Abstract
Serratognathus diversus An, Cornuodus longibasis (Lindström), Drepanodus arcuatus Pander, and eleven other less common conodonts, including Cornuodus? sp., Oistodus lanceolatus, Protopanderodus gradatus, Protoprioniodus simplicissimus, Juanognathus variabilis, Nasusgnathus dolonus, Paltodus? sp., Scolopodus houlianzhaiensis, Semiacontiodus apterus, Semiacontiodus sp. cf. S. cornuformis and Serratognathoides? sp., are described and illustrated from the Honghuayuan Formation in Guizhou, South China, concluding revision of the conodont fauna from this unit, which comprises 24 species in total. The most distinctive species in the fauna, S. diversus, consists of a trimembrate apparatus, including symmetrical Sa, asymmetrical Sb and strongly asymmetrical Sc elements. This species concept is supported by the absence of any other element types in a large collection represented by nearly 500 specimens of this species. The fauna indicates a late Tremadocian to mid-Floian age (Early Ordovician) for the Honghuayuan Formation, which was widely distributed on the Yangtze Platform in shallow water environments. Previously published biostratigraphic zonations for the Honghuayuan Formation are reviewed, and revised on the basis of our knowledge of the entire conodont fauna, supporting the establishment of three biozones, Triangulodus bifidus, Serratognathus diversus, and Prioniodus honghuayanensis biozones in ascending order. Species of Serratognathus enable correlation between Ordovician successions of South China, North China (North China Platform and Ordos Basin), Tarim Basin, and further afield into Malaysia and northwestern Australia.

CONODONTS of the Honghuayuan (Hunghuayuan) Formation were collected and studied from the type Honghuayuan Section and two other sections (at Ganxi and Huanghuachong) in Guizhou Province, South China, as part of an international cooperation project, ongoing since 2000, involving palaeontologists from Australia, Peking University and the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences. The present contribution is the final report of the project following five earlier publications on the fauna from the Honghuayuan Formation (Zhen et al. Citation2005, Citation2006a, Citation2006b, Citation2007b, Zhen Citation2007), with emphasis on conodont biostratigraphy and regional correlation of the fauna. The Honghuayuan fauna, represented by 24 conodont species in total, 14 of which are documented herein, is one of the best-preserved and most diverse Early Ordovician faunas in South China. Of particular importance to correlations with North China, Malaysia and Australia, are species of the distinctive genus Serratognathus, one of which (S. diversus) ranges through the middle and upper part of the Honghuayuan Formation in sufficient numbers to permit its reconstruction as a trimembrate apparatus.
Geological setting
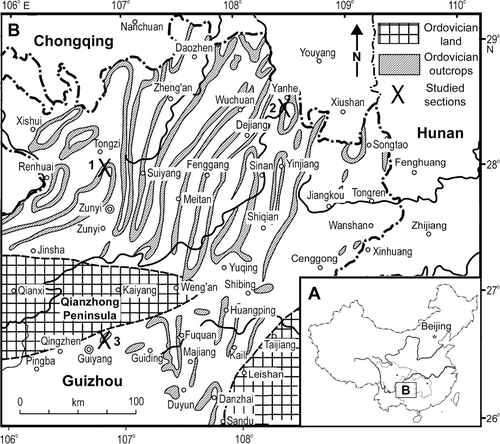
During the Ordovician, South China, as an independent plate in close association with eastern Gondwana, comprised three sedimentary-structural units. From northwest to southeast, these were the carbonate-dominated Yangtze Platform succession with a thickness of 300 to 500 m (Zhang & Zhen Citation1991), a transitional belt occupying the Jiangnan Slope, and the deep-water Zhujiang Basin with turbidite successions up to 7000 m thick. The Honghuayuan Formation, consisting of shallow inner-shelf carbonates dominated by thick-bedded, bioclastic limestones with local development of biostromes and bioherms or patch reefs (Zhu et al. Citation1995, Li et al. Citation2004), is one of the most widely distributed Ordovician carbonate units on the Yangtze Platform (Chen Xu et al. Citation1995). The formation is best developed in Guizhou Province. At its type locality near Honghuayuan village, 7 km south of Tongzi County town (), it consists of mainly medium- to thick-bedded limestone 32 m thick. In north and northeast Guizhou, the formation has a thickness of 20–50 m. It is generally thinner (20–30 m) in the area adjacent to the ‘Qianzhong Peninsula’ (an easterly extension of the Dianqian Land), but thickens to more than 130 m northeastwards. The Honghuayuan Formation also extends eastwards from Guizhou–Sichuan through Hunan–Hubei to the Lower Yangtze Valley (Anhui and Jiangsu), and thins southwards into siliciclastic successions of the outer shelf-slope facies in the Jiangnan area (An Citation1987, Zhou et al. Citation2007, Zhan & Jin Citation2007).
Fig. 1 Locality maps of the studied areas. A, Map of China showing the location of Guizhou Province. B, Map of northeast Guizhou showing the locations of three sections sampled for this study (Geological information modified from Gao, Citation1976): (1) Honghuayuan Section of Tongzi; (2) Ganxi Section of Yanhe; (3) Huanghuachong section of Guiyang in Guizhou.
The top of the Honghuayuan Formation in Guizhou is diachronous, and grades lithologically into the overlying graptolitic shale-dominated Meitan Formation. Here the base of the Honghuayuan Formation can be recognized by a shale marker bed at the top of the underlying Tongzi (Tungtzu) Formation, which is composed predominantly of dolomite and dolomitic limestone intercalated with shale and siltstone ().
Conodont biostratigraphy
An (Citation1981) and An et al. (Citation1981) established the Serratognathus Zone, based on a fauna consisting of 12 conodont species from the Honghuayuan Formation in the Huanghuachang area of Yichang, Hubei Province. They correlated it with similar faunas from the Liangjiashan (Liangchiashan) Formation of North China and from Korea, and indicated that the Serratognathus Zone might be divided into an upper subzone correlating with the Prioniodus elegans Zone and a lower subzone correlating with the Paroistodus proteus Zone of the Baltic successions.
Based on later more comprehensive studies of conodonts from South China, An (Citation1987, pp. 70–71) established the Serratognathus diversus Zone, which was further subdivided into lower and upper subzones, within the Honghuayuan Formation. The lower boundary of this Zone was defined by the first appearance (FAD) of S. diversus, with its upper boundary marked by the first appearance of Oepikodus communis or Prioniodus elegans. An (Citation1987, table 2) correlated the S. diversus Zone with the Paroistodus proteus Zone of the Baltic successions, implying a slightly narrower age span than the original concept of An et al. (Citation1981) and An (Citation1981). Subsequently, the widely distributed S. diversus Zone fauna has provided the basis for biostratigraphic correlation of the Honghuayuan Formation throughout the Yangtze Platform, and also with other coeval units in North China (An et al. Citation1983, An & Zheng Citation1990) and the Tarim Basin (Zhao et al. Citation2000). However, precise regional correlations remained uncertain to some degree (Zhen Citation2007), as S. diversus typically occurs only in the middle and upper part of the Honghuayuan Formation at its type locality and is absent from the lower and topmost parts. As both the lower and upper boundaries of the formation are regionally diachronous, the stratigraphic distribution of S. diversus may vary from place to place (e.g. in Hubei, the first appearance of S. diversus is in the underlying Fenxiang Formation). Furthermore, the ranges of O. communis and P. elegans are not entirely coeval and these species likely lived in different environments; the former occurring typically in the North American mid-continent successions (Tropical Domain of the Shallow-Sea Realm) and the latter in the Balto-Scandian successions (Cold Domain of the Shallow-Sea Realm) or in deep water successions such as the Cow Head Group of Newfoundland (Open-Sea Realm). Confirmed P. elegans specimens are rare in the Lower Ordovician of South China but Prioniodus honghuayuanensis Zhen in Zhen et al., Citation2005 occurs abundantly at the top of the Honghuayuan Formation [slightly lower stratigraphically than O. communis, to which it was assigned by An (Citation1987) as B. communis—see discussion by Zhen et al. Citation2005, Citation2007a, and is closely related morphologically to P. elegans.
Zeng et al. (Citation1983) proposed the Triangulodus bicostatus Zone to represent the conodont fauna from the Honghuayuan Formation in the Yangtze Gorges area of Hubei Province, and suggested a ‘mid-early Arenig’ age for the formation. As S. diversus is more abundant and distinctive in the fauna and is confined to only the Honghuayuan Formation, this latter proposal has been rejected by subsequent conodont workers. Zhen (in Zhen et al. Citation2006a), who regarded T. bicostatus Ni (in Zeng et al. Citation1983) as a nomen nudum since no formal description or diagnosis of the species was given by the original author, proposed Triangulodus bifidus as the replacement (see Zhen et al. Citation2006a). After the publication of T. bifidus, it was brought to our attention that a brief description of T. bicostatus had been subsequently published by Ni & Li (Citation1987, pp. 444–445), implying that T. bicostatus might still have nomenclatorial priority over T. bifidus. The holotype of T. bicostatus (Zeng et al. Citation1983, pl. 10 fig. 41) is a symmetrical specimen that is likely assignable to the Sd element of T. bifidus as defined by Zhen (in Zhen et al. Citation2006a), and was actually from the upper part of the Fenxiang Formation immediately underlying the Honghuayuan Formation in the Yangtze Gorges area. However, as the type material of T. bicostatus Ni is still poorly known and not easily accessible for further examination, we contend that the fully documented species T. bifidus Zhen, Citation2006 (in Zhen et al. Citation2006a) is the preferred name for this distinctive form occurring in the Honghuayuan Formation.
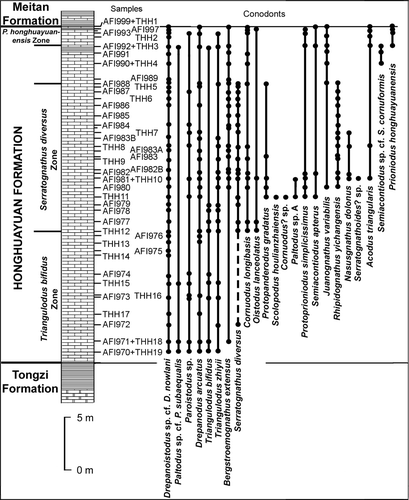
The conodont fauna reported by An (Citation1987, p. 29) from the Honghuayuan Formation in the type section exposed near Honghuayuan village consists of around 12 species, including (using An's original assignments) Cornuodus longibasis, Drepanodus arcuatus, Drepanoistodus sp., Paltodus sp., ?Tripodus variabilis and Serratognathus diversus in the lower to middle part, and Baltoniodus communis, Bergstroemognathus extensus robustus, B. hubeiensis, Hunghuayuangnathus ichangensis, Scalpellodus sp. and Scolopodus rex from the top 2 m. Our current investigation of the Honghuayuan Formation at its type locality () has resulted in the recovery of a much more diverse conodont fauna consisting of 24 species, indicative of a late Tremadocian to mid-Floian age. Three conodont biozones are recognized within the formation (Zhen Citation2007), in ascending order as outlined below.
Fig. 2 Stratigraphic section of the Honghuayuan Formation at Honghuayuan village near Tongzi County Town, showing the sampling locations and the stratigraphic ranges of the conodont species.
Triangulodus bifidus Biozone (Zhen Citation2007)
Definition. This is an interval zone with its base defined by the first occurrence (FAD) of T. bifidus and the top defined by the first appearance (FAD) of S. diversus. It was proposed and briefly discussed by Zhen (Citation2007).
Distribution
At its type locality (type section of the Honghuyuan Formation), this biozone is marked by the first appearance of T. bifidus near the base of the formation and extends to the middle part of the formation, with the top of the biozone (13 metres above the base) defined by the first appearance of S. diversus. Triangulodus bifidus has not been recorded in any of the samples from the underlying Tongzi Formation in the type locality (YYZ, unpublished collection). Regionally, the T. bifidus Biozone is distributed widely in the lower part of the Honghuayuan Formation on the Yangtze Platform. However, due to the diachronous nature of the base of the Honghuayuan Formation, the lower part of this biozone may extend laterally into the upper part of the underlying Fenxiang Formation in the Yangtze Gorges area (Zeng et al. Citation1983, Ni & Li Citation1987).
Characteristics and suggested age
The conodont fauna from the type section of the Honghuayuan Formation has a low diversity in this zone. Seven conodont species, including Bergstroemognathus extensus, Drepanoistodus sp. cf. D. nowlani, Drepanodus arcuatus, Paltodus sp. cf. P. subaequalis, Paroistodus sp., Triangulodus bifidus and T. zhiyii, were recovered from this interval (), and all extend into the middle and upper part of the formation. The occurrence of P. sp. cf. P. subaequalis in the lower part of the Honghuayuan Formation is crucial in regional correlation. This species was restricted to the latest Tremadocian, from the Drepanoistodus sp. aff. D. amoenus subzone to the lower part of the Tripodus subzone of the lower P. proteus Zone in Sweden (Löfgren Citation1997). The first appearance of this species near the base of the Honghuayuan Formation in the type section, therefore, suggests a late Tremadocian age correlated with the lower P. proteus Zone of the Baltoscandian successions (Zhen Citation2007). Triangulodus bifidus is morphologically distinctive and ranges through much of the Honghuayuan Formation, but is more common in the lower and middle part.
Serratognathus diversus Biozone (An 1981)
Definition
This biozone was initially named the Serratognathus Zone (An Citation1981, p. 108), and later renamed as the S. diversus Zone (An Citation1987, p. 18). An (Citation1981, Citation1987) defined it as an assemblage zone including conodont fauna from the entire Honghuayuan Formation in South China. This zone is redefined herein as a range zone with its base and the top defined by the first (FAD) and the last (LAD) appearances of S. diversus.
Distribution
At the type section of the Honghuayuan Formation (), the base of this biozone (corresponding to the entire range of S. diversus) is defined by the first confirmed occurrence of S. diversus in sample THH12, about 13 m above the base of the formation. Although a single specimen was recovered from sample AFI972, this species is absent from nine samples collected in the nine metres of limestone succession between this sample (AFI972) and the next sample (THH12) containing S. diversus (). The occurrence of a single specimen in the sample AFI972 may be due to contamination. The last appearance of S. diversus at the type section was recorded from sample AFI988 at about 5.5 m below the top of the formation (). Characterized by one of the most dominant species occurring in the Honghuayuan Formation of South China, this biozone can be widely recognized within the Honghuayuan Formation and other coeval stratigraphic units on the Yangtze Platform of South China, and typically occupies the middle part of the Honghuayuan Formation. However, as both lower and upper boundaries of the Honghuayuan Formation are regionally diachronous, the extent of the biozone within the formation may vary in different localities. This biozone can also be recognized in the Ordos Basin of North China (An & Zheng Citation1990), and in the outcrop and subsurface successions in the Tarim Basin (Zhao et al. Citation2000; see ).
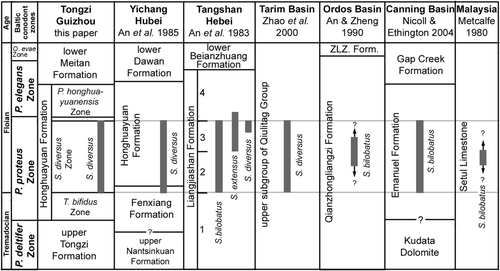
Fig. 3 Conodont zonation for the Honghuayuan Formation (latest Tremadocian to mid-Floian, Early Ordovician) on the Yangtze Platform, and correlation with coeval successions in other regions in China, Australia and Malaysia (ZLZ. Form.—Zhongliangzi Formation).
Characteristics and suggested age
At the Honghuayuan Formation type section, this biozone has a much higher diversity (21 conodont species) than the underlying zone (). Of these, S. diversus is the most distinctive and dominant species. Other species making their first appearance in this interval include Acodus triangularis, Cornuodus longibasis, C.? sp., Juanognathus variabilis, Nasusgnathus dolonus, Oistodus lanceolatus, Paltodus sp. A, Protopanderodus gradatus, Protoprioniodus simplicissimus, Rhipidognathus yichangensis, Scolopodus houlianzhaiensis, Semiacontiodus apterus and Serratognathoides? sp. Seven species, viz. Bergstroemognathus extensus, Drepanoistodus sp. cf. D. nowlani, Drepanodus arcuatus, Paltodus sp. cf. P. subaequalis, Paroistodus sp., Triangulodus bifidus and Triangulodus zhiyii, extend into this zone from the lower part of the formation ().
The S. diversus Zone is recognized in the Liangjiashan Formation of North China (), where the nominate species is associated with S. bilobatus and S. extensus. Occurrence of S. bilobatus and its association with several pandemic taxa in the Emanuel Formation of the Canning Basin in Western Australia allows correlation with the upper P. proteus Zone of the Baltoscandian successions (Zhen & Nicoll 2009). Therefore, an early Floian age is suggested for the S. diversus Zone.
Prioniodus honghuayuanensis Biozone (Zhen 2007; emend. herein)
Definition
The base of this interval zone is defined by the first appearance (FAD) of P. honghuayuanensis, with the top defined by the first appearance (FAD) of O. communis.
Distribution
At the type locality (type section of the Honghuayuan Formation, see ), this zone spans the top part of the formation (within an interval of 2.5 m), characterized by the occurrence of P. honghuayuanensis. This zone is widely distributed in the uppermost part of the Honghuayuan Formation on the Yangtze Platform (Zhen et al. Citation2005).
Prioniodus honghuayuanensis and O. communis are found in an overlapping stratigraphic relationship in the samples examined in this investigation. In the Honghuayuan Section, P. honghuayuanensis occurs abundantly in the top part of the Honghuayuan Formation, but neither P. honghuayuanensis nor O. communis was recovered from the basal part of the overlying Meitan Formation. However, in the Ganxi Section of Yanhe, P. honghuayuanensis was recovered from the very top of the Honghuayuan Formation (Zhen et al. Citation2005, Citation2007b), and O. communis was reported from the basal part of the overlying Meitan Formation (Zhen et al. Citation2007a). As both species were recorded within a very short interval in the Ganxi section, it is considered likely that they might have been adapted to different environments, in particular water depth or temperature.
Characteristics and suggested age
Fifteen conodont species are recorded in this interval (), most of them ranging from the underlying biozone. Morphologically, P. honghuayuanensis shows some resemblance to P. elegans (Zhen et al. Citation2005) and might be closely related to that species, but the former occurs in relatively shallower water environments. It is correlated with the lower P. elegans Zone (Floian, see ).
Remarks
At the type locality, there is a gap of about 3 m between the last appearance of S. diversus and the first appearance of P. honghuayuanensis (). However, at the Guanyinqiao Section of Qijiang, Sichuan Province, An (Citation1987, p. 33) reported the occurrence of both S. diversus and P. honghuayuanensis (assigned to B. communis) in an interval 4–7 m below the top of the Honghuayuan Formation.
Correlation of Serratognathus faunas
Three species of Serratognathus have been recognized that were widely distributed within eastern Gondwanan and peri- Gondwanan plates and terranes. The type species, S. bilobatus Lee, Citation1970, was originally reported from the Dumugol Formation of South Korea, which was correlated with an interval from the base of the P. elegans Zone to the lower O. evae Zone (Lee Citation1975). This species occurs in Korea, North China, South China, Australia and Malaysia. A second species, S. extensus Yang (in An et al. Citation1983) with a robust cusp and much more extended lateral processes, was reported only from the Liangjiashan Formation of the North China Platform. The species present in the Honghuayuan Formation, S. diversus, is apparently restricted in distribution to South China, North China (An et al. Citation1983) and the Tarim Basin (Gao Citation1991, Zhao et al. Citation2000). These species of Serratognathus are important in regional correlations of stratigraphic units in China, Korea, Southeast Asia and Australia ().
In South China, the S. diversus Zone is defined by the occurrence of S. diversus, commonly co-occurring with S. bilobatus in the Lower Yangtze Valley (An & Ding Citation1982, Citation1985), where it was correlated with the T. approximatus graptolite Zone (early Floian age) in deeper water graptolitic facies laterally equivalent to the carbonate facies of the Honghuayuan Formation (Chen & Wang Citation1993, Chen Xu et al. Citation1995). In Guizhou and Sichuan, this zone typically characterizes the middle and lower upper part of the Honghuayuan Formation. However, due to the regionally diachronous nature of the Honghuayuan Formation on the Yangtze Platform, S. diversus was recognized only in the lower part of the Honghuayuan Formation and in the upper part of the underlying Fenxiang (Fenhsiang) Formation in the Yangtze Gorges area of Hubei (An Citation1987, p. 42). In Anhui, Serratognathus species were found extending through most of the 24 m thick Honghuayuan Formation in the Hexian Section (An & Ding Citation1985, ). Serratognathus diversus was also reported from the Shanpianshan Formation of Wenshan, eastern Yunnan (Dong & Wang Citation2006), representing the southernmost distribution of the species in South China.
Yellow River Area (North China Platform)
The Ordovician succession on the North China Platform, deposited in a tropical, semi-enclosed epicontinental sea, is dominated by carbonates and evaporites about 500–1000 m thick. These rocks conformably or disconformably overlie upper Cambrian strata, and are disconformably overlain by Carboniferous coal measures. The Upper Ordovician is either not preserved or was not deposited over most of the area due to uplift of the platform in the Katian. However, the age of the youngest preserved Ordovician varies considerably across the region (Zhang & Zhen Citation1991).
The Liangjiashan Formation, consisting of a lower limestone member and an upper dolomite member, is widely distributed in North China and has conformable contacts with the underlying Yeli (Yehli) Formation and the overlying Beianzhuang Formation. At its type locality at Liangjiashan, about 25 km north of Qinhuangdao city, Liaoning, the formation might be incompletely preserved due to its disconformable contact with overlying Upper Carboniferous rocks. Hence, the Zhaogezhuang Section near Tangshan, Hebei Province, was selected as the auxiliary section, where the Liangjiashan Formation is a 160 m succession of thick-bedded limestone and dolomite (Chen Xu et al. Citation1995). Although much less abundant in each sample compared with yields in samples from the Honghuayuan Formation in South China, Serratognathus is widely distributed in the Liangjiashan Formation with all three species recognized. An et al. (Citation1983) established four conodont biozones within the Liangjiashan Formation in ascending order (): (1) Scalpellodus tarsus Zone; (2) Serratognathus bilobatus Zone; (3) Serratognathus extensus Zone; and (4) Paraserratognathus paltodiformis Zone. In the Zhaogezhuang Section of Tangshan studied by An et al. (Citation1983), S. bilobatus has a relatively long range, extending from the upper beds of the lower part (upper 6 m of Bed 18, see An et al. Citation1983, table 6) to the middle part of the Liangjiashan Formation (lower part of Bed 23, see An et al. Citation1983, table 6). Serratognathus extensus first appears in the upper part of Bed 19 and ranges to the middle part of Bed 23. Serratognathus diversus had a comparatively short range, limited to the top of Bed 22 and lower part of Bed 23. Therefore, the three species of Serratognathus co-occur only in Bed 23. The S. bilobatus Zone is confined to the lower part of the Liangjiashan Formation (upper part of Bed 18 and lower part of Bed 19) with a total thickness of 30 m in the Zhaogezhuang Section, and the overlying S. extensus Zone ranges through the middle part of the formation (upper 10 m of Bed 19, continuing to Bed 23) with a total thickness of 50 m (An et al. Citation1983, p. 26). Based on the occurrences of Serratognathus, Bergstroemognathus, Acodus, and Scolopodus, An et al. (Citation1983, table 8) correlated the S. bilobatus Zone and the S. extensus Zone in the Liangjiashan Formation with the Honghuayuan Formation in South China, and with the P. proteus and P. elegans zones of the Balto-Scandian successions.
Conodonts from the Liangjiashan Formation in North China represent faunas from a typical shallow water tropical setting and show a close biogeographic link with eastern Gondwana (Australia) and peri-Gondwanan plates and terranes, evidenced by the common occurrence of Serratognathus. However, the occurrence of Paraserratognathus An (in An et al. Citation1983), which is more closely related to Scolopodus than Serratognathus, in the upper part of the Liangjiashan Formation, also suggests links with coeval North American mid-continent faunas (see further discussion in Systematics section regarding Serratognathus).
Ordos Basin
The Ordos Basin is located in the border area of Shaanxi, Gansu, Ningxia and Inner Mongolia, encircled by the big bend of the Yellow River from the north. It is infilled by Mesozoic–Cenozoic deposits and loess, with Palaeozoic rocks exposed only in the mountains along the basin margins. A complete succession of Ordovician rocks is well exposed and formed the west marginal belt of the North China Platform.
A Serratognathus fauna, represented by S. bilobatus, Bergstroemognathus sp., Scolopodus houlianzhaiensis and other species, was reported from the middle part of the Qianzhongliangzi Formation exposed in the Helanshan area at the western edge of the Ordos Basin (An & Zheng Citation1990, p. 43). This formation consists of medium- to thick-bedded limestone about 400 m thick, and correlates to the Yeli, Liangjiashan, and Beianzhuang formations on the North China Platform. The occurrence of S. bilobatus forms the basis for direct correlation with the Liangjiashan Formation of Hebei and Liaoning in North China and the Honghuayuan Formation of South China.
Tarim Basin
The Tarim Basin is now covered by one of the world's largest sandy deserts (the Taklamakan Desert) in its interior. It is encircled by the Tianshan Mountains along its northern and western margins, and is bounded by the Kunlun Mountains to the south and the Aerjin Mountains to the southeast. The Tarim palaeo-plate consists of a complex Precambrian basement overlain by thick Phanerozoic strata. During the Ordovician, carbonate-dominated platform successions were deposited in much of the region with marginal basin or trough deposits fringing its north (Zhou et al. Citation2007). Intensive biostratigraphic studies of the Ordovician rocks in the region over the last three decades have been stimulated by its high potential for fossil fuel reserves (Zhao et al. Citation2000).
Serratognathus diversus was reported from the Lower Ordovician (upper subgroup of the Qiulitag Group) exposed in the Bachu area and from subsurface core samples in the Central Tarim and Lunnan areas (Wang & Zhou Citation1998, Zhao et al. Citation2000, Wang & Qi Citation2001). Zhao et al. (Citation2000, p. 7) noted that the S. diversus Zone spanned a much greater thickness of the carbonate successions (with a maximum thickness of nearly 600 m) in the Tarim Basin than in South China, and recorded an apparent thickness of 377 m for the range of S. diversus in core samples of Central Tarim Well 162 (Zhao et al. Citation2000, pp. 112–114). Zhao et al. subdivided this zone into three subzones (in ascending order): (1) Colaptoconus hemisphaericus Subzone; (2) Colaptoconus tarimensis Subzone; and (3) Pteracontiodus exilis–Bergstroemognathus extensus Subzone. The much thicker succession occupied by the S. diversus Zone in the Tarim Basin suggests that during the Early Ordovician, the Tarim Plate may have been located in a tropical setting with much higher depositional rates. The most important characteristic of the Tarim S. diversus fauna is the common appearance of Colaptoconus Kennedy, Citation1994 (a replacement name for Glyptoconus Kennedy, Citation1980), which is interpreted as being restricted to very shallow (peri-tidal) and warm environments (Ji & Barnes Citation1994b). This genus was recognized as a typical element of the Laurentian Province (North American mid-continent) of the Tropical Domain within the Shallow-Sea Realm during the Floian and late Tremadocian (Zhen & Percival Citation2003). In the recent revision of Ulrichodina Furnish, Citation1938, Landing (see Landing et al. Citation2003, Landing & Westrop Citation2006) suggested that Colaptoconus should be a junior synonym of Ulrichodina.
Australia
A diverse Serratognathus fauna, including S. bilobatus, Lissoepikodus nudus, Bergstroemognathus extensus, ?Microzarkodina adentata, Jumudontus brevis, Paroistodus parallelus, Paracordylodus gracilis and Stiptognathus sp. A, was reported from the middle member of the Emanuel Formation in the Canning Basin of Western Australia (Nicoll & Ethington Citation2004, Zhen & Nicoll 2009). Although S. bilobatus is relatively rare in the fauna, it spans a great stratigraphical thickness in the formation. Nicoll & Ethington (Citation2004) correlated this fauna with the upper P. proteus Zone to lower P. elegans Zone (). A similar but less diverse fauna bearing S. bilobatus, B. extensus, Scolopodus sp., Drepanoistodus sp. and others was recently recovered from core samples of petroleum exploration wells drilled in the Arafura Basin off the northern coast of Australia (Nicoll Citation2006, Nicoll et al. in prep.). The occurrence of S. bilobatus in Early Ordovician shallow marine carbonates in Australia suggests a close biogeographic link with Chinese faunas, particularly those from North China.
Malaysia
A small fauna including Serratognathus bilobatus was reported from the lower part of the Setul Limestone (unit 1, Lower Ordovician), exposed about 3.5 km northwest of Kaki Bukit, west Malaysia (Metcalfe Citation1980, Citation2003, Nicoll & Metcalfe Citation2001).
Sampling localities
Fifty-four conodont samples (with prefix THH and AFI) were collected from the Honghuayuan Formation type section in a 32 m thick limestone succession exposed on the hill slope to the southeast of Honghuayuan village (; location coordinates: 28°04.27′N 106°50.91′E), about 7 km south of Tongzi County town. Fifty of them () were productive and form the basis of this study, together with material obtained from samples collected from two other sections in Guizhou (). Of 35 samples collected from the Honghuayuan Formation (71.8 m thick) exposed southwest of Yanhe County town (, 28°22.72′N 108°25.70′E) at the Ganxi Section along the road from Ganxi village to Shichangao, 24 were productive and yielded a similar fauna (). Nine samples collected from the Honghuayuan Formation (27.4 m thick) in the Huanghuachong Section at Wudang, near Guiyang (; top of the Honghuayuan Formation at 26°37.83′N 106°46.21′E) were all productive (). However, as samples were collected at a much wider spacing in the Ganxi and Huanghuachong sections, only a general distribution pattern with much less detail has been revealed in comparison with the distribution of the fauna from the Honghuayuan type section ().
Table 1. Distribution of conodonts (numbers refer to quantity of specimens) in samples collected from the Honghuayuan Formation in the Honghuayuan Section (THH and AFI) of Tongzi, in the Ganxi Section (YTH) of Yanhe, and in the Huanghuachong Section (WHC) of Guiyang, Guizhou Province, South China
All samples (1–3 kg each) were completely dissolved in 10% acetic acid, with residues washed through sieves of upper 1.2 mm metal (14 mesh) and lower 0.25 mm cloth mesh sizes, and separated and concentrated using sodium polytungstate.
Systematic palaeontology
All photographic illustrations shown in to are SEM photomicrographs captured digitally (numbers with the prefix IY are the file names of the digital images). Figured specimens bearing the prefix AMF. are deposited in the collections of the Palaeontology Section at the Australian Museum, Sydney. Conodont terminology and notation employed in this contribution are conventional as defined in the Treatise Part W (Clark et al. Citation1981), except for the M element (makellate), whose orientation, morphology and terminology were introduced by Nicoll (Citation1990, Citation1992).
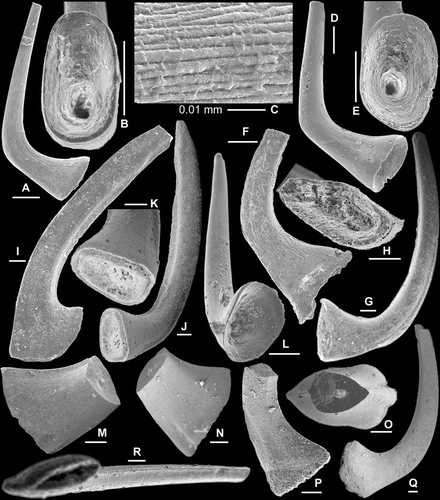
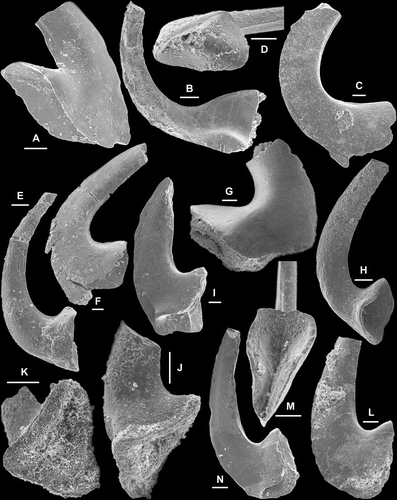
Fig. 4 A–L, Cornuodus longibasis (Lindström, Citation1955); A–C, Sa element, AMF.134989, AFI997, A, lateral view (IY119–046), B, basal view (IY119–048), C, close up showing fine surface striae (IY119–049); D–E, Sb element, AMF.134990, AFI997, D, inner lateral view (IY119–053), E, basal view (IY119–054); F, Sc element, AMF.134991, WHC36, inner lateral view (IY121–005); G–H, Sd element, AMF.134992, WHC36, G, inner lateral view (IY121–008), H, basal view (IY121–009); I, Pa element, AMF.134993, THH12, inner lateral view (IY120–033); J–L, Pb element; J–K, AMF.134994, WHC36, J, inner lateral view, K, close up of the basal part (IY121–006); L, AMF.134995, AFI997, basal inner lateral view (IY119–045). M–R, Cornuodus? sp.; M–O, Sa element, AMF.134996, THH11, M–N, lateral views (IY122002, IY122003), O, upper view (IY122001); P, Sb element, AMF.134997, THH11, inner lateral view (IY122006); Q–R, Pa element, AMF.134998, THH11, Q, inner lateral view (IY122004), R, basal view (IY122005). Scale bars=100 μm, unless indicated otherwise.
Phylum CHORDATA Balfour, 1880
Class CONODONTA Pander, 1856
Cornuodus Fåhraeus, 1966
Type species
Cornuodus erectus Fåhraeus, 1966 (=Drepanodus longibasis Lindström, Citation1955).
Cornuodus longibasis (Lindström, Citation1955) (–L)
1955 Drepanodus longibasis Lindström, p. 564, pl. 3, fig. 31.
1993 Scolopodus sunanensis An & Ding; Ding et al. in Wang, p. 206, pl. 6, –, 14, 15.
1999 Cornuodus longibasis (Lindström); Löfgren, p. 175–184, pls 1–3 (cum syn.). [1999a]
2004 Cornuodus longibasis (Lindström); Zhen et al., p. 52, pl. 2, fig. 19 (cum syn.).
2004 Cornuodus longibasis (Lindström); Zhen & Percival, p. 91, –J.
2005 Cornuodus longibasis (Lindström); Löfgren et al., –H.
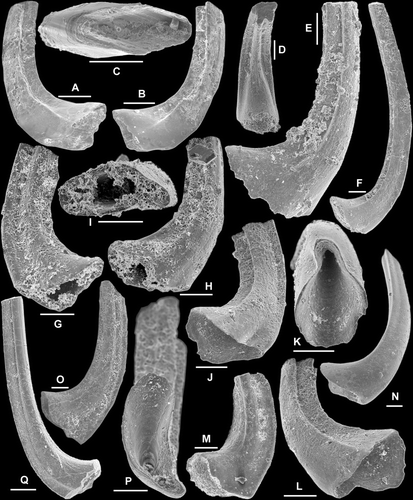
Fig. 5 Drepanodus arcuatus Pander, 1856. A, M element, AMF.134999, THH12, anterior view (IY120–014). B–C, Sa element; B, AMF.135000, YTH13, lateral view (IY62–006); C, AMF.135001, THH12, lateral view (IY120–032). D–E, Sb element, AMF.135002, AFI976, D, basal view (IY120–034), E, outer lateral view (IY120–035). F–G, Sc element; F, AMF.135003, AFI980, inner lateral view (IY120–038); G, AMF.135004, WHC36, outer lateral view (IY121–012). H, Sd? Element, AMF.135005, YTH23, outer lateral view (IY121–033). I–K, Pa element; I, AMF.135006, AFI980, outer lateral view (IY121–001); J–K, AMF.135007, AFI982, J, outer lateral view (IY55–037), K, basal view (IY55–036). L–N, Pb element; L, AMF.135008, THH10, outer lateral view (IY119–066); M–N, AMF.135009, AFI976, M, basal view (IY120–016), N, outer lateral view (IY120–017). Scale bars=100 μm.
2009 Cornuodus longibasis (Lindström); Zhen et al. in press, .
Material
One hundred and thirty-six specimens from 27 samples (see ).
Remarks
Cornuodus longibasis was revised by Löfgren (Citation1999a) as consisting of a seximembrate apparatus, and her species concept is applied herein. It occurs in many samples of the Honghuayuan Formation as a minor component of the fauna (). The P elements have a shorter base and can be differentiated into the Pa element with a recurved strongly laterally compressed cusp and an antero-posteriorly expanded base having an arched basal margin (). The Pb element has an erect to slightly reclined cusp bearing sharp anterior and posterior margins, and a less laterally compressed base (–L). The S elements have a longer base and an erect to slightly reclined cusp; they form a symmetry transition series, from symmetrical Sa, slightly asymmetrical Sb, to asymmetrical and laterally more compressed Sc. The Sd element is like the Pb element, but with the cusp twisted distally and typically having a longer, laterally more compressed base ().
Cornuodus? sp. (–R)
1993 Scandodus mecynus Gao; Ding et al. in Wang, p. 201, pl. 9, –.
Material
Three specimens from THH11.
Description
Elements recovered include symmetrical Sa (–O), asymmetrical Sb () and short-based Pa (–R); all are laterally compressed coniform elements with sharp anterior and posterior margins and a diamond- or oval-shaped cross-section of the cusp.
Remarks
Specimens referred herein to Cornuodus? sp. (–R) differ from C. longibasis in having a shorter base and a more laterally compressed cusp with a sharp anterior margin. These elements are comparable with those assigned to S. mecynus by Ding et al. (in Wang Citation1993) from the Honghuayuan Formation and Kunshan Formation in the Lower Yangtze area. However, the type material of the poorly known S. mecynus from the Honghuayuan Formation in Hubei displays sharp posterior and anterior margins and a postero-laterally located groove on each side (An et al. Citation1985, pl. 6, , 7). This latter feature is lacking both in our current material and that illustrated by Ding et al. (in Wang Citation1993).
Drepanodus Pander, 1856
Type species
Drepanodus arcuatus Pander, 1856.
Drepanodus arcuatus Pander, 1856 (–N)
1856 Drepanodus arcuatus Pander, p. 20, pl. 1, , 4–5, 17, 30, ?31.
1856 Drepanodus flexuosus Pander, p. 20, pl. 1, –.
Fig. 6 A–I, Juanognathus variabilis Serpagli, Citation1974; A–C, Sa element; A–B, AMF.135010, AFI991, A, posterior view (IY54–005), B, postero-basal view (IY54–006); C, AMF.135011, YTH4, postero-basal view (IY61–042); D–E, Sb element, AMF.135012, AFI991, D, posterior view (IY54–008), E, postero-outer lateral view (IY54–007); F–I, Sc element, F, AMF.135013, AFI991, inner lateral view (IY53–040); G–H, AMF.135014, AFI991, G, basal view (IY54–002), H, outer lateral view (IY54–003); I, AMF.135015, AFI980, outer lateral view (IY56–021). J–O, Oistodus lanceolatus Pander, 1856; J–M, Sa element, AMF.135016, THH10, J, anterior view (IY59–028), K, M, lateral views (IY59–029, IY59–031), L, posterior view (IY59–030). N, M element, AMF.135017, AFI977, outer lateral view (IY65–025). O, Sc element, AMF.135018, AFI997, posterior view (IY119–041). Scale bars=100 μm.
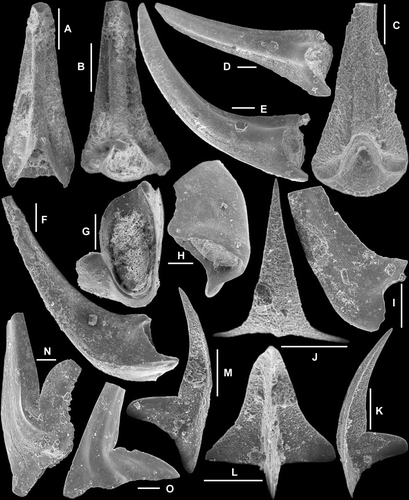
Fig. 7 A–I, Nasusgnathus dolonus (An, Citation1981); A–C, Sc element, A, AMF.135019, THH10, inner lateral view (IY120–004); B–C, AMF.135020, THH10, B, outer lateral view (IY120–002), C, basal view (IY120–001); D–E, Sb element, D, AMF.135021, THH10, outer lateral view (IY120–005), E, AMF.135022, THH7, outer lateral view (IY59–009); F–I, Sd element; F–G, AMF.135023, THH10, F, basal view (IY120–011), G, outer lateral view (IY120–010); H–I, AMF.135024, THH10, H, inner lateral view (IY120–007), I, close up showing fine surface striae (IY120–008). J–K, Paltodus sp. Sa element, AMF.135025, YTH16, J, basal view (IY121–029), K, lateral view (IY121–031). L–O, Protoprioniodus simplicissimus McTavish, 1973. Sb element, AMF.135026, AFI982, L, inner lateral view (IY55–027), M, upper view (IY55–028), N, anterior view (IY55–024), O, outer lateral view (IY55–025). Scale bars=100 μm, unless indicated otherwise.

Fig. 8 Protopanderodus gradatus Serpagli, 1974. A–C, Sa element, AMF.135027, YTH1, A, B, lateral views (IY61–031, IY61–032), C, basal view (IY61–030). D–F, Sb element; D, AMF.135028, AFI981, posterior view (IY56–002); E, AMF.135029, THH11, outer lateral view (IY120–031); F, AMF.135030, AFI980, inner lateral view (IY120–041). G–I, Sc element, AMF.135031, THH10, G, inner lateral view (IY120–023), H, outer lateral view (IY120–022), I, basal view (IY120–021). J–L, Sd element, AMF.135032, YTH5, J, inner lateral view (IY121–024), K, basal view (IY121–023), L, outer lateral view (IY121–025). M, Pb element, AMF.135033, AFI980, inner lateral view (IY57–008). N–Q, Pa element; N, AMF.135034, WHC31, outer lateral view (IY121–017); O–P, AMF.135035, YTH4, O, inner lateral view (IY61–045), P, basal view (IY61–043); Q, AMF.135036, WHC31, inner lateral view (IY121–016). Scale bars=100 μm.
1981 Drepanodus perlongus An in An et al., p. 108, pl. 1, .
1982 Drepanodus perlongus An in An et al.; An & Ding, pl. 1, .
1985 Drepanodus perlongus An in An et al.; An et al., p. 39, pl. 5, , 6, text-.8.
1987 Drepanodus perlongus An in An et al.; An, p. 144, pl. 9, , pl. 10, –, pl. 15, fig. 15 (cum syn.).
1993 Drepanodus perlongus An in An et al.; Ding et al. in Wang, p. 172, pl. 10, –, 13–14, 21, 25.
2003 Drepanodus arcuatus Pander; Löfgren & Tolmacheva, p. 211–215, , 3A–C, E–H, 5K–V, 6M–U, 7H–N, 8A–G (cum syn.).
2004 Drepanodus arcuatus Pander; Zhen et al., p. 52–53, pl. 3, –
Fig. 9 A–C, Protoprioniodus simplicissimus McTavish, Citation1973; A, Pa element, AMF.135037, THH11, outer lateral view (IY120–029), B, Pb element, AMF.135038, AFI981, outer lateral view (IY55–040), C, M element, AMF.135039, AFI980, posterior view (IY56–028). D–F, Semiacontiodus sp. cf. S. cornuformis (Sergeeva, Citation1963); D–E, Sb element, AMF.135040, AFI992, D, posterior view (IY53–032), E, inner lateral view (IY53–033); F, Sa element, AMF.135041, AFI992, upper posterior view (IY53–029). G–L, Scolopodus houlianzhaiensis An & Xu in An et al., Citation1983; G–I, Sc element, AMF.135042, WHC38, G, basal-posterior view (IY62–013), H, inner lateral view (IY62–014), I, outer lateral view (IY62–015); J–L, Sb element, AMF.135043, THH11, J, inner lateral view (IY120–028), K, outer lateral view (IY120–027), L, upper-anterior view (IY120–026). Scale bars=100 μm.
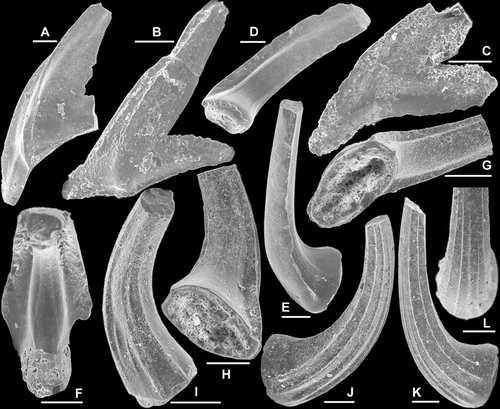
Fig. 10 Semiacontiodus apterus (Gao in An et al., Citation1985). A–C, Sb element, AMF.135058, AFI982, A, outer lateral view (IY55–034), B, postero-basal view (IY55–033), C, inner lateral view (IY55–035). D–F, Sa element, AMF.135059, THH3, D, posterior view (IY59–006), E–F, lateral views (IY59–008, IY59–007). G–H, Sc element, AMF.135060, WHC34, G, inner lateral view (IY62–020), H, basal view (IY62–021). I–K, Sd element, AMF.135053, YTH1, I, lateral view (IY61–028), J, posterior view, and K, enlargement showing the outline of the base (IY61–026). Scale bars=100 μm.
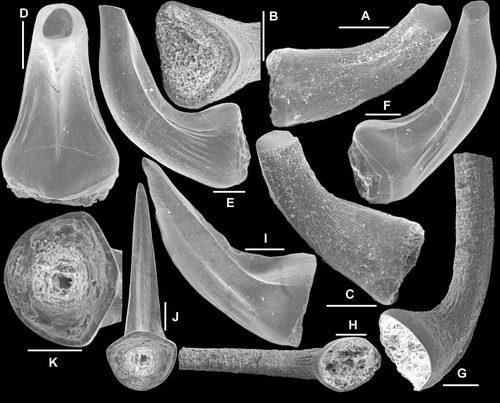
Fig. 11 A–C, Serratognathoides? sp. Sb-c element, AMF.135054, THH10; A–B, oblique upper views (IY49–019, IY49–017), C, close up showing the posterior edge of the unit (IY49–018). D–M, Serratognathus diversus An, 1981. Sc element; D–F, AMF.132591, THH12, D, posterior view (IY12–049), E, basal view (IY12–050), F, anterior view (IY48–018); G, AMF.135055, THH6, inner lateral view (IY12–029); H–I, AMF.135056, THH7, H, basal view (IY48–015), I, upper view (IY12–033); J–M, AMF.135057, THH10, J, posterior view (IY119–037), K, upper view (IY119–033), L, basal-posterior view, close up showing striae structure (IY119–031), M, upper view, close up showing cross-section of the cusp and adjacent denticles (IY119–036). Scale bars=100 μm, unless indicated otherwise.

Fig. 12 Serratognathus diversus An, 1981. Sa element. A, AMF.135044, THH7, upper view (IY119–001); B, AMF. 132592, THH6, anterior view (IY12–020); C, AMF.135045, THH6, anterior view (IY48–005); D, AMF. 132594, THH8, basal view (IY12–038); E, AMF.135046, THH6, posterior view (IY48–008); F, AMF.135047, THH10, lateral view (IY119–038); G–K, AMF.135048, THH7, G, basal view (IY119–008), H, posterior view (IY119–005), I, posterior view, enlargement showing surface striae (IY119–006), J, posterior view, enlargement showing laminated structure of posterior margin of the cusp (IY119–006), K, basal view, enlargement showing the circular ring structure of the initial growth disk (IY119–009). Scale bars=100 μm, unless indicated otherwise.
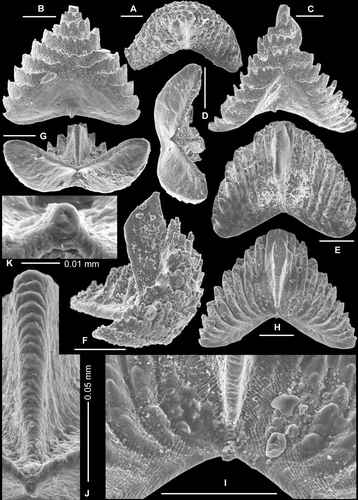
Fig. 13 Serratognathus diversus An, 1981. Sb element. A–B, AMF.135049, THH7, A, antero-outer lateral view (IY119–018), B, upper-anterior view (IY119–017). C–F, AMF.135050, THH7, C, posterior view, close up showing laminated structure on posterior margin of the cusp (IY119–022), D, posterior view (IY119–020), E, antero-basal view (IY119–019), F, posterior view, enlargement of Fig. 13C, showing laminated posterior margin of the cusp, and circular ring structure of the initial growth disk (IY119–022). G, AMF.135051, THH7, anterior view (IY48–006). H, AMF.135052, THH6, posterior view (IY12–025). I–K, AMF.132593, THH6, I, posterior view (IY12–022), J, posterior view, enlargement showing rows of denticles on the lateral process (IY12–024), K, posterior view, close up showing laminated structure on the posterior margin of the cusp and circular ring structure of the initial growth disk (IY12–023). Scale bars=100 μm.

Material
One hundred and forty-one specimens from 36 samples (see ).
Remarks
In the recent comprehensive revision of D. arcuatus by Löfgren & Tolmacheva (Citation2003), they selected a neotype (as Pander's original types are lost) and defined the species as consisting of a seximembrate apparatus. This species concept has been followed herein, with all element types being recognized in the collection from the Honghuayuan Formation. Some specimens we have ascribed to D. arcuatus (e.g. ) are identical with Drepanodus perlongus An, 1981. The holotype of this latter species from the Dawan Formation in Hubei was initially illustrated by An et al. (Citation1981, pl. 1, ), and later described (An et al. Citation1985; An Citation1987) as a form species similar to D. arcuatus but with a longer base. In fact, it is very similar to the symmetrical (Sa) element of the original types of D. arcuatus illustrated by Pander (see Löfgren & Tolmacheva Citation2003, a), and hence D. perlongus is considered to be a junior synonym of D. arcuatus.
Juanognathus Serpagli, 1974
Type species
Juanognathus variabilis Serpagli, 1974.
Juanognathus variabilis Serpagli, Citation1974 (–I)
1974 Juanognathus variabilis Serpagli, p. 49, pl. 11, –b, pl. 22, figs 6–17, text-.
2004 Juanognathus variabilis Serpagli; Zhen et al., pp. 53–54, pl. 4, figs 1–14 (cum syn.).
Material
Eighty-six specimens from 15 samples (see ).
Remarks
As discussed recently (Zhen et al. Citation2004), this species might have either a seximembrate or septimembrate apparatus. However, as only three juanognathiform S elements including symmetrical Sa (–C), asymmetrical Sb (–E) and strongly asymmetrical Sc (–I) elements were recovered, mainly from the upper part of the Honghuayuan Formation, we are unable to resolve on this question.
Nasusgnathus Ni in Zeng et al., 1983
1987 Nasusgnathus Zeng et al., Citation1983; An, p. 159.
1987 Nasusgnathus Ni, 1983; Ni & Li, p. 417.
2000 Nasusgnathus Zeng et al., Citation1983; Zhao et al., p. 206.
Type species
Drepanodus dolonus An in An et al., 1981.
Remarks
Nasusgnathus was informally introduced by Zeng et al. (Citation1983, p. 54) in the caption to a figure without formal generic diagnosis or designation of type species, and was subsequently used by several Chinese conodont workers (e.g. An Citation1987, Zhao et al. Citation2000). However, the genus was later formally published with Ni as the authority rather than Zeng et al., Citation1983 (see Ni & Li Citation1987). The genus includes N. dolonus and N. giganteus Ni (in Ni & Li Citation1987) that are interpreted to include symmetrical–asymmetrical albid, laterally compressed, coniform elements.
Nasusgnathus dolonus (An in An et al., Citation1981) (–I)
1981 Drepanodus dolonus An in An et al., pl. 1, –.
1983 Nasusgnathus dolonus (An); Ni in Zeng et al., p. 54, pl. 11, fig. 29.
1984 Drepanodus dolonus An; An & Xu, pl. 1, .
1985 Drepanodus dolonus An; An et al., pp. 38–39, pl. 5, figs 24–26, text-fig. 5.6.
1987 Nasusgnathus dolonus (An); An, p. 159, pl. 12, –, 7, pl. 30, .
1987 Nasusgnathus dolonus (An); Ni & Li, p. 418, pl. 53, figs 21–23, text-fig. 5–80.
1995 Nasusgnathus dolonus (An); Chen X.Y. et al., p. 2, .
2000 Nasusgnathus dolonus (An); Zhao et al., p. 207, pl. 19, –.
Material
Ten specimens from five samples (see ).
Description. Coniform species having at least four elements forming a symmetry transition series: Sa element (not recognized in the current collection); Sc element most strongly compressed laterally with a posteriorly extended base (–C), Sb element resembling Sc, but without posterior extension of the base (–E); Sd element less laterally compressed with a narrower outline in lateral view (–G); all elements albid, ornamented with fine striae on the lateral surface ().
Remarks
Nasusgnathus dolonus was initially named by An in the caption to an illustration of the designated holotype (in An et al. Citation1981, pl. 1, ), and was formally described by An et al. (Citation1985, p. 38–39) as a form species of symmetrical or nearly symmetrical coniform elements that they doubtfully ascribed to Drepanodus. Ni & Li (Citation1987) recognized a bimembrate apparatus for N. dolonus including symmetrical and asymmetrical elements, and designated it as the type species of Nasusgnathus. Based on the general morphology, deep basal cavity, and oval cross-section, they suggested that Nasusgnathus might have a close phylogenetic relationship with Scalpellodus. However, the symmetrical element illustrated by Ni & Li (Citation1987, pl. 53, figs 21–22) is larger with a less laterally compressed cusp, and is only doubtfully assigned to the Sa position. Three asymmetrical morphotypes of N. dolonus recovered from the Honghuayuan Formation are considered to represent the Sb, Sc and Sd elements, which form a transitional series.
Oistodus Pander, 1856
Type species
Oistodus lanceolatus Pander, 1856.
Oistodus lanceolatus Pander, 1856 (–O)
1955 Oistodus lanceolatus Pander; Lindström, p. 577, pl. 3, figs 58–60.
1955 Oistodus delta Lindström, p. 573, pl. 3, –.
1955 Oistodus triangularis Lindström, p. 581, pl. 4, figs 14–18.
1974 Oistodus lanceolatus Pander; Lindström in Ziegler, p. 201, Oistodus–plate 1, –.
1985 Oistodus lanceolatus Pander; An et al., pl. 6, .
1987 Oistodus lanceolatus Pander; An, p. 160, pl. 12, figs 10–15, 24.
1993 Oistodus lanceolatus Pander; Löfgren, –X.
1994 Oistodus lanceolatus Pander; Löfgren, .38–6.40.
2001 Oistodus lanceolatus Pander; Viira et al., –y.
2001 Oistodus lanceolatus Pander; Tolmacheva & Fedorov, –R.
2005 Oistodus lanceolatus Pander; Löfgren et al., –Y.
Material
Ten specimens from six samples (see ).
Remarks
Oistodus lanceolatus is rare in the Honghuayuan Formation, and only symmetrical Sa (–M), asymmetrical Sc () and geniculate M () elements have been recovered.
Paltodus Pander, 1856
Type species
Paltodus subaequalis Pander, 1856.
Paltodus? sp. (–K)
Material
Two specimens from two samples, YTH16 and YTH20 (see ).
Remarks
In a recent description of drepanoistodontid conodonts from the Honghuayuan Formation (Zhen et al. Citation2007b), we documented a species of Paltodus referred to as P. sp. cf. P. subaequalis, but no Sa element was then recognized. Subsequently, two well-preserved specimens of the Sa element have been identified in two samples and one is illustrated herein (–K). It shows some resemblence to the Sa elements of P. subaequalis (see Löfgren Citation1997, S) and P. sp. cf. P. subaequalis (see Löfgren Citation1997, K), except that the Honghuayuan specimen has a more strongly proclined cusp, weaker development of the lateral costae and a stronger anticusp-like extension of the antero-basal corner compared with the Swedish material. However, its general morphology also resembles N. dolonus. It is possible that this element may represent the symmetrical Sa element of the latter species. This taxonomic uncertainty will have to remain until there is more material available. Therefore, it is doubtfully assigned to Paltodus pending further study.
Protopanderodus Lindström, 1971
Type species
Acontiodus rectus Lindström, 1955.
Protopanderodus gradatus Serpagli, Citation1974 (–Q)
1974 Protopanderodus gradatus Serpagli, p. 75, pl. 15, –b, pl. 26, figs 11–15, pl. 30, –b, text-fig. 17.
2004 Protopanderodus gradatus Serpagli; Zhen et al., pp. 56–57, pl. 5, – (cum syn.).
2006 Protopanderodus gradatus Serpagli; Zhen & Percival, –C.
Material
Thirty-three specimens from 11 samples (see ).
Description
A species of Protopanderodus consisting of a seximembrate apparatus; Sa element (–C) symmetrical, with a sharp posterior margin and a sharp costa on each side; Sb element (–F) like Sa but asymmetrical, with costa on the less convex inner lateral face typically located more posteriorly (); Sc element (–I) strongly asymmetrical with a mid-costa on the outer lateral face, and two costae on the inner lateral face; Sd element (–L) like Sc but less laterally compressed; P elements with a smooth convex outer lateral face and a costa on the inner lateral face; Pa element typically with a proclined cusp (–Q), and Pb with a suberect cusp ().
Remarks
Serpagli (Citation1974) documented the symmetry transition in detail and recognized three basic morphotypes (symmetrical, asymmetrical with twisted cusp, and asymmetrical–subsymmetrical) and intermediate forms among the type material of this species from the San Juan Formation of Precordilleran Argentina. It was then interpreted as consisting of a quadrimembrate (Dzik Citation1994) or quinquimembrate (Albanesi in Albanesi et al. Citation1998, Zhen et al. Citation2004) apparatus. With recognition of the two P elements, P. gradatus is defined herein as constituting a seximembrate apparatus.
Protoprioniodus McTavish, 1973
Type species
Protoprioniodus simplicissimus McTavish, 1973.
Remarks
Cooper (Citation1981) suggested that Oelandodus van Wamel, Citation1974 was a junior synonym of Protoprioniodus, a direct descendent of Acodus, whereas some subsequent authors (e.g. Bagnoli et al. Citation1988, Stouge & Bagnoli Citation1988, Sweet Citation1988, Rasmussen Citation2001, Tolmacheva et al. Citation2001) treated Oelandodus as a valid genus. Van Wamel (Citation1974) noted the close resemblance between Oelandodus and Oistodus. The latter consists of dominantly hyaline elements. Moreover, some specimens ascribed to Oelandodus elongatus (Lindström, Citation1955) and Oelandodus? costatus van Wamel, Citation1974 by Bagnoli et al. (Citation1988, pl. 39, , 9–10) seem more similar to Oistodus than either Protoprioniodus or Acodus, both of which comprise dominantly small, albid elements. As discussed by Zhen (in Zhen et al. Citation2003), Protoprioniodus differs from Cooperignathus in having pastinate P elements with a robust cusp.
Protoprioniodus simplicissimus McTavish, Citation1973 (–O; 9A–C)
1973 Protoprioniodus simplicissimus McTavish, pp. 48–49, pl. 2, , 8–9.
1984 Protoprioniodus costatus (van Wamel); An & Xu, pl. 1, .
1985 Oelandodus costatus van Wamel; An et al., pl. 6, figs 19–21.
1987 Protoprioniodus costatus (van Wamel); An, p. 174–175, pl. 14, , 4–5, pl. 16, fig. 25.
1987 Protoprioniodus aff. simplicissimus McTavish; An, pl. 16, partim only fig. 18.
1993 Protoprioniodus simplicissimus McTavish; Ding et al. in Wang, p. 197, pl. 15, , 11.
1993 Protoprioniodus costatus (van Wamel); Ding et al. in Wang, p. 197, pl. 14, fig. 27.
1994 Protoprioniodus simplicissimus McTavish; Ji & Barnes, pp. 54–55, partim only pl. 16, , 12–18. [1994a]
2001 Protoprioniodus simplicissimus McTavish; Tolmacheva et al., .17, 5.22.
Material
Ten specimens from eight samples (see ).
Remarks
Only M, Sb, and P elements of this species have been recovered in our collections. Specimens referrable to this species from the Honghuayuan Formation and the overlying Dawan Formation in South China were previously ascribed to either P. simplicissimus or O. costatus (see synonymy list).
Scolopodus Pander, 1856
Type species
Scolopodus sublaevis Pander, 1856.
Scolopodus houlianzhaiensis An & Xu in An et al., Citation1983 (–L)
1983 Scolopodus rex houlianzhaiensis An & Xu in An et al., p. 148, pl. 12, figs 23–27, text-.7, 11.8.
?1987 Scolopodus rex houlianzhaiensis An & Xu in An et al.; Ding, pl. 6, fig. 23.
non 1993 Scolopodus rex houlianzhaiensis An & Xu in An et al.; Ding et al. in Wang, Citation1993, p. 205, pl. 14, figs 13–15.
Material
Two specimens from two samples (see ).
Remarks
This species is rare in the Honghuayuan Formation and is only represented in the present collection by two specimens referable to the strongly asymmetrical Sc (–I) and weakly asymmetrical Sb (–L) elements. The symmetrical and weakly asymmetrical multicostate Sa and Sb elements of S. houlianzhaiensis show some resemblance to S. striatus Pander, 1856, but the former typically bear more costae. Moreover, S. houlianzhaiensis has a distinctive Sc element with a strongly compressed cusp and with lateral costae restricted to the outer lateral face, more commonly near the anterior margin. Scolopodus striatus was revised recently on material, considered to be part of Pander's original type collection from the Lower Ordovician in the vicinity of St. Petersburg, as having a quinquimembrate apparatus (Tolmacheva Citation2006). Both S. quadratus Pander, 1856, and S. rex Lindström, Citation1955, which are widely cited in the Ordovician conodont literature, were regarded as junior synonyms of S. striatus.
Semiacontiodus Miller, 1969
Type species
Semiacontiodus nogamii Miller, 1969.
Remarks
In a detailed study of several Ordovician species of Semiacontiodus from Baltoscandia, Löfgren (Citation1999b) suggested a seximembrate apparatus including Sa, Sb, Sc, Sd, Pa, and Pb elements. This concept is accepted, although a complete species apparatus has not yet been recovered for either of the two species documented herein. In our collections, this genus is only represented by a few specimens that are dominantly hyaline, rather than albid as are those described from western Newfoundland (Ji & Barnes Citation1994a). Semiacontiodus can be differentiated from hyaline Colaptoconus Kennedy, Citation1994 (a replacement name for Glyptoconus Kennedy, Citation1980) by lacking deep grooves and coarse costae.
Semiacontiodus apterus (Gao in An et al., Citation1985) (–K)
1985 Scolopodus apterus Gao in An et al., p. 41, pl. 8, , 18, text-.3, 5.7.
1987 Scolopodus apterus Gao in An et al.; An, p. 181, pl. 7, , 27.
Material
Five specimens from five samples (see ).
Description
Sa element (–F) symmetrical, having a proclined cusp with a prominent costa along the posterior margin and a broad anterior face, lateral face ornamented with a strong costa, and commonly several weaker and shorter secondary costae. Sb element (–C) asymmetrical, with broad anterior and posterior faces and a costa on each side, that on the inner lateral face is located more towards anterior. Sc element (–H) weakly asymmetrical and laterally compressed, with a proclined to nearly suberect cusp, and a shorter base with oval outline in basal view. Sd element (–K) like Sa, symmetrical with a proclined cusp, but more antero-posteriorly compressed and lateral faces with two costae, base more or less pentagonal in outline. Fine striae are better developed on the lateral face of the asymmetrical Sb and Sc elements.
Remarks
This is a rare species in the Honghuayuan Formation, originally described as a form species with only a symmetrical Sa element recovered from this unit in the Huanghuachang Section of Hubei (Gao in An et al. Citation1985). The holotype (An et al., Citation1985, pl. 8, fig. 18) has a short, non-flared base, a reclined cusp with rounded anterior face, and a prominent costa along the posterior margin and on each side. It differs from our specimens only in having a reclined rather than proclined cusp (–F). Gao (in An et al. Citation1985) noted that short finer costae may be developed on the basal part of the type specimens, a feature also observed on the current material ().
Semiacontiodus sp. cf. S. cornuformis (Sergeeva, Citation1963) (–F)
1987 Scolopodus cornuformis Sergeeva; An, p. 183, pl. 7, –, 13–16.
1993 Scolopodus cornuformis Sergeeva; Ding et al. in Wang, p. 202, pl. 5, fig. 33.
Material
Eight specimens from three samples (see ).
Remarks
These specimens are comparable with those illustrated as Scolopodus cornuformis from the Dawan Formation in Sichuan (An Citation1987) and Anhui (Ding et al. in Wang Citation1993) in South China. Löfgren (Citation1999b) revised S. cornuformis in detail and considered it to be a typical species of Semiacontiodus. Löfgren (Citation1999b) also doubtfully included An's material in her synonymy of S. cornuformis. However, the species from the Honghuayuan Formation has sharper lateral costae and less prominent striae on the surface.
Semiacontiodus sunanensis (An & Ding, Citation1982), with less strong costa on the lateral faces, is reported from the Honghuayuan and Dawan formations in South China (An Citation1987; non Ding et al. in Wang 1993=C. longibasis) and the Liangjiashan Formation of North China (see An et al. Citation1983, pl. 13, figs 8–14). It might be closely related to S. cornuforms, particularly those early forms of this species that Löfgren (Citation1999b) assigned to S. sp. cf. S. cornuformis.
Serratognathoides An, 1987
Type species
Serratognathoides chuxianensis An, 1987.
Remarks
Serratognathoides is closely related morphologically to Serratognathus, but its elements are much more flattened with an indistinct cusp and less prominent rows of denticles on the upper face. Two species have been recognized from the Lower Ordovician in China. The type species S. chuxianensis, from the Dawan Formation in Anhui Province, was established as a form species consisting of a single element. An (Citation1987, p. 189) recognized its resemblance to Serratognathus bilobatus, and noted that S. chuxianensis had the rows of denticles only weakly developed. This species was also reported from the upper subgroup of the Qiulitag Group (Lower Ordovician) in the Tarim Basin (Gao Citation1991, Zhao et al. Citation2000). The second species, S. aksuensis Gao, Citation1991, was recorded only from subsurface rocks of the upper Qiulitag Group in the Tarim Basin (Gao Citation1991, Zhao et al. Citation2000). Similar to Serratognathus, both species of Serratognathoides may have a trimembrate apparatus including symmetrical Sa (e.g. Zhao et al. Citation2000, pl. 28, ), asymmetrical Sb (e.g. Zhao et al. Citation2000, pl. 28, ) and strongly asymmetrical Sc (e.g. Zhao et al. Citation2000, pl. 28, fig. 15) elements.
Serratognathoides? sp. (–C)
2000 Gen. et sp. nov. Zhao et al., pp. 232, 282, pl. 28, –.
Material
One specimen (AMF.135054) from sample THH10, Honghuayuan Formation (S. diversus Biozone), Honghuayuan Section, Tongzi, Guizhou.
Diagnosis
A serratognathoid species consisting of symmetrical to asymmetrical disk-like elements with a concave upper face ornamented with needle-like denticles that form radiating rows outwards from the cusp and a smooth, convex (or flat) basal face bearing radiating ridges and grooves along the basal-anterior margin; small to indistinct, conical cusp located near the posterior margin; basal cavity represented by a small, shallow pit underneath the cusp.
Description
Sa element, represented by a specimen figured by Zhao et al. (Citation2000, pl. 28, –), is symmetrical, disk-like, fan-shaped (or shell-like) in upper view with a broad anterior-lateral platform, and with a concave upper face and convex or flat basal face. Cusp small, conical, positioned nearly medially on posterior margin of the wide fan-like platform. Basal face smooth, ornamented with fine concentric striae centred around the basal cavity represented by a small, shallow pit; short radiating ridge-like denticles and grooves developed along the outer margin, particularly on the basal-anterior margin. Sb-c element resembling Sa, but asymmetrical (–C).
Remarks
Zhao et al. (Citation2000) briefly described and illustrated this species in open nomenclature, while recognizing its close relationship with Serratognathoides, from which it differs in having a thin, shell-like unit with convex or flat basal face rather than a concave basal face. A single specimen has also been recovered from the Honghuayuan Formation in Guizhou (–C), and is identical with the Tarim specimens, except that it is apparently asymmetrical.
As a rare species represented by only a single specimen in the current collection from the Honghuayuan Formation in Guizhou and two specimens from the upper subgroup of the Qiulitag Group in the Tarim Basin (Zhao et al. Citation2000), morphologically it is easily distinguished from other confirmed species of both Serratognathus and Serratognathoides. The limited material prevents current recognition as a new genus and species, and it is doubtfully attributed to Serratognathoides pending further studies.
Serratognathus Lee, 1970
Type species
Serratognathus bilobatus Lee, 1970.
Remarks
The following three species are assigned to Serratognathus: S. bilobatus Lee, Citation1970, S. diversus An, Citation1981 and S. extensus Yang in An et al. (Citation1983). Serratognathus armadillodus Pohler, Citation1994 from reworked clasts in Beds 12 and 14 of the Cow Head Group of western Newfoundland is herein re-assigned to Paraserratognathus An (in An et al. Citation1983), recently revised by Landing & Westrop (Citation2006) who considered Wandelia Smith, Citation1991 and Stultodontus Ji & Barnes, Citation1994a to be junior synonyms of Paraserratognathus. Furthermore, we regard S. armadillodus Pohler, Citation1994 as conspecific with P. ovatus (Ji & Barnes, Citation1994a), the type species of Stultodontus, from the lower part of the Catoche Formation of western Newfoundland. It was based on much more abundant and in situ material. Although both species were published in the same year, the publication date (31 March, 1994) of P. ovatus (Ji & Barnes, Citation1994a) was slightly earlier than that (1 July, 1994) of S. armadillodus Pohler, 1994. Therefore, the former should be regarded as the valid name and S. armadillodus is considered herein as a junior synonym of P. ovatus.
However, the situation is further complicated by the possibility that Eoserratognathus An & Zheng, Citation1990 is a junior synonym of Paraserratognathus. Both genera were recorded from the Qianzhongliangzi Formation in Ningxia, North China. The type species of Eoserratognathus, E. ovatus An & Zheng, Citation1990, closely resembles P. ovatus Ji & Barnes, Citation1994a, although they are not necessarily conspecific. As a consequence, E. ovatus An & Zheng, Citation1990 (the type and only species of Eoserratognathus) becomes a senior homonym of P. ovatus (Ji & Barnes, Citation1994a), and the latter should be renamed, or alternatively P. armadillodus (Pohler, Citation1994) rather than P. ovatus (Ji & Barnes, Citation1994a) should be used as the name of the Newfoundland species.
Serratognathus diversus An, Citation1981 (–M; 12A–K; 13A–K)
1981 Serratognathus sp. A; An et al., pl. 1, .
1981 Serratognathus diversus An, p. 216, pl. 2, figs 23, 27, 30.
1981 Serratognathus diversus An; Ni, pl. 1, .
1982 Serratognathus diversus An; An & Ding, pl. 5, fig. 25.
1983 Serratognathus diversus An; An et al., p. 150, pl. 17, fig. 34, pl. 33, –.
1984 Serratognathus diversus An; An & Xu, pl. 2, , 13, 16.
1985 Serratognathus diversus An; An & Ding, pl. 1, .
1985 Serratognathus diversus An; An et al., pl. 4, fig. 16, pl. 6, figs 22–23.
1987 Serratognathus diversus An; An, p. 190, pl. 18, –.
1990 Serratognathus diversus An; Duan, pl. 2, figs 18–19.
1991 Serratognathus diversus An; Gao, p. 142, pl. 6, figs 14–15.
1993 Serratognathus diversus An; Ding et al. in Wang, p. 207–208, pl. 20, figs 12–14, 16.
1993 Serratognathus diversus An; Chen & Wang, .
1995 Serratognathus diversus An; Chen X.Y. et al., p.1, fig. 21.
1996 Serratognathus diversus An; Wang et al., pl. 2, figs 10–12.
1998 Serratognathus diversus An; Wang & Zhou, pl. 2, .
2000 Serratognathus diversus An; Zhao et al., p. 225, pl. 28, figs 1–8.
2001 Serratognathus diversus An; Wang & Qi, pl. 2, –.
2007 Serratognathus diversus An; Zhen, pl. 2, –.
Material
Four hundred and seventy-three specimens from 34 samples (see ).
Diagnosis
Species of Serratognathus with a trimembrate apparatus, including symmetrical Sa, asymmetrical Sb, and strongly asymmetrical Sc elements; all elements, semi-conical in outline with fan-shaped array of small, closely spaced denticles along anterior and lateral edges of overlapping laminar layers; cusp prominent, distally compressed laterally and enclosed antero-laterally by typically 10–15 vertically overlapping laminar layers that are antero-basally divided by a broad median groove into two lobe-like lateral processes; basal cavity absent.
Description
Trimembrate apparatus, including symmetrical Sa, asymmetrical Sb, and strongly asymmetrical Sc elements forming a symmetry transition series; all elements semi-conical with up to 20 vertically overlapping layers enclosing the distinctive cusp and forming a gently, posteriorly curved, lobe-like lateral process on each side, resembling a half-cut onion; central cusp prominent, laterally compressed, particularly distally, blade-like in lateral profile; with fine upwardly overlapping microstructure best developed on the posterior face, and in the mature specimens, cusp surrounded antero-laterally by 6–20 vertically overlapping layers that are ornamented by small, closely spaced denticles on the edge; posterior face of the lateral processes ornamented by rows of denticles formed along the edge of the overlapping layers; denticulate anterior margin of each layer turned upwards; basal face wide, smooth, distally arched, and divided by a broad, median groove into two lobes; basal cavity absent, typically with a small ring-like node underneath the cusp.
Sa element () symmetrical, crescentic outline in upper view with convex anterior margin and concave posterior margin (), and tower-like in anterior view (–C), formed by the vertically overlapping layers with up-turned, denticulate edges; cusp prominent, laterally compressed (), blade-like in lateral profile (), with overlapping laminar structure observed along the posterior margin of the cusp (–J); posterior face of the unit ornamented by rows of discrete denticles surrounding the cusp in a semicircle, representing the lateral edge of each overlapping layer; fine striae developed on the posterior face (); in posterior view, two processes extend posteriorly and downward, forming an angle of about 100° or more, with an arched basal margin (, H); basal face smooth, crescentic in outline in basal view, bisected by antero-posteriorly directed median groove () dividing basal face into two symmetrical lobes, each extending under the lateral processes; no basal cavity, with basal end of the cusp represented by a small ring-like node (–C, K).
Sb element () like Sa, but asymmetrical; in posterior view (, I, H), inner lateral process extending laterally with basal margin nearly horizontal, and outer lateral process extending laterally and downward; basal margin of the two processes forming an angle of 120° or more; basal face asymmetrical, with a longer lobe under outer lateral process and a shorter lobe under inner lateral process ().
Sc element (–M) similar to Sb, but strongly asymmetrical with a longer, more strongly posteriorly extended outer lateral process; in posterior view, the basal margin of the two lateral processes forming more or less a right angle (, J).
Remarks. Serratognathus diversus differs from S. bilobatus in having a more prominent cusp that is laterally compressed distally, and by its more strongly divergent lateral processes (An et al. Citation1981, Citation1985). The holotype (BJ79-10, Peking University) of S. diversus (see An Citation1981, pl. 2, fig. 27) was collected from the upper part of the Honghuayuan Formation in the Huanghuachang Section of Yichang, Hubei, and represents the strongly asymmetrical Sc element (in our notation). However, a specimen (also a Sc element) with the same registration number as the holotype of B. diversus (BJ79-10) was also illustrated by An et al. (Citation1981, pl. 1, ) as Serratognathus sp. A. As the photograph of this specimen was taken from a different angle, it is difficult to be certain whether it represents the holotype of S. diversus.
We interpret each element as growing upwards by addition of overlapping layers surrounding the cusp antero-laterally, progressively forming a conical structure. The smooth basal face presumably was in direct contact with soft tissue of the conodont animal, with this interface representing the generation layer of new skeletal material.
Acknowledgements
Research by YYZ was funded by the CAS/SAFEA International Partnership Program for Creative Research Teams. Fieldwork by YYZ in Guizhou was undertaken with the support of the Australian Academy of Sciences and Chinese Academy of Sciences. Further collecting in Tongzi was undertaken by JBL with the support of the Special Funds for Major State Basic Research Project (G200077700) of China. Professors Zhou Zhiyi, Rong Jiayu and Chen Xu (Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing) are thanked for their logistic support during the fieldwork in China (NSFC projects 40272001, and Major Basic Research Projects of MST, China G2000077703). G. Dargan (Geological Survey of New South Wales, Sydney) assisted with acid leaching and residue separation. Scanning electron micrographs were prepared in the Electron Microscope Unit of the Australian Museum (Sydney). IGP publishes with permission of the Director, Geological Survey of NSW. J. Repetski and J. Rasmussen are thanked for their careful and constructive reviews of the manuscript. This is a contribution to IGCP Project 503: Ordovician Palaeogeography and Palaeoclimate.
Related Research Data
References
- Albanesi , G. L. , Hünicken , M. A. and Barnes , C. R. 1998 . Bioestratigrafia, biofacies y taxonomia de conodontes de las secuencias Ordovicicas del cerro porterillo, Precordillera central de San Juan, R. Argentina , 249 Córdoba, , República Argentina : Academia Nacional de Ciencias .
- An , T. X. 1981 . Recent progress in Cambrian and Ordovician conodont biostratigraphy of China . Geological Society of America Special Paper , 187 : 209 – 226 .
- An , T. X. 1987 . Early Paleozoic conodonts from south China , 238 Beijing : Peking University Publishing House . (in Chinese)
- An , T. X. and Ding , L. S. 1982 . Preliminary studies and correlations on Ordovician conodonts from the Ningzhen Mountains, China . Acta Petroleum Sinica , 3 ( 4 ) : 1 – 11 . (in Chinese)
- An , T. X. and Ding , L. S. 1985 . Ordovician conodont biostratigraphy in Hexian, Anhui Province . Geological Review , 31 ( 1 ) : 1 – 12 . (in Chinese with English abstract)
- An , T. X. and Xu , B. Z. 1984 . Ordovician System and conodonts of Tungshan and Xianning, Hubei . Acta Scientiarum Naturalium Universitatis Pekinensis , 5 : 73 – 87 . (in Chinese with English abstract)
- An , T. X. and Zheng , S. C. 1990 . The Conodonts of the Marginal Areas Around the Ordos Basin, North China , 199 Beijing : Science Press . (in Chinese with English abstract)
- An , T. X. , Du , G. Q. , Gao , Q. Q. , Chen , X. B. and Li , W. T. 1981 . “ Ordovician conodont biostratigraphy of the Huanghuachang area of Yichang, Hubei ” . In Selected Papers of the First Symposium of the Micropalaeontological Society of China , Edited by: Micropalaeontological Society of China . 105 – 113 . Beijing : Science Press . (in Chinese)
- An , T. X. , Zhang , F. , Xiang , W. D. , Zhang , Y. Q. , Xu , W. H. , Zhang , H. J. , Jiang , D. B. , Yang , C. S. , Lin , L. D. , Cui , Z. T. and Yang , X. C. 1983 . The Conodonts of North China and the Adjacent Regions , 223 Beijing : Science Press . (in Chinese with English abstract)
- An , T. X. , Du , G. Q. and Gao , Q. Q. 1985 . Ordovician Conodonts from Hubei , 64 Beijing : Geological Publishing House . (in Chinese with English abstract)
- Bagnoli , G. , Stouge , S. and Tongiorgi , M. 1988 . Acritarchs and conodonts from the Cambro–Ordovician Furuhäll (Köpingsklint) Section (Öland, Sweden) . Rivista Italiana di Paleontologia e Stratigrafia , 94 : 163 – 248 .
- Balfour , F. M. 1880–1881 . A Treatise on Comparative Embryology , Two Volumes , London : Macmillan & Co. .
- Chen , Xu and Wang , Z. H. 1993 . The base of Arenig Series in China . Newsletters on Stratigraphy , 29 : 159 – 164 .
- Chen , Xu , Rong , J. Y. , Wang , X. F. , Wang , Z. H. , Zhang , Y. D. and Zhan , R. B. 1995 . Correlation of the Ordovician rocks of China: charts and explanatory notes . International Union of Geological Sciences, Publication , 31 : 1 – 104 .
- Chen , X. Y. , Peng , M. H. and Jin , C. S. 1995 . Lower Ordovician conodonts from Tudi'ao, Yanhe County, Guizhou . Acta Micropalaeontologica Sinica , 12 : 323 – 332 . (in Chinese with English summary)
- Clark , D. L. , Sweet , W. C. , Bergström , S. M. , Klapper , G. , Austin , R. L. , Rhodes , F. H.T. , Müller , K. J. , Ziegler , W. , Lindström , M. , Miller , J. F. and Harris , A. G. 1981 . “ Conodonta ” . In Treatise on Invertebrate Paleontology, Part W, Miscellanea, Supplement 2 , Edited by: Robison , R. A. 202 Lawrence : The Geological Society of America, Boulder, and the University of Kansas .
- Cooper , B. J. 1981 . Early Ordovician conodonts from the Horn Valley Siltstone, central Australia . Palaeontology , 24 : 147 – 183 .
- Ding , L. S. . Preliminary probes into Ordovician conodont biostratigraphy from the Kunshan area, Jiangsu, China . Proceedings of Petroleum, Stratigraphy and Palaeontology 1987. Geological Publishing House . Beijing. pp. 41 – 53 . (in Chinese with English abstract)
- Dong , D. Y. and Wang , W. 2006 . The Cambrian–Triassic Conodont Faunas in Yunnan, China—Correlative Biostratigraphy and the Study of Palaeobiogeographic Province of Conodont , 347 Kunming : Yunnan Science and Technology Press . (in Chinese with English summary)
- Duan , J. Y. 1990 . Ordovician conodonts from northern Jiangsu and indices of their colour alteration . Acta Micropalaeontologica Sinica , 7 ( 1 ) : 19 – 41 . (in Chinese with English abstract)
- Dzik , J. 1994 . Conodonts of the Mójcza Limestone . Palaeontologia Polonica , 53 : 43 – 128 .
- Fåhræus , L. E. 1966 . Lower Viruan (Middle Ordovician) conodonts from the Gullhögen Quarry, southern central Sweden . Sveriges Geologiska Undersökning C , 610 : 1 – 40 .
- Furnish , W. M. 1938 . Conodonts from the Prairie du Chien (Lower Ordovician) beds of the upper Mississippi valley . Journal of Paleontology , 12 : 318 – 340 .
- Gao , D. D. 1976 . “ Guizhou de Aotaoxi[The Ordovician System of Guizhou]. Stratigraphy and Palaeontology Team ” . 104 Huishui : Geological Bureau of Guizhou . (in Chinese)
- Gao , Q. 1991 . “ Conodonts ” . In Sinian to Permian Stratigraphy and Palaeontology of the Tarim Basin II, Keping-Bachu area , Edited by: Xinjiang Petroleum Administration Bureau and the Jianghan Petroleum Administration Bureau . 125 – 149 . Beijing : Petroleum Industry Press . (in Chinese with English abstract)
- Ji , Z. L. and Barnes , C. R. 1994a . Lower Ordovician conodonts of the St. George Group, Port au Port Peninsula, western Newfoundland, Canada . Palaeontographica Canadiana , 11 : 1 – 149 .
- Ji , Z. L. and Barnes , C. R. 1994b . Conodont paleoecology of the Lower Ordovician St. George Group, Port au Port Peninsula, western Newfoundland . Journal of Paleontology , 68 : 1368 – 1383 .
- Kennedy , D. J. 1980 . A restudy of conodonts described by Branson and Mehl, 1933, from the Jefferson City Formation, Lower Ordovician, Missouri . Geologica et Palaeontologica , 14 : 45 – 76 .
- Kennedy , D. J. 1994 . Colaptoconus (Conodonta), a replacement name for Glyptoconus Kennedy, 1981, non Glyptoconus von Moellendorff, 1894 . Journal of Paleontology , 68 : 1417
- Landing , E. and Westrop , S. R. 2006 . Lower Ordovician faunas, stratigraphy, and sea level history of the middle Beekmantown Group, northeastern New York . Journal of Paleontology , 80 : 958 – 980 .
- Landing , E. , Westrop , S. R. and van Aller Hernick , L. 2003 . Uppermost Cambrian–Lower Ordovician faunas and Laurentian platform sequence stratigraphy, eastern New York and Vermont . Journal of Paleontology , 77 : 78 – 98 .
- Lee , H. Y. 1970 . Conodonten aus der Choson-Gruppe (Unteres Ordovizium) von Korea . Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen , 136 : 303 – 344 .
- Lee , H. Y. 1975 . Conodonts from the Dumugol Formation (Lower Ordovician), South Korea . Journal of the Geological Society of Korea , 11 : 75 – 98 .
- Li , Y. , Kershaw , S. and Mu , X. N. 2004 . Ordovician reef systems and settings in South China before the Late Ordovician mass extinction . Palaeogeography, Palaeoclimatology, Palaeoecology , 205 : 235 – 254 .
- Lindström , M. 1955 . Conodonts from the lowermost Ordovician strata of south-central Sweden . Geologiska Föreningens i Stockholm Förhandlingar , 76 : 517 – 604 .
- Lindström , M. 1971 . “ Lower Ordovician conodonts of Europe ” . In Symposium on Conodont Biostratigraphy , Edited by: Sweet , W. C. and Bergström , S. M. Vol. 127 , 21 – 61 . Memoir : Geological Society of America .
- Löfgren , A. 1993 . Arenig conodont successions from central Sweden . Geologiska Föreningens i Stockholm Förhandlingar , 115 : 193 – 207 .
- Löfgren , A. 1994 . Arenig (Lower Ordovician) conodonts and biozonation in the eastern Siljan District, central Sweden . Journal of Paleontology , 68 : 1350 – 1368 .
- Löfgren , A. 1997 . Conodont faunas from the upper Tremadoc at Brattefors, south-central Sweden, and reconstruction of the Paltodus apparatus . GFF , 119 : 257 – 255 .
- Löfgren , A. 1999a . “ A septimembrate apparatus model for the Ordovician conodont genus Cornuodus Fåhræus, 1966 ” . In Studies on Conodonts—Proceedings of the Seventh European Conodont Symposium, Bologna-Modena, 1998 Edited by: Serpagli , E. Bollettino della Società Paleontologica Italiana 37, 175–186
- Löfgren , A. 1999b . The Ordovician conodont Semiacontiodus cornuformis (Sergeeva, 1963) and related species in Baltoscandia . Geologica et Palaeontologica , 33 : 71 – 91 .
- Löfgren , A. and Tolmacheva , T. J. 2003 . Taxonomy and distribution of the Ordovician conodont Drepanodus arcuatus Pander, 1856, and related species . Paläontologische Zeitschrift , 77 : 203 – 221 .
- Löfgren , A. , Viira , V. and Mens , K. 2005 . Conodont biostratigraphy and sedimentary history in the upper Tremadoc at Uuga, Cape Pakri, NW Estonia . GFF , 127 : 283 – 293 .
- McTavish , R. A. 1973 . Prioniodontacean conodonts from the Emanuel Formation (Lower Ordovician) of Western Australia . Geologica et Palaeontologica , 7 : 27 – 58 .
- Metcalfe , I. 1980 . Ordovician conodonts from the Kaki Bukit area, Perlis, West Malaysia . Warta Geologi , 6 : 63 – 68 .
- Metcalfe , I. 2003 . Colour and textural alteration of Palaeozoic and Triassic conodonts from Peninsular Malaysia: implications for tectonic evolution and hydrocarbon generation . Courier Forschungsinstitut Senckenberg , 245 : 261 – 279 .
- Miller , J. F. 1969 . Conodont fauna of the Notch Peak Limestone (Cambro-Ordovician), House Range, Utah . Journal of Paleontology , 43 : 413 – 439 .
- Ni , S. Z. 1981 . “ Discussion on some problems of Ordovician stratigraphy by means of conodonts in eastern part of Yangtze Gorges Region ” . In Selected Papers on the 1st Convention of Micropalaeontological Society of China , Edited by: Micropalaeontological Society of China . 127 – 134 . Beijing : Science Press . (in Chinese)
- Ni , S. Z. and Li , Z. H. 1987 . “ Conodonts ” . In Biostratigraphy of the Yangtze Gorge Area 2: Early Palaeozoic Era , Edited by: Wang , X. F. , Ni , S. Z. , Zeng , Q. L. , Xu , G. H. , Zhou , T. M. , Li , Z. H. , Xiang , L. W. and Lai , C. G. 386 – 447 . Beijing : Geological Publishing House . 549–555, 619–632, pls 52–62 (in Chinese with English abstract)
- Nicoll , R. S. 1990 . The genus Cordylodus and a latest Cambrian–earliest Ordovician conodont biostratigraphy . BMR Journal of Australian Geology & Geophysics , 11 : 529 – 558 .
- Nicoll , R. S. 1992 . Analysis of conodont apparatus organisation and the genus Jumudontus (Conodonta), a coniform–pectiniform apparatus structure from the Early Ordovician . BMR Journal of Australian Geology & Geophysics , 13 : 213 – 228 .
- Nicoll , R. S. 2006 . Cambrian and Ordovician sediments and biostratigraphy of the Arafura Basin, offshore Northern Territory, Australia . Geoscience Australia Record , 2006/06 : 1 – 16 . (unpublished)
- Nicoll , R. S. and Ethington , R. L. 2004 . Lissoepikodus nudus gen. et sp. nov. and Oepikodus cleftus sp. nov., new septimembrate conodont taxa from the Early Ordovician of Australia and Nevada . Courier Forschungsinstitut Senckenberg , 245 : 427 – 461 .
- Nicoll , R. S. and Metcalfe , I. 2001 . “ Cambrian to Permian conodont biogeography in East Asia-Australasia ” . In Faunal and Floral Migrations and Evolution in SE Asia–Australasia , Edited by: Metcalfe , I. , Smith , J. M.B. , Morwood , M. and Davidson , I. 59 – 72 . Lisse : Balkema .
- Nicoll , R. S. , Zhen , Y. Y. and Laurie , J. in prep. Cambrian and Ordovician stratigraphy and biostratigraphy of the Arafura Basin, offshore Northern Territory . The Association of Australasian Palaeontologists, Memoir ,
- Pander , C. H. 1856 . Monographie der fossilen Fische des Silurischen Systems der Russisch-Baltischen Gouvernements , 91 St. Petersburg : Akademie der Wissenschaften .
- Pohler , S. M.L. 1994 . Conodont biofacies of Lower to lower Middle Ordovician megaconglomerates, Cow Head Group, Western Newfoundland . Geological Survey of Canada, Bulletin , 459 : 1 – 71 .
- Rasmussen , J. A. 2001 . Conodont biostratigraphy and taxonomy of the Ordovician shelf margin deposits in the Scandinavian Caledonides . Fossils and Strata , 48 : 1 – 180 .
- Sergeeva , S. P. 1963 . Conodonts from the Lower Ordovician of the Leningrad region . Paleontologicheskiy Zhurnal, Akademiya Nauk SSSR , 2 : 93 – 108 .
- Serpagli , E. 1974 . Lower Ordovician conodonts from Precordilleran Argentina (Province of San Juan) . Bollettino della Società Paleontologica Italiana , 13 : 17 – 98 .
- Smith , M. P. 1991 . Early Ordovician conodonts of East and North Greenland . Meddelelser om Gr⊘nland, Geoscience , 26 : 1 – 81 .
- Stouge , S. and Bagnoli , G. 1988 . Early Ordovician conodonts from Cow Head Peninsula, western Newfoundland . Palaeontographica Italica , 75 : 89 – 179 .
- Sweet , W. C. 1988 . The Conodonta: Morphology, Taxonomy, Paleoecology, and Evolutionary History of a Long-Extinct Animal Phylum , 212 Oxford : Clarendon Press .
- Tolmacheva , T. Y. 2006 . Apparatus of the conodont Scolopodus striatus Pander, 1856 and a re-evaluation of Pander's species of Scolopodus . Acta Palaeontologica Polonica , 51 : 247 – 260 .
- Tolmacheva , T. Y. and Fedorov , P. 2001 . The Ordovician Billingen/Volkhov boundary interval (Arenig) at Lava River, northwestern Russia . Norsk Geologisk Tidsskrift , 81 : 161 – 168 .
- Tolmacheva , T. Y. , Koren , T. N. , Holmer , L. E. , Popov , L. E. and Raevskaya , E. 2001 . The Hunneberg Stage (Ordovician) in the area east of St. Petersburg, north-western Russia . Paläontologische Zeitschrift , 74 : 543 – 561 .
- van Wamel , W. A. 1974 . Conodont biostratigraphy of the Upper Cambrian and Lower Ordovician of north-western Öland, south-eastern Sweden . Utrecht Micropalaeontological Bulletins , 10 : 1 – 125 .
- Viira , V. , Löfgren , A. , Mägi , S. and Wickström , J. 2001 . An Early to Middle Ordovician succession of conodont faunas at Mäekalda, northern Estonia . Geological Magazine , 138 : 699 – 718 .
- Wang C. Y. Conodonts of the Lower Yangtze Valley—An Index to Biostratigraphy and Organic Metamorphic Maturity Science Press Beijing 1993 326 (in Chinese with English summary)
- Wang , Z. H. and Qi , Y. P. 2001 . Ordovician conodonts from drillings in the Taklamakan Desert, Xinjiang, NW China . Acta Micropalaeontologica Sinica , 18 : 133 – 148 .
- Wang , Z. H. and Zhou , T. R. 1998 . Ordovician conodonts from western and northeastern Tarim and their significance . Acta Palaeontologica Sinica , 37 : 173 – 193 .
- Wang , Z. H. , Bergström , M. and Lane , H. R. 1996 . Conodont provinces and biostratigraphy in Ordovician of China . Acta Palaeontologica Sinica , 35 : 26 – 59 .
- Zeng , Q. L. , Ni , S. Z. , Xu , G. H. , Zhou , T. M. , Wang , X. F. , Li , Z. H. , Lai , C. G. and Xiang , L. W. 1983 . Subdivision and correlation on the Ordovician in the eastern Yangtze Gorges, China . Bulletin of the Yichang Institute of Geology & Mineral Resources, Chinese Academy of Geological Sciences , 6 : 1 – 68 .
- Zhan , R. B. and Jin , J. S. 2007 . Ordovician–Early Silurian (Llandovery) Stratigraphy and Palaeontology of the Upper Yangtze Platform, South China , 169 Beijing : Science Press .
- Zhang , S. X. and Zhen , Y. Y. 1991 . “ Chapter 5, China ” . In The Phanerozoic Geology of the World I, the Palaeozoic, A , Edited by: Moullade , M. and Nairn , A. E.M. 219 – 274 . Elsevier : Amsterdam .
- Zhao , Z. X. , Zhang , G. Z. and Xiao , J. N. 2000 . Paleozoic stratigraphy and conodonts in Xinjiang , 340 Beijing : Petroleum Industry Press . (in Chinese with English abstract)
- Zhen , Y. Y. 2007 . Conodont biostratigraphy of the Honghuayuan Formation (late Early Ordovician) in Guizhou, South China . Acta Palaeontologica Sinica , 46 ( Supplement ) : 537 – 542 .
- Zhen , Y. Y. and Nicoll , R. S. 2009 . Biogeographic and biostratigraphic implications of the Serratognathus bilobatus fauna (Conodonta) from the Emanuel Formation (Early Ordovician) of the Canning Basin, Western Australia . Records of the Australian Museum , 61 : 1 – 30 .
- Zhen , Y. Y. and Percival , I. G. 2003 . Ordovician conodont biogeography—reconsidered . Lethaia , 36 : 357 – 369 .
- Zhen , Y. Y. and Percival , I. G. 2004 . Middle Ordovician (Darriwilian) conodonts from allochthonous limestones in the Oakdale Formation of central New South Wales, Australia . Alcheringa , 28 : 77 – 111 .
- Zhen , Y. Y. and Percival , I. G. 2006 . Late Cambrian–Early Ordovician conodont faunas from the Koonenberry Belt of western New South Wales, Australia . Association of Australasian Palaeontologists Memoir , 32 : 267 – 285 .
- Zhen , Y. Y. , Percival , I. G. and Webby , B. D. 2003 . Early Ordovician conodonts from far western New South Wales, Australia . Records of the Australian Museum , 55 : 169 – 220 .
- Zhen , Y. Y. , Percival , I. G. and Webby , B. D. 2004 . Early Ordovician (Bendigonian) conodonts from central New South Wales, Australia . Courier Forschungsinstitut Senckenberg , 245 : 39 – 73 .
- Zhen , Y. Y. , Liu , J. B. and Percival , I. G. 2005 . Revision of two prioniodontid species (Conodonta) from the Early Ordovician Honghuayuan Formation of Guizhou, South China . Records of the Australian Museum , 57 : 303 – 320 .
- Zhen , Y. Y. , Percival , I. G. and Liu , J. B. 2006a . Early Ordovician Triangulodus (Conodonta) from the Honghuayuan Formation of Guizhou, South China . Alcheringa , 30 : 191 – 212 .
- Zhen , Y. Y. , Percival , I. G. and Liu , J. B. 2006b . Rhipidognathid conodonts from the Early Ordovician Honghuayuan Formation of Guizhou, South China . Palaeoworld , 15 : 194 – 210 .
- Zhen , Y. Y. , Liu , J. B. and Percival , I. G. 2007a . Revision of the conodont Erraticodon hexianensis from the upper Meitan Formation (Middle Ordovician) of Guizhou, South China . Palaeontological Research , 11 : 145 – 162 .
- Zhen , Y. Y. , Percival , I. G. , Löfgren , A. and Liu , J. B. 2007b . Drepanoistodontid conodonts from the Early Ordovician Honghuayuan Formation of Guizhou, South China . Acta Micropalaeontologica Sinica , 24 : 125 – 148 .
- Zhen , Y. Y. , Zhang , Y. D. and Percival , I. G. 2009 . Early Sandbian (Late Ordovician) conodonts from the Yenwashan Formation, western Zhejiang, South China . Alcheringa , 33 : 133 – 161 .
- Zhen , Y.Y. , Percival , I.G. , Cooper , R.A. , Simes , J.E. and Wright , A.J. in press . Darriwilian (Middle Ordovician) conodonts from Thompson Creek, northwest Nelson, New Zealand . Association of Australasian Palaeontologists, Memoir 37 , 27–55
- Zhou , Z. Y. , Zhen , Y. Y. , Zhou , Z. Q. and Yuan , W. W. 2007 . A new approach to the division of Ordovician geographic units of China . Acta Palaeontologica Sinica , 46 ( Supplement ) : 558 – 563 .
- Zhu , Z. D. , Jiang , Y. W. and Liu , B. L. 1995 . “ Palaeoecology of late Tremadocian reef-bearing strata in western Hubei province of China ” . In Ordovician Odyssey: Short Papers for the Seventh International Symposium on the Ordovician System , Edited by: Cooper , Droser J.M.D.L. and Finney , S. C. 427 – 428 . Fullerton, California : SEPM Pacific Section, Book 76 .
- Ziegler W. Catalogue of Conodonts I E. Schweizerbart'sche Stuttgart 1974 504