Abstract
The aim of the present study was to evaluate a potential role of different passerine birds species in Mazurian Lake region (northeast Poland) in the spread of Borrelia burgdorferi sensu lato, the spirochaete that causes Lyme disease. A total number of 1254 birds (representing 42 species) were captured during the 3-year study period. Blood samples were collected from birds and analyzed with a nested polymerase chain reaction technique in order to detect fragments of the pathogen DNA. Positive results were obtained in 4.2% of all blood samples. Specifically, B. burgdorferi s.l. were detected in tree pipit (Anthus Erivialis; 21.1% of 19 birds), dunnock (Prunella modularis; 15.8% of 19 birds), chaffinch (Fringilla coelebs; 12.7% of 166 birds), song thrush (Turdus philometos; 9.3% of 54 birds), nuthatch (Sitta euopea; 7.7% of 26 birds), hawfinch (Coccothraustes coccothroustes; 6.7% of 15 birds), robin (Erithacus rebecula; 5.1% of 256 birds), blackbird (Turdus merula; 4.2% of 71 birds) and wren (Troglodytes troglodytes; 3.7% of 27 birds). Additionally, the incidence of the infection was analyzed in relation to the habitat in which the birds resided (mixed coniferous forest or alder swamp forest), months of the study (from April to October), age and sex, but the differences were not statistically significant.
1 Introduction
Lyme disease, caused by the spirochaete Borrelia burgdorferi sensu lato, is the most common tick-borne zoonosis in the world, manifesting with multisystemic disorders in humans (Steere, Citation1989). B. burgdorferi s.l. is transmitted mainly by ticks of the Ixodes ricinus group. These tick species parasitize various mammals, birds, and reptiles. Infected nymphs or adult females may transmit B. burgdorferi s.l. to vertebrates, which have a reservoir role (i.e. the spirochaetes persist for a considerable time), and they can infect engorging larval ticks, non-infected nymphs as well as adult ticks. Small and medium-sized mammals are the main vertebrate reservoirs of B. burgdorferi s.l. (Mather et al., Citation1989; Humair et al., Citation1993a; Tälleklint & Jaenson, Citation1994). The importance of birds as additional reservoir hosts is a subject of growing interest.
It is well documented that numerous species of passerine birds are frequently infested with ticks (Pruett-Jones & Pruett-Jones, Citation1991; Battaly & Fish, Citation1993; Humair et al., Citation1993b; Hubálek et al., Citation1996; Kinsey et al., Citation2000). Moreover, the role of these avian hosts in spreading ticks to other sites has been shown (Olsén et al., Citation1995; Smith et al., Citation1996; Durden et al., Citation2001; Scott et al., Citation2001; Slowik & Lane, Citation2001). On the contrary, B. burgdorferi s.l. has been isolated from the blood (Burgess et al., Citation1993; Humair et al., Citation1993b; McLean et al., Citation1993; Olsén et al., Citation1996; Durden et al., Citation1997), from skin biopsies (Miyamoto et al., Citation1997; Humair et al., Citation1998; Durden et al., Citation2001) and internal organs (e.g. liver; Anderson et al., Citation1986; Miyamoto et al., Citation1997) of passerine birds. The pathogen has been also detected in blood and foot web tissues of seabirds (Olsén et al., Citation1993; Gylfe et al., Citation1999). However, there is insufficient data to assess whether birds have the ability not only to maintain and multiply, but also to transmit the pathogen to ticks.
The aim of the present study was to establish whether the spirochaete is found in the passerine bird community resident in the Mazurian Lake region, and if there are any differences between avian species in the presence of B. burgdorferi s.l.
The most valuable investigative approach is larval xenodiagnosis (Humair et al., Citation1998; Kurtenbach et al., Citation1998; Richter et al., Citation2000), but not all vertebrate species are amenable to it because they are difficult to capture and maintain under controlled conditions. For the latter reasons, in the present study detection of B. burgdorferi s.l. DNA in the birds blood was employed.
The Mazurian Lake region, located in northeastern Poland, was selected as the study site as it is considered as an endemic area of B. burgdorferi s.l. infection in ticks (Sinski & Karbowiak, Citation1994; Wegner et al., Citation1995). Second, it is also a region of abundance of the most common tick (I. ricinus L.) in Poland (Lachmajer, Citation1967; Siuda, Citation1993). Furthermore, many studies revealed B. burgdorferi or antibodies to B. burgdorferi in humans living in this region and reporting frequent contacts with ticks (Prokopowicz, Citation1995; Prokopowicz et al., Citation1995; Pancewicz et al., Citation1996).
2 Materials and Methods
2.1 Bird sampling
Birds were captured in two types of habitats: mixed coniferous forest (Pino—Quercetum) and alder swamp forest (Carici elongatae—Alnetum), in places where the abundance of undergrowth and shrub plants was similar. The catches were performed in 1996, 1997 and 1998 during the seasons of the highest tick activity; that is, from April to October (permissions OP 4072/230/97, OPog 4201/103/98). Twenty standard ‘mist nets’ were placed in both habitats and examined at 1-h intervals. Investigations of both forests took place during a subsequent full 6 days and 6 nights for 7 consecutive months.
The species and, if possible, the age and sex of the captured birds were determined according to Svensson (Citation1992). A small amount of blood (approximately 50 μl) was collected from the brachial vein by the method of Kruszewicz (Citation1995) from individuals presenting in good physical condition. The blood samples were frozen at –20°C. After blood sampling the birds were ringed. Then the birds were released into the same habitats in which they had just been caught. Individuals recaptured within the same month were released immediately and were not recorded in the data analysis, whereas birds recaptured in other months or years of the study were analyzed separately and the distinct samples were tested each time.
2.2 Polymerase chain reaction
All the blood samples were analyzed with a nested plymerase chain reaction (PCR) technique in order to detect any fragments of B. burgdorferi s.l. DNA. The DNA was isolated by the procedure of Stanczak et al. (Citation1995). Briefly, the DNA was extracted once with an equal volume of water-saturated phenol and once with an equal volume of chloroform. Centrifugation was at 15 000×g for 15 min at 4°C. The supernatant was placed into a fresh tube and an equal volume of isopropyl alcohol was added to precipitate the DNA. Then the reaction mixture was incubated overnight at –20°C and the DNA was spun down at 15 000×g for 15 min at 4°C. The supernatant was then removed, DNA was washed with 200 μl cold 70% ethanol, centrifuged (at 15 000 x g for 15 min at 4°C), dried (for 3 min at 72°C) and dissolved in 60 μl sterile water. Amplification of DNA was performed by the nested PCR technique according to Valsangiacomo et al. (Citation1996) and based on two-stage amplification of the fragment of hbb gene coding a highly preserved histon protein of B. burgdorferi s.l. The first stage product is 433 base pairs in size and contains the whole hbb gene, and the second stage product is a 184 base pairs fragment of the hbb gene. Genomic DNA of B. burgdorferi (DNA; Gdansk II) was used as a positive control, and reagents as a negative control. The cycling conditions were 94°C for 1 min, 52°C for 1 min and then 72°C for 30 sec for a total of 35 cycles in the first stage and 25 cycles in the second stage in an automated thermal cycler (GeneAmp PCR System 9600; Perkin Elmer).
The second-stage products were separated in 2% agarose gel and visualized by 3 μl ethidium bromide staining. Electrophoresis was performed during 30 min with the current intensity of 120 mA. The M1 marker (pUC19/MspI DNA; Gdansk II) was used as a molecular mass standard of DNA ().
Fig. 1 Agarose gel demonstrating PCR detection of Borrelia burgdorferi s.l. DNA in passerine bird blood.
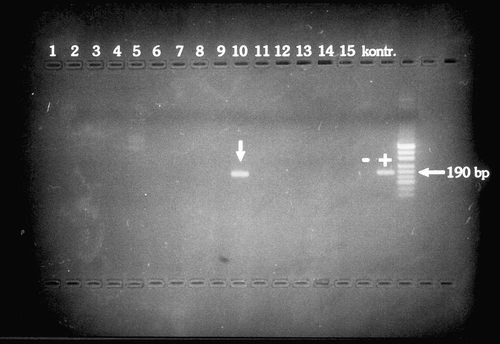
3 Results
3.1 Birds investigated
During the 3-year study period a total number of 1254 blood samples were collected () (277 blood samples in 1996, 448 in 1997 and 529 in 1998). Of these, 100 individuals were recaptures. In the study site, the most numerous species were robin, great tit and chaffinch; 256, 183 and 166 specimens were caught and investigated, respectively.
Passerine birds examined for B. Burgdorferi s.l. infection in northeastern Poland
3.2 Detection of B. burgdorferi s.l. by PCR
B. burgdorferi s.l. was found in blood of 4.2% of all analyzed birds (n=1254). Positive samples were observed in nine of the 42 avian species examined (). Of the recaptured birds (n=100) only one bird (chaffinch) that was caught in July and August 1998 was positive for B. burgdorferi s.l. each time. The general prevalence of bird infections with B. burgdorferi s.l. was similar within the three consecutive seasons: in 1996 it was 3.6% of the 277 individuals, in 1997 it was 5.1% of 448 individuals, and in 1998 it was 3.8% of 528 individuals.
The highest prevalence (more then 10%) was measured for the tree pipit, dunnock and chaffinch; 21.1% of 19 birds, 15.8% of 19 birds and 12.7% of 166 birds, respectively, were infected (). For groups of frequently captured birds (more than 15 representatives), statistical analysis revealed significant differences of infection rates between the bird species (χ2=75, degrees of freedom (df)=19, P<0.0001).
Of all infected birds, 57% (n=53) were captured in the mixed coniferous forest. These infected birds represented 5.2% of the total number of the birds captured and examined in this type of forest (n=577), compared with 3.4% of total number of the birds captured and examined in alder swamp forest (n=677); however, the difference was not statistically significant (χ2=2.5, df=1, not significant (NS)). In the next step, analysis was made only for the bird species that significantly preferred one of the forest habitat. This analysis revealed that birds preferring mixed coniferous forest were more frequently B. burgdorferi s.l. carriers than birds preferring alder swamp forest (χ2=13.85, df=1, P<0.0002).
The infection rates changed by month; however, the alterations were not statistically significant (χ2=4.4, df=6, NS) ().
Fig. 2 Changes in Borrelia burgdorferi s.l. infection by season.
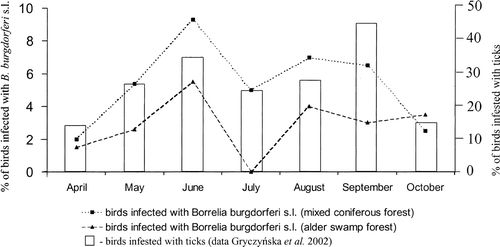
Analyses with respect to age and sex were also performed. Young birds (up to 1 year old; n=520) were infected in 5% of cases, and the older population (n=662) was infected in 4.2%; however, the difference was not statistically significant (χ2=0.4, df=1, NS). Similarly, the sex analysis (only species with sex dimorphism were considered) revealed no significant differences. Bacterial DNA was found in 4.3% of 185 females and 5.2% of 347 males (χ2=0.2, df=1, NS).
4 Discussion
The etiology and transmission of tick-borne diseases are subjects of rapidly growing interest during recent years. Research studies have attempted to determine not only which vertebrate species are the most frequent tick hosts, but have also focused on their potential role as reservoir hosts; that is, their ability to participate in the circulation of the pathogen in nature (Mather et al., Citation1989; Tälleklint & Jaenson, Citation1994). Another important factor is also quantitative analysis to determine the infection rate with the pathogen. The overall findings of the present study suggest that the infection rate with B. burgdorferi s.l. is markedly lower in birds when compared with rodents (4.2% versus 34.8 to 75%) (Humair et al., Citation1993a; Sinski & Karbowiak, Citation1994). This may suggest the lower importance of birds as hosts and reservoirs of B. burgdorferi s.l. in comparison with small forest rodents. A key consideration, apart from the low average prevalence rates for all the birds, is whether there are differences in infection rate between different bird species. The present results suggest that within a resident passerine bird community of the study area there are some bird species potentially spreading the bacteria B. burgdorferi s.l. more than others (), making it possible that some avian species could be considered important in maintaining the B. burgdorferi s.l. population in nature. The European Union Concerted Action on Lyme Borreliosis (EUCALB) (Smith et al., Citation1998) has published a list of vertebrate species considered effective reservoirs of the bacteria B. burgdorferi s.l. (Gern et al., Citation1998). In the present study, seven of the species of passerine birds listed by EUCLAB as reservoirs were found to be infected. Additionally, infection was detected in two other bird species frequently found in the study area (i.e. nuthatch and dunnock). Some bird species listed by EUCALB as participating in the survival of B. burgdorferi s.l. population were not found to have the pathogen in the present study. However, three of these species (whitethroat, redstart and thrush nightingale) are only incidentally found in the study area (up to five caught individuals), which may explain negative results. However, both the great tit and the blackcap were very frequently captured birds in this study (183 and 120 specimens, respectively), but none was shown to be pathogen-positive. This may reflect very low rates of infestation of both bird species by ticks in the study area (Gryczynska et al., Citation2002). Great tits are of a bird group preferring upper zones of forest that limit their contact with ticks. Blackcaps do not avoid bottom zones of the forest; their low level of infestation may be due to their migration behavior. These birds are found in the study area relatively late (i.e. during May) and start their autumn return rather early (i.e. August). As a consequence they may evade contact with ticks, which are not found to show marked activity before September.
The differences in infection rate between bird species may result from many reasons. However, the most frequently infected bird species are those with the highest prevalence of infestation of the vector (i.e. I. ricinus tick) (Gryczynska et al., Citation2002).
In mixed coniferous forest, there was a higher percentage of B. burgdorferi s.l.-infected birds than in the alder swamp forest. Probably, this results from a higher prevalence of tick infestation in birds of this type of forest (Gryczynska et al., Citation2002). The hypothesis that passerine birds play a greater role in spreading of B. burgdorferi s.l. in mixed coniferous forest than in alder swamp forest is supported by the differences in infection prevalence between two bird species preferring one habitat or the other. Such comparison was performed for tree pipits and blackbirds, preferring mixed coniferous forest and alder swamp forest, respectively. The presence of B. burgdorfresi s.l. in blood was found more frequently in tree pipits than in blackbirds (χ2=5.92, df=1, P<0.015), even though tick infestation was found at similar frequency in these two species (Gryczynska et al., Citation2002).
The changes in the incidence of infected birds during consecutive months suggest that passerine birds may be a carrier of the pathogen only for short period of time. The incidence of infected birds increased during the first 3 months of the study in the both type of a forest, and this corresponds with the increase of the total population of ticks in their environment (Gryczynska et al., Citation2002). Then, the incidence of infected birds went down during the period of lower tick activity (July, August). Accordingly, the incidence of infected individuals of the birds living in the same area does not increase as months go by, but it strictly depends on the presence (and quantity) of infected ticks in this area. This hypothesis is also supported by the fact that older birds did not appear to be more frequently infected than 1-year-old individuals. On the contrary, however, it may be that the reducing pathogen incidence in birds with time may be related to an increased mortality of birds carrying the pathogen and earlier elimination of infected individuals in the environment. This hypothesis is supported by the fact that of 100 birds captured more then once only the one individual was positive both during the first and second catching (1-month interval), whereas other infected birds were not caught again.
In conclusion, it is suggested that passerine birds may play a role as a reservoir in the process of B. burgdorferi s.l. spread in the forest environment; however, their host competence is dependent on the bird species. There is a need to determine whether B. burgdorferi s.l.-positive avian species are able to transmit the spirochaete to ticks and thus to have reservoir competence. There is also a need to analyze B. burgdorferi s.l. genomospecies, as a few reports suggest that different groups of vertebrates are reservoirs for specific bacteria genomospecies (Hanincova et al., Citation2003a,bCitation, Humair et al., Citation1995; Olsén et al., Citation1995). It is also known that ticks may be carriers both of one and many bacteria genomospecies (mixed infections) (Strle et al., Citation1995; Rijpkema et al., Citation1995; Saint Girons et al., Citation1998), and the possibility exists that there are two or more independent cycles maintaining stable specific populations of the bacteria genomospecies in their environment providing distinct zoonotic reservoirs (Kurtenbach et al., Citation2002). Various genomospecies of bacteria B. burgdorferi s.l. may be responsible for different forms of borrelliosis (Canica et al., Citation1993; van Dam et al., Citation1993).
Acknowledgments
The study was partially supported by Warsaw University grants BW-1345/15, BW-1385/9 and BW-98/1. M.S. is supported by a Young Scientist Award from the Foundation for Polish Science, Warszawa, Poland.
Related Research Data
References
- Anderson , J.F. , Johnson , R.C. , Magnarelli , L.A. and Hyde , F.W. 1986 . Involvement of birds in the epidemiology of the Lyme disease agent Borrelia burgdorferi . Infection and Immunity , 51 : 394 – 396 .
- Battaly , G.R. and Fish , D. 1993 . Relative importance of bird species as hosts for immature Ixodes dammini (Acari: Ixodidae) in a suburban residential landscape of southern New York State . Journal of Medical Entomology , 30 : 740 – 747 .
- Burgess , E.C. , Wachal , M.D. and Cleven , T.D. 1993 . Borrelia burgdorferi infection in dairy cows, rodents, and birds from four Wisconsin dairy farms . Veterinary Microbiology , 35 : 61 – 77 .
- Canica , M.M. , Nato , F. , du Merle , L. , Mazie , J.C. , Baranton , G. and Postic , D. 1993 . Monoclonal antibodies for identification of Borrelia afzelii sp.nov. associated with late cutaneous manifestations of Lyme borreliosis . Scandinavian Journal of Infectious Diseases , 25 : 441 – 448 .
- Durden , L.A. , McLean , R.G. , Oliver , J.H., Jr. , Ubico , S.R. and James , A.M. 1997 . Ticks, Lyme disease spirochetes, trypanosomes, and antibody to encephalitis viruses in wild birds from coastal Georgia and South Carolina . Journal of Parasitology , 83 : 1178 – 1182 .
- Durden , L.A. , Oliver , J.H., Jr. and Kinsey , A.A. 2001 . Ticks (Acari: Ixodidae) and spirochaetes (Spirochaetaceae: Spirochaetales) recovered from birds on a Georgia Barrier Island . Journal of Medical Entomology , 38 : 231 – 236 .
- Gern , L. , Estrada-Peň , A. , Frandsen , F. , Gray , J.S. , Jaenson , T.G.T. , Jongejan , F. , Kahl , O. , Korenberg , E. , Mehl , R. and Nuttall , P.A. 1998 . European reservoir hosts of Borrelia burgdorferi sensu lato . Zentralblatt für Bakteriologie , 287 : 196 – 204 .
- Gryczyńska , A. , Barkowska , M. and Siemiatkowski , M. 2002 . Analysis of Ixodes ricinus (L.) tick burdens in a resident passerine bird community in the Mazurian Lake region (Northeastern Poland) . Acta Parasitologica , 47 : 51 – 57 .
- Gylfe , Å. , Olsen , B. , Straševičius , D. , Marti Ras , N. , Weihe , P. , Noppa , L. , Östberg , Y. , Baranton , G. and Bergström , S. 1999 . Isolation of Lyme Disease Borrelia from puffins (Fratercula arctica) and seabird ticks (Ixodes uriae) on the Faeroe Islands . Journal of Clinical Microbiology , 37 : 890 – 896 .
- Hanincova , K. , Schafer , S.M. , Etti , S. , Sewell , H.S. , Taragelova , V. , Ziak , D. , Labuda , M. and Kurtenbach , K. 2003a . Association of Borrelia afzelii with rodents in Europe . Parasitology , 126 : 11 – 20 .
- Hanincova , K. , Taragelova , V. , Koci , J. , Schafer , S.M. , Hails , R. , Ullmann , A.J. , Piesman , J. , Labuda , M. and Kurtenbach , K. 2003b . Association of Borrelia garinii and B. valaisiana with songbirds in Slovakia . Applied and Environmental Microbiology , 69 : 2825 – 2830 .
- Hubálek , Z. , Anderson , J.F. , Halouzka , J. and Hájek , V. 1996 . Borreliae in immature Ixodes ricinus (Acari: Ixodidae) ticks parasitizing birds in the Czech Republic . Journal of Medical Entomology , 33 : 766 – 771 .
- Humair , P.F. , Turrian , N. , Aeschlimann , A. and Gern , L. 1993a . Borrelia burgdorferi in a focus of Lyme borreliosis: epizootiologic contribution of small mammals . Folia Parasitologica , 40 : 65 – 70 .
- Humair , P.F. , Turrian , N. , Aeschlimann , A. and Gern , L. 1993b . Ixodes ricinus immatures on birds in a focus of Lyme borreliosis . Folia Parasitologica , 40 : 237 – 242 .
- Humair , P.F. , Peter , O. , Wallich , R. and Gern , L. 1995 . Strain variation of Lyme disease spirochaetes isolated from Ixodes ricinus ticks and rodents collected in two endemic areas in Switzerland . Journal of Medical Entomology , 32 : 433 – 438 .
- Humair , P.F. , Postic , D. , Wallich , R. and Gern , L. 1998 . An avian reservoir (Turdus merula) of the Lyme borreliosis spirochetes . Zentralblatt für Bakteriologie , 287 : 521 – 538 .
- Kinsey , A.A. , Durden , L.A. and Oliver , J.H., Jr . 2000 . Tick infestations of birds in coastal Georgia and Alabama . Journal of Parasitology , 86 : 251 – 254 .
- Kruszewicz , A.G. 1995 . Zwia˛zki parazytologii z ornitologia˛. II Przeglad systematyczny pasożytów i możliwości diagnozowania inwazji . Notatki Ornitologiczne , 36 : 141 – 150 .
- Kurtenbach , K. , Peacey , M. , Rijpkema , S.G.T. , Hoodless , A.N. , Nuttall , P.A. and Randolph , S.E. 1998 . Differential transmission of the genospecies of Borrelia burgdorferi sensu lato by game birds and small rodents in England . Applied and Environmental Microbiology , 64 : 1169 – 1174 .
- Kurtenbach , K. , De Michelis , S. , Etti , S. , Schafer , S.M. , Sewell , H.S. , Brade , V. and Kraiczy , P. 2002 . Host association of Borrelia burgdorferi sensu lato—the key role of host complement . Trends in Microbiology , 10 : 74 – 79 .
- Lachmajer , J. 1967 . Species composition and distribution of Ixodoidea (Acarina) in Poland . Wiadomosci Parazytologiczne , 13 : 511 – 515 .
- Mather , T.N. , Wilson , M.L. , Moore , S.I. , Ribeiro , J.M.C. and Spielman , A. 1989 . Comparing the relative potential of rodents as reservoirs of the Lyme disease spirochaete (Borrelia burgdorferi) . American Journal of Epidemiology , 130 : 143 – 150 .
- McLean , R.G. , Ubico , S.R. , Norton Hughes , C.A. , Engstrom , S.M. and Johnson , R.C. 1993 . Isolation and characterization of Borrelia burgdorferi from blood of a bird captured in the Saint Croix River Valley . Journal of Clinical Microbiology , 31 : 2038 – 2043 .
- Miyamoto , K. , Sato , Y. , Okada , K. , Fukunaga , M. and Sato , F. 1997 . Competence of a migratory bird, red-bellied thrush (Turdus chrysolaus), as an avian reservoir for the Lyme disease spirochaetes in Japan . Acta Tropica , 65 : 43 – 51 .
- Olsén , B. , Jaenson , T.G.T. , Noppa , L. , Bunikis , J. and Bergström , S. 1993 . A Lyme borreliosis cycle in seabirds and Ixodes uriae ticks . Nature , 362 : 340 – 342 .
- Olsén , B. , Jaenson , T.G.T. and Bergström , S. 1995 . Prevalence of Borrelia burgdorferi sensu lato-infected ticks on migrating birds . Applied and Environmental Microbiology , 61 : 3082 – 3087 .
- Olsén , B. , Gylfe , Å. and Bergström , S. 1996 . Canary finches (Serinus canaria) as an avian infection model for Lyme borreliosis . Microbial Pathogenesis , 20 : 319 – 324 .
- Pancewicz , S.A. , Januszkiewicz , A. and Hermanowska-Szpakowicz , T. 1996 . Obecnosc przeciwcial przeciwko Borrelia burgdorferi wśród mieszkańców pólnocno-wschodniej Polski . Przegla˛d Epidemiologiczny , 50 : 375 – 381 .
- Prokopowicz, D. (1995). Choroby przenoszone przez kleszcze 1st edn. Warszawa: Wydawnictwo Fundacji Büchnera.
- Prokopowicz, D., Grzeszczuk, A., Wiercińska-Drapalo, A. & Flisiak, R. (1995). The role of tick-vector in transmission of tick-borne encephalitis and Lyme borreliosis in North-Eastern Poland. Programme and Abstracts IX European Meeting of Society For Vector Ecology (p. 44). Prague, Czech Republic.
- Pruett-Jones, M. & Pruett-Jones, S. (1991). Analysis and ecological correlates of tick burdens in a New Guinea avifauna. In J.E. Loye & M. Zuk (Eds.), Ecology, Behaviour and Evolution of Bird–Parasite Interactions (pp. 154–176). Ames, IA: Oxford University Press.
- Richter , D. , Spielman , A. , Komar , N. and Matuschka , F.R. 2000 . Competence of american robins as reservoir hosts for Lyme disease spirochetes . Emerging Infectious Diseases , 6 : 133 – 138 .
- Rijpkema , S.G.T. , Molkenboer , M.J.C.H. , Schouls , L.M. , Jongejan , F. and Schellekens , J.F.P. 1995 . Simultaneous detection and genotyping of three genomic groups of Borrelia burgdorferi sensu lato in Dutch Ixodes ricinus ticks by characterization of the amplified intergenic spacer region between 5S and 23S rRNA genes . Journal of Clinical Microbiology , 33 : 3091 – 3095 .
- Saint Girons , I. , Gern , L. , Gray , J.S. , Guy , E.C. , Korenberg , E. , Nuttall , P.A. , Rijpkema , S.G.T. , Schönberg , A. , Stanek , G. and Postic , D. 1998 . Identification of Borrelia burgdorferi sensu lato species in Europe . Zentralblatt für Bakteriologie , 287 : 190 – 195 .
- Scott , J.D. , Fernando , K. , Banerjee , S.N. , Durden , L.A. , Byrne , S.K. , Banerjee , M. , Mann , R.B. and Morshed , M.G. 2001 . Birds disperse Ixodid (Acari: Ixodidae) and Borrelia burgdorferi-infected ticks in Canada . Journal of Medical Entomology , 38 : 493 – 500 .
- Siński , E. and Karbowiak , G. 1994 . Zarazenie Borrelia burgdorferi w wybranej populacji drobnych ssaków w pólnocnej Polsce . Biuletyn Instytutu Medycyny Morskiej i Tropikalnej , 27 : 70
- Siuda, K. (1993). Kleszcze Polski (Acari: Ixodida). Czesc II. Systematyka i rozmieszczenie 1st edn. Warszawa: Polskie Towarzystwo Parazytologiczne.
- Slowik , T.J. and Lane , R.S. 2001 . Birds and their ticks in northwestern California: minimal contribution to Borrelia burgdorferi enzootiology . Journal of Parasitology , 87 : 755 – 761 .
- Smith , M. , Gettinby , G. , Granström , M. , Gray , J.S. , Guy , E.C. , Revie , C. , Robertson , J.N. and Stanek , G. 1998 . The European Union Concerted Action World Wide Web Site for Lyme Borreliosis . Zentralblatt für Bakteriologie , 287 : 266 – 269 .
- Smith , R.P., Jr. , Rand , P.W. , Lacombe , E.H. , Morris , S.R. , Holmes , D.W. and Caporale , D.A. 1996 . Role of bird migration in the long-distance dispersal of Ixodes dammini, the vector of Lyme disease . Journal of Infectious Diseases , 174 : 221 – 224 .
- Stańczak , J.J. , Brojer , E. , Radlińska , M. , Medyńska , J. and Seyfried , H. 1995 . Partial nucleotide sequences and genotypes of hepatitis C virus (HCV) isolated in Polish blood donors and patients with hepatitis . Hepatologia Polska , 2 : 87 – 92 .
- Steere , A.C. 1989 . Lyme disease . New England Journal of Medicine , 321 : 586 – 596 .
- Strle , F. , Cheng , Y. , Nelson , J.A. , Picken , M.M. , Bouseman , J.K. and Picken , R.N. 1995 . Infection rate of Ixodes ricinus ticks with Borrelia afzelii, Borrelia garinii, and Borrelia burgdorferi sensu stricto in Slovenia . European Journal of Clinical Microbiology and Infectious Diseases , 14 : 994 – 1001 .
- Svensson L. Identification Guide to European Passerines 4th edn Lars Svensson, Sweden: Stockholm 1992
- Tälleklint , L. and Jaenson , T.G.T. 1994 . Transmission of Borrelia burgdorferi s.l. from mammal reservoirs to the primary vector of Lyme borreliosis, Ixodes ricinus (Acari: Ixodidae), in Sweden . Journal of Medical Entomology , 31 : 880 – 886 .
- Wegner, Z., Stańczak, J., Kubica-Biernat, B., Racewicz, M. & Kruminis-Lozowska, W. (1995). Some aspects of ecology of Ixodes ricinus and Borrelia burgdorferi in North-Eastern Poland. Programme and Abstracts IX European Meeting of Society For Vector Ecology (p. 46). Prague, Czech Republic.
- Valsangiacomo , C. , Balmelli , T. and Piffaretti , J-C. 1996 . A nested polymerase chain reaction for the detection of Borrelia burgdorferi sensu lato based on a multiple sequence analysis of the hbb gene . FEMS Microbiology Letters , 136 : 25 – 29 .
- van Dam , A.P. , Kuiper , H. , Vos , K. , Widjojokusumo , A. , de Jongh , B.M. , Spanjaard , L. , Ramselaar , A.C.P. , Kramer , M.D. and Dankert , J. 1993 . Different genospecies of Borrelia burgdorferi are associated with distinct clinical manifestations of Lyme borreliosis . Clinical Infectious Diseases , 17 : 708 – 717 .