
ABSTRACT
The European Union for Bird Ringing (EURING) coordinates bird ringing at a continental scale and operates the EURING Data Bank (EDB) to facilitate large-scale analyses of movements and demography. The EDB contains over 10 million individual encounter records which are summarised on a publicly available website, the EDB index. EURING welcomes applications to analyse these data. Ring-recovery data from the EDB contribute to research on many ecological issues, particularly migration and movements, hunting, mortality causes, disease transmission, population dynamics and dispersal. Recent developments will facilitate the incorporation of comprehensive sets of first-encounter records and local recaptures which are essential for robust quantitative studies of population dynamics and movements. Furthermore, the recent inclusion of fields for moult, measurements and weights within the EURING code will facilitate novel research at a European scale. We expect increasing use of the EDB for quantitative studies of avian demography and movements with high applied value. Wherever possible, this research should also incorporate complementary ecological information. EURING’s immediate priority is the production of a European Migration Atlas that would provide an up-to-date synthesis of the movements of European bird populations, with many direct implications for their conservation.
Studies of marked individuals form an essential part of pure and applied research in avian biology, particularly with respect to migration, movements and population dynamics. Furthermore, there is an increasing realisation that large-scale analyses of ecological data, supported by online data gathering, integrated database technologies and novel statistical and computational approaches can offer powerful new insights into the ecology of birds and other organisms. Thus the application of concepts such as big data (Schimel Citation2011, Hampton et al Citation2013), hierarchical modelling (Kéry & Schaub Citation2012), integrated population modelling (Robinson et al Citation2014) and machine-learning techniques (Fink et al Citation2010) offer many exciting opportunities for studies of marked birds to advance our knowledge of avian ecology over the coming decade. Linked to the development of these large-scale analytical approaches there is an increasing move towards the provision of open data that will allow analysts to combine data from a wide range of different sources quickly and easily (Reichman et al Citation2011, Michener & Jones Citation2012).
It is evident that many of the processes that determine the distributions, abundances and movement patterns of birds operate across political boundaries, and therefore studies covering groups of countries, continents or flyways are needed. From its inception, bird ringing has always addressed movements across political boundaries. The resulting need for a common language to support standardised data gathering, quality control and data exchange necessitated close cooperation between institutions that use bird ringing as a tool for avian science. Thus the European Union for Bird Ringing (EURING) was founded in 1963. Today, almost all European countries are represented in the EURING community through some 40 ringing centres, usually one per country. EURING provides guidelines and standards for bird ringing, maintains a standard code for the computerisation and exchange of mark–reencounter data and operates a multi-language website (www.ring.ac) for the reporting of ring recoveries. It facilitates communication between schemes, ringers and members of the public through the internet and other media, undertakes analyses of mark–reencounter data at a European scale and coordinates a network of over 500 Constant Effort Sites throughout Europe (Eglington et al Citation2015, Johnston et al Citation2016). It also promotes the development of statistical and computing methods for the analysis of mark–reencounter data, particularly through the influential series of EURING analytical meetings that bring together leading statisticians and ecologists working on this topic (Francis et al Citation2014). Last, but certainly not least, it runs the EURING Data Bank (EDB). This unique set of mark–reencounter data on European birds recently reached the milestone of 10 million encounter records and is at the disposal of the scientific and conservation communities.
The EURING Data Bank
Terminology used for mark–reencounter data
Here, first-encounter records refer to the initial encounter with an individual when it was first ringed. Recoveries comprise all dead (including sick and dying) encounters, many of which are from members of the public, live recaptures away from the ringing place (nearly all by qualified ringers), and live resightings away from the ringing place (mainly by ringers and other birdwatchers). Live recoveries include both recaptures and resightings away from the ringing place, normally with a threshold of 5 km, although there is some variation with respect to particular species and ringing centres (). Depending on context, the term ‘recoveries’ may refer to both a finding record and the original ringing record. Local recaptures and local resightings are those below the relevant distance threshold. All record types comprising ringing records, dead recoveries, recaptures and resightings are referred to as mark–reencounter data. An encounter history comprises all encounters of a particular individual. Following the normal convention, reencounter data based on live recaptures and resightings only are referred to as capture–recapture data.
Table 1. EURING Data Bank holdings by ringing scheme and 20-year period. Numbers of recovery records held by the EDB in June 2015. This includes all birds recovered dead or injured together with recaptures and resightings more than 10 km from the original ringing location. Initial ringing records and live recaptures close to the ringing location are excluded. These figures should be interpreted in relation to differing data management and submission practices between schemes (see text). Additional data may exist in the files of individual schemes or individual ringers, but may not have been submitted or computerised. In addition to the records summarised here the EDB holds some local recaptures for schemes marked a and some first-encounter data for birds with no reencounters for schemes marked b. Where appropriate, data from defunct schemes are included with those from the scheme currently covering the relevant country.
Historical development of ringing in Europe
Scientific bird ringing began in Denmark in 1889 when H.C.C. Mortensen attached metal rings to Starlings Sturnus vulgaris in order to study their migration. Each ring carried a return address and a unique number – vital items required for identifying the individual and reporting the information about subsequent encounters. There are now some 40 formally constituted ringing schemes in Europe, several having achieved their first century. Studies of migration necessarily involve communication between ringers in one country and finders in another. In 1966, only three years after it was founded, EURING published a Code Manual which, for the first time, defined a coding scheme for recording information about ring-reencounter histories (Anon Citation1966). Technology limited the information held for each recovery (both ringing and finding information) to one 80-column punch card. At that time, most schemes did not have access to computers and the future power of information technology could not be foreseen. It is thus to the great credit of the code’s authors that they provided such a firm foundation for subsequent developments. The code was revised in 1979, but still with the 80-character limitation (Mead et al Citation1979). Computerisation of current and historical recovery information was enhanced in 1982 by grants from the European Community coordinated through EURING. Thanks to this and other inputs, we now have access to ring recoveries dating back half a century before the development of digital computers.
In 1977, with support from the Netherlands Institute of Ecology, the EURING Data Bank (EDB) was founded. It aspired to hold all ring-recovery data for birds ringed or recovered in Europe so as to make them available for research. Initially, technology limited the scope of the Data Bank, with data transfers relying on low-capacity floppy disks sent through the post and very limited processing power, making data manipulation and analysis difficult.
In 2000, a major revision of the code was published, with the character constraint being increased from 80 to 255 per record (Speek et al Citation2001). At the same time, the ringing and reencounter information were split into separate records for each encounter, giving vastly increased capability to record relevant information and avoiding the duplication of first-encounter data. This new format is better suited to recording comprehensive sets of encounter histories, including the many records of marked individuals that are not subsequently reencountered. Space was reserved for data on measurements and moult – recognising that ringing data could now be used to address many research questions in addition to the description of migration routes and basic analyses of demographic rates.
The EDB was reorganised in 2005 and moved to the British Trust for Ornithology (BTO) in the UK. All records are now stored in an ORACLE database which facilitates rapid and effective data access, presentation and analysis. The EURING web site became the gateway to the EDB for researchers in search of data and also made the EDB more prominent and accessible.
In 2010 an extension of the 2000 exchange code was released, called the 2000+ code (du Feu et al Citation2010). This defined, for the first time, morphometric and moult variables and contained all the fields required for data exchange between schemes during recovery processing. Codes for some fields (Scheme, Place, Species, Finding Circumstances) are now available from interactive tables on the EURING website (Appendix), rather than being incorporated in the text of the code manual. The release of the 2000+ code realised the far-sighted vision of the authors of the 1966 code – and much more besides.
Current content
A full description of the contents of the EDB is provided by the EDB Index (du Feu et al Citation2016; Appendix) which is available online and is updated as new data are received. This includes summaries of numbers of dead and live recoveries by species and time period. It also provides maps showing the distributions of ringing and finding places of birds recovered dead and alive for each species (). This information allows potential analysts to assess whether the data are likely to be suitable to address a specific research question. The figures presented in the online EDB index need careful interpretation, due to variation in the submission practices of different schemes, largely driven by differences in data-management practices and the extent of computerisation. Some 14 schemes submit data for all reencounters of ringed birds, at least for more recent years, while two have recently started to include ringing records for birds with no subsequent reencounters (). Most schemes submit data for all ringed birds found dead and those live recoveries beyond a certain distance from the point of ringing (see www.euring.org/data-and-codes/reportable-recovery-definitions for details). Although most records are relatively recent, a few are over 100 years old. For some species, there are sufficient older records to enable analyses of long-term temporal changes in recovery and movement patterns (Fiedler Citation2001, Ambrosini et al Citation2011, Calladine et al Citation2012).
Figure 1. Distribution of ringing and recovery locations for Ospreys Pandion haliaetus ringed by European Ringing Schemes and recovered either dead or alive: (a) ringing locations of birds recovered dead; (b) ringing locations of birds recovered alive; (c) finding locations of birds recovered dead; (d) finding locations of birds recovered alive. Redrawn from du Feu et al (Citation2016).
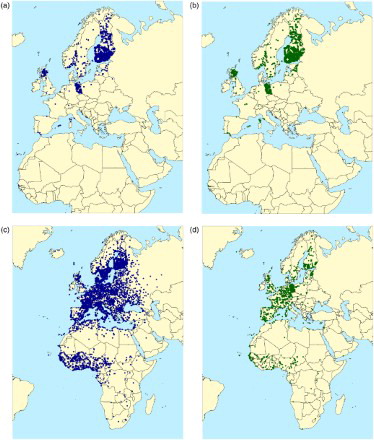
Here we provide a brief summary of the information available. Our latest analysis (June 2015) indicated that the EDB contains over ten million encounter records contributed by 37 ringing schemes. If we consider conventional recovery records, as defined above, the Data Bank contained 3 968 522 recoveries (). Eight schemes (Brussels, Hiddensee, Wilhelmshaven, Thetford, Arnhem, Gdansk, Helsinki and Stockholm) each contributed over 4% of the total and between them these schemes supplied 81.3% of all recovery records. Only 0.1% of these records are from before 1920, with 94.7% being since 1960 and 42.2% since the turn of the century ().
Eight species groups (ducks, raptors, gulls, owls, titmice, hirundines, Acrocephalus warblers, thrushes and finches) each contributed over 4% of individual birds recorded and between them these groups comprise over 55.7% of such records in the EDB (). The representation of different species groups within the EDB is due to a range of factors including species abundances and distributions, ease of capture, recovery probabilities, research priorities and the distribution and interests of volunteer ringers and professional ecologists. A high proportion of bird ringing is dependent on the efforts of highly trained volunteers, and thus tends to focus on small birds that can be captured in at least reasonable numbers using mist nets. This high representation of small passerines will become even more pronounced as more first-capture data and local recaptures are added to the EDB. In contrast, larger species such as ducks and gulls are ringed in much smaller numbers but have relatively high reencounter rates. For analyses of movements and survival that depend on reencounter records, the information content of the records for these larger species may be relatively high while, as information on measurements and moult is increasingly incorporated, the information on small birds, with large numbers of first captures and local recaptures, will become more prominent.
Table 2. EURING Data Bank holdings by species group. Number of records of individual birds held within the EDB in June 2015. Here one individual counts only once, regardless of the number of reencounters. The figures include local recaptures, but not individuals that were captured only once.
Data-provision services
Potential analysts are advised to use the initial enquiry form on the EURING website (www.euring.org/data-and-codes/edb-application-procedure) in order to establish the number of records available, to refine the request and to agree the charge. EURING endeavours to keep these charges as low as possible, but they are necessary to cover the basic running costs of the EDB, which receives no external funding. After the initial enquiry has been completed, a formal request can be made using the relevant form available from the same web page. This is sent, automatically, to all schemes who are thereby invited to agree to the release their data. It most cases this procedure is straightforward and once agreements have been obtained and the charge paid, data are supplied promptly. The process from submitting a formal request to receipt of the data can normally be completed in under a month. Data are supplied in a standard format defined in the EURING exchange code (du Feu et al Citation2010). Requests relating to particular species, areas, times or other selection criteria can be accommodated readily.
Future plans
The EDB was created to promote research into bird movements, demography and conservation using ring-recovery data on a European or flyway scale. The scope of possible research is being enhanced through the inclusion of records of birds which have been ringed but never reencountered, of local recaptures and of data on morphometrics and moult. The total number of records in the latter category is small, as yet, but is expected to increase rapidly in the near future. This will enable a wider range of studies into avian life histories, population processes, morphometrics and moult than has been possible to date. Some of the results together with ideas for novel future research are outlined in subsequent sections of this review.
Research based on large-scale studies of marked birds
Describing migration routes
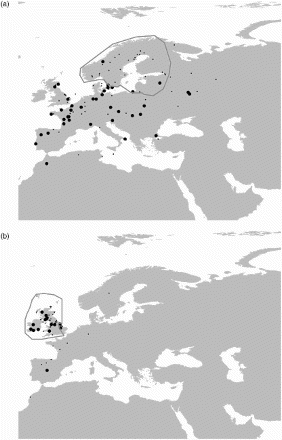
The traditional and most widely understood aim of bird ringing is the description of migration routes. Understanding the spatiotemporal structure of bird movements is a prerequisite for species conservation as well as for understanding the drivers and consequences of migration. Ring-recovery atlases provide overviews of bird movements in space and time as shown by ring recoveries, sometimes also supplemented by other material such as counts or tracking data. Some 13 comprehensive national ring-recovery atlases have been published for European countries, ten in the last decade. Recent additions to the list include Croatia, Faroes, Finland, Hungary, Italy and Germany (Csörgő et al Citation2009, Spina & Volponi Citation2009, Kralj et al Citation2013, Bairlein et al Citation2014, Valkama et al Citation2014). Building on this basic knowledge of bird movements derived from ring recoveries, recent developments of loggers and transmitters facilitate the collection of information about individuals at much higher spatial and temporal resolutions (Fiedler Citation2009, Bridge et al Citation2011). Numerous studies have used ring-recovery data to describe patterns of connectivity between areas used by particular bird species, and also to document different migration routes of subpopulations, age or sex classes (eg Mason & Allsop Citation2009, Reichlin et al Citation2009, Liker & Nagy Citation2009, Kylin et al Citation2011, Ambrosini et al Citation2014). For example, Calladine et al (Citation2012) used records from the EDB to describe the passage and wintering areas of different breeding populations of Short-eared Owls Asio flammeus (). There is also increasing evidence of changes in migration routes over time, as Ambrosini et al (Citation2011) showed for Barn Swallow Hirundo rustica and Calladine et al (Citation2012) for Short-eared Owl.
Figure 2. Recovery locations of Short-eared Owls Asio flammeus with natal/breeding areas in (a) the Boreal region and (b) the Insular Atlantic region. Regions are outlined; large dots show recoveries in winter, small dots in passage periods. Redrawn from Calladine et al (Citation2012).
Mortality causes
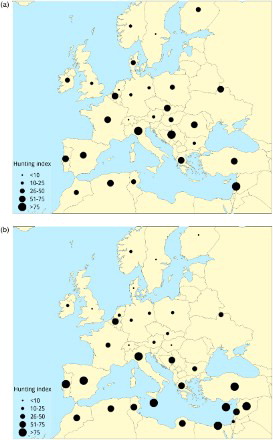
Dead recoveries are stored with the information about the reported cause of death whenever this information is provided by the finder. This typically provides information on conspicuous causes of death related to human activities such as hunting, oil pollution and mortality in fishing nets. Natural causes such as starvation or disease can only rarely be assessed adequately by finders, for example as part of specific studies. Although the finder’s assessment of the reason that a bird died may not always be correct, data on reported causes of death allow some useful insights into mortality patterns (Hüppop & Hüppop Citation2002). Such analyses have been used to document geographical and temporal variation in the hunting of migratory birds across Europe (McCulloch et al Citation1992; ) and to relate variation in the demographic rates of seabirds to differences in exposure to oiling (Votier et al Citation2008).
Figure 3. Geographical variation in hunting indices (a measure of the relative proportion of recoveries attributable to hunting) for (a) raptors and (b) passerines, in the pre-1980 period. Redrawn from McCulloch et al (Citation1992).
A major problem is that the probability of finding a dead individual depends on the mortality cause, and therefore the frequency of mortality causes related to humans (eg hunting, traffic accidents, electrocution) is usually overestimated within the raw data. Schaub & Pradel (Citation2004) developed a probabilistic model which provides unbiased estimates of mortality proportions based on recoveries of marked individuals. They applied their model to recovery data for Swiss White Storks Ciconia ciconia and found that about 35% of individuals died due to the collisions with power lines. Once cause-specific mortality probabilities and their temporal variations are known, hypotheses about whether a specific mortality cause is additive to the remaining (natural) mortality, or is (partially) compensated by lowering of other mortality causes through density-dependent population processes, can be tested (Schaub & Lebreton Citation2004, Servanty et al Citation2010). Such an assessment is often important for the management of populations, in particular for setting hunting bags. The degree of compensation of hunting mortality has been a key research focus for the regulation of the hunting of Mallard Anas platyrhynchos and other duck species in the USA (Burnham & Anderson Citation1984, Nichols et al Citation2007) and there is an urgent need to apply such approaches within a European context (Elmberg et al Citation2006).
Disease transmission
Birds are vectors for a multitude of pathogens, ranging from those that are mainly interesting for understanding host–pathogen interactions or epidemiological mechanisms (Hochachka & Dhondt Citation2006) to those of direct significance for the health of humans or livestock (Globig et al Citation2009a). Since few other vertebrate hosts are as mobile as migrating birds, the latter may play an important role in long-distance transmission of some diseases. While case studies can be undertaken using tracking and geo-logging devices to follow the migration routes of individual birds, at present such measures cannot fully replace the long-term ring-recovery data sets covering large areas and many species (Fiedler Citation2009).
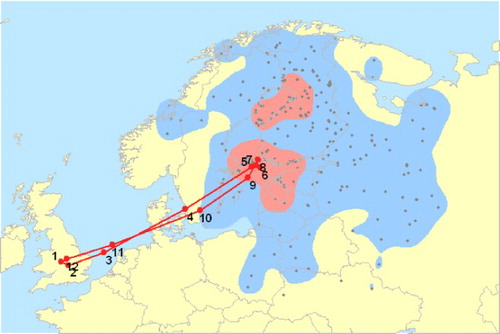
In the context of highly pathogenic avian influenza virus H5N1, EURING, together with BTO and Wetlands International, and with financial support from DG Environment of the European Commission, developed a ‘Migration Mapping Tool’ as a mechanism for policymakers and others to assess migratory connectivity of waterbirds and the consequent potential for disease transmission between various parts of Europe, North Africa and the Middle East (Atkinson et al Citation2007; ). When, in February 2006, H5N1 reached the northeastern German island of Rügen, and shortly afterwards the southern German side of Lake Constance, existing ring-recovery data allowed the identification of the main origins of wintering waterbirds using this area. By combining this information with genetic data on the virus, it could be shown that two independent influx events happened at almost the same time (Globig et al Citation2009b). These findings provide an important illustration of the way that an epidemic involving this particular pathogen may develop.
Figure 4. Movements of Tufted Ducks Aythya fuligula between the Baltic region and Britain and Ireland as shown by the Migration Mapping Tool. Coloured kernels indicate the main concentrations of recoveries (50% of all recoveries in red, 95% in blue). The average migration route is shown by a line with numbers indicating the average location in each month. This is one of nine flyways identified for Tufted Ducks occurring in Britain and Ireland. From Atkinson et al (Citation2007).
Dispersal
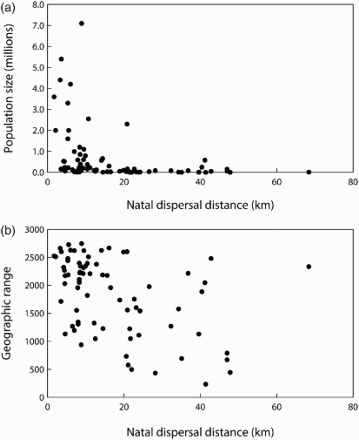
Dispersal is often defined as the movement of an individual between its location of birth and the place of first breeding (natal dispersal) and between subsequent breeding locations (breeding dispersal; Greenwood & Harvey Citation1982); it is an important individual behaviour affecting the spatial structure of populations and their dynamics, as well as gene flow. Dispersal studies typically focus either on the distribution of distances moved or on the probability of conducting a dispersal movement. Either way, such studies are possible only if individuals can be followed, and ringing has been used widely for this purpose (Bennetts et al Citation2001). For example, Paradis et al (Citation1998) compared dispersal distances of 75 European bird species based on recoveries of ringed individuals from Britain and Ireland. They found that interspecific differences in dispersal patterns were related to species ecology. Both natal and breeding dispersal were lower among abundant species and among species with large geographic ranges (), while migratory species showed greater dispersal than residents. Wetland species showed greater breeding dispersal than others, presumably reflecting the relatively patchy nature of wetland habitats.
Figure 5. Relationships of (a) population size and (b) range size (number of occupied 10-km squares in Britain) with mean natal dispersal, estimated from ring recoveries for 75 British bird species (population size: r = -0.794, P < 0.001; range size: r = -0.601, P < 0.001: both after controlling for phylogeny). Redrawn from Paradis et al (Citation1998).
Live recaptures or resightings of marked individuals sampled at different locations have been used to estimate dispersal probabilities between these locations by means of multistate capture–recapture models (Blums et al Citation2003, Schaub & Von Hirschheydt Citation2009). This provides a more precise approach than one based on dead recoveries but is generally limited to smaller spatial scales. A general limitation of using mark–recapture/resighting data for studying dispersal is imperfect detection: dispersal can be inferred only from some individuals, and these individuals are unlikely to be a random sample. If not corrected for, this typically results in a bias towards small dispersal distances and in an underestimation of dispersal probabilities, as individuals dispersing to locations that are not included in the study remain undetected. Promising approaches to reduce potential biases are the calculation of distance-related recruitment rates (van Noordwijk Citation2011) and the inclusion of spatial information in capture–recapture models (Schaub & Royle Citation2014). Thus, ring-recovery data and mark–recapture data offer complementary advantages and disadvantages for the study of dispersal. Ring-recovery data provide relatively crude dispersal distributions but can be applied to many species and are not constrained by sampling being restricted to particular locations. In contrast, mark–recapture data can provide much more detailed assessments but inference is normally limited to specific sampling locations and such studies are available for fewer species.
Survival, recruitment and breeding propensity
Survival, age at first breeding as a component of recruitment and breeding propensity are usually studied with ringed individuals, since individual identification is crucial for measuring these variables. The master data sources are dead recoveries of marked individuals and capture–recapture data. The main challenge in all these analyses is that observations are imperfect. Marked individuals that are still alive may not be seen and not all dead individuals are found. Therefore, statistical models that contain nuisance parameters such as the probability that a dead marked bird is found and reported or the probability that a marked individual is captured/sighted alive have to be used to analyse the data. There has been tremendous progress in the development of statistical methods for estimating demographic parameters from such data (Brownie et al Citation1985, Lebreton et al Citation1992, Williams et al Citation2002). User-friendly software is available which has resulted in wide use of these methods in demographic studies of birds (eg White & Burnham Citation1999). A website (www.phidot.org) providing links to software, discussion groups and future workshops is also available.
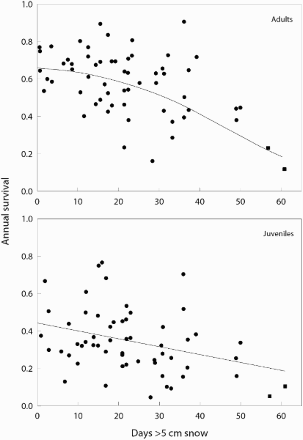
Although many demographic studies use capture–recapture data or ring recoveries collected by the researchers themselves, the use of data from the EDB and similar continental or national data sets have been particularly important for the study of long time series (eg Altwegg et al Citation2006, Robinson Citation2010) or for analyses at large spatial scales (Siriwardena et al Citation1998). Many of these studies reveal relationships with environmental factors that are likely to drive large-scale population changes (; Altwegg et al Citation2006). In Britain, national ring-recovery data sets have made an important contribution to explaining the demographic causes of population changes amongst a range of widespread species such as Lapwing Vanellus vanellus, Song Thrush Turdus philomelos and Blackbird T. merula (Peach et al Citation1994, Robinson et al Citation2004, Citation2012). More recently, the explanatory power of such analyses has been further improved through the use of integrated population models (Robinson et al Citation2012, Citation2014). Use of such data from the EDB itself has been seriously limited by the lack of information on cohort sizes of ringed samples, which is required to avoid an unrealistic assumption of constant ring-reporting rates. In the future this issue will be resolved through the incorporation of first-encounter data.
Figure 6. Survival of adult and juvenile Barn Owls Tyto alba in Switzerland, estimated from large-scale recapture/recovery data, in relation to the severity of winter weather. The two most extreme winters led to population crashes and are indicated with square symbols. Redrawn from Altwegg et al (Citation2006).
While traditional mark–recapture studies, such as those based on Constant Effort Sites or Retrapping Adults for Survival projects, focus mainly on survival (Peach et al Citation1999, Robinson et al Citation2008) more elaborate study designs allow a wider range of questions to be addressed. For example, if data are gathered from several locations, movement rates among locations can be estimated (Blums et al Citation2003). Other studies have addressed a wide range of issues including breeding propensity (Reed et al Citation2004), age at first breeding (Tavecchia et al Citation2001) and disease dynamics (Conn & Cooch Citation2009). All these analyses require the application of multistate capture–recapture models (Lebreton et al Citation2009). Methods have been developed that allow the estimation of the target parameters even if the states are partially unknown or recorded with errors (Kendall Citation2004, Pradel Citation2005).
Capture–recapture data from properly designed studies provide a rich source of information. Besides demographic parameters, the size of the population (Schwarz & Seber Citation1999), population density (Royle & Young Citation2008) and the population growth rate (Pradel Citation1996) can also be estimated. EURING analytical meetings have played an important role in the development of these methods (Thomson et al Citation2009, Schaub & Kendall Citation2012, Francis et al Citation2014).
Future opportunities and challenges
Mechanisms for data sharing
Large-scale sharing of data and information forms an essential part of contemporary research in many areas, and is particularly important when addressing large-scale processes such as bird migration and the effects of land use and climate change on population processes. However, it is essential to acknowledge the contributions of those individuals and organisations that collected the data, and to ensure that the outcomes of data sharing provide positive enhancement of the data-gathering process. These considerations are particularly important in the case of the long-term population studies which are so critical to our understanding of population processes and require the dedicated commitment of researchers over decades (Mills et al Citation2015). Where data gathering is funded wholly by public bodies, it may be appropriate for data to be available without restriction to anyone who wishes to use them. However, in many cases the collection of bird-ringing data is funded from a wide range of sources in addition to public funding, including charitable foundations, research councils, private individuals, independent organisations coordinating data gathering and analysis, and the ringers themselves. Most importantly, European ringing data gathering relies on a huge input of unpaid time from volunteers. Given all of these stakeholders, and the complex interrelationships between them, EURING believes that it has a responsibility to balance the desirability of open data with some limited controls that give appropriate priority to the interests of data gatherers. This is done by acknowledging that individual ringing centres are the primary owners of the data sets that they contribute to the EDB. These centres will normally be best placed to balance the requirements of data contributors and potential users, taking account of expectations, funding streams and specific circumstances amongst their own ringers, funders and data users.
This rationale provides the basis for operating the EDB. Its data holdings and availability are widely advertised via the EDB index. On receipt of a formal application all schemes are automatically consulted and have a set time in which to respond. Where necessary any issues around data access, conflicts with existing work and the need for collaboration can be discussed and resolved. In the vast majority of cases the data are released in a timely manner. The EDB does make some charges in order to cover part of its costs. In recent years it has had no other external source of funding, with the balance of costs being made up through modest contributions from EURING’s member schemes. This is despite much of the work being undertaken by a volunteer (CRdF) and the costs of building a new database having been funded by the BTO.
EURING is very keen to increase the availability of results from the EDB further, building particularly on success of the EDB index (du Feu et al Citation2016) and the Migration Mapping Tool (Atkinson et al Citation2007). The most obvious opportunity is to produce a significant online component towards a European Migration Atlas, with EURING providing maps and tables documenting the movements and migratory connectivity of many species. As the availability of first-capture data, moult data and measurements within the EDB develops, it may also be possible to provide online continental-scale information on annual cycles and demographic monitoring, complementing online information that is now available within some individual countries (eg Baillie et al Citation2014, Robinson et al Citation2015).
Quantifying movement patterns
Ringing and recovery data are not uniformly distributed in space or time. They depend on local or clustered catching activities as well as on heterogeneous finding and reporting probabilities of ringed birds. To date, most analyses of movement patterns based on ring recoveries have focused on the description of observed recovery distributions combined with qualitative interpretation of the observed patterns based on knowledge of human population density, recovery circumstances and other factors (examples above). It has been known for some time that, where numbers ringed and recovered across several areas are known, then quantitative inferences are possible (Kania & Busse Citation1987), but use and development of such methods has been hindered by the lack of availability of data on numbers ringed in different areas. Recently there has been increased interest in quantitative analyses of movement patterns based on ring recoveries, resulting in the development of a number of complementary statistical approaches (Korner-Nievergelt et al Citation2010, Thorup et al Citation2014). Inferences from such analyses may also be strengthened by incorporating multiple species so as to provide additional information on geographical variation in reporting rates (Cohen et al Citation2014).
There remain some circumstances in which recoveries can never provide a complete picture. In remote areas finding and reporting probabilities can be so low that the use of these areas by a certain bird species is unlikely to be recorded. In one of the first large-scale satellite-tracking studies of birds, Berthold et al (Citation2001) found that White Storks using the eastern Mediterranean flyway have an important early-winter resting area in Chad and western Sudan – an area where almost no ring recoveries originated. In these cases, tracking and geo-logging approaches provide vital additions to ring-recovery data. However, tracking studies remain expensive and in most cases will provide high-quality data for only relatively small numbers of individuals. Hence interpretation of these data from a few individuals – often carrying quite significant loads of electronic devices – will often need to be evaluated in combination with the findings revealed by ring recoveries. We envisage that future statistical developments will allow joint analyses of these different data types. Detailed tracking data then help to reduce bias while the ring-recovery data will continue to provide information at large temporal and spatial scales.
Demographic modelling
Demographic analyses based entirely or partially on marked individuals are likely to develop in the following directions. First, more sophisticated models will be applied more frequently, allowing some of the underlying assumptions of standard models to be relaxed. The development of Bayesian analyses in particular offers extreme flexibility in modelling (Kéry & Schaub Citation2012). For example, for modelling purposes it has usually been assumed that individuals are subject to identical demographic parameters (eg probability of surviving over a particular time period). The application of models incorporating individual random effects now makes it possible to study variability among individuals (Cam et al Citation2013) or the impact of morphological traits on demography (Gimenez et al Citation2006). Second, the combination of information originating from different data sources will become more widely used. For example, integrated population models (Besbeas et al Citation2002, Schaub & Abadi Citation2011, Robinson et al Citation2014) combine population survey data with demographic data (capture–recapture, recoveries, reproductive success). Advantages of joint analyses include more precise parameter estimates, the ability to estimate parameters that cannot be estimated with a separate analysis of the available data and the ability to test the consistency of different data sources. Third, the development of spatially explicit models is advancing rapidly (Royle et al Citation2013) offering the possibility of obtaining new insights into dispersal, together with improved estimates of survival and population density.
The main challenge for the EDB in meeting future needs in this area is that comprehensive first-encounter and local reencounter data should be stored, in addition to conventional live and dead recoveries. Survival analyses are often impossible if data on cohort sizes of ringed birds are not available. Ideally, capture data will be stored with individual covariates such as morphometrics, exact location and, where appropriate, factors such as breeding status. This would facilitate analyses that that incorporate the effects of factors such as morphology, sex or state. Finally, since the design of the studies has consequences for their analyses, there is a need to develop methods for storing relevant meta-data about study design.
Morphometrics, body condition and moult
The collection of data on measurements, weights and moult through structured networks of volunteer ringers is a potentially powerful way to address a range of biological questions (Bairlein Citation2003, Cresswell Citation2009). Thus the planned addition of such data to the EDB will open up a huge range of additional studies that were previously very difficult to undertake at a European scale. In a brief review of this kind it is not possible to detail all the potential research areas where such data could make an important contribution. Here, we highlight a few key examples of research based on these kinds of data.
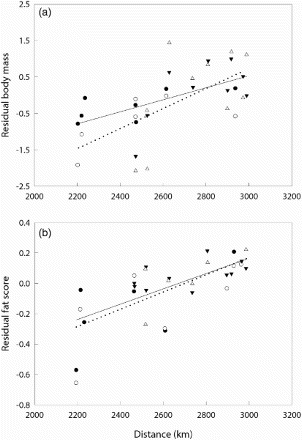
Body size usually varies geographically both within and between species and subspecies, so measurements can provide important supplementary information about the origins of migratory populations (Summers et al Citation1988, Pérez-Tris et al Citation1999). Weights and measurements also provide critical information on the accumulation of body reserves prior to migration, as has recently been illustrated by studies of Barn Swallow at both a country and European scale (Rubolini et al Citation2002, Coiffait et al Citation2011; ). To understand migration strategies fully and the role of stopover sites, such large-scale studies need to be complemented with more intensive mark–recapture/resighting investigations at a limited number of sites to measure length of stay and rate of fattening, and to understand the stopover decision rules used by migratory species (Schaub et al Citation2008, Bairlein & Schaub Citation2009).
Figure 7. Mean residual body mass (a) and fat score (b) of Barn Swallows Hirundo rustica for each sampling location (n = 10 or more individuals) in relation to linear distance between the sampling location and the southern margin of the Sahara Desert. Triangles, adults; circles, juveniles; filled symbols, Italian sites; open symbols, Iberian sites. Redrawn from Rubolini et al (Citation2002).
Another topical area of research drawing on large-scale data on weights and measurements involves the accumulation of winter fat reserves and the trade-off between the risks of starvation and predation (Cresswell Citation2009). In temperate and northern regions most small passerines accumulate substantial winter fat reserves to allow them to survive the longer winter nights and as an insurance against sudden cold spells and other unpredictable food shortages. An analysis of long-term trends in the weights of Great Tits Parus major across southern Britain found that winter weights increased when Sparrowhawk Accipiter nisus numbers were reduced due to pesticides, and then declined as the Sparrowhawk population recovered (Gosler et al Citation1995). These results were subsequently extended to show an interaction between the effects of predation and winter temperature on fat reserves. In colder winters, higher Sparrowhawk abundance was associated with lower Great Tit body mass while in warmer winners it was associated with higher Great Tit body mass (Cresswell et al Citation2009). This and other studies suggest that body-mass responses to environmental conditions may be subject to change as a result of both phenotypic plasticity and micro-evolutionary adaptations (Salewski et al Citation2010). Thus understanding the large-scale response of such processes to climate change is likely to be an important area for future research that the EDB can help to facilitate.
Moult, and in particular the main annual moult of the primary feathers, is a key component of the annual cycle of birds that can easily be studied by bird ringers (Kjellén Citation1994, Newton Citation2009). For most European bird species moult is a discrete phase of the annual cycle undertaken at a separate time from breeding or migration such that its timing and duration may be critical with respect to these other life-history events (Holmgren & Hedenström Citation1995, Hedenström Citation2008). Indeed, for many both resident and migratory populations the onset of primary moult may provide a good measure of the end of the nesting season (Newton Citation2009). In some migratory populations moult may be arrested to enable migration to proceed, with the extent of such arrested moult differing between populations in different parts of Europe in relation to specific life-history constraints (Mead & Watmough Citation1976, Swann & Baillie Citation1979). A recent study of 15 species undertaking primary moult within Britain found that, within species, the speed of primary moult varied according to the length of the breeding season, illustrating the importance of understanding the extent to which species are able to vary their moult patterns in relation to environmental change (Morrison et al Citation2015). In general, there is huge potential to increase our understanding of moult patterns through the types of large-scale analyses that could be facilitated by the EDB.
Synthesis
The EURING Data Bank provides a substantial and reasonably comprehensive resource for conducting analyses of what may be described as traditional ring-recovery data – ringing and reencounter records for birds that have subsequently been recovered. Such data have been used to address a wide range of questions involving the description of movements and mortality patterns, including the documentation of migration patterns, temporal and spatial variation in anthropogenic causes of death such as oil pollution and hunting, and the potential role of birds in disease transmission. With careful analysis and interpretation, such data can also be used to measure average survival rates and dispersal patterns, and can therefore contribute to studies of population dynamics designed to elucidate the demographic and environmental causes of population changes.
In order to model patterns of movements and survival formally and in more detail, it is usually necessary to have information on the numbers of first-encounter records, so that variation in ring reporting rates can be accounted for in the models. Furthermore, mark–recapture analyses can often provide more precise estimates of survival than ring recoveries, and for these local recaptures and a clear study design are needed. Thus a desirable future enhancement for the EDB, and also many individual ringing centres, would be to add data on the design of mark–recapture studies. Recent developments in the EDB, building on advances within individual ringing centres, will soon make it possible to incorporate large volumes of first captures and local recaptures within the Data Bank. This in turn will facilitate many more high-quality analyses at a European scale, addressing issues such as the causes of population changes in migratory species, the impacts of climate change, the role of protected areas in a changing environment and the effects of landscape configuration and connectivity on avian distributions and abundances. Further capability will be provided by the addition of data on measurements, weights and moult. While such data might appear to be of mainly academic interest they may in fact be crucial for understanding a range of applied issues, ranging from the effects of climate-related phenological changes on population dynamics to changes in the quality of wintering and staging areas under the influence of environmental change.
While we develop the capacity to address these important, challenging and exciting questions, EURING has identified an immediate priority for the production of a European Migration Atlas. This would provide a synthesis of information on movement patterns at a flyway scale that would be of immense value to policymakers, conservationists, ecological researchers and ornithologists. Its listing as one of the key evidence needs for the African–Eurasian Migrant Landbird Action Plan developed under the Convention on the Conservation of Migratory Species (CMS) clearly demonstrates its policy relevance. We envisage a series of high-level summaries of the movement patterns of individual species, backed by an online resource that would allow users to explore recovery patterns in more detail. We aim to support this with a series of cross-species reviews addressing issues such as the importance of key stopover areas for migratory species, the wise management of hunted populations, changing migratory strategies in the face of climate change and relationships between migratory behaviour and conservation status. In addressing all of these issues it must be stressed that ringing data are of most value when combined with other information sources, including counts, tracking data, genetic studies and many other types on information. Bird ringing has a great deal to offer conservation science, and the EDB provides a mechanism through which such research can be delivered at a range of scales.
Acknowledgements
We are very grateful to all of the European Ringing Centres, their ringers, staff, volunteers and Committee members, without whose input the EDB would not be a reality. We particularly thank the Netherlands Institute for Ecological Research (NIOO) and the BTO who have hosted the EDB and provided the technical and support infrastructure needed for it to operate. Rinse Wassenaar, former EDB manager at NIOO, made a huge contribution to the development of the EDB while Gerrit Speek led the development of the EURING 2000 code. Many former and present members of the EURING board and committees have made important contributions to developing the EDB, but we would particularly like to mention Franz Bairlein, the late Chris Mead, Arie van Noordwijk, the late Albert Perdeck, Pertti Saurola, the late Bob Spencer, Fernando Spina and the late Gerhard Zink for their unstinting support.
References
- Altwegg, R., Roulin, A., Kestenholz, M. & Jenni, L. (2006) Demographic effects of extreme winter weather in the barn owl. Oecologia 149, 44–51. doi: 10.1007/s00442-006-0430-3
- Ambrosini, R., Rubolini, D., Møller, A.P., Bani, L., Clark, J.A., Karcza, Z., Vangeluwe, D., du Feu, C., Spina, F. & Saino, N. (2011) Climate change and the long-term northward shift in the African wintering range of the barn swallow Hirundo rustica. Climate Research 49, 131–141. doi: 10.3354/cr01025
- Ambrosini, R., Borgoni, R., Rubolini, D., Sicurella, B., Fiedler, W., Bairlein, F., Baillie, S.R., Robinson, R.A., Clark, J.A., Spina, F. & Saino, N. (2014) Modelling the progression of bird migration with conditional autoregressive models applied to ringing data. PLoS ONE 9, e102440. doi: 10.1371/journal.pone.0102440
- Anon (1966) EURING Code Manual. European Union for Bird Ringing, Tring, UK.
- Atkinson, P.W., Robinson, R.A., Clark, J.A., Miyar, T., Downie, I.S., du Feu, C.R., Fiedler, W., Fransson, T., Grantham, M.J., Gschweng, M., Spina, F. & Crick, H.Q.P. (2007) Migratory movements of waterfowl: a web-based mapping tool. EURING report to the EU commission. http://blx1.bto.org/ai-eu
- Baillie, S.R., Marchant, J.H., Leech, D.I., Massimino, D., Sullivan, M.J.P., Eglington, S.M., Barimore, C., Dadam, D., Downie, I.S., Harris, S.J., Kew, A.J., Newson, S.E., Noble, D.G., Risely, K. & Robinson, R.A. (2014) BirdTrends 2014: trends in numbers, breeding success and survival for UK breeding birds. Research Report 624. BTO, Thetford. http://www.bto.org/about-birds/birdtrends/2014
- Bairlein, F. (2003) Large-scale networks in bird research in Europe: pitfalls and prospects. Avian Science 3, 49–63.
- Bairlein, F. & Schaub, M. (2009) Ringing and the study of mechanisms of migration. Ringing & Migration 24, 162–168. doi: 10.1080/03078698.2009.9674387
- Bairlein, F., Dierschke, J., Dierschke, V., Salewski, V., Geiter, O., Hüppop, K. & Köppen, U. (2014) Atlas des Vogelzugs: Ringfunde deutscher Brut- und Gastvögel. AULA-Verlag GmbH, Wiebelsheim.
- Bennetts, R.E., Nichols, J.D., Lebreton, J.-D., Pradel, R., Hines, J.E. & Kitchens, W.M. (2001) Methods for estimating dispersal probabilities and related parameters using marked animals. In Dispersal (ed Clobert, J.), 3–17. Oxford University Press, Oxford.
- Berthold, P., van den Bossche, W., Fiedler, W., Kaatz, C., Kaatz, M., Leshem, Y., Nowak, E. & Querner, U. (2001) Detection of a new important staging and wintering area of the White Stork Ciconia ciconia by satellite tracking. Ibis 143, 450–455. doi: 10.1111/j.1474-919X.2001.tb04946.x
- Besbeas, P., Freeman, S.N., Morgan, B.J.T. & Catchpole, E.A. (2002) Integrating mark–recapture–recovery and census data to estimate animal abundance and demographic parameters. Biometrics 58, 540–547. doi: 10.1111/j.0006-341X.2002.00540.x
- Blums, P., Nichols, J.D., Hines, J.E., Lindberg, M.S. & Mednis, A. (2003) Estimating natal dispersal movement rates of female European ducks with multistate modelling. Journal of Animal Ecology 72, 1027–1042. doi: 10.1046/j.1365-2656.2003.00774.x
- Bridge, E.S., Thorup, K., Bowlin, M.S., Chilson, P.B., Diehl, R.H., Fléron, R.W., Hartl, P., Kays, R., Kelly, J.F., Robinson, W.D. & Wikelski, M. (2011) Technology on the Move: Recent and Forthcoming Innovations for Tracking Migratory Birds. BioScience 61, 689–698. doi: 10.1525/bio.2011.61.9.7
- Brownie, C., Anderson, D.R., Burnham, K.P. & Robson, D.S. (1985) Statistical inference from band recovery data – a handbook. Resource Publication 156. US Fish and Wildlife Service, Washington, DC.
- Burnham, K.P. & Anderson, D.R. (1984) Tests of compensatory vs additive mortality in Mallards. Ecology 65, 105–112. doi: 10.2307/1939463
- Calladine, J., du Feu, C. & du Feu, R. (2012) Changing migration patterns of the Short-eared Owl Asio flammeus in Europe: An analysis of ringing recoveries. Journal of Ornithology 153, 691–698. doi: 10.1007/s10336-011-0786-y
- Cam, E., Gimenez, O., Alpizar-Jara, R., Aubry, L.M., Authier, M., Cooch, E.G., Koons, D.N., Link, W. A., Monnat, J.Y., Nichols, J.D., Rotella, J.J., Royle, J.A. & Pradel, R. (2013) Looking for a needle in a haystack: Inference about individual fitness components in a heterogeneous population. Oikos 122, 739–753. doi: 10.1111/j.1600-0706.2012.20532.x
- Cohen, E.B., Hostetler, J.A., Royle, J.A. & Marra, P.P. (2014) Estimating migratory connectivity of birds when re-encounter probabilities are heterogeneous. Ecology and Evolution 4, 1659–1670. doi: 10.1002/ece3.1059
- Coiffait, L., Robinson, R.A., Clark, J.A. & Griffin, B.M. (2011) Fattening strategies of British & Irish Barn Swallows Hirundo rustica prior to autumn migration. Ringing & Migration 26, 15–23. doi: 10.1080/03078698.2011.586583
- Conn, P.B. & Cooch, E.G. (2009) Multistate capture–recapture analysis under imperfect state observation: An application to disease models. Journal of Applied Ecology 46, 486–492. doi: 10.1111/j.1365-2664.2008.01597.x
- Cresswell, W. (2009) The use of mass and fat reserve measurements from ringing studies to assess body condition. Ringing & Migration 24, 227–232. doi: 10.1080/03078698.2009.9674396
- Cresswell, W., Clark, J.A. & Macleod, R. (2009) How climate change might influence the starvation-predation risk trade-off response. Proceedings. Biological sciences of the Royal Society of London B 276, 3553–3560.
- Csörgő, T., Karcza, Z., Halmos, G., Magyar, G., Gyurácz, J., Szép, T., Bankovics, A., Schmidt, A. & Schmidt, E. (2009) Hungarian Bird Migration Atlas. Kossuth Kiadó, Budapest.
- du Feu, C.R., Clark, J.A., Fiedler, W. & Baillie, S.R. (2010) EURING exchange code 2000+. Radolfzell, Germany.
- du Feu, C.R., Joys, A.C., Clark, J.A., Fiedler, W. & Baillie, S.R. (2016) EURING Databank species index. www.euring.org/data-and-codes/euring-databank-index
- Eglington, S.M., Julliard, R., Gargallo, G., van der Jeugd, H.P., Pearce-Higgins, J.W., Baillie, S.R. & Robinson, R.A. (2015) Latitudinal gradients in the productivity of European migrant warblers have not shifted northwards during a period of climate change. Global Ecology and Biogeography 24, 427–436. doi: 10.1111/geb.12267
- Elmberg, J., Nummi, P., Pöysä, H., Sjöberg, K., Gunnarsson, G., Clausen, P., Guillemain, M., Rodrigues, D. & Väänänen, V.-M. (2006) The scientific basis for new and sustainable management of migratory European ducks. Wildlife Biology 12, 121–127. doi: 10.2981/0909-6396(2006)12[121:TSBFNA]2.0.CO;2
- Fiedler, W. (2001) Large-scale ringing recovery analysis of European White Storks (Ciconia ciconia). The Ring 23, 73–79.
- Fiedler, W. (2009) New technologies for monitoring bird migration and behaviour. Ringing & Migration 24, 175–179. doi: 10.1080/03078698.2009.9674389
- Fink, D., Hochachka, W.M., Zuckerberg, B., Winkler, D.W., Shaby, B., Munson, M.A., Hooker, G., Riedewald, M., Sheldon, D. & Kelling, S. (2010) Spatiotemporal exploratory models for broad-scale survey data. Ecological Applications 20, 2131–2147. doi: 10.1890/09-1340.1
- Francis, C.M., Barker, R.J. & Cooch, E.G. (2014) Modelling demographic processes in marked populations. Proceedings of the EURING 2013 analytical meeting. Methods in Ecology and Evolution 5, 1265–1268.
- Gimenez, O., Covas, R., Brown, C.R., Anderson, M.D., Brown, M.B. & Lenormand, T. (2006) Nonparametric estimation of natural selection on a quantitative trait using mark–recapture data. Evolution 60, 460–466. doi: 10.1111/j.0014-3820.2006.tb01127.x
- Globig, A., Staubach, C., Beer, M., Köppen, U., Fiedler, W., Nieburg, M., Wilking, H., Starick, E., Teifke, J.P., Werner, O., Unger, F., Grund, C., Wolf, C., Roost, H., Feldhusen, F., Conraths, F.J., Mettenleiter, T.C. & Harder, T.C. (2009a) Epidemiological and ornithological aspects of outbreaks of highly pathogenic avian influenza virus H5N1 of Asian lineage in wild birds in Germany, 2006 and 2007. Transboundary and Emerging Diseases 56, 57–72. doi: 10.1111/j.1865-1682.2008.01061.x
- Globig, A., Baumer, A., Revilla-Fernández, S., Beer, M., Wodak, E., Fink, M., Greber, N., Harder, T.C., Wilking, H., Brunhart, I., Matthes, D., Kraatz, U., Strunk, P., Fiedler, W., Fereidouni, S.R., Staubach, C., Conraths, F.J., Griot, C., Mettenleiter, T.C. & Stärk, K.D.C. (2009b) Ducks as sentinels for avian influenza in wild birds. Emerging Infectious Diseases 15, 1633–1636. doi: 10.3201/eid1510.090439
- Gosler, A.G., Greenwood, J.J.D. & Perrins, C. (1995) Predation risk and the cost of being fat. Nature 377, 621–623. doi: 10.1038/377621a0
- Greenwood, P.J. & Harvey, P.H. (1982) The natal and breeding dispersal of birds. Annual Review of Ecology and Systematics 13, 1–21. doi: 10.1146/annurev.es.13.110182.000245
- Hampton, S.E., Strasser, C.A., Tewksbury, J.J., Gram, W.K., Budden, A.E., Batcheller, A.L., Duke, C.S. & Porter, J.H. (2013) Big data and the future of ecology. Frontiers in Ecology and the Environment 11, 156–162. doi: 10.1890/120103
- Hedenström, A. (2008) Adaptations to migration in birds: behavioural strategies, morphology and scaling effects. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences 363, 287–299. doi: 10.1098/rstb.2007.2140
- Hochachka, W.M. & Dhondt, A.A. (2006) House finch Carpodacus mexicanus population- and group-level responses to a bacterial disease. Ornithological Monographs 60, 30–43. doi: 10.1642/0078-6594(2006)60[30:HFCMPG]2.0.CO;2
- Holmgren, N. & Hedenström, A. (1995) The scheduling of molt in migratory birds. Evolutionary Ecology 9, 354–368. doi: 10.1007/BF01237759
- Hüppop, K. & Hüppop, O. (2002) Atlas zur Vogelberingung auf Helgoland Teil 1: Zeitliche und regionale Veränderungen der Wiederfundraten und Todesursachen auf Helgoland beringter Vögel (1909 bis 1998). Die Vogelwarte 41, 161–181.
- Johnston, A., Robinson, R.A., Gargallo, G., Julliard, R., van der Jeugd, H. & Baillie, S.R. (2016) Survival of Afro-Palaearctic passerine migrants in western Europe and the impacts of seasonal weather variables. Ibis 158, 465–480.
- Kania, W. & Busse, P. (1987) An analysis of the recovery distribution based on finding probabilities. Acta Ornithologica 23, 121–128.
- Kendall, W. (2004) Coping with unobservable and mis-classified states in capture–recapture studies. Animal Biodiversity and Conservation 27, 97–107.
- Kéry, M. & Schaub, M. (2012) Bayesian population analysis using WinBUGS. Academic Press, Amsterdam.
- Kjellén, N. (1994) Moult in relation to migration in birds – a review. Ornis Svecica 4, 1–24.
- Korner-Nievergelt, F., Sauter, A., Atkinson, P.W., Guélat, J., Kania, W., Kéry, M., Köppen, U., Robinson, R.A., Schaub, M., Thorup, K., van der Jeugd, H. & van Noordwijk, A.J. (2010) Improving the analysis of movement data from marked individuals through explicit estimation of observer heterogeneity. Journal of Avian Biology 41, 8–17. doi: 10.1111/j.1600-048X.2009.04907.x
- Kralj, J., Barišić, S., Ćiković, D. & Tutiš, V. (eds) (2013) Croatian Bird Migration Atlas. Croatian Academy of Sciences and Arts, Zagreb.
- Kylin, H., Bouwman, H. & Louette, M. (2011) Distributions of the subspecies of Lesser Black-backed Gulls Larus fuscus in sub-Saharan Africa. Bird Study 58, 186–192. doi: 10.1080/00063657.2011.555517
- Lebreton, J., Burnham, K.P., Clobert, J. & Anderson, D.R. (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecological Monographs 62, 67–118. doi: 10.2307/2937171
- Lebreton, J.-D., Nichols, J.D., Barker, R.J., Pradel, R. & Spendelow, J.A. (2009) Modeling individual animal histories with multistate capture–recapture models. In Advances in Ecological Research 41 (ed Caswell, H.), 87–173. Academic Press, Burlington.
- Liker, A. & Nagy, L. (2009) Migration of Mallards Anas platyrhynchos in Hungary: migration phenology, the origin of migrants, and long-term changes. Ringing & Migration 24, 259–265. doi: 10.1080/03078698.2009.9674400
- Mason, P. & Allsop, J. (2009) The Golden Oriole. T. & A.D. Poyser, London.
- McCulloch, M.N., Tucker, G.M. & Baillie, S.R. (1992) The hunting of migratory birds in Europe: a ringing recovery analysis. Ibis 134 (suppl. 1), 55–65.
- Mead, C.J. & Watmough, B.R. (1976) Suspended moult of trans-Saharan migrants in Iberia. Bird Study 23, 187–196. doi: 10.1080/00063657609476500
- Mead, C.J., Speek, B.J., Busse, P. & Jacquat, B. (1979) Code Manual: New EURING. European Union for Bird Ringing, Tring, UK.
- Michener, W.K. & Jones, M.B. (2012) Ecoinformatics: supporting ecology as a data-intensive science. Trends in Ecology and Evolution 27, 88–93. doi: 10.1016/j.tree.2011.11.016
- Mills, J.A., Teplitsky, C., Arroyo, B., Charmantier, A., Becker, P.H., Birkhead, T.R., Bize, P., Blumstein, D.T., Bonenfant, C., Boutin, S., Bushuev, A. & 52 others (2015) Archiving primary data: solutions for long-term studies. Trends in Ecology and Evolution 30, 581–589. doi: 10.1016/j.tree.2015.07.006
- Morrison, C.A., Baillie, S.R., Clark, J.A., Johnston, A., Leech, D.I. & Robinson, R.A. (2015) Flexibility in the timing of post-breeding moult in passerines in the UK. Ibis 157, 340–350. doi: 10.1111/ibi.12234
- Newton, I. (2009) Moult and plumage. Ringing & Migration 24, 220–226. doi: 10.1080/03078698.2009.9674395
- Nichols, J.D., Runge, M.C., Johnson, F.A. & Williams, B.K. (2007) Adaptive harvest management of North American waterfowl populations: a brief history and future prospects. Journal of Ornithology 148 (suppl. 2), 343–349. doi: 10.1007/s10336-007-0256-8
- Paradis, E., Baillie, S.R., Sutherland, W.J. & Gregory, R.D. (1998) Patterns of natal and breeding dispersal in birds. Journal of Animal Ecology 67, 518–536. doi: 10.1046/j.1365-2656.1998.00215.x
- Peach, W., Thompson, P. & Coulson, J. (1994) Annual and long-term variation in the survival rates of British lapwings Vanellus vanellus. Journal of Animal Ecology 63, 60–70. doi: 10.2307/5583
- Peach, W.J., Siriwardena, G.M. & Gregory, R.D. (1999) Long-term changes in over-winter survival rates explain the decline of reed buntings Emberiza schoeniclus in Britain. Journal of Applied Ecology 36, 798–811. doi: 10.1046/j.1365-2664.1999.00445.x
- Pérez-Tris, J., Carbonell, R. & Tellería, J.L. (1999) A method for differentiating between sedentary and migratory Blackcaps Sylvia atricapilla in wintering areas of southern Iberia. Bird Study 46, 299–304. doi: 10.1080/00063659909461142
- Pradel, R. (1996) Utilization of capture–mark–recapture for the study of recruitment and population growth rate. Biometrics 52, 703–709. doi: 10.2307/2532908
- Pradel, R. (2005) Multievent: an extension of multistate capture–recapture models to uncertain states. Biometrics 61, 442–447. doi: 10.1111/j.1541-0420.2005.00318.x
- Reed, E.T., Gauthier, G. & Giroux, J.F. (2004) Effects of spring conditions on breeding propensity of Greater Snow Goose females. Animal Biodiversity and Conservation 27, 35–46.
- Reichlin, T.S., Schaub, M., Menz, M.H.M., Mermod, M., Portner, P., Arlettaz, R. & Jenni, L. (2009) Migration patterns of Hoopoe Upupa epops and Wryneck Jynx torquilla: an analysis of European ring recoveries. Journal of Ornithology 150, 393–400. doi: 10.1007/s10336-008-0361-3
- Reichman, O.J., Jones, M.B. & Schildhauer, M.P. (2011) Challenges and opportunities of open data in ecology. Science 331, 703–705. doi: 10.1126/science.1197962
- Robinson, R.A. (2010) Estimating age-specific survival rates from historical ringing data. Ibis 152, 651–653. doi: 10.1111/j.1474-919X.2010.01032.x
- Robinson, R.A., Green, R.E., Baillie, S.R., Peach, W.J. & Thomson, D.L. (2004) Demographic mechanisms of the population decline of the song thrush Turdus philomelos in Britain. Journal of Animal Ecology 73, 670–682. doi: 10.1111/j.0021-8790.2004.00841.x
- Robinson, R.A., Balmer, D.E. & Marchant, J.H. (2008) Survival rates of hirundines in relation to British and African rainfall. Ringing & Migration 24, 1–6. doi: 10.1080/03078698.2008.9674375
- Robinson, R., Baillie, S. & King, R. (2012) Population processes in European blackbirds Turdus merula: a state–space approach. Journal of Ornithology 152, 419–422. doi: 10.1007/s10336-010-0612-y
- Robinson, R.A., Morrison, C.A. & Baillie, S.R. (2014) Integrating demographic data: towards a framework for monitoring wildlife populations at large spatial scales. Methods in Ecology and Evolution 5, 1361–1372. doi: 10.1111/2041-210X.12204
- Robinson, R.A., Leech, D.I. & Clark, J.A. (2015) The Online Demography Report: bird ringing and nest recording in Britain & Ireland in 2014. BTO, Thetford. www.bto.org/ringing-report [created 7 Sep 2015]
- Royle, J.A. & Young, K.V. (2008) A hierarchical model for spatial capture–recapture data. Ecology 89, 2281–2289. doi: 10.1890/07-0601.1
- Royle, J.A., Chandler, R.B., Sollmann, R. & Gardner, B. (2013) Spatial capture–recapture. Academic Press, New York.
- Rubolini, D., Pastor, A.G., Pilastro, A. & Spina, F. (2002) Ecological barriers shaping fuel stores in barn swallows Hirundo rustica following the central and western Mediterranean flyways. Journal of Avian Biology 33, 15–22.
- Salewski, V., Hochachka, W.M. & Fiedler, W. (2010) Global warming and Bergmann’s rule: do central European passerines adjust their body size to rising temperatures? Oecologia 162, 247–260. doi: 10.1007/s00442-009-1446-2
- Schaub, M. & Abadi, F. (2011) Integrated population models: a novel analysis framework for deeper insights into population dynamics. Journal of Ornithology 152 (suppl. 1), 227–237. doi: 10.1007/s10336-010-0632-7
- Schaub, M. & Kendall, W.L. (2012) Estimation, modelling and conservation of vertebrate populations using marked individuals. Journal of Ornithology 152 (suppl. 2), 291–292. doi: 10.1007/s10336-011-0738-6
- Schaub, M. & Lebreton, J.-D. (2004) Testing the additive versus the compensatory hypothesis of mortality from ring recovery data using a random effects model. Animal Biodiversity and Conservation 27, 73–85.
- Schaub, M. & Pradel, R. (2004) Assessing the relative importance of different sources of mortality from recoveries of marked animals. Ecology 85, 930–938. doi: 10.1890/03-0012
- Schaub, M. & Royle, J.A. (2014) Estimating true instead of apparent survival using spatial Cormack–Jolly–Seber models. Methods in Ecology and Evolution 5, 1316–1326. doi: 10.1111/2041-210X.12134
- Schaub, M. & Von Hirschheydt, J. (2009) Effect of current reproduction on apparent survival, breeding dispersal, and future reproduction in barn swallows assessed by multistate capture–recapture models. Journal of Animal Ecology 78, 625–635. doi: 10.1111/j.1365-2656.2008.01508.x
- Schaub, M., Jenni, L. & Bairlein, F. (2008) Fuel stores, fuel accumulation, and the decision to depart from a migration stopover site. Behavioral Ecology 19, 657–666. doi: 10.1093/beheco/arn023
- Schimel, D. (2011) The era of continental-scale ecology. Frontiers in Ecology and the Environment 9, 311. doi: 10.1890/1540-9295-9.6.311
- Schwarz, C.J. & Seber, G.A.F. (1999) Estimating animal abundance: review III. Statistical Science 14, 427–456. doi: 10.1214/ss/1009212521
- Servanty, S., Choquet, R., Baubet, É., Brandt, S., Gaillard, J., Schaub, M., Toïgo, C., Lebreton, J., Buoro, M. & Gimenez, O. (2010) Assessing whether mortality is additive using marked animals: a Bayesian state-space modeling approach. Ecology 91, 1916–1923. doi: 10.1890/09-1931.1
- Siriwardena, G.M., Baillie, S.R. & Wilson, J.D. (1998) Variation in the survival rates of some British passerines with respect to their population trends on farmland. Bird Study 45, 276–292. doi: 10.1080/00063659809461099
- Speek, G., Clark, J.A., Rohde, Z., Wassenaar, R. & van Noordwijk, A.J. (2001) The EURING exchange-code 2000. Heteren, Netherlands.
- Spina, F. & Volponi, S. (2009) Atlante della Migrazione degli Uccelli in Italia. Volume 2: Passeriformi. ISPRA-MATTM, Rome.
- Summers, R.W., Nicoll, M., Underhill, L.G. & Petersen, A. (1988) Methods for estimating the proportions of Icelandic and British Redshanks Tringa totanus in mixed populations wintering on British coasts. Bird Study 35, 169–180. doi: 10.1080/00063658809476986
- Swann, R.L. & Baillie, S.R. (1979) The suspension of moult by trans-Saharan migrants in Crete. Bird Study 26, 55–58. doi: 10.1080/00063657909476617
- Tavecchia, G., Pradel, R., Boy, V., Johnson, A.R. & Cézilly, F. (2001) Sex- and age-related variation in survival and cost of first reproduction in Greater Flamingos. Ecology 82, 165–174. doi: 10.1890/0012-9658(2001)082[0165:SAARVI]2.0.CO;2
- Thomson, D.L., Cooch, E.G. & Conroy, M.J. (2009) Modeling demographic processes in marked populations. Proceedings of EURING 2007 Analytical Conference. Springer, New York.
- Thorup, K., Korner-Nievergelt, F., Cohen, E.B. & Baillie, S.R. (2014) Large-scale spatial analysis of ringing and re-encounter data to infer movement patterns: a review including methodological perspectives. Methods in Ecology and Evolution 5, 1337–1350. doi: 10.1111/2041-210X.12258
- Valkama, J., Saurola, P., Lehikoinen, A., Lehikoinen, E., Piha, M., Sola, P. & Velmala, W. (2014) The Finnish Bird Ringing Atlas. Volume 2. Finnish Museum of Natural History and Ministry of Environment, Helsinki.
- van Noordwijk, A.J. (2011) Measuring dispersal as distance-dependent recruitment rates: testing the performance of DDRR on simulated data. Journal of Ornithology 152 (suppl. 1), 239–249. doi: 10.1007/s10336-010-0637-2
- Votier, S.C., Birkhead, T.R., Oro, D., Trinder, M., Grantham, M.J., Clark, J.A., McCleery, R.H. & Hatchwell, B.J. (2008) Recruitment and survival of immature seabirds in relation to oil spills and climate variability. Journal of Animal Ecology 77, 974–983. doi: 10.1111/j.1365-2656.2008.01421.x
- White, G.C. & Burnham, K.P. (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46 (suppl. 1), 120–139. doi: 10.1080/00063659909477239
- Williams, B.K., Nichols, J.D. & Conroy, M.J. (2002) Analysis and management of animal populations. Academic Press, San Diego.
Appendix
Useful websites referred to in the text: