Abstract
Sub-lingual oral fistulas are a consistently observed lesion affecting the New Zealand stitchbird (hihi: Notiomystis cincta). This lesion, which has not been reported in other species, is usually only recognized when the tongue protrudes below the bird's mandible from a hole in the oral-cavity floor. In this study, we surveyed the prevalence of oral fistulas in a free-living population of stitchbirds on Tiritiri Matangi Island in 2002, 2003 and 2005. Between surveys, individuals with a fistula were caught and the progress of their lesion was monitored. The majority of birds with a fistula had a small localized lesion alongside the edge of the mandible without the tongue protruding. Oral fistulas were generally not associated with any reduction in the bird's condition or productivity, but if the tongue consistently deviated through the fistula it affected nectar-feeding efficiency. No fistulas were found in nestlings, but 9% to 10% of adult birds had some form of oral fistula, suggesting that it developed after fledging. Repeated measurement of birds showed that the size of the fistulas did not progress beyond the formation of the initial hole unless the tongue protruded. This protrusion resulted in continuous rubbing and erosion of the oral cavity floor and, ultimately, the mandible itself. Histopathology confirmed that fistulas occur in the thinnest part of the floor of the oral cavity, at the attachment point of the skin to the mandible. Despite long-term monitoring of this population, the formation of an oral fistula has never been observed and its aetiology remains elusive.
Fistules orales sublinguales chez des méliphages hihi (Notiomystis cincta) vivant en liberté
Des fistules orales sublinguales sont des lésions fréquemment observées affectant les méliphages hihi (Notiomystis cincta) en Nouvelle Zélande. Ces lésions qui n'ont pas été rapportées chez d'autres espèces, ne deviennent généralement visibles que quand la langue dépasse sous la mandibule de l'oiseau à partir d'un trou dans le plancher de la cavité orale. Dans cette étude, nous avons enquêté sur la prévalence des fistules orales dans une population de méliphages hihi vivant en liberté sur l’île de Tiritiri Matangi en 2002, 2003 et 2005. Entre les enquêtes, des individus, avec une fistule, ont été capturés et l’évolution de leur lésion a été étudiée. La majorité des oiseaux avec une fistule présentaient une petite lésion localisée le long du bord de la mandibule sans que la langue dépasse. Les fistules orales n’étaient généralement pas associées à une diminution de l’état de l'oiseau ou de sa productivité, mais si la langue déviait en permanence à travers la fistule cela affectait l'efficacité de son alimentation en nectar. Aucune fistule n'a été trouvée chez les oisillons, mais 9–10% des oiseaux adultes présentaient une fistule orale d'une forme ou d'une autre, suggérant qu'elle s’était développée après le jeune âge. Des mesures répétées des oiseaux ont montré que la taille des fistules ne progressait pas au-delà de la formation d'un trou initial, sauf quand la langue dépassait. Ce phénomène est le résultat du frottement et de l’érosion du plancher de la cavité orale et, ultérieurement, de la mandibule elle-même. L'histopathologie a confirmé que les fistules apparaissaient dans la partie la plus fine du plancher de la cavité orale, au point d'attachement de la peau à la mandibule. Malgré l’étude à long terme de cette population, la formation d'une fistule orale n'a jamais été observée et son étiologie reste indéfinie.
Sublinguale Fisteln in der Schnabelhöhle von wildlebenden Gelbbandhonigessern (Notiomystis cincta)
Bei den Neuseeland-Gelbbandhonigessern oder Hihi (Notiomystis cincta) sind sublinguale Fisteln in der Schnabelhöhle häufig zu beobachtende Veränderungen. Diese Läsion, die bei anderen Vogelarten nicht beschrieben ist, wird gewöhnlich nur erkannt, wenn die Zunge unterhalb der Mandibel des Vogels durch ein Loch im Boden der Schnabelhöhle herausragt. In dieser Studie untersuchten wir in den Jahren 2002, 2003 und 2005 das Vorkommen der oralen Fisteln in einer wildlebenden Population von Gelbbandhonigessern auf der Insel Tiritiri Matangi. Zwischen den Reihenuntersuchungen wurden Einzeltiere mit Fistel eingefangen und der Fortschritt der Veränderungen protokolliert. Die Mehrzahl der Vögel mit einer Fistel hatten eine kleine umschriebene Läsion neben der Unterkieferkante, ohne dass die Zunge vorfiel. Die oralen Fisteln waren im Allgemeinen nicht mit einer Einschränkung des Allgemeinbefindens oder der Produktivität des Vogels verbunden, aber wenn die Zunge ständig durch die Fistel abgelenkt wurde, war die Nektaraufnahme behindert. Bei Nestlingen wurden keine Fisteln gefunden, aber 9–10% der adulten Vögel wiesen unterschiedliche Stadien der Oralfisteln auf, was vermuten ließ, dass sie sich erst nach dem Flüggewerden entwickeln. Wiederholte Untersuchungen der Vögel ließen erkennen, dass die Größe der Fisteln nicht über die Bildung eines anfängliche Lochs hinausging, sofern die Zunge nicht herausgestreckt werden konnte. Dieses Herausstrecken führte zu fortschreitendem Abrieb und Aushöhlung des Schnabelhöhlenbodens und letztendlich auch des Unterkiefers selber. Histopathologisch konnte bestätigt werden, dass sich die Fisteln an der dünnsten Stelle des Schnabelhöhlenbodens, nämlich der Anheftungsstelle der Haut an den Unterkiefer ausbildeten. Trotz der Langzeitstudie an dieser Population konnte die Entstehung der oralen Fistel kein einziges Mal beobachtet werden und ihre Ätiologie bleibt somit unklar.
Fístulas sublinguales en Pájaros Puntados (Notiomystis cincta)
Las fístulas orales sublinguales son una lesión observada frecuentemente en Pájaros Puntados de Nueva Zelanda (Notiomystis cincta). Esta lesión, que no se ha descrito en otras especies, generalmente sólo se reconoce cuando la lengua sobresale de un orificio en el suelo de la cavidad oral por debajo de la mandíbula del ave. En este estudio, investigamos la prevalencia de las fístulas orales en la población de Pájaros Puntados salvajes en la isla de Tiritiri Matangi en los años 2002, 2003 y 2005. Entre cada estudio se capturaban los individuos con fístula y se monitorizaba la progresión de esta lesión. La mayoría de aves con fístula tenían una pequeña lesión a lo largo del borde de la mandíbula sin protusión de la lengua. Generalmente, la presencia de fístulas orales no se asociaba a reducción de la condición corporal del ave o de su productividad, pero si la lengua se desviaba de manera importante a través de la fístula afectaba la eficiencia en la ingestión de néctar. No se observaron fístulas en los pollitos jóvenes, pero entre un 9–10% de las aves adultas tenían alguna forma de fístula oral, lo cual sugiere que ésta se desarrolla después del emplume. Medidas repetidas en las aves mostraron que el tamaño de las fístulas no progresaba más allá de la formación inicial del orificio si no se producía protusión lingual. Esta protusión producía un roce contínuo y erosión del suelo de la cavidad oral, y finalmente, incluso de la mandíbula. La histopatología confirmó que las fístulas ocurrían en la parte más fina del suelo de la cavidad oral, en el punto de unión entre la piel y la mandíbula. A pesar del largo período de monitorización de esta población, nunca se ha observado la formación de una fístula oral y su etiología sigue siendo desconocida.
Introduction
The New Zealand stitchbird or hihi (Notiomystis cincta) is a range-restricted endangered passerine for which past conservation efforts have primarily focussed on establishing new populations via translocation (Higgins et al., Citation2001). Currently, there are four free-living populations: the source population on Little Barrier Island and three small translocated populations—Kapiti Island, Tiritiri Matangi Island and the Karori Wildlife Sanctuary in Wellington. Previous studies have highlighted the stitchbird's susceptibility to aspergillosis and atoxoplasmosis, diseases that cause significant morbidity and mortality in both wild and captive populations (Alley et al., Citation1999; Cork et al., Citation1999). Because translocation provides a means by which disease agents could be transmitted between populations (Viggers et al., Citation1993), there is a requirement in New Zealand that diseases at the source and release locations be identified and their potential impacts assessed (Jakob-Hoff et al., Citation2004). While much is understood about the risks associated with aspergillosis and atoxoplasmosis, there is one abnormality that has been widely observed in stitchbirds and yet almost nothing is known about its cause, prevalence, or effects on survival: sub-lingual oral fistulas.
Sub-lingual oral fistulas have been observed in stitchbirds since the early 1990s and were first described by Castro & Taylor (Citation2001). In their five reported cases, Castro and Taylor observed each bird's tongue hanging out of a large hole in the tissue comprising the floor of the oral cavity. Observations of similar tongue protrusions have now been reported in all recent translocated stitchbird populations—Mokoia Island, Kapiti Island, Tiritiri Matangi Island, Mt Bruce National Wildlife Centre (Castro & Taylor, Citation2001; Price, Citation2006; R. Collen, personal communication)—as well as the natural population on Little Barrier Island (J. Ewen, personal communication). Despite the apparent ubiquity of this pathology in stitchbird populations, to our knowledge it has not been observed in other New Zealand species, nor has it been reported in birds from other parts of the world. While Castro & Taylor (Citation2001) used the term “bill abnormality” as a label for the condition, it is not clear to what extent the bill is involved; because of this, we have adopted the term “sub-lingual oral fistula” (henceforth “oral fistula”; see also Castro & Taylor, Citation2001) to describe any lesion in the floor of the oral cavity that creates an abnormal communication between the oral cavity and the external surface of the lower jaw.
Hypotheses proposed by Castro & Taylor (Citation2001) for the existence of these oral fistulas include: genetic abnormality, possibly associated with small founder populations on islands where stitchbird translocations have taken place; nutritional deficiencies; and injury, either associated with their acrobatic feeding behaviour or aggression during the breeding season. A long-term study of the Tiritiri Matangi Island stitchbird population provided an opportunity to evaluate predictions arising from these hypotheses and to collect data in order to better understand the prevalence, impact and progression of the abnormality over a period of several years.
Materials and Methods
Study population
Birds in this study were observed during four breeding seasons between 2001/02 and 2004/05 on Tiritiri Matangi Island (36°36′ S, 174°53′ E), a scientific reserve located off the northeast coast of New Zealand's North Island. The island is approximately 220 hectares in area, with stitchbirds restricted to remnant and regenerating closed-canopy forest patches totalling around 30 hectares. All birds on the island were uniquely colour banded, with their ages and social parentage known. Stitchbirds on Tiritiri Matangi Island breed during the spring and summer (September to February) and may successfully raise up to two broods of between one and five chicks each season. Stitchbirds were translocated to the island in 1995 as part of the ongoing management of the species by the New Zealand Department of Conservation where they now form a closed population. The population is small but expanding (32 breeding females in 2001/02, increasing to 53 breeding females in 2004/05), allowing all breeding attempts to be monitored. Stitchbirds feed on fruits, insects and nectar (Wilson, Citation1997), and have a specialized tongue for extracting nectar from flowers (Higgins et al., Citation2001). Supplementary food in the form of a 20% (w/v) sugar solution was provided year round at nine feeding stations and used by all birds on the island. These feeding stations were necessary because of a shortage of natural food, and were situated at the forest edges and not contained within birds’ territories. Stitchbirds usually require tree cavities for successful nesting; however, these are not readily available on the island as the vegetation is predominantly young regenerating forest. Hence, small groups of two or three nest boxes were placed throughout potential nesting areas (78 in 2000, increased to 86 in 2001). Each nest box was attached to a tree trunk approximately 1.5 metres off the ground and had a hinged lid that allowed easy monitoring of nesting.
Capture and measurement
During three periods of the study (February 2002, June 2003, February to April 2005), a large proportion of the stitchbird population on the island was caught and examined for the presence of oral fistulas. In 2002, this represented 100% of the adults present; in 2003, 96% of all stitchbirds present; and in 2005, approximately 60% of the birds were caught and examined. Specific birds were not targeted for capture during these periods, and thus the number of oral fistulas described is likely to represent their true prevalence in this population. Birds known to have oral fistulas were targeted for capture at other times to monitor their fistulas and note any changes; these capture periods coincided with captures for other studies (for example, Low et al., Citation2005).
Stitchbirds were caught as they came to feed at supplementary feeding stations by quickly closing the door to the feeder and then extracting the bird by hand. The ability to target specific birds in this way prevented the same bird being caught more than once during each of the capture periods. Upon capture, all birds were weighed to within 0.5 g using a Pesola spring balance and had their body condition checked by palpation of the keel bone. During the 2002/03 breeding season, birds were weighed on a daily basis using a set of electronic scales (Weighing Systems Ltd., Nelson, New Zealand) to an accuracy of ±0.5 g (see Low, Citation2004). The scales were attached to a hummingbird feeder containing artificial nectar so that when birds came to drink they stood on a perch linked to the weighing mechanism. An electronic readout of the bird's weight was displayed, and this was recorded the first time each bird came to the feeder.
Observations of oral fistulas
A free-ranging bird was recorded as suffering from an oral fistula if it was observed with its tongue protruding from a hole in its lower bill (the criterion used by Castro & Taylor, Citation2001). In addition to this, we closely examined birds at capture that were not displaying any detectable tongue protrusion, and, thus, would otherwise be presumed to be “normal”. These birds were held on their backs and the floor of the lower bill was examined in the following way: a smooth-ended probe was used to lift feathers originating from the midline ventral feather tract, the line of feathers that extends from the tip of the mandible to the midline of the ventral neck, to expose the underlying skin; the skin was then examined on both the left-hand and right-hand sides of the ventral feather tract, using the probe to stretch any skin folds that might be hiding lesions; if a fistula was detected, its location was recorded and its size determined to ±0.05 mm using vernier callipers, with any lesions associated with the beak also noted; and, finally, if the tongue was protruding from the hole, the beak was opened and the probe was used to thread the tongue back into its proper position—exiting the oral cavity between the tips of the maxilla and mandible. From these data, birds were then assigned into one of five categories ( and ).
Figure 1. Ventral views of stitchbirds with oral fistulas (OF). 1a: A 4-month-old female with a small stage 1 OF. 1b: A male with a stage 2 OF; photograph taken immediately after the tongue had been replaced through the fistula. In both cases the OF was restricted to one side of the midline ventral feather tract. 1c: The tongue was permanently protruding through the fistula (stage 3 OF), and its action had eroded the ventral feather tract and the fistula incorporated the entire floor of the oral cavity. 1d: The same bird 2 years later with a stage 4 OF; the action of the tongue had eroded a significant piece (12 mm) of the rostral left mandible (LM indicates the rostral edge of the remaining left mandible).
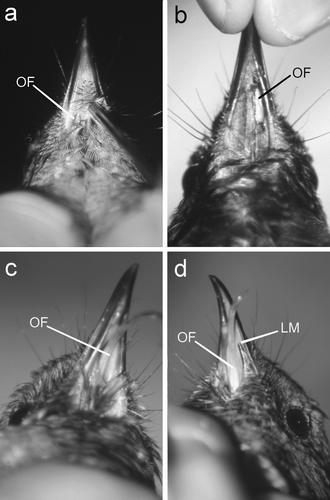
Table 1. Categories for classifying oral fistulas
Histological examination
Histological sections were prepared from the lower beaks of three male stitchbirds that died from other causes and were not suffering from any fistulas or mandibular abnormalities. Particular attention was paid to the area of skin and oral mucosa adjoining the mandible where fistulas appear to form. Histological sections were also prepared from the oral cavity of an adult female stitchbird from the Mt Bruce National Wildlife Centre that had a chronic oral fistula incorporating the entire floor of the oral cavity, as well as the lower beak and pharynx of a juvenile male stitchbird from Kapiti Island that had died from complications arising from a 6×6 mm2 abscess in the floor of the oral cavity 12 mm caudal to the symphysis of the mandible. All birds had been preserved in 10% buffered formalin soon after death. For the three males with no oral lesions, the mandible was removed and a transverse section of the lower bill was cut 15 mm from the anterior tip of the mandible; this is the approximate location of most oral fistulas. For the female with the oral fistula and the male with the oral abscess, a sagittal section of the head and beak was cut to provide a good cross-section of the point where the oral fistula or abscess began. All tissues were cut at 4 µm and stained with haematoxylin and eosin.
Data analysis
Data were not normally distributed; non-parametric statistics were therefore used for statistical comparisons. For the comparison of oral fistula prevalence, a mean was derived from the three studies for each age classes. Means are displayed±1 standard error unless otherwise indicated.
Results
During the study, 11 birds were discovered to have oral fistulas: five females and six males. Two young birds exhibited tongue protrusion through a hole in the lower beak, but these birds could not be caught for examination. Of the remaining nine birds, five had a small fistula along the medial edge of the left mandible, two along the medial edge of the right mandible, and two had lost the floor of their oral cavity at the time of first examination (). With one exception, there did not appear to be any active inflammation or infection associated with the fistulas, with the edges of the holes appearing to have completely re-epithelialized. In the one bird with some minor lesions at the fistula, this was associated with rubbing of the tongue on the tissue of the ventral feather tract. Rubbing occurred because of the angle at which the tongue exited the fistula.
Prevalence of oral fistulas
During the 4 years of this study, the prevalence of oral fistulas in the adult population remained relatively stable at between 9% and 10% (). While there were higher proportions of adult males with oral fistulas in all surveys when compared with females (), this difference was not significant (Fisher's exact test, P > 0.1). Juveniles were significantly less likely to be found with an oral fistula during these surveys when compared with adults (1 out of 71 versus 10 out of 111; χ2=4.4, P<0.05). In 2001/02, of the 103 chicks banded at 21 days of age, none had oral fistulas at the time of examination. This was also the case for the 67 dead chicks that were recovered from inside and outside of nests. The lack of any detectable lesions between hatching and fledging was significantly different to that seen in adults (0 out of 170 versus 10 out of 111; χ2=15.8, P<0.001).
Table 2. Prevalence of oral fistulas in adults and juvenilesa during the three population surveys
Of the 11 birds with oral fistulas, six came from first clutch nests and five from second clutches. For 10 of the birds whose parents where still alive and were able to be caught and examined, none were ever recorded as suffering from an oral fistula during the period of the study. However, two birds (“bm/rg” and “bm/ry”) were siblings from the same nest ().
Table 3. Oral fistula scores (range from stage 0 to stage 4) and dimensions (mm) for the 11 birds in this study relative to their age at examination
Longitudinal changes
During the study, seven of the birds with an oral-fistula were caught more than once, and this allowed the progression of their lesions to be monitored over time (). Three of these birds were first examined at a young age (<2 months old) when they did not have an oral fistula. An additional four birds were first caught when their fistulas were small and not associated with any tongue protrusion. Two of these birds did not have any change in the appearance of their lesion for several years, while the other two enlarged slightly and this coincided with a stage 2 tongue protrusion (). In no cases did the fistula ever become smaller or heal over. Four of the 11 birds did not display tongue protrusion at any time.
Examinations of birds with tongue protrusions provided evidence for the mechanism by which the fistulas progress from stage 1 to stage 4. Birds with no reported tongue protrusion during the study had no measurable changes in their fistulas. Birds in stage 2 or 3 had wider and longer lesions; this change appeared to be because the tongue provided constant pressure and rubbing on the anterior and medial surfaces of the hole. For the birds with long-term tongue protrusion, the beak fractured along a weak point created by the action of the tongue on the underside of the mandible. In the female “ym/wy”, this continued until 12 mm of the left anterior mandible had been eroded ().
Impacts on foraging and productivity
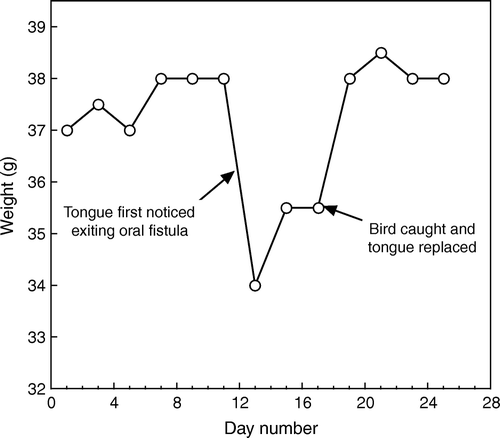
The presence of an oral fistula did not have any measurable effect on the birds’ body condition at the end of the breeding season (February 2002; condition score 3.8±0.1, n=79 versus 3.7±0.1, n=3), immediately post-moult (April 2005; condition score 3.8±0.1, n=101 versus 4.0±0.0, n=7), or during the winter (June 2003; condition score 3.6±0.1, n=50 versus 3.4±0.2, n=5) (Mann-Whitney U tests, all P>0.1). All birds with oral fistulas were observed feeding on fruits, nectar and invertebrates. However, two birds with tongue protrusions had markedly different supplementary feeder usage when compared with other birds in 2002/03. The female “ym/wy” (stage 4) spent more than 50% of her time within 5 m of the supplementary feeding station during October and November 2002, in contrast to an average of 6% for other birds. In 2005, this female was caught for examination and her faeces contained many Coprosma robusta seeds showing that she was capable of taking fruit. The male “bg/wm” changed his feeding pattern when he progressed from stage 1 to stage 2 (the stage 1 fistula was unobserved, the first sign that he was suffering from an oral fistula was when his tongue protruded; see ); once his tongue protruded, the time he spent at the feeder during each visit increased threefold. During this same period his weight dropped markedly and did not recover until he was caught and his tongue was replaced ().
Figure 2. Weight fluctuations of male “bg/wm” relative to a tongue protrusion.
For the six pairs in 2001/02 where one member had a stage 1 fistula, there were no differences between their productivity and that of the general population—first clutches: eggs laid, 4±0.4 versus 4.02±0.1; chicks fledged, 2.3±0.9 versus 2.2±0.3; chick weight at 21 days, 41.5±1.4 g versus 41.5±0.5 g; second clutches: eggs laid, 4±0.4 versus 4.2±0.2; chicks fledged, 1.2±0.7 versus 1.3±0.3; chick weight at 21 days, 40.8±0.3 g versus 40.3±1.0 g (Mann-Whitney U test, all P>0.5). For the two females with stage 3/4 oral fistulas, very poor breeding success was observed (3.2±0.3 eggs laid per clutch (n=5) with no chicks successfully hatched); however, this was partly confounded by their late nesting and being the secondary female to a polygynous male (see Low et al., Citation2006).
Histology
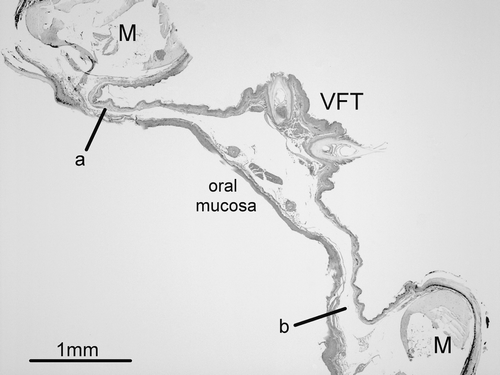
Histological examination of the mandibles of the three males not afflicted by fistulas confirmed that the area where the fistulas occur corresponds to where the floor of the oral cavity is thinnest (as little as 0.2 mm) (). These thin attachment points to the mandible are in contrast to the mid-way point between the two bones of the mandible where the tissue is significantly thicker (>1 mm) as it provides the anchor point for the feathers and bristles of the ventral feather tract.
Figure 3. Photomicrograph of a haematoxylin and eosin-stained transverse section of the lower jaw of a male stitchbird. The left and right mandibular bones (M) are joined by a thin section of tissue that comprises the floor of the oral cavity. The ventral feather tract (VFT) runs along the midline and here the tissue is significantly thicker than at the margins of the mandible where oral fistulas develop (a and b).
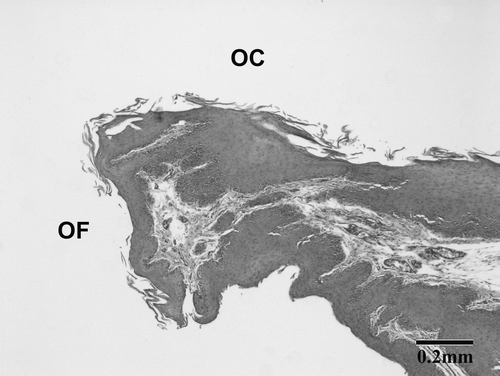
For the female stitchbird with the chronic oral fistula, the ventral skin beneath the oral cavity was continuous with the ventral oral mucosa at the caudal margin of the oral fistula (). Approximately 5 mm caudal to the fistula opening, the epidermis of the throat became acanthotic (from two to four cells in thickness to 10 to 20 cells) and showed extensive hyperkeratosis. This thickening continued into the oral mucosa where the ventral oral epithelium was approximately 30 cells in thickness (normal thickness ∼10 cells). The submucosa showed no significant changes. As the ventral oral mucosa extended towards the mid-line, it increased further in thickness and formed extended irregular epithelial pegs; a few lymphocytes had accumulated around the submucosal blood vessels in this area. A section near the mid-line showed a small submucosal cyst (approximately 0.5 mm diameter) present in the ventral oral mucosa, 3 mm caudal to the fistula opening. No inflammatory changes were present in the submucosal tissues.
Figure 4. Photomicrograph of a haematoxylin and eosin-stained sagittal section of the oral cavity (OC) of a female stitchbird with an oral fistula. OF indicates the caudal edge of the oral fistula and the transition from the skin to the ventral oral mucosa.
The ventral pharyngeal region of the juvenile male contained a large (6 mm diameter) abscess that extended from the base of the tongue into the left lower mandible and sublingual salivary glands. It consisted of a central accumulation of necrotic exudate containing numerous colonies of bacteria (both Gram-positive and Gram-negative coccobacilli). This was surrounded by a thick zone of fibrous granulation tissue that extended into the lingual muscle and adjacent mandibular bone. The affected bone showed varying degrees of lysis and remodelling of the cortex, and periosteal proliferation. A more lateral section of the oral cavity revealed a large nodule of ulcerating granulation tissue attached to the left ventral oral mucosa. This had a small central area of ossification and cartilage formation.
Discussion
Until now, it has been assumed that sub-lingual oral fistulas in stitchbirds are manifest by a visible deviation of the bird's tongue through the hole in the lower bill (Castro & Taylor, Citation2001). From our study, it is apparent that birds with a tongue protrusion represent a minority of the birds affected by oral fistulas; thus, estimates based only on sightings of birds with tongue protrusions will be significantly lower than the true prevalence.
Despite the causal factors responsible for the development of oral fistulas in stitchbirds remaining elusive, data collected from this study have helped evaluate the three non-exclusive hypotheses suggested by Castro & Taylor (Citation2001). The first hypothesis proposes that reduced genetic variability due to inbreeding has fixed detrimental alleles within a limited gene pool; small founder populations at translocation sites were suggested as being a factor because of the observation that tongue protrusions had only been observed at translocation sites. However, the fact that a stitchbird with an oral fistula and tongue protrusion has recently been observed at the source population on Little Barrier Island and that all four of the intensively monitored translocation sites have reported oral fistulas, is highly suggestive that the prevalence of oral fistulas in the source population is similar to that in translocated populations. Chicks and juveniles are significantly less likely to exhibit oral fistulas when compared with adults, and this, combined with the data from two birds who developed their fistulas after 2 months of age, demonstrate that the condition is not congenital. Rather, it develops after birds leave the nest. While this does not necessarily mean that there is not a genetic component to the disease, it does suggest that if inbreeding plays a role, a second factor is probably needed to induce the pathology. This interpretation is further supported from the lack of any genealogical association between birds with oral fistulas in this study.
The second hypothesis is that translocation sites are nutritionally deficient and that oral fistulas are a clinical manifestation of a nutritional deficiency. With the observation of the condition in the Little Barrier population, a self-sustaining population in a diverse forest habitat, and at the Mt Bruce National Wildlife Centre, where the birds are given a nutritionally balanced diet, this explanation seems unlikely. Additionally, in none of the birds examined in this study were there any clinical signs indicating a nutritional deficiency, and no defects of this type are currently known to be caused by nutritional deficiencies (Ritchie et al., Citation1994).
The third hypothesis proposed by Castro & Taylor (Citation2001) was that oral fistulas were a result of an injury or infection associated with feeding or aggression during the breeding season. This still remains a possibility, but the mechanism by which this could occur is unclear. No adult birds were ever found to develop fistulas from one season to the next; the six birds that developed fistulas during the study were young, and the fistulas had most probably formed prior to their first breeding season. In other avian species, crop fistulas are known to occur more commonly in neonates presumably because the crop is more fragile and susceptible to injury than in adults (Ritchie et al., Citation1994). One possibility that has not been previously considered is a relationship between Knemidocoptes infections and the development of the fistulas (Low et al., Citation2007). However, there is currently no evidence that birds displaying mite lesions are any more likely to suffer from an oral fistula. Histopathology showed that the fistulas form in the area of the oral cavity where the tissue is at its thinnest and that, once formed, there is little active inflammation associated with them. The juvenile male from Kapiti with the abscess on the floor of the oral cavity suggests a possible aetiology for the fistulas. Necrosis associated with an abscess or insect sting may produce a fistula where the floor of the oral cavity is thin; however, this hypothesis currently has no supporting evidence.
While it is still not known how or why the fistulas form, this study provides a much clearer indication of when and where they occur as well as the progression of the disease over time. All fistulas were found to originate in the same place; that is, against the medial surface of the mandible (on the left or right), approximately mid-way along its length (). In a number of cases the fistula did not develop beyond this stage and did not appear to inconvenience the bird in any way. However, occasionally, the tongue managed to exit the oral cavity through the fistula rather than through the opening of the beak. If this occurred, the fistula began to grow in size due to the action of the tongue on the surrounding tissue. The result of this was that the fistula became so large that the tongue could not be held inside the beak and it permanently protruded through the fistula. Once this had occurred, a segment of the mandible was eroded through the continual movement of the tongue on the underside of the beak. The impact of this change in tongue direction, and presumably feeding efficiency, was dramatic (in the short term at least) for the male who progressed from a stage 1 to a stage 2 oral fistula during the 2002/03 breeding season. This was most probably because of his difficulty in ingesting nectar from the supplementary feeders. Despite this, there was little evidence that birds with oral fistulas or tongue protrusions could not maintain their body weight in the long term, at least in a habitat like Tiritiri Matangi Island where ad libitum supplementary food may help compensate for foraging inefficiencies associated with oral fistulas. Even with one-half of the left side of her mandible missing, the female stitchbird in this study with a stage 4 oral fistula was in good condition and managing to take supplementary nectar as well as fruit (as evidenced by the seeds in her faeces). However, there was some evidence in support of severely affected stage 4 birds having difficulty in successfully hatching and raising offspring. Despite this, evolutionary selection pressures against this disease may be weak, especially when one considers that most birds will breed a number of times before a tongue protrusion has any impact on their ability to forage and regurgitate food to offspring.
The current focus of conservation medicine is to limit the spread or impact of diseases in vulnerable animal populations (Deem et al., Citation2002). Currently there is no evidence to suggest that sub-lingual oral fistulas are the result of an infectious disease, and therefore birds with this condition do not need to be isolated. Also, the idea that this is a disease exacerbated by inbreeding at translocation sites is not supported by current evidence, and, thus, additional translocations of birds as recommended by Castro & Taylor (Citation2001) to improve genetic diversity and minimize the incidence of this lesion are currently not justified. Because a number of birds eventually do progress from a stage 1 fistula to one with a tongue protrusion, which might impact on their ability to forage for offspring, it would be wise to inspect the underside of the bill of any bird targeted for translocation and remove them from the group of birds to be moved. More work is still required to understand the pathogenesis of this abnormality in the stitchbird, and to answer the question of why it has not been reported in other species.
Acknowledgement
The authors thank Rose Collen, Greg Moorcroft and John Ewen for recovery of stitchbird carcasses, Troy Makan, Tamara Henry, Ian Price, Clare Miller, Kirsty Chalmers, Åsa Berggren, Sandra Jack, Su Sinclair and John Ewen for assistance in the field and reporting observations of birds, and Barbara Walter, Ray Walter, Ian Price, Rosalie Stamp and Richard Griffiths for logistical support. The expertise of Pat Davey and Evelyn Lupton in processing the histopathology was invaluable. They also thank the comments of two anonymous referees on the manuscript. This research was supported by the New Zealand Lotteries Commission, J.S. Watson Conservation Trust and the Supporters of Tiritiri Matangi Inc. All work was carried out under a research permit from the New Zealand Department of Conservation and had Massey University animal ethics approval.
References
- Alley , M.R. , Castro , I. and Hunter , J.E.B. 1999 . Aspergillosis in hihi (Notiomystis cincta) on Mokoia Island . New Zealand Veterinary Journal , 47 : 88 – 91 .
- Castro , I. and Taylor , J. 2001 . Survival and reproductive success of stitchbird (hihi, Notiomystis cincta) suffering from a bill abnormality (oral fistula) . Notornis , 48 : 241 – 244 .
- Cork , S.C. , Alley , M.R. , Johnstone , A.C. and Stockdale , P.H. 1999 . Aspergillosis and other causes of mortality in the stitchbird in New Zealand . Journal of Wildlife Diseases , 35 : 481 – 486 .
- Deem , S.L. , Kilbourn , A.M. , Wolfe , N.D. , Cook , R.A. and Karesh , W.B. 2002 . Conservation medicine . Annals of the New York Academy of Sciences , 916 : 370 – 377 .
- Higgins , P.J. , Peter , J.M. and Steele , WK. 2001 . “ Stitchbird ” . In Handbook of Australian, New Zealand and Antarctic Birds , Edited by: Higgins , P.J. , Peter , J.M. and Steele , W.K. Vol. 5 , 954 – 966 . Melbourne : Oxford University Press .
- Jakob-Hoff , R. , McInnes , K. , Frank , M . & Cromarty P . ( 2004 ). Translocation and quarantine health workbook. Unpublished report , New Zealand Department of Conservation , Wellington , , Lake Worth, FL .
- Low , M. 2004 . Female weight predicts the timing of forced copulation attempts in stitchbirds, Notiomystis cincta . Animal Behaviour , 68 : 637 – 644 .
- Low , M. , Castro , I. and Berggren , Å. 2005 . Cloacal erection promotes vent apposition during forced copulation in the New Zealand stitchbird . Behavioural Ecology and Sociobiology , 58 : 247 – 255 .
- Low , M. , Joy , M.K. and Makan , T. 2006 . Using regression trees to predict patterns of male provisioning in the stitchbird (hihi) . Animal Behaviour , 71 : 1057 – 1068 .
- Low , M. , Alley , M.R. & Scott , I. ( 2007 ). Pruritic facial dermatitis in a population of free-living stitchbirds . Journal of Wildlife Diseases , In press .
- Price , I. ( 2006 ). Kapiti Island hihi (stitchbird) monitoring report. Unpublished report , New Zealand Department of Conservation , Wellington .
- Ritchie , B.W. , Harrison , G.J. and Harrison , L.R. 1994 . Avian Medicine: Principles and Application , FL : Wingers .
- Viggers , K.L. , Lindenmayer , D.B. and Spratt , D.M. 1993 . The importance of disease in reintroduction programmes . Wildlife Research , 20 : 687 – 698 .
- Wilson , L.R. ( 1997 ). The ecology and management of honeyeaters in Northern New Zealand . M.Sc. thesis , University of Auckland , New Zealand .