Abstract
The unstreaked Acrocephalus warblers can be challenging to identify, even in the hand. Previous studies have highlighted wing length, second-primary notch length, and second-primary notch position as criteria for separating Eurasian Reed Warbler A. scirpaceus and Marsh Warbler A. palustris. We present data for wing length, second-primary notch length, and second-primary notch position for Blyth's Reed Warbler A. dumetorum and Paddyfield Warbler A. agricola, obtained from museum skins, to complement data published previously for Scandinavian populations of Eurasian Reed Warbler and Marsh Warbler. These data provide a reliable and non-subjective method for the separation of Blyth's Reed Warbler and Paddyfield Warbler from Eurasian Reed Warbler and Marsh Warbler in Scandinavia.
Blyth's Reed Warbler Acrocephalus dumetorum, Eurasian Reed Warbler A. scirpaceus and Marsh Warbler A. palustris can prove difficult to identify, even in the hand. The plumages of these unstreaked Acrocephalus warblers are similar and there is overlap in structural size measurements. Walinder et al Citation(1988) showed that individuals of Reed and Marsh Warblers caught in southern Scandinavia could be separated reliably using two sets of combined measurements: wing length and notch length on the second outer primary (P2, numbered ascendently), and wing length and P2 notch position in relation to other flight feathers.
In the hand, the identification of Blyth's Reed Warbler, a rare vagrant to southern Scandinavia, relies on a combination of plumage and bare-part coloration, measurements and wing formula (Svensson Citation1992, Cramp Citation1992, Kennerley & Pearson Citation2010). The authors have been involved in discussions surrounding five birds ringed in Sweden since 2007, including both spring and autumn birds, identified using current literature as Blyth's Reed Warbler. All proved to be Eurasian Reed Warbler, including a bird subsequently identified using genetic markers (www.artportalen.se/birds/gallery_imageinfo.asp?imageid=94875); this highlights the potential for error when using the current criteria.
Plumage and bare-part coloration is subjective and can be hard to assess on a lone bird, a bird in, for example, early morning or late evening lighting conditions when orange tones become more prominent, or when observers have little previous experience of the relevant species. Plumage patterns are also hard to quantify, while behavioural and vocal differences between the species are not relevant for identifying a bird in the hand. Svensson Citation(1992) suggests initially checking wing formula, length of wing and bill length. However, there is overlap in wing length and bill length between Eurasian Reed Warbler and Blyth's Reed Warbler, and all misidentified birds in the Swedish examples had wing lengths within the range given by Svensson Citation(1992) for Blyth's Reed Warbler. In addition, we have found bill length one of the hardest measurements to reproduce accurately on a live bird.
Some Eurasian Reed Warblers can show a (weak) emargination, perhaps better referred to as a tapering of the outer web rather than a true emargination, on P4 (Pearson et al Citation2002), inviting confusion with the emarginated P4 typically shown by Blyth's Reed Warbler, while a mist-net thread wrapped around the primaries during trapping can cause an indentation on the outer web of a primary similar in appearance to an emargination (pers obs). Furthermore, Blyth's Reed Warbler may lack an emargination on P4, and be similar to Eurasian Reed Warbler: in a recent report, 11% of 44 juvenile birds lacked emargination on P4 (and P5), although all of 69 adult birds had emargination on P4 (with 7% showing an emargination on P5; Malmhagen Citation2013). Thus, while emarginations are valuable for identification, at times they may be misleading.
Van Swelm & de Kniff Citation(1984) concluded that the position of the notch on the second primary is “[one of the two] most reliable characters” for identifying Blyth's Reed Warbler but few data were presented. Svensson Citation(1992) presents P2 notch position as a measure of distance from secondary tips, a measurement that is hard to take on live birds in the field.
To extend the data presented by Walinder et al Citation(1988) for Eurasian Reed Warbler and Marsh Warbler, three measurements were taken by BM from skins of Blyth's Reed Warbler and from the related Paddyfield Warbler A. agricola; P2 notch length, P2 notch position, and wing length. Paddyfield Warbler is the most distinctly plumaged of the four species compared here (Kennerley & Pearson Citation2010) but aspects of wing morphology are still likely to prove helpful in identifying a vagrant bird, and these measurements appear to be useful in separating Paddyfield Warblers from agricola-like Acrocephalus hybrids (cf van Eerde Citation1999; Malmhagen & Menzie pers. obs.). The skins used for this study were at the British Natural History Museum, Tring, Universitetets Zoologisk Museum, Copenhagen, and Naturhistoriska riksmuseet, Stockholm, and had been collected across a relatively broad area (). Measurements taken by BM and Göran Walinder on museum skins have been calibrated on live birds whilst working together at Falsterbo Bird Observatory and data (including those taken by Walinder in Walinder et al Citation1988) should be considered comparable in this respect.
Table 1. Sample sizes of Blyth's Reed Warbler and Paddyfield Warbler museum skins measured, by country of origin.
Wing length was taken as maximum chord (method 3 in Svensson Citation1992) and measured to the nearest 1 mm; P2 notch length refers to the length of the notch on the inner web of the second primary, measured to the nearest 0.5 mm from the point of inflection to the feather tip (; Svensson Citation1992; Deutsche Ornithologen-Gesellschaft Citation2011). Notch position was taken on the closed wing and was scored according to the remex whose tip the inflection of the notch fell closest to; where the level of the notch fell between the tip of two remiges, it was scored as x/y (falling between feather x and feather y; see Svensson Citation1992).
Figure 1. P2 notch length (left-hand image; line A) should be measured from the tip of the feather to the point of inflection (right-hand image; B).

The preparation and drying of museum skins can cause wing length to decrease due to removal of muscle tissue and skin shrinkage. The degree of shrinking varies between individual skins (Winker Citation1993) but must be taken into account when comparing live birds (in this study, Reed Warber and Marsh Warbler from Walinder et al Citation1988) with museum specimens. Winker Citation(1993) showed that the average wing length of Alder/Willow Flycatcher Empidonax alnorum/traillii and Tennessee Warbler Oreothlypis peregrina, two species with similar wing lengths to the Acrocephalus warblers studied in this paper, decreased by 0.57% and 1.44% respectively, representing significant changes in wing length. Studies on other taxa have shown that shrinkage can be an average 4% of the live or freshly dead wing length (eg in grebes, Fjeldså Citation1980). Winker Citation(1993) suggests a crude general correction factor of 0.983, based on an average shrinkage of 1.7%, to be used when comparing taxa for which skin shrinkage has not been specifically studied. However, without specific study on the skin shrinkage of Acrocephalus warblers, we present our data unadjusted. Nevertheless, individual feather length is virtually unaffected by skin shrinkage (Jenni & Winkler Citation1989) and the P2 notch length on museum skins should be comparable with that on live birds.
We treat the birds of each species as two data sets: juvenile and adult. Birds were aged according to criteria in Svensson Citation(1992). Blyth's Reed Warbler undergoes a complete post-breeding/post-juvenile moult at stopover sites in northwest India during their southbound autumn migration (early October to early November). Paddyfield Warbler undergoes a complete post-breeding/post-juvenile moult upon arrival on the wintering grounds; active moult generally occurs during September and October, and is completed by November. After completion of full moult, birds are classified as “adult”.
Juveniles of all four species show on average shorter wing length and shorter notch length than adults (Walinder et al Citation1988, Svensson Citation1992). Data for the two age classes should therefore be considered separately and it is of critical importance that an individual bird is aged before the methods presented here are used.
Paddyfield Warbler is considered polytypic with two races currently recognised. However, Kennerley & Pearson Citation(2010) state that differences are slight and consider there to be no consistent differences in plumage coloration or measurements between the two races. Therefore, we make no effort to differentiate between the two recognised races. Blyth's Reed Warbler is considered monotypic (Kennerley & Pearson Citation2010), though we acknowledge that this does not imply uniform distribution of measurements across the species' range.
Data for Blyth's Reed Warbler and Paddyfield Warbler are shown in & , separately for juvenile and adult birds, together with data from Walinder et al Citation(1988) for Eurasian Reed Warbler and Marsh Warbler to allow direct comparison. Analysis of the data was carried out by MK using R (R Core Team Citation2012) and mean, sample size, range and standard deviation are summarised in . To calculate 95% confidence regions, shown as coloured cells in & , the R-function ‘data.Ellipse’ was used. The assumption that data were normally distributed along the x- and y-axes was verified by frequency histogram plots of each variable.
Figure 2. ‘Wing/notch’ method for adults (a) and juveniles (b). Sample sizes for the relationship between notch length (P2) (mm) and wing length (mm) in adult Paddyfield Warbler (red numbers), Blyth's Reed Warbler (blue), Eurasian Reed Warbler (orange) and Marsh Warbler (green) with coloured areas representing 95% confidence ellipses. Hatched areas represent areas of overlap between species.
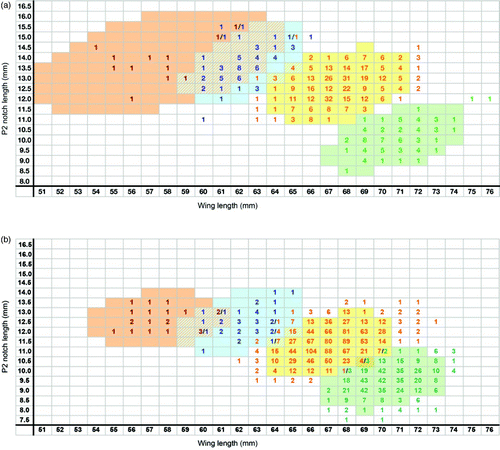
Figure 3. ‘Notch position’ method for adults (a) and juveniles (b). Sample sizes for the relationship between notch length (P2) (mm) and wing length (mm) in adult Paddyfield Warbler (red numbers), Blyth's Reed Warbler (blue), Eurasian Reed Warbler (orange) and Marsh Warbler (green) with coloured areas representing 95% confidence ellipses. Hatched areas represent areas of overlap between species.
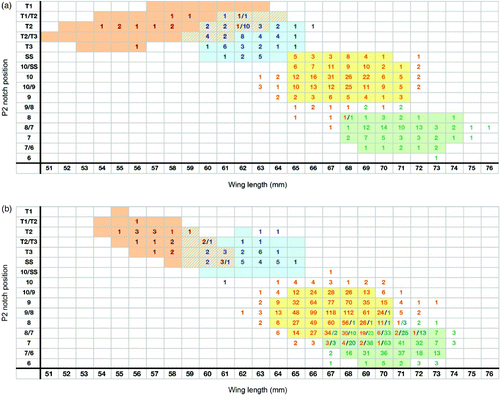
Table 2. Mean values and ranges of wing length, P2 notch length and notch position, with sample size (n) and standard deviation (sd). All measurements in mm. P = primary, numbered ascendently (distal to proximal); T = tertial, numbered descendently (proximal to distal); SS = secondaries.
As the origins of vagrant Blyth's Reed Warblers and Paddyfield Warblers in Europe are unknown, we made no effort to separate the data based on origin of specimens. The data for Eurasian Reed Warbler and Marsh Warbler (Walinder et al Citation1988) were collected during autumn migration in southern Sweden and these birds are likely to be of Scandinavian origin. Wilson et al Citation(2001) found that the measurements of Walinder et al Citation(1988) were unsatisfactory for determining the identification of Eurasian Reed Warblers (nominate and fuscus races) and Marsh Warblers passing through Lesvos, Greece. Therefore, for these two species, it may be unwise to use the measurements presented here away from Scandinavia.
When using these methods, care must be taken to avoid applying criteria to birds in active moult (adults and juveniles in winter) or recently fledged birds still growing primaries. Wing length is at its maximum immediately following completion of growth before the effects of wear have abraded the feather (Francis & Wood Citation1989) and, for the species treated here, wing length should be greatest in early spring and least in late autumn. Using Marsh Warbler data, we compared data for wing length and for notch length collected at Falsterbo Bird Observatory during spring (13 May to 10 June) to that collected during autumn (21 July to 25 August). Although mean spring wing length was 0.21 mm longer than mean autumn wing length, this difference was not statistically significant (P > 0.09) and there was no significant difference in mean notch length between spring and autumn (P > 0.56). Therefore, feather wear may not be an important factor to consider when applying the criteria presented here.
In all measurements there is a risk of error caused by slight variation in measuring techniques between people. We strongly advise that the methods described here and in Walinder et al Citation(1988) and Svensson Citation(1992) are followed as accurately as possible. Our view is that the three measurements of wing length, P2 notch length and P2 position used here are less prone to error than alternative methods for the identification of Blyth's Reed Warbler from Eurasian Reed Warbler and Marsh Warbler.
We recommend that the criteria presented here should be regarded as additional tools for the identification of unstreaked Acrocephalus warblers in the hand. For birds where measurements fall into the overlap zones (hatched areas in & ), traditional identification methods such as plumage and bare-part coloration or full wing formula will need to be used. Hybrids are not unknown (eg Marsh Warbler x Blyth's Reed Warbler: see Lindholm et al Citation2007, Pearson et al Citation2012) and some birds may need to be left unidentified or identified using genetic material. We acknowledge that these methods can be improved with the addition of data from live Blyth's Reed Warblers and Paddyfield Warblers.
ACKNOWLEDGEMENTS
We would like to thank several people who made this article possible. First and most of all, these are the long-term permanent staff at Falsterbo Bird Observatory who wrote the article in 1988, Göran Walinder, Lennart Karlsson and Karin Persson. Especially we want to thank Göran Walinder for rewarding discussions and Lennart Karlsson for valuable comments on the manuscript. We also want to thank Mark Adams (Natural History Museum, Tring), Jan Bolding Kristensen (Zoological Museum in Copenhagen) and Thord Fransson (Swedish Museum of Natural History in Stockholm) for giving us access to the skin collection in each museum. This is report number 277 from Falsterbo Bird Observatory.
REFERENCES
- Cramp , S. 1992 . The birds of the Western Palearctic. Volume 6, Warblers , Edited by: Cramp , S. Oxford : Oxford University Press .
- Deutsche Ornithologen-Gesellschaft . 2011 . Measuring Birds/Vögel Vermsessen , Wilhelmshaven : Deutsche Ornithologen-Gesellschaft .
- van Eerde , K. A. 1999 . Hybride Rietzanger x Kleine Karekiet te Makkum in augustus 1997 . Dutch Birding , 21 : 34 – 37 .
- Fjeldså , J. 1980 . Post-mortem changes in measurements of grebes . Bulletin of the British Ornitholgists' Club , 100 : 151 – 154 .
- Francis , C. M. and Wood , D. S. 1989 . Effects of age and wear on the wing length of wood-warblers . Journal of Field Ornithology , 60 : 495 – 503 .
- Jenni , L. and Winkler , R. 1989 . The feather-length of small passerines: a measurement for wing-length in live birds and museum skins . Bird Study , 36 : 1 – 15 . (doi:10.1080/00063658909476996)
- Kennerley , P. and Pearson , D. 2010 . Reed and Bush Warblers , London : Christopher Helm .
- Lindholm , A. , Bensch , S. , Dowsett-Lemaire , F. , Forsten , A. and Kärkkäinen , H. 2007 . Hybrid Marsh x Blyth's Reed Warbler with mixed song in Finland in June 2003 . Dutch Birding , 29 : 223 – 231 .
- Malmhagen , B. 2013 . Artbestämning av tre ostreckade Acrocephalus . Roadrunner , 2013 : 26 – 32 .
- Pearson , D. J. , Small , B. J. and Kennerley , P. R. 2002 . Eurasian Reed Warbler: the characters and variation associated with the Asian form fuscus . British Birds , 95 : 42 – 61 .
- Pearson , D. J. , Kerton , I. , Amakobe , B. and Bensch , S. 2012 . Presumed hybrid Blyth's Reed Warbler Acrocephalus dumetorum x Marsh Warbler A. palustris trapped at Ngulia, Kenya, in December 2009 . Bulletin of the British Ornithologists' Club , 132 : 124 – 127 .
- R Core Team . 2012 . R: a language and environment for statistical computing , Vienna , , Austria : R Foundation for Statistical Computing .
- Svensson , L. 1992 . Identification Guide to European Passerines. Fourth edition , Stockholm : Lars Svensson .
- van Swelm , N. and de Kniff , P. 1984 . A new method for identifying A. palustris, A. scirpaceus and A. dumetorum in hand . Ostvorne Ringing Report , 1983 : 23 – 26 .
- Walinder , G. , Karlsson , L. and Persson , K. 1988 . A new method for separating Marsh Warbler (Acrocephalus palustris) from Reed Warbler (Acrocephalus scirpaceus) . Ringing & Migration , 9 : 55 – 62 . (doi:10.1080/03078698.1988.9673924)
- Wilson , J. D. , Akriotis , T. , Balmer , D. E. and Kyrkos , A. 2001 . Identification of Marsh Warblers Acrocephalus palustris and Reed Warblers A. scirpaceus on autumn migration through the eastern Mediterranean . Ringing & Migration , 20 : 224 – 232 . (doi:10.1080/03078698.2001.9674250)
- Winker , K. 1993 . Specimen shrinkage in Tennessee Warblers and “Traill's” Flycatchers . Journal of Field Ornithology , 64 : 331 – 336 .