) Tufted Duck and f) Goldeneye respectively. Ringing recoveries are shown as closed circles and their dates of recovery (coloured scale) range from the 1950s (dark blue) to the 2000s (dark red) and have been spatially interpolated using kriging to illustrate any trend. Evidence of migratory short-stopping would be taken as a southwest to northeast trend progressing from red to blue indicating that the southwestern range edge was contracting and the northeastern one expanding, consistent with global climate change and observed positive trends in winter temperatures.
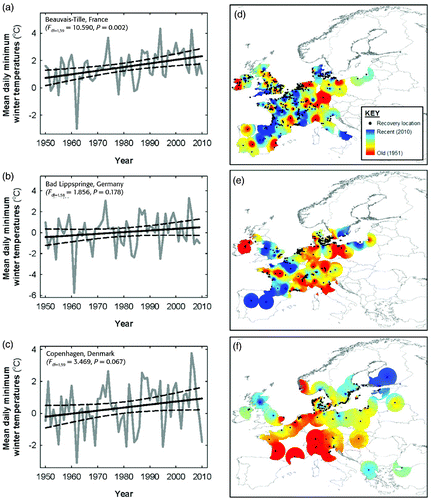
Figure 2. Relative importance of explanatory variables in explaining variation in the recovery distance of a) Pochard, b) Tufted Duck and c) Goldeneye using i) direct recoveries only and ii) direct plus indirect recoveries. Variables are ranked in order of the sum of their Akaike weights (Σωi ) within the top set of models, ie models with ΔAIC≤2. Black bars indicate those variables that were retained in the best single approximating model (ie that with the lowest AIC value) and grey bars indicate variables included in all other models within the top set. Variables judged significant at P<0.05 in the top model are shown in bold where ** indicates P<0.01 and *** P<0.001. Notation to the right indicates the strength of the slopes for each standardised covariate.
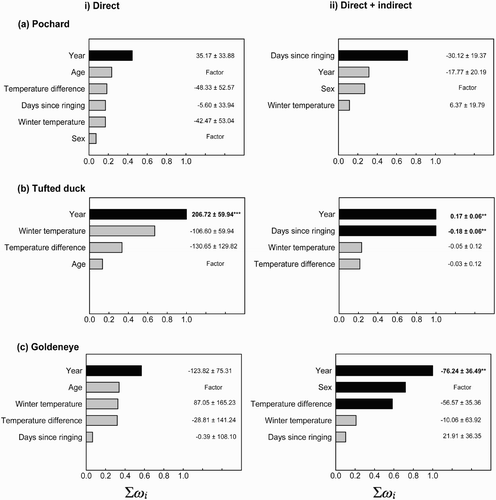
In Latvia, the mean ringing location for Tufted Duck was 57°6.042'N 24°9.177'E. The mean recovery position for Tufted Duck was in northwestern Germany (51°51.505'N 7°47.527'E) but recoveries were dispersed throughout western Europe. There was no significant change in winter temperatures between 1950 and 2010 at this location but there was a weakly positive trend (F 1,58 = 1.856, P = 0.178; . However, there was an apparent trend in the spatial location of recoveries with more recent recoveries tending to be near the southwestern edge of the wintering range in Iberia, southwestern and northeastern France and Great Britain (. Thus, there was a significant positive trend in the recovery distances of Tufted Duck with year using both direct and all recoveries ( with the strongest trend exhibited by direct recoveries (, b). On average, the recovery distance of Tufted Duck extended by approximately 400 km between 1950 and 2010 (. Tufted Duck recovery distances were negatively influenced by days since ringing, suggesting that the longer the period between ringing and recovery the more likely the bird was to be recovered closer to its ringing location (.
Figure 3. Trends in recovery distances for a) direct recoveries of Tufted Duck, b) all recoveries of Tufted Duck and c) all recoveries of Goldeneye.
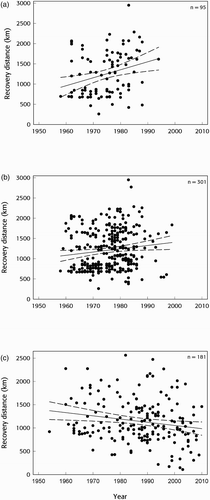
In Finland, the mean ringing location for Goldeneye was 62°7.365'N 25°40.471'E. The mean recovery position for Goldeneye was located in the Baltic Sea just off the Danish island of Møn (54°58.629'N 12°14.265'E) reflecting a clustered distribution of recovery positions and limited recoveries throughout Europe. There was a strong trend for winter temperatures to increase at this location between 1950 and 2010 (F 1,59 = 3.469, P = 0.067; . Moreover, there was an apparent spatiotemporal trend for old recoveries to have been collected from near the southwestern edge of the species' wintering range and more recent recoveries to have been collected near the northeastern edge ( whilst there was a significant negative relationship between recovery distances and time using direct and indirect recoveries (. On average, the recovery distance of Goldeneye decreased by approximately 400 km between 1950 and 2010 (.
None of the species exhibited consistent or significant relationships between their recovery distances and either winter temperatures or the difference in temperatures on the breeding and wintering grounds.
DISCUSSION
The recovery distance for Goldeneye decreased significantly between 1950 and 2010 but, contrary to our hypothesis, the recovery distance for Tufted Duck increased and recovery distances for Pochard remained variable and unchanged. Although there was wide dispersal from a single breeding source for all three species, there were no species-consistent changes in the winter distribution during the second half of the 20th century. Lehikoinen et al Citation(2013), using International Waterbird Census (IWC) data, convincingly demonstrated a northeastern shift, ie closer to the breeding grounds, in the wintering distribution of Tufted Duck and Goldeneye during the last 30 years. On the other hand, Švažas et al Citation(2001), using ringing and recovery data, found no changes in the recovery distance of Pochard migrating from their breeding grounds in Lithuania to their wintering grounds. He postulated this may have been due to his small sample size of only 33 individuals. Here, using a much larger sample (564 individuals), we were similarly unable to find any evidence of migratory short-stopping in Pochard breeding in neighbouring Latvia. Thus, despite evidence for warmer winter conditions throughout Europe and migratory short-stopping of diving duck species using count data, ringing and recovery data provided little utility in capturing and describing movement contraction associated with climatic change.
Recovery distances were independent of mean daily minimum winter temperatures on the wintering grounds and the difference in temperatures between the breeding and wintering grounds. In a comparable study, winter recovery distances for 12 of 24 bird species were shown to have decreased over the past 70 years and recovery distances were negatively correlated with temperature in the winter of recovery (Visser et al Citation2009). This effect was most pronounced for species from dry and open areas and least so for species from wet and open areas (Visser et al Citation2009), such as Pochard and Tufted Duck. Keller et al Citation(2009) showed that Pochard travelled longer distances in harsh winters compared to mild ones, a pattern possibly triggered by the freezing of lakes. We did not find this to be the case, as Pochard did not shorten their recovery distance even though the mean daily minimum winter temperatures at their wintering grounds significantly increased. Tufted Duck, on the other hand, increased their recovery distance while the temperatures at their wintering grounds exhibited only marginal warming. Fiedler et al Citation(2004) demonstrated that, of 30 species of short distance or partial migrants, eight reduced and five increased their migratory distance from their breeding to wintering grounds, while for 17 species migratory distance did not change significantly over time. Whilst a link between recent global climate change and migration distances may be plausible, it has yet to be shown to be causal (Fiedler et al Citation2004).
Other studies have shown that European winters have become milder (Moberg & Jones Citation2005, Lehikoinen et al Citation2013). On a regional scale, the annual and February temperatures in southwestern Sweden have increased significantly since the mid 1970s; this was not the case for January temperatures which remained constant (Nilsson et al Citation2006). Overall, the majority of studies have demonstrated an effect of warmer winter temperatures on changes in the wintering distribution of migratory species (Austin & Rehfisch Citation2005, Maclean et al Citation2008, Visser et al Citation2009, Lehikoinen et al Citation2013). Wildfowl in particular have been shown to respond rapidly to short-term climatic fluctuations (Ridgill & Fox Citation1990, Wernham et al Citation2002, Hofer et al Citation2005, Rainio et al Citation2006). Other factors, however, such as hunting pressure, wetland availability or availability of winter feeding are also likely to play an important role in determining the wintering distribution of wildfowl (see Dalby et al Citation2013).
Birds ringed or recovered in Russia were not included in the current study as data were not available. It is possible that Russian birds and those nesting further east may exhibit a greater propensity for migratory short-stopping behaviour, especially if they perform a leapfrog migration which would render their breeding and wintering grounds further apart. Leapfrog migration has been demonstrated in female Pochard (Hofer et al Citation2006), and it is plausible that it could apply to the other diving duck species.
We advocate caution in the use of ringing and recovery data to assess the role of climate change on migratory short-stopping. Ducks as quarry species have a recovery rate of 5–10% (Guillemain et al Citation2011) which is much higher than those of species that are not hunted; hence ducks should be ideal candidates for this type of analysis. However, it follows that the distribution of ringing recoveries is heavily influenced by hunting pressure throughout most of Europe (Geis & Atwood Citation1961, Hofer et al Citation2006). For example, the hunting pressure on Pochard is higher in western than in central Europe (Švažas et al Citation2001); therefore, we would assume that more of these ducks would be recovered from western Europe, biasing analytical results.
Recovery rates also vary temporally (Baillie & Green Citation1987, Robinson et al Citation2009, Guillemain et al Citation2011). We found a decline in the number of recoveries of Pochard and Tufted Duck, with records sparse after 2000. This trend was not as pronounced in Goldeneye, which was sourced from a different breeding area and was the only species in which a reduction in the recovery distances could be demonstrated. We have no data on the numbers of Goldeneye ringed in Finland, but the numbers of Pochard and Tufted Duck ringed on Lake Engure, where the vast majority of Latvian diving ducks were ringed, decreased steeply towards the end of our study period due to a declining breeding population. During 1980–95, the mean numbers of Pochard and Tufted Duck ringed per year were 1,544 and 1,150 individuals, respectively. However, during 1996–2011, these numbers fell to 54 and 167 individuals respectively (J.A. Viksne, pers comm). Hence, it is clear that the declining number of recoveries from this source was at least partially caused by fewer birds being ringed, because of the large decreases in breeding numbers (J.A. Viksne, pers comm).
The lack of data for ringing recoveries for Pochard and Tufted Duck in the last 15 years will likely present a problem, as any effect of climate change would be expected to be more pronounced in recent years. We detected a warming of winter temperatures in all three sites examined, which was most pronounced in France and to a lesser extent Denmark. The extent of ice cover in the Baltic has generally decreased in the last 20 years (Nilsson Citation2008). This seems linked to increasing numbers of Pochard, Tufted Duck and Goldeneye wintering in Sweden (Nilsson Citation2008). Therefore, we might conclude that the period when a foreshortening in the recovery distance of wintering diving ducks is most likely to occur coincides with the period when both ringing effort and reporting rates of recovered birds have fallen. While changes in the recovery distances for Tufted Duck and Goldeneye were detected, there was no evidence that these changes were temperature dependent and thus causally linked to climate change. It is possible that this is due to the limitations of ringing and recovery data sets and the confounding variables of uneven hunting pressure and declining recovery rates, rather than to a lack of any clear trend. Therefore, it must be accepted that ringing and recovery data are extremely noisy and potentially of limited utility in studies of this kind.
Continued changes in the distribution and abundance of these waterbirds along the flyway will necessitate vigilance to ensure that protected site networks are modified to take account of the responses of quarry duck species to changes in climate and other associated environmental factors. We would encourage more constant efforts to individually mark diving duck species in the immediate future and to improve reporting rates as we enter a period of accelerating environmental change.
ACKNOWLEDGEMENTS
This project was funded by the Natural Heritage Research Partnership (NHRP) between the Northern Ireland Environment Agency and Quercus, Queen's University Belfast in collaboration with the Aarhus University, Denmark under an AGSoS PhD School Visiting Grant to IT. We are grateful to the European Union for Bird Ringing (EURING) which made the recovery data available through the EURING data bank and to the very many ringers and ringing scheme staff who gathered and prepared the data. We are especially grateful to the team at Lake Engure in Latvia for their inspiring long-term ringing and monitoring of bird populations there. Thanks go also to Kendrew Colhoun (RSPB NI) for his support and encouragement.
Related Research Data
REFERENCES
- Austin , G. E. and Rehfisch , M. M. 2005 . Shifting distributions of migratory fauna in relation to climatic change . Global Change Biology , 11 : 31 – 38 . (doi:10.1111/j.1529-8817.2003.00876.x)
- Baillie , S. R. and Green , R. E. 1987 . The importance of variation in recovery rates when estimating survival rates from ringing recoveries . Acta Ornithologica , 23 : 41 – 46 .
- Berry , J. 1941 . International Wildfowl Inquiry: factors affecting the general status of wild geese and wild duck, Volume 1 , Cambridge : Cambridge University Press .
- Burnham , K. P. and Anderson , D. R. 2002 . Model Selection and Multimodel Inference: a practical information-theoretic approach , New York , , USA : Springer-Verlag .
- Carbone , C. and Owen , M. 1995 . Differential migration of the sexes of Pochard Aythya ferina: results from a European survey . Wildfowl , 46 : 99 – 108 .
- Dalby , L. , Fox , A. D. , Petersen , I. K. , Delany , S. and Svenning , J.-K. 2013 . Temperature does not dictate the wintering distributions of European dabbling duck species . Ibis , 155 : 80 – 88 . (doi:10.1111/j.1474-919X.2012.01257.x)
- du Feu , C. R. , Joys , A. C. , Clark , J. A. , Fiedler , W. , Downie , I. S. , van Noordwijk , A. J. , Spina , F. , Wassenaar , R. and Baillie , S. R. 2009 . EURING Data Bank geographical index. , www.euring.org/edb (accessed 23 May 2012)
- EURING . 2010 . EURING Exchange Code 2000+ , Thetford : European Union for Bird Ringing . www.euring.org/edb (accessed 23 May 2012)
- Fiedler , W. , Bairlein , F. and Köppen , U. 2004 . Using large-scale data from ringed birds for the investigation of effects of climate change on migrating birds: pitfalls and prospects . Advances in Ecological Research , 35 : 49 – 67 . (doi:10.1016/S0065-2504(04)35003-8)
- Geis , A. D. and Atwood , E. L. 1961 . Proportion of recovered waterfowl bands reported . Journal of Wildlife Management , 25 : 154 – 159 . (doi:10.2307/3798673)
- Guillemain , M. , Devineau , O. , Gauthier-Clerc , M. , Hearn , R. , King , R. , Simon , G. and Grantham , M. 2011 . Changes in ring recovery rates over the last 50 years: shall we continue to ring ducks? . Journal of Ornithology , 152 : 55 – 61 . (doi:10.1007/s10336-010-0546-4)
- Hofer , J. , Korner-Nievergelt , F. , Korner-Nievergelt , P. , Kestenholz , M. and Jenni , L. 2005 . Herkunft und Zugverhalten von in der Schweiz überwinternden Reiherenten Aythya fuligula: eine Ringfundanalyse . Ornithologische Beobachter , 102 : 181 – 204 .
- Hofer , J. , Korner-Nievergelt , F. , Korner-Nievergelt , P. , Kestenholz , M. and Jenni , L. 2006 . Herkunft und Zugverhalten von in der Schweiz überwinternden oder durchziehenden Tafelenten Aythya ferina . Ornithologische Beobachter , 103 : 65 – 86 .
- Keller , I. , Korner-Nievergelt , F. and Jenni , L. 2009 . Within-winter movements: a common phenomenon in the Common Pochard Aythya ferina . Journal of Ornithology , 150 : 483 – 494 . (doi:10.1007/s10336-008-0367-x)
- La Sorte , F. A. and Thompson , F. R. III . 2007 . Poleward shifts in winter ranges of North American birds . Ecology , 88 : 1803 – 1812 . (doi:10.1890/06-1072.1)
- Lehikoinen , A. , Jaatinen , K. , Vähätalo , A. V. , Clausen , P. , Crowe , O. , Deceuninck , B. , Hearn , R. , Holt , C. A. , Hornman , M. , Keller , V. , Nilsson , L. , Langendoen , T. , Tománková , I. , Wahl , J. and Fox , A. D. 2013 . Rapid climate driven shifts in wintering distributions of three common waterbird species . Global Change Biology , 19, 2071–2081
- Maclean , I. M.D. , Austin , G. E. , Rehfisch , M. M. , Blew , J. , Crowe , O. , Delany , S. , Devos , K. , Deceuninck , B. , Günther , K. , Laursen , K. , Van Roomen , M. and Wahl , J. 2008 . Climate change causes rapid changes in the distribution and site abundance of birds in winter . Global Change Biology , 14 : 2489 – 2500 .
- McAlpine , C. A. , Rhodes , J. R. , Callaghan , J. G. , Bowen , M. E. , Lunney , D. , Mitchell , D. L. , Pullar , D. V. and Possingham , H. P. 2006 . The importance of forest area and configuration relative to local habitat factors for conserving forest mammals: a case study of koalas in Queensland, Australia . Biological Conservation , 132 : 153 – 165 . (doi:10.1016/j.biocon.2006.03.021)
- Moberg , A. and Jones , P. D. 2005 . Trends in indices for extremes in daily temperature and precipitation in central and western Europe, 1901–99 . International Journal of Climatology , 25 : 1149 – 1171 . (doi:10.1002/joc.1163)
- Nichols , J. D. and Haramis , G. M. 1980 . Sex-specific differences in winter distribution patterns of Canvasbacks . Condor , 82 : 406 – 416 . (doi:10.2307/1367565)
- Nilsson , A. L.K. , Lindström , Å. , Jonzén , N. , Nilsson , S. G. and Karlsson , L. 2006 . The effect of climate change on partial migration – the blue tit paradox . Global Change Biology , 12 : 2014 – 2022 . (doi:10.1111/j.1365-2486.2006.01237.x)
- Nilsson , L. 2008 . Changes in numbers and distribution of wintering waterfowl in Sweden during forty years, 1967–2006 . Ornis Svecica , 18 : 135 – 226 .
- Quinn , G. P and Keough , M. J. 2002 . Experimental Design and Data Analysis for Biologists , Cambridge : Cambridge University Press .
- Rainio , K. , Laaksonen , T. , Ahola , M. , Vähätalo , A. V. and Lehikoinen , E. 2006 . Climatic responses in spring migration of boreal and arctic birds in relation to wintering area and taxonomy . Journal of Avian Biology , 37 : 507 – 515 . (doi:10.1111/j.0908-8857.2006.03740.x)
- Ridgill , S. C. and Fox , A. D. 1990 . Cold weather movements of waterfowl in western Europe. Special Publication 13 , Slimbridge : IWRB .
- Robinson , R. A. , Grantham , M. J. and Clark , J. A. 2009 . Declining rates of ring recovery in British birds . Ringing & Migration , 24 : 266 – 272 . (doi:10.1080/03078698.2009.9674401)
- Stroud , D. A. , Mudge , G. P. and Pienkowski , M. W. 1990 . Protecting Internationally Important Bird Sites: a review of the EEC Special Protection Area network in Great Britain , Peterborough : Nature Conservancy Council .
- Švažas , S. , Patapavičius , R. and Dagys , M. 2001 . Recent changes in distribution of wintering populations of waterfowl established on the basis of Lithuanian ringing recoveries . Acta Zoologica Lituanica , 11 : 235 – 242 . (doi:10.1080/13921657.2001.10512455)
- Visser , M. E. , Perdeck , A. C. , van Balen , J. H. and Both , C. 2009 . Climate change leads to decreasing bird migration distances . Global Change Biology , 15 : 1859 – 1865 . (doi:10.1111/j.1365-2486.2009.01865.x)
- Wernham , C. V. , Toms , M. P. , Marchant , J. H. , Clark , J. A. , Siriwardena , G. M. and Baillie , S. R. 2002 . The Migration Atlas: movements of the birds of Britain and Ireland , London : T. & A.D. Poyser .