Abstract
West Yunnan in southwestern China formed an important part of the Sibumasu Terrane during the Ordovician. The Ordovician rocks of the region are exposed at many localities but have been strongly affected by several phases of tectonic activity, especially by the most recent Himalayan phase that commenced in the Paleogene. For these reasons, together with the difficult access to the localities, the Ordovician graptolites and biostratigraphy of this region are rather poorly known. Herein, we describe a Darriwilian graptolite fauna from the Shihtien Formation at Baoshan and Shidian, western Yunnan. The fauna comprises 11 genera and 15 species, including Didymograptus artus, D. murchisoni, D. spinulosus, Pterograptus sp., Hustedograptus vikarbyensis, H. teretiusculus, Archiclimacograptus angulatus, A. riddellensis, Haddingograptus oliveri, Proclimacograptus angustatus, and the dendroids Dictyonema, Palaeodictyota, Dendrograptus, Callograptus and Ptilograptus, indicating an age of middle to late Darriwilian. Two graptolite biozones are recognized: the Didymograptus artus and Didymograptus murchisoni biozones, in ascending order. The graptolite fauna is substantially similar to those of Baltica and South China.

YUNNAN PROVINCE in southwestern China encompasses three distinct geological domains (). The east Yunnan region, bounded to the west by the Ailaoshan Mts (China)–Song Da (Viet Nam) suture, has been interpreted as belonging to the South China Palaeoplate in the Ordovician, and the marine faunas display a great uniformity across this terrane (Chen & Rong Citation1992). The central part of Yunnan has been widely regarded as a part of the Indo-China Block (Annamia), although the Ordovician faunas of the region are poorly known (Zhou et al. Citation1998). The west Yunnan region has been widely regarded as a constituent of the Sibumasu Terrane (also called the Shan-Thai Terrane or Palaeoplate) during the Ordovician (Chen & Rong Citation1992, Fortey & Cocks Citation1998, Citation2003, Metcalfe Citation1998). The Sibumasu Terrane embraces west Yunnan, East Burma (mainly Shan State), most of the Malay Peninsula and Sumatra, and is bounded to the east within Yunnan by the Changning-Menglian suture (Fang et al. Citation1992, Liu et al. Citation1993, Metcalfe Citation1998). The biogeographic affinities of the Sibumasu Terrane with other blocks or continents, such as South China, North China, and Gondwana, are still disputed owing to the poorly surveyed fossil faunas in Sibumasu. Fortey & Cocks (Citation1998, Citation2003) concluded that the carbonate succession of the Cambrian–Ordovician transition interval in the terrane displays strong similarities to those in North China and central Australia, whereas the younger Ordovician trilobite faunas are nearly identical to those of South China, despite the fact that the Caradoc brachiopods from Burma show a modest degree of endemism (Cocks & Zhan Citation1998, Zhan & Cocks Citation1998). Zhou & Zhen (Citation2008) proposed that, during the Ordovician, the Shan State (Burma)–Baoshan (west Yunnan) region was in low latitudes and not far from South China, whereas the West Malaysia–Thailand Peninsula block was located close to the North China Plate on the palaeo-equator. Here we report Darriwillian graptolites from west Yunnan that, together with recent studies of other groups (e.g. Popov et al. 2008, Fang & Cope 2008), enhance understanding of the biogeographic relationships between Sibumasu and surrounding areas during the Ordovician.
Fig. 1. Location of the Laojianshan section, Baoshan and the Banpo section, Shidian, west Yunnan, China. A, Location of the study area (triangle) and the Sibumasu Block on a reconstruction map of the Southern Hemisphere Ordovician (see Fortey & Cocks Citation2003 for abbreviations). B, Location and accessibility of the Laojianshan (1) and Banpo (2) sections in west Yunnan, China.
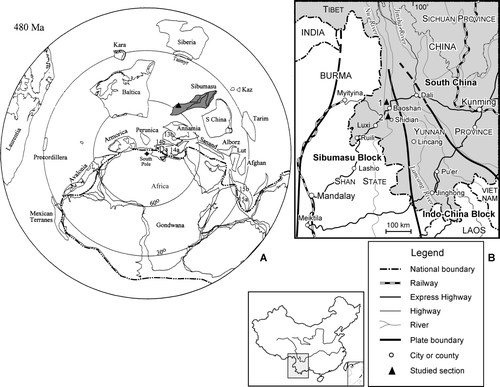
The Ordovician rocks of west Yunnan have been poorly documented to date. The earliest geological work in west Yunnan was carried out by Loczy in 1880 (results published by Loczy 1893) and Brown in 1908–1910. Loczy first reported Ordovician trilobites in the region and Brown published a preliminary introduction to the Ordovician and Silurian rocks of the region (Brown Citation1913, Citation1916). The abundant fossils collected by Brown, including cystoids, crinoids, brachiopods, gastropods, cephalopods, trilobites and graptolites, were published by Reed (Citation1917). The graptolites in Brown's collections, all from Silurian rocks, were identified by G.L. Elles to be of Llandovery age. Yin & Lu (1936–1937) collected fossils from, and described, the Jenhochiao section (now called the Banpo section) of Shihtien (now called Shidian). They reported the discovery of an Ordovician graptolite fauna including specimens they identified as Didymograptus nanus Lapworth in Hopkinson & Lapworth, 1875, Didymograptus sp. indet., Dictyonema sp. and Desmograptus? sp. (original identifications), together with some brachiopods, trilobites and cephalopods. The graptolites were described and illustrated briefly by Yin (Citation1937). A subsequent investigation of the Baoshan region, western Yunnan by Sun in 1938 led to a subdivision of the Ordovician rocks into three formations (Sun Citation1945; ): the Shihtien Formation [Arenig and Llanvirn, in which Didymograptus nanus and Didymograptus murchisoni (Beck, 1839) were reported], lower Hungshuitang Formation (Llandeilo) and upper Hungshuitang Formation (Caradoc). Yu (Citation1962) described some additional graptolite species, which he assigned to Ptilograptus Hall, 1865 and a new genus Zigzagigraptus Yu, 1962. Herein, we describe a much more diverse Ordovician graptolite fauna from the Laojianshan section in Baoshan and the Banpo section in Shidian (). These graptolite specimens were largely compressed, but without significant tectonic distortions. Some specimens are pyritic and partially three-dimensional, with most details of the proximal structures and thecae available.
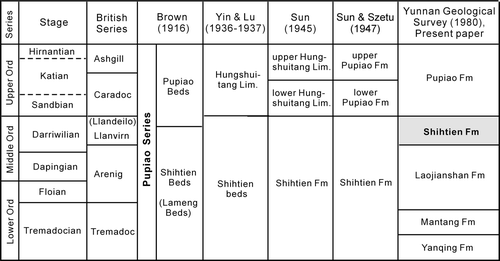
Fig. 2. Historical development of the lithostratigraphic scheme for the Ordovician rocks in west Yunnan.
Laojianshan section
The Laojianshan section is located at Laojianshan Mountain near the Upper Yanqing village, ca 17 km northwest of Baoshan city centre (). The section, exposed along the road from Zhugeyin Town to Yangliu Town, includes a complete sequence of Ordovician rocks (; Yunnan Geological Survey Citation1980). In the Shihtien Formation, we discovered a graptolite fauna in association with some shelly fossils. The Shihtien Formation conformably overlies the Laojianshan Formation and conformably underlies the Pupiao Formation. It consists of greenish grey and green-yellow siltstones and shales, with a few layers of thin-bedded micrites, containing fairly abundant graptolites, trilobites and brachiopods. The formation was derived from the original ‘Shihtien Beds’ of Brown (Citation1916), which was redefined by Yin & Lu (1936) to include the Shihtien Beds and the basal graptolite-bearing part of the overlying Pupiao Beds (the rest of the Pupiao Beds was called Hungshuitang Limestone). The ‘Shihtien bed’ was further subdivided into four formations (), whereas the ‘Hungshuitang Limestone’ was renamed the Pupiao Formation (Yunnan Geological Survey Citation1980). The Pupiao Formation is typified by purple-red and greenish grey siltstones, shales and micrites, with its base defined by the first appearance of purple-red layers. It is overlain by the black shales and cherts of the Llandovery Jenhochiao Formation.
Banpo section
The Banpo section is located at Banpo village near Renhoqiao (formerly known as Jenhochiao) Town, ca 6 km to the north of Shidian County town, and ca 60 km south of Baoshan city centre (). The section includes the Shihtien, Pupiao and Jenhochiao formations in ascending order, but the succession is somewhat tectonically distorted. We discovered a graptolite fauna in the upper part of the Shihtien Formation.
Biostratigraphy
The graptolites collected by Brown in his pioneering expedition were identified by Elles as including Didymograptus murchisoni, D. murchisoni geminus (Hisinger, 1840), D. indentus (Hall, 1858) and Climacograptus sp. cf. C. scharenbergi (Lapworth, 1876), which suggested an age equivalent to the Didymograptus murchisoni Biozone (Brown Citation1916, Reed Citation1917). The rocks bearing these graptolites were assigned to the basal part of the ‘Pupiao Beds’ by Brown, but later included into the upper part of the redefined ‘Shihtien Formation’ by Yin & Lu (Citation1936) who found additional Didymograptus nanus and some dendroids indicating an earliest Llanvirn age of the ‘Didymograptus bifidus’ Biozone (=D. artus Biozone). Our collection includes a more diverse graptolite fauna, including Didymograptus artus Elles & Wood, Citation1901, D. murchisoni, D. spinulosus Perner, 1895, Pterograptus sp., Hustedograptus vikarbyensis (Jaanusson, Citation1960), H. teretiusculus (Hisinger, 1840), Archiclimacograptus angulatus (Bulman, Citation1953), A. riddellensis (Harris, Citation1924), Haddingograptus oliveri (Bouček, Citation1973), Proclimacograptus angustatus (Ekström, Citation1937), and the dendroids Dictyonema Hall, 1851, Palaeodictyota Whitfield, Citation1902, Dendrograptus Hall, 1858, Callograptus Hall, 1865 and Ptilograptus Hall, 1865. Based on the stratigraphic distribution of the species, two graptolite biozones are recognized herein, the Didymograptus artus Biozone and the Didymograptus murchisoni Biozone in ascending order.
Didymograptus artus Biozone
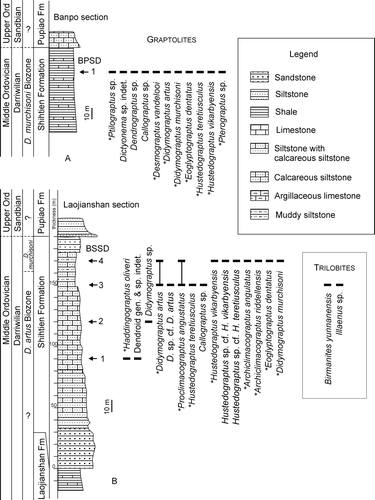
This biozone is recognized by the co-occurrences of the eponymous species, together with Haddingograptus oliveri, Proclimacograptus angustatus, Hustedograptus teretiusculus, some other uncertain species of Didymograptus, and some dendroids (in some samples, the zonal index may be absent but other elements characteristic of the assemblage are present; ). These graptolites occur mainly in the upper part of the Shihtien Formation, whereas the lower part comprising sandstones and calcareous sandstones is barren of graptolites. However, Undulograptus austrodentatus (Harris & Keble, Citation1932) was found in the top part of the underlying Laojianshan Formation in some sections near Shidian (Yunnan Geological Survey Citation1980), hence the lower part of the Shihtien Formation possibly corresponds to the transition interval between the U. austrodentatus Biozone and the D. artus Biozone, and is probably equivalent to the Undulograptus intersitus Biozone in the Yangtze Region of South China (Zhang & Chen Citation2003, Zhang et al. Citation2007) and Australasia (VandenBerg & Cooper Citation1992), and the Undulograptus dentatus Biozone in North America (Maletz Citation2005).
Fig. 3. Lithological logs of the Banpo (A) and Laojianshan (B) sections, west Yunnan, showing the distribution of Darriwilian graptolites in the sampled horizons. The asterisks (∗) indicate the species described in the present paper.
Equivalent assemblages of the D. artus Biozone are recognized in Changning, Sichuan, South China (Zhang et al. Citation2007) and in the British D. artus Biozone (Fortey et al. Citation1995, Citation2000). The D. artus Biozone of west Yunnan may also correspond to the Nicholsonograptus fasciculatus Biozone and the lower part of Pterograptus elegans Biozone in the Jiangnan Region of South China (Zhang et al. Citation2007) and in Baltica (Maletz Citation1997).
Didymograptus murchisoni Biozone
This biozone is characterized by the abundance of the eponymous species, together with common biserial graptolites including Didymograptus artus, Proclimacograptus angustatus, Hustedograptus teretiusculus, H. vikarbyensis, Archiclimacograptus riddellensis, A. angulatus, Eoglyptograptus dentatus, and some dendroids. Didymograptus artus and Proclimacograptus angustatus first appear in the underlying D. artus Biozone and persist in this biozone. Hustedograptus teretiusculus is fairly common in this biozone, but the species is long-ranging in both Baltica and South China; hence it is biostratigraphically less significant.
Among the biserial graptolites of this biozone, Hustedograptus vikarbyensis, Archiclimacograptus riddellensis and A. angulatus, together with Eoglyptograptus dentatus (sensu Bulman Citation1963), first appear in the upper Pterograptus elegans Biozone of Norway, Baltica province (Maletz Citation1997). The D. murchisoni Biozone is also recognized in Britain (Cooper et al. Citation1995, Fortey et al. Citation2000), and correlates with the lower part of the Archiclimacograptus riddellensis Biozone of Australia (VandenBerg & Cooper Citation1992), and the Pterograptus elegans Biozone of Argentina (Brussa et al. Citation2003a, b).
No graptolites have been found from the uppermost Shihtien Formation or the entire Pupiao Formation in the region, hence no graptolite zones younger than the Didymograptus murchisoni Biozone can be identified.
Biogeography
The graptolite fauna of the Shihtien Formation is very similar to that of Baltica (Berry Citation1964, Maletz Citation1997) and the Yangtze region of South China (Zhang et al. Citation2007), but has a lower diversity (). All the Shihtien Formation species (except the dendroids) occur in either or both the Yangtze or Baltica regions. Most of the common species are represented in each of these regions (e.g. Didymograptus artus, D. murchisoni, Hustedograptus vikarbyensis, H. teretiusculus, Archiclimacograptus riddellensis, Proclimacogaptus angustatus and species of Pterograptus). The other species of the Shihtien Formation are shared with either the Yangtze or Baltica regions. However, quite a few species occurring in either the Yangtze or Baltica regions, such as Didymograptus conflectus, D. stabilis, D. changningensis, D. nanus, Hustedograptus denticulatus, H. dashabensis, Archiclimacograptus caelatus, Haddingograptus scharenbergi, and Acrograptus, Expansograptus, Xiphograptus, and Normalograptus species, have not been found in west Yunnan, indicating a much lower graptolite diversity in the latter region (albeit with less extensive sampling). The graptolite fauna of the Shihtien Formation indicates that west Yunnan was biogeographically closely related to South China and Baltica, and represented a shallow-water environment in the Darriwilian. The Darriwilian trilobites from Baoshan of west Yunnan also suggest a close biogeographic and physical proximity to South China (Fortey & Cocks Citation1998, Citation2003, Zhou & Zhen Citation2008, p. 185).
Table 1. Occurrences of graptolite taxa in the Didymograptus artus and Didymograptus murchisoni biozones, or the equivalent horizons in west Yunnan and some other major regions
The graptolite species of the Shihtien Formation have also been recorded in the Jiangnan region of South China, which yields a much more diverse fauna including many deep-water forms (Zhang et al. Citation2007). Several species are shared with the equivalent rocks of Great Britain (e.g. Didymograptus artus, D. murchisoni, D. spinulosus, Archiclimacograptus angulatus, Eoglyptograptus dentatus and Proclimacograptus angustatus), but some other taxa including Pterograptus, Hustedograptus vikarbyensis, H. teretiusculus, Archiclimacograptus riddellensis and Haddingograptus oliveri are absent in the latter, indicating a relatively weak biogeographic link between these regions.
Some graptolite species of the Shihtien Formation fauna (e.g. Didymograptus artus, D. spinulosus and Proclimacograptus angustatus) were also recorded in northern Turkey, which is considered to have been situated in the vicinity of central European terranes along the western margin of Gondwana (Dean et al. Citation2000). However, the other species, especially biserial graptolites such as Archiclimacograptus, Hustedograptus and Haddingograptus, have not been reported in the latter region. Both these regions have been interpreted to have lain high in southern latitudes, but with a considerable separation (see also Fortey & Cocks Citation2003).
Systematic palaeontology
Order DENDROIDEA Nicholson, 1872
Family PTILOGRAPTIDAE Hopkinson in Hopkinson & Lapworth, 1875
Ptilograptus Hall, 1865
Type species
Ptilograptus plumosus Miller, 1889
Ptilograptus poctai Ruedemann, Citation1908 ()
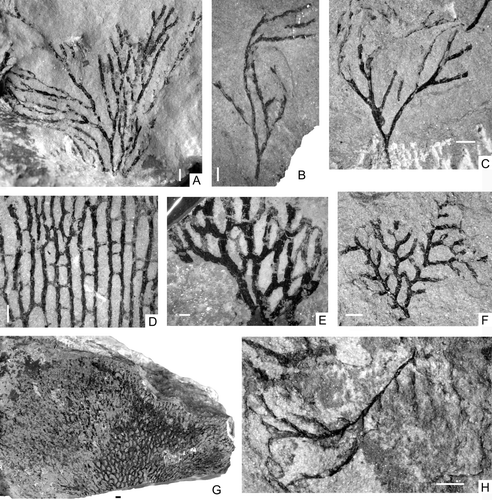
Fig. 4. Darriwilian graptolites from west Yunnan, China. A, Callograptus sp. NIGP149232 (BPSD-1-48). B–C, Dendrograptus sp., B, NIGP149233 (BSSD-1-3), C, NIGP149234 (BPSD-1-27). D, Dictyonema sp., NIGP149235 (BPSD-1-45). E, Palaeodictyota sp., NIGP149236 (BPSD-1-40). F, Ptilograptus poctai Ruedemann, NIGP149237 (BPSD-1-9). G, Desmograptus vandelooi Ruedemann, NIGP149238 (BPSD-1-42). H, Pterograptus sp., NIGP149239 (BPSD-1-25). All scale bars=1 mm.
1908 Ptilograptus poctai Ruedemann, pp. 148–149, text-fig. 53, pl. 1, .
1947 Ptilograptus poctai Ruedemann; Ruedemann, p. 247, pl. 34, figs 15, 16.
1962 Zigzagigraptus yunnanensis Yu, pp. 50–51, pl. 1, –.
Material
Two flat and incomplete specimens, one of which is fairly well preserved and contains four branches (), whereas the other one is a broken branch. The specimens are from the Didymograptus murchisoni Biozone, Shihtien Formation at the Banpo section, Shidian, Yunnan.
Description. Rhabdosome small; with the preserved part roughly 7.2 mm wide, containing four main branches of three orders. The proximal part is not preserved. The four main branches (stems) are all zigzagged, bifurcating alternately and regularly. Bifurcation angles are uniformly 70–90°, most commonly 75–85°. The distances between adjacent bifurcations are approximately 0.5 mm throughout. The main and lateral branches are uniformly 0.2 mm in lateral width. The branches derived from the same side of the main branch are parallel to each other, and a few of the lateral branches bifurcate in the same way, to become a main branch. No detailed thecal features are available.
Remarks
The present specimens match the types from North America in the general morphology of the rhabdosome, especially the branching style, but are preserved with more branches of more orders. The branches are slightly thinner than in the types (0.2 mm vs 0.3 mm), but this difference is regarded as intraspecific variation. No details of thecal morphologies are evident in the present specimens; hence a comparison with that of the types is not possible. The types were reported in association with Dicranograptus diapason, and are thus of early Sandbian age, whereas the present specimens are from the D. murchisoni Biozone of the Darriwilian, a slightly older age.
Family DENDROGRAPTIDAE Roemer in Frech, 1897
Desmograptus Hopkinson in Hopkinson & Lapworth, 1875
Type species
Dictyograptus cancellatus Hopkinson in Hopkinson & Lapworth, 1875
Desmograptus vandelooi Ruedemann, Citation1947 ()
1908 Desmograptus vandelooi Ruedemann, pp. 181–182, pl. 5, , text-figs 87, 88.
1947 Desmograptus vandelooi Ruedemann; Ruedemann, p. 225, pl. 23, figs 14–16.
Material
Only one incomplete, flattened specimen without proximal part is available (), from the Didymograptus murchisoni Biozone, Shihtien Formation at the Banpo section, Shidian, Yunnan.
Description
Rhabdosome is possibly flabellate, consisting of evidently flexuous stipes, 47 mm wide and 30 mm wide. The flexuous stipes are uniformly 0.3–0.5 mm wide, connected by largely regular anastomoses and rare dissepiments. The formed meshes are elliptical or irregularly rectangular, approximately 0.7 mm wide. There are 10–12 branches per 10 mm transversely. Details of the stipes and thecae are not available.
Order GRAPTOLOIDEA Lapworth in Hopkinson & Lapworth, 1875
Family DICHOGRAPTIDAE Lapworth, 1873
Pterograptus Holm, 1881
Type species
Pterograptus elegans Holm, 1881
Pterograptus sp. (; )
Material
Only one incomplete specimen (, ), with two main stipes and alternating lateral branches is available from the Didymograptus murchisoni Biozone, Shihtien Formation at the Banpo section, Shidian, Yunnan.
Description
The preserved rhabdosome is 8.8 mm long, containing two main stipes. The proximal part is indistinctly preserved: the sicula and proximal end are indistinguishable. The longer main stipe exceeds 6.5 mm and is nearly straight; the distal part is curved dorsally due to compression. Lateral branches are derived regularly (0.45–0.5 mm apart) and alternatively from the main stipe at 50°. The stipes are slender proximally and slightly wider distally (0.15–0.2 mm wide on average). The details of thecae are not available.
Didymograptus M'Coy in Sedgwick & M'Coy, 1851
Type species
Graptolithus murchisoni Beck in Murchison, 1839
Didymograptus artus Elles & Wood, Citation1901 (, R–T; 6A–H)
Fig. 5. Camera-lucida illustrations of the Darriwilian graptolites from the Shihtien Formation in west Yunnan. A–B, Archiclimacograptus riddellensis (Harris), A, NIGP149274 (BSSD-4-8), B, NIGP 149275 (BSSD-4-24). C–D, Archiclimacograptus angulatus (Bulman), C, NIGP 149279 (BSSD-4-21a), D, NIGP 149277 (BSSD-4-21b). E–F, L, Hustedograptus vikarbyensis (Jaanusson), E, NIGP 149261 (BPSD-1-19), F, NIGP 149259 (BSSD-4-38), L, NIGP149260 (BSSD-4-58). G–H, M, Hustedograptus teretiusculus (Hisinger), G, NIGP149272 (BPSD-1-46), H, NIGP149270 (BPSD-1-55), M, NIGP149273 (BSSD-4-37). I, K, N, Q, Didymograptus murchisoni (Beck), I, proximal part of mature specimen, NIGP149281 (BPSD-1-32), K, proximal part of mature specimen with developed membrane, NIGP149282 (BPSD-1-15), N, proximal part of mature specimen with typical pakrianus-like membrane, NIGP149256 (BPSD-1-18), Q, NIGP149255 (BSSD-4-40). J, Pterograptus sp. NIGP149239 (BPSD-1-25). O, R–T, Didymograptus artus Elles & Wood, O, proximal part of the specimen showing the origination of th21 from th11, NIGP149283 (BSSD-3-24), R, NIGP149246 (BSSD-4-8), S, NIGP149284 (BSSD-4-1), T, Juvenile specimen showing slender proximal part, NIGP149240 (BPSD-1-3). P, Eoglyptograptus dentatus (Brongniart), NIGP149264 (BSSD-4-21). All scale bars=1 mm.

1901 Didymograptus artus Elles & Wood, pp. 48–49, pl. 4, –d, text-fig. 30.
Fig. 6. Darriwilian Didymograptus from the Shihtien Formation in west Yunnan, China. A–H, Didymograptus artus Elles & Wood, A, Juvenile specimen showing the artus-type proximal end, NIGP149240 (BPSD-1-3). B, NIGP149241 (BSSD-3-15). C, NIGP149242 (BSSD-3-19). D, Two slightly distorted specimens on the same slab, NIGP 149243 (upper left), 149244 (BSSD-4-11). E, Two specimens on the same slab, NIGP149245, 149246 (BSSD-4-8). F, NIGP149247 (BSSD-4-14). G, NIGP149248 (BSSD-4-15). H, NIGP149249 (BSSD-4-12). I, Didymograptus spinulosus Perner, NIGP 149250 (BPSD-4-42). All scale bars=1 mm.

1973 Didymograptus artus Elles & Wood; Skevington, pp. 46–47, text-.
1973 Didymograptus artus Elles & Wood; Bouček, pp. 85–87, pl. 15, , text-fig. 26e–g.
Fig. 7. Darriwilian graptolites from the Shihtien Formation in west Yunnan, China. A–E, Didymograptus murchisoni (Beck), A, NIGP149251 (BPSD-1-2), B, NIGP149252 (BPSD-1-8), C, NIGP149253, 149254 (BPSD-1-10), D, mature specimen with partially developed proximal membrane, NIGP149255 (BSSD-4-40), E, NIGP149256 (BPSD-1-18). F, Proclimacograptus angustatus (Ekström), NIGP149257 (BSSD-4-52). G–H, L–N, Hustedograptus vikarbyensis (Jaanusson), G, NIGP149258 (BSSD-4-3), H, NIGP149259 (BSSD-4-38), L, NIGP149260 (BSSD-4-58), M, NIGP149261 (BPSD-1-19), N, NIGP149262 (BSSD-4-36). I–J, Eoglyptograptus dentatus (Brongniart), I, NIGP149263 (BPSD-1-9), J, NIGP149264 (BSSD-4-21). K, P, Haddingograptus oliveri (Bouček), K, enlargement of P, NIGP149265 (BSSD-1-2). O, Archiclimacograptus angulatus (Bulman), NIGP149266 (BSSD-4-6). All scale bars=1 mm.

1973 Didymograptus cf. artus Elles & Wood; Bouček, pp. 87–88, text-fig. 26h–l.
?1979 Didymograptus cf. artus Elles & Wood; Mu et al., pp. 64–65, pl. 21, .
1987 Didymograptus (Didymograptus) artus Elles & Wood; Fortey & Owens, pp. 258–259, figs 112b–d, 113.
1997 Didymograptus (Didymograptus) artus Elles & Wood; Maletz, p. 29, text-figs 9b, g, j, k, pl. 1, fig. B.
2000 Didymograptus (Didymograptus) artus Elles & Wood; Dean et al., p. 568, fig. 11b, c.
Material
More than 20 specimens; most are flat and preserve the proximal part; a few are partially pyritized. The specimens commonly contain 6–9 thecae on each stipe, with a maximum of 13 thecae (, R–T, ). The specimens are from the Didymograptus artus Biozone in the Shihtien Formation at the Laojianshan section, Baoshan; and the D. murchisoni Biozone, Shihtien Formation at the Banpo section, Shidian, Yunnan.
Description
The fork-like rhabdosome is composed of two pendent stipes. The maximum length is 8.0 mm. The two stipes diverge at angles of 90–120° at the proximal end, but curve ventrally to a common divergent angle of 25–30°, or become nearly parallel. The distal part of the stipes may be straight or slightly curved dorsally. The stipe width at the curvature point of th11 is 0.3 mm and at the aperture is commonly 0.6 mm. The most slender part of the stipes is the prothecal portion of th12 (0.15–0.2 mm). The stipe width increases to 0.9–1.0 mm at the aperture of th51, and then gradually to 1.2 mm at the aperture of th101 (). The stipe seems to widen slightly distally, but the exact width is unknown because no specimens preserving more than 12 thecae occur in the present collection.
Table 2. Stipe widths (mm) measured in five specimens of D. artus
The cone-shaped sicula is relatively short, ca 1.5–1.8 mm long and 0.3–0.4 mm wide at the sicular aperture. It is almost straight, but in some cases gently curves dorsally approaching the aperture, resulting in an arcuate form of the dorsal margin. The sicular apex is commonly detached and not preserved. The sicular aperture is gently concaved with development of a prominent rutellum, and opens slightly towards the second stipe. The first theca (th11) originates from the rutellar side of the sicula at a level approximately 0.9 mm above the sicular aperture, then extends downwards along the rutellar side until it curves away in the declined direction at 0.5 mm above the aperture; the basal free length of the sicula is 0.2–0.25 mm. The distal portion of th11 extends in the declined direction for 0.7 mm and curves ventrally with a gently concave ventral margin. The aperture of th11 is straight, at right angles to the axis of the stipe, and forms an acute angle of ca 65° with the ventral margin. Th12 originates from the left side of th11 at a level similar to the curvature point of th11, and extends nearly horizontally across the sicula on the reverse side with its ventral margin matching the cross-point of the dorsal margin and aperture of the sicula. It then curves ventrally, forming a prominent arc with an expanding distal portion and an obviously concave ventral margin. The thecal aperture of th12 is nearly straight or slightly concave, forming a sharply acute angle with the concave ventral margin, and a remarkable apertural process. The th21 originates from the dorsal side of th11 at the position just after the curvature of th11 from the sicula. The proximal development is of typically artus-type, with th11 being the dicalycal theca.
Normal thecae are tube-like, nearly straight or very gently curved ventrally, expanding slightly distally. Both the ventral and apertural margins of the thecae are nearly straight or slightly concave, forming prominent acute angles and apertural processes. The theca is 0.6 mm long and 0.25 mm wide at th22, and 1.4 mm long and 0.4 mm wide at th10. Thecal inclinations increase from 25° at the proximal end to 30–35° distally. Thecae overlap by 1/2 proximally and 2/3 distally. There are 8 thecae per 5 mm. The 2TRD is consistently 1.2–1.3 mm throughout.
Remarks. The species is distinctive in its small rhabdosome, relatively narrower stipes, slender sicula, and dicalycal th11 (artus-type proximal development). Our specimens from west Yunnan conform with the British types (Elles & Wood Citation1901). It is distinguished from D. spinulosus by its shorter sicula, and the artus-type proximal development (th12 dicalycal in the latter; see Jenkins Citation1983, text-, and Rickards & Khashogji Citation2000, ). It differs from D. pandus mainly in having a shorter sicula and a greater divergence angle at the proximal end. The species resembles Didymograptus acutus Ekström, Citation1937 in general morphology, but can be distinguished in having a shorter sicula (3 mm long in the latter). Rickards & Khashogji (Citation2000) regarded the latter (together with D. pandus and D. stabilis) as junior synonyms of D. murchisoni.
Fig. 8. Darriwilian graptolites from the Shihtien Formation in west Yunnan, China. A–E, G–H, Hustedograptus teretiusculus (Hisinger), A, NIGP 149267 (BSSD-4-62), B, NIGP149268 (BPSD-1-37), C, NIGP149269 (BSSD-4-35), D, NIGP149270 (BPSD-1-55), E, NIGP149271 (BPSD-1-33), G, NIGP 149272 (BPSD-1-46), H, NIGP149273 (BSSD-4-37). F, M, Archiclimacograptus riddellensis (Harris), F, NIGP149274 (BSSD-4-8), M, NIGP 149275 (BSSD-4-24). I–L, N, Archiclimacograptus angulatus (Bulman), I, NIGP149276 (BSSD-4-21a-6), J, NIGP149277 (BSSD-4-21b), K, NIGP149278 (BSSD-4-24), L, NIGP 149279 (BSSD-4-21a), N, juvenile form showing the proximal development, NIGP 149280 (BSSD-4-21c). All scale bars=1 mm.

Didymograptus murchisoni (Beck, 1839) (, K, N, Q; 7A–E)
1839 Graptolithus murchisoni Beck (in Murchison), pl. 26, .
1860 Didymograptus pakrianus Jaanusson, pp. 310–314, text-fig. 4, pl. 1, –, pl. 3, –.
1964 Didymograptus murchisoni (Beck); Berry, pp. 94–95, pl. 3, –.
1964 Didymograptus murchisoni geminus (Hisinger); Berry, pp. 95–96, pl. 3, .
1964 Didymograptus pakrianus Jaanusson; Berry, pp. 103–104, pl. 5, –, 4–5.
1973 Didymograptus murchisoni (Beck); Skevington, pp. 47–48, pl. 8, –, text-fig. 3a.
1979 Didymograptus murchisoni (Beck); Mu et al., pp. 69–70, pl. 23, figs 9–10.
1986 Didymograptus murchisoni (Beck); Strachan, pp. 12–13, pl. 3, , 9, 11–14.
1997 Didymograptus (Didymograptus) murchisoni (Beck); Maletz, pp. 27–29, pl. 1, fig. G, text-figs 9A, C, D, F, H–I.
2000 Didymograptus murchisoni (Beck); Rickards & Khashogji, pp. 182–186, –, , 9, 10a–d.
Material
More than fifteen flat specimens. Most of them are mature, preserved with proximal ends, and are developed with outgrowth of the periderm. The specimens are from the D. murchisoni Biozone in the Shihtien Formation at the Banpo section, Shidian, Yunnan.
Description
The fork-like rhabdosome is composed of two pendent stipes (, , , , ). The maximum length of the stipes in the present collection is 55.3 mm (commonly 15–35 mm), with over 60 thecae on each stipe. The stipe is 0.6–0.7 mm wide (1.1–1.3 mm with a pakrianus-like proximal sheath) in the proximal part (at th11). The stipes widen gradually to their maximum of 2.9–3.0 mm at a distance of 30 mm from the proximal end; they retain this width for 15 mm then sharply taper distally ( , 4). The two stipes diverge at 70–90° at the very proximal end but immediately curve ventrally and become parallel or nearly parallel at small diverging angles of 10–15°. The distal parts of the stipes become slightly more divergent (20°) in some specimens.
Table 3. Stipe width (mm) measured in five specimens of D. murchisoni. The position here refers to the distance (mm) from the proximal end (∗developed with outgrowth of periderm)
The detailed morphology of the proximal end is unknown due to the development of a pakrianus-like sheath. The sicula is robust, cone-shaped and straight, 2.3 mm in maximum length as measured in a few specimens (BPSD-1-32), although in most other specimens the apex parts are commonly detached. The exact origination position of th11 from the sicula is unknown. The dorsal margins of two stipes seem to be at the same level, forming a symmetrical proximal end. The pakrianus-like sheath is well developed in some mature specimens, linking the dorsal margin of the two stipes with the sicula, forming a triangular proximal part. The proximal sheath (membrane) also links the adjacent thecal apertures in the proximal part of the rhabdosome, making the thecal morphologies indiscernible.
The thecae in the proximal part of the rhabdosome curve prominently ventrally. The remarkably concave ventral margin and apertural margin of thecae form an acute angle on the ventral side. Thecal apertural processes are well developed in the immature specimens (especially for the proximal 10 pairs of thecae), but become obscure in the adult specimens due to the development of a proximal membrane. The thecae seem to expand gradually towards the apertures. The thecae are remarkably arc-shaped proximally but become gently curved to nearly straight in the middle and distal parts of the rhabdosome.
The thecae are 1 mm long, 0.4 mm wide at the apertures and inclined at angles of 40° proximally. They are 3.3 mm long and 0.7 mm wide (apertural part), inclined at angles of 50° in the distal part of the rhabdosome. Thecal overlap increases from only 1/2 in the proximal part to over 5/6 distally. There are 7.5 thecae per 5 mm in the proximal part of the rhabdosome (2TRD=1.2 mm), and 6 thecae per 5 mm in the distal part (2TRD=1.8 mm).
Remarks
The species is distinguished by its robust and long sicula, and the development of a pakrianus-like sheath in the proximal part. It also differs from D. artus in its much wider stipes and larger rhabdosome. No well-preserved juvenile forms of D. murchisoni are available in our collections, therefore, the distinctions between immature forms without a pakrianus-like sheath and the similar D. spinulosus, as proposed by Rickards & Khashogji (Citation2000), are unclear. The specimens described by Jenkins (Citation1983) as Didymograptus pluto, comprise at least two species, some belonging to D. murchisoni, others to D. spinulosus (see Fortey & Owens Citation1987, pp. 255–257; Maletz Citation1997, p. 27).
Didymograptus spinulosus Perner, 1895 ()
1895 Didymograptus spinulosus Perner, p. 22, pl. 5, figs 9, 10.
1973 Didymograptus spinulosus Perner; Bouček, pp. 88–90, pl. 13, –, text-fig. 27a–d.
1987 Didymograptus (Didymograptus) spinulosus Perner; Fortey & Owens, pp. 255–258, figs 110, 111, 112.
2000 Didymograptus (Didymograptus) spinulosus Perner; Dean et al., p. 568, fig. 10i–l.
Material
Only one flat specimen with the proximal end and eight thecae on each stipe is available (). The specimen is from the D. murchisoni Biozone in the Shihtien Formation at the Laojianshan section, Baoshan, Yunnan.
Description
The tuning fork-like rhabdosome is composed of two pendent stipes, 8.8 mm long and 3.7 mm wide. The stipes diverge primarily at an angle of ca 90°, but abruptly curve ventrally to become nearly parallel. The stipes are relatively slender, 6.5 mm long, 0.7 mm wide at th11 and 1.2 mm wide at th81.
The sicula is long and slender, ca 2.1 mm long and 0.4 mm wide at the aperture. The metasicula gently curves dorsally, and expands prominently approaching the sicular aperture. The aperture is smooth and flat but slightly inclined towards the second stipe, with an ambiguous rutellum. Th11 originates from the rutellar side of the sicula at 1 mm above the sicular aperture, and then extends downwards along the sicula until it turns to the declined-pendent direction at a 0.7 mm above the sicular aperture. The basal free length of the sicula is ca 0.3 mm. The free distal portion of th11 is 0.9 mm long and 0.3 mm wide at the aperture, with the ventral margin slightly concave to nearly straight. The aperture of th11 is gently concave with inconspicuous processes. The prothecal portion of th12 is very thin (0.2 mm wide), with its origination from th11 0.6–0.7 mm above the sicular aperture. The free ventral margin of th12 is nearly straight or inconspicuously concave. Th21 appears to originate from th11, but the exact point is unknown. The proximal end appears somewhat asymmetrical, the second stipe being slightly lower than the first one. The proximal development is of artus-type.
The normal thecae are nearly straight, tube-like with gently concave apertures. Thecae are inclined at angles of 35–40°, and overlap 1/2 proximally and 2/3 distally. There are 6 thecae per 5 mm in the proximal part (2TRD decreases from 1.5 mm proximally to 1.2 mm distally).
Remarks
The species closely resembles Didymograptus artus but has a much longer sicula.
Family DIPLOGRAPTIDAE Lapworth, 1873
Hustedograptus Mitchell, 1987
Type species
Diplograptus uplandicus Wiman, 1895
Hustedograptus vikarbyensis (Jaanusson, Citation1960) (–F, L; 7G–H, L–N)
1960 Glyptograptus vikarbyensis Jaanusson, pp. 323–324, pl. 3, , , , text-.
1960 Glyptograptus cernuus Jaanusson; pp. 324–325, pl. 3, fig. 9, text-.
?1963 Glyptograptus vikarbyensis Jaanusson; Geh, pp. 251–252, fig. 11b.
1997 Hustedograptus vikarbyensis (Jaanusson); Maletz, pp. 41–43, pl. 2, figs G, I, M, text-figs 16D, 18A–I.
Material
Eight flat specimens with the proximal ends preserved (, , , ), from the D. murchisoni Biozone in the Laojianshan section, Baoshan, and the Banpo section, Shidian, Yunnan.
Description
Biserial rhabdosome straight, with a maximum length of 10.8 mm. The rhabdosome is 1.0–1.2 mm wide at the proximal end, widening gradually to a maximum of 1.9 mm distally.
The sicula is 0.2 mm wide at the aperture, but the length is unknown due to enclosure by thecae. The robust virgella tapers but is typically straight; it is slightly inclined to the dorsal side in some specimens; 0.5 mm long. The sicular aperture is conspicuously concave, and no inconspicuous anti-virgellar spines are evident. Details of the proximal end are unavailable due to incomplete preservation. The distal portions of the first pair of thecae are gently curved, extending in an outward-reclined direction, each with a prominent and robust spine (0.2 mm long). The spine of th12 is derived from the free ventral wall somewhat below the thecal aperture, hence being a mesial spine, whereas that of th11 is from the aperture-apertural spine. No spines are present beyond the first pair of thecae. The proximal end appears symmetrical and broadly rounded.
The median suture is nearly straight or gently undulate. Thecae are apparently sigmoidal in the proximal part, and become less prominent distally. Thecal apertures are even and horizontal, forming obvious acute angles with the free ventral walls. There are 6.5–7 thecae per 5 mm in the proximal part. The 2TRD is 1.2 mm proximally and increases to a maximum of 1.7 mm distally.
Remarks
The morphology of the studied specimens agrees closely with the Swedish types described by Jaanusson (Citation1960), although some specimens have a smaller rhabdosome width (1.0–1.2 mm vs 1.4–1.5 mm at the proximal end). The specimens have a similar rhabdosome width to the types of Glyptograptus cernuus (see Jaanusson Citation1960), which is herein regarded as conspecific with H. vikarbyensis. The proximal end of H. vikarbyensis also resembles that of Orthograptus uplandicus (Wiman), but can be distinguished by the absence of anti-virgellar spines. The specimens found in the Miaopo Formation of the Yichang area and assigned to H. vikarbyensis by Geh (Citation1963) possess a much wider rhabdosome at the proximal end (2.1 mm), and may belong to another species.
Hustedograptus teretiusculus (Hisinger, 1840) (, , ; , )
1840 Prionotus teretiusculus (Nob.) Hisinger, p. 5, pl. 38, .
1907 Diplograptus (Glyptograptus) teretiusculus (Hisinger); Elles & Wood, pp. 250–252, pl. 31, .
1960 Glyptograptus cf. teretiusculus (Hisinger); Jaanusson, p. 322, pl. 3, figs 10–11.
1963 Glyptograptus cf. teretiusculus (Hisinger); Geh, p. 250–251, fig. 11a.
?1986 Glyptograptus teretiusculus (Hisinger); Strachan, p. 38, pl. 5, fig. 10, text-fig. 32.
1997 Hustedograptus teretiusculus (Hisinger) sensu Jaanusson, Citation1960; Maletz, pp. 39–41, pl. 2, fig. H, pl. 7, figs F, G, text-figs 16A, B, 17A–H.
2003 Hustedograptus teretiusculus (Hisinger); Toro & Brussa, p. 461, pl. 13, fig. 17, pl. 14, .
Material
More than 15 flat specimens with proximal ends. Most of them are mature, with over eight pairs of thecae (, , ; , ). The specimens are from the D. murchisoni Biozone in the Shihtien Formation at the Laojianshan section, Baoshan, and the Banpo section, Shidian, Yunnan.
Description
Rhabdosome robust and rigid, 144 mm in maximum length. The rhabdosome is 0.8–0.9 mm wide (spines excluded) at the proximal end, and widens to 1.4 mm at the 5th pair of thecae and 1.7 mm at the 10th pair of thecae, and reaches a maximum of 2 mm at the 13th pair of thecae, beyond which this width is retained.
The detailed morphology of the sicula is unknown due to incomplete preservation. The virgella is thick and straight, with a prominently thickened basal part, 0.8 mm long. The distal portion of th11 extends in the reclined direction and bears a rigid apertural spine, which is nearly straight, 0.5 mm long and in the reclined direction. The distal portion of th12 curves to nearly upwards, with a rigid mesial spine derived from a position slightly below the aperture. The spines of th11 and th12 are slightly elevated, and the aperture of th12 is remarkably higher than that of th11. The proximal part of the rhabdosome is obviously asymmetrical.
The normal thecae are gently sigmoidal, and originate in an atypical alternating pattern with their apertures slightly uneven. The geniculae are inconspicuous, and the thecae become nearly straight distally. The apertures of the proximal thecae are characterized by the development of prominent tongue-shaped dorso-lateral notches and lateral lappets that become progressively less prominent distally. In the middle and distal part of the rhabdosome, the thecal apertures are nearly straight, horizontal or slightly inclined dorsally, forming sharp acute angles of 30–50° with the supragenicular walls. The median suture is straight, and no projecting nema beyond the distal end is evident in our collection.
Thecae are inclined at angles of 30° proximally and 40° distally. Thecae overlap by 1/2–2/3. There are eight thecae per 5 mm proximally (2TRD=1.2 mm), and six thecae within the same distance distally (2TRD=1.6–1.7 mm).
Remarks
This species differs from Hustedograptus vikarbyensis (Jaanusson) in having an obviously asymmetrical proximal end with the aperture of th12 prominently higher than that of th11, and deeper excavations relative to the width of the rhabdosome.
Proclimacograptus Maletz, 1997
Type species
Proclimacograptus bulmani Maletz, 1997
Proclimacograptus angustatus (Ekström, Citation1937) ()
1937 Climacograptus angustatus Ekström, p. 36, pl. 7, –.
1937 Climacograptus celsus Ekström, p. 37, pl. 7, figs 7–11.
1944 Climacograptus paradoxus Bouček, p. 2, pl. 1, figs 1–11, pl. 2, figs 1–9.
1953 Climacograptus pauperatus Bulman, p. 512, pl. 1, figs 10–12, pl. 2, figs 13–16, text-.
1960 Climacograptus pauperatus Bulman; Jaanusson, p. 332, pl. 3, figs 12–14.
1964 Climacograptus pauperatus Bulman; Berry, pp. 134–135, pl. 13, , 11.
1964 Climacograptus angustatus Ekström; Berry, pp. 132–133, pl. 13, figs 12–13.
1997 Proclimacograptus angustatus (Ekström); Maletz, pp. 69–70, pl. 5, figs M–R, text-figs 33A–T.
Material
There are three flat specimens, two of which retain the proximal ends. The specimens are all from the D. artus and D. murchisoni biozones in the Shihtien Formation at the Laojianshan section, Baoshan.
Description
The rhabdosome is slender (), 20 mm in maximum length. It widens gradually from 0.5 mm at the proximal end, to 0.8 mm at th5, to a maximum of 1.0 mm at th9, and thereafter the width remains consistent until a slight narrowing at the very distal end. The median septum is straight, and the nema projects far beyond the distal end (for 6 mm).
The detailed morphology of the sicula is unknown due to incomplete preservation; only the straight virgella is clearly evident, being 0.8 mm long. The proximal end is narrowly rounded, with distinctively upturned distal portions of the first pair of thecae. No thecal spines are evident. Normal thecae are geniculate and of typical climacograptid type, with shallow excavations that occupy only 1/4 of the rhabdosome width. The supragenicular walls are parallel to the axis of the rhabdosome. Two thecal series are arranged alternately. There are six or seven thecae per 5 mm in the proximal part (2TRD=1.7 mm), and only five distally (2TRD=2.0 mm).
Remarks
These specimens agree well with the Swedish types described by Ekström (Citation1937). The present specimens differ from the pyritized specimens of Climacograptus pauperatus Bulman, Citation1953 found in the Oslo region only in having a relatively wider rhabdosome, which is regarded here as simply due to their different states of preservation and intraspecific variation.
Eoglyptograptus Mitchell, 1987
Type species. Fucoides dentatus Brongniart, 1828
Eoglyptograptus dentatus (Brongniart) sensu Bulman, Citation1963 (; )
1963 Glyptograptus dentatus (Brongniart); Bulman, pp. 673–675, pl. 96, –, text-, 4.
1965 Glyptograptus dentatus (Brongniart); Skevington, pp. 55–56, figs 60–62.
1987 Eoglyptograptus dentatus (Brongniart) sensu Bulman; Mitchell, text-, .
1992 Eoglyptograptus dentatus (Brongniart); Mitchell, , .
1997 Eoglyptograptus dentatus (Brongniart) sensu Bulman; Maletz, pp. 45–47, pl. 2, figs A–D, F, J, pl. 6, fig. H, pl. 7, figs H, K, text-figs 16F, 20A–B, D–H.
Material
Only two flat specimens with proximal ends (, ) were recovered from the D. murchisoni Biozone in the Shihtien Formation at the Laojianshan section, Baoshan, and the Banpo section, Shidian, Yunnan.
Description
Rhabdosome straight with narrowly round and asymmetrical proximal end, 8 mm long. The rhabdosome width is 0.8 mm at the proximal end, and increases sharply to 1.5 mm at the fifth theca, and thereafter increases slightly or remains nearly unchanged.
The detailed morphology of the sicula is indeterminate; virgella straight and 0.7–0.8 mm long. The distal portion of th11 curves away from the sicula at a level significantly lower than the sicular aperture, and extends in a reclined direction, being 0.4 mm long. The thecal aperture is nearly straight or slightly convex, with a prominent sub-apertural spine extending in the declined direction. Th12 turns from downwards to upwards at the sicular aperture with its aperture opening upwards. No apertural spines are evident on th12. The distal upward-extending portion is 0.6–0.7 mm long. Th11 and th12 are obviously asymmetrical, and their apertures differ by 0.5 mm in height. Th21 and subsequent thecae originate alternately, and are gently sigmoid with inconspicuous geniculae, gently convex free ventral walls and upwards-opening apertures. The free distal portion is 0.7–0.8 mm long. The morphologies of the thecal apertures are indeterminate.
Normal thecae overlap 1/3–1/2 proximally and are inclined at 20–40°. The 2TRD is 1.5–1.6 mm throughout.
Archiclimacograptus Mitchell, 1987
Type species
Pseudoclimacograptus angulatus sebyensis Jaanusson, 1960.
Archiclimacograptus angulatus (Bulman, Citation1953) (; ; , )
1953 Climacograptus scharenbergi var. angulatus Bulman, pp. 511–512, pl. 1, , text-.
1997 Archiclimacograptus angulatus angulatus (Bulman); Maletz, pp. 53–54, pl. 3, figs A–C, E, G, L, text-figs 25R, T–X.
2003 Archiclimacograptus angulatus angulatus (Bulman); Toro & Brussa, p. 462, pl. 16, fig. 11.
Material
There are over eight specimens with proximal ends (, , , ), some of which are partially to fully three-dimensional, from the Didymograptus murchisoni Biozone of the Shitien Formation at the Laojianshan Formation, Baoshan, Yunnan.
Description
The rhabdosome is straight, relatively slender and 5 mm in maximum length. The rhabdosome is almost uniform throughout with the two ventral sides nearly parallel, being 0.7 mm wide proximally and 0.8 mm at the fifth pair of thecae, remaining consistent thereafter to the distal end. The median septum is nearly straight or slightly undulate throughout.
Three spines have been found in the proximal end: the virgella and the apertural spines of the first pair of thecae. The detailed morphology of the sicula is unknown due to enclosure by subsequent thecae. The virgella is commonly preserved, 0.3 mm long, slightly inclined to the second series of thecae. The proximal end of the rhabdosome is broadly and obliquely squared, and the first pair of thecae is asymmetrical with the ventral wall forming an oblique line with the axis of the rhabdosome. The distal portion of th11 curves outwards broadly at a level slightly below the sicular aperture and then extends in a reclined direction with a 0.5 mm long sub-apertural (mesial) spine that extends in a horizontal-declined direction. The aperture of th11 is somewhat introverted. Th12 curves outwards from the sicula at the level of the sicular aperture, and the distal portion extends upwards with its aperture introverted as th11. A thick and robust, 0.6 mm long, sub-apertural (mesial) spine is present. The proximal development type is C-pattern in the present collection.
Subsequent thecae are sigmoidal but with obvious geniculae and gently convex to nearly straight supragenicular ventral walls that are nearly parallel to the axis of the rhabdosome. No apertural spines have been found except in the first pair of thecae. The apertural excavations are narrowly U-shaped and inclined dorsally, occupying approximately 1/4 of the rhabdosome width. There are eight thecae per 5 mm in the proximal part (the 2TRD is 1.1 mm proximally and 1.5 mm distally).
Remarks
The studied west Yunnan specimens agree well with the types from Oslo described by Bulman (Citation1953). No apertural spines at the first pair of thecae were observed in the Oslo specimens, probably due to their pyritized preservation. The species is distinguished from Archiclimacograptus riddellensis (Harris, Citation1924) in having more widely spaced thecae, and from Pseudoclimacograptus scharenbergi (Lapworth in Armstrong et al., 1876) in its gently geniculate thecae, the obliquely arranged excavations and in the structure of the proximal end (see Mitchell Citation1987).
Archiclimacograptus riddellensis (Harris, Citation1924) (; , )
1924 Climacograptus riddellensis Harris, pp. 100–101, pl. 8, figs 11, 12.
1960 Pseudoclimacograptus angulatus sebyensis Jaanusson, pp. 330–331, pl. 4, figs 5–9, text-.
1964 Climacograptus angulatus Bulman; Berry, pp. 125–128, pl. 13, .
1964 Climacograptus angulatus magnus Berry, pp. 128–130, pl. 11, , 6, 9–11, pl. 12, –.
1966 Climacograptus riddellensis Harris; Berry, pp. 439–440, pl. 50, –.
1987 Pseudoclimacograptus (Archiclimacograptus) angulatus sebyensis Jaanusson; Mitchell, p. 387, figs 21N, Q.
1989 Pseudoclimacograptus angulatus sebyensis Jaanusson; Hughes, p. 70, pl. 3, figs j, k, text-figs 23i–j, 28k–l.
1992 Pseudoclimacograptus (Archiclimacograptus) riddellensis (Harris); VandenBerg & Cooper, , .
1997 Archiclimacograptus riddellensis (Harris); Maletz, pp. 55–56, pl. 3, figs D, F, H–K, M, O, Q, R, T, U, text-figs 21A, 25A–Q, S.
Material. There are two flat, mature specimens with proximal ends and nine pairs of thecae (–B, 8F, M). They are from the D. murchisoni Biozone of the Shitien Formation, Baoshan.
Description
Rhabdosome small with broadly squared proximal end, with a maximum length of 4.1 mm. The rhabdosome has nearly uniform width with a very gentle increase distally, 0.9 mm wide in the proximal end (apertural spines excluded), 1.0 mm wide at the fifth pair of thecae, 1.1 mm wide at the ninth pair of thecae.
The detailed morphology of the sicula is unknown; but the thick virgella (over 0.3 mm long) is inclined obliquely to the dorsal side of the sicula. The distal portion of th11 curves outwards at a level slightly below the sicular aperture. The aperture of th11 opens upwards with a robust, horizontal subapertural spine 0.6 mm long. Th12 curves outwards at the level of the sicular aperture and its apertural portion extends upwards with a prominently convex ventral margin and introverted aperture. The apertural margin of th12 is slightly convex and inclined towards the dorsal direction, forming a long and narrow, convex-upwards excavation together with the infragenicular ventral wall. A thick, horizontal subapertural spine is developed, more than 0.4 mm long. The subsequent thecae possess no apertural spines.
The normal thecae are prominently genicular, with a convex-upwards infragenicular wall and a gently convex-outwards supragenicular wall that is 0.5 mm long, and a convex-upwards apertural margin. Inconspicuous lateral lappets and collars are present in most thecae. The apertural excavations are deep and narrow, prominently inclined dorsally, occupying one-third of the rhabdosome width. The median septum is zigzagged. The thecae are densely spaced, 2TRD being 0.9–1.0 mm throughout.
Remarks
The specimens described here agree closely with the types described by Harris (Citation1924), but have a slightly broader proximal end of the rhabdosome, which may be due to a preservational artefact. The species resembles Archiclimacograptus angulatus (Bulman, Citation1953), but differs in having more closely spaced thecae (2TRD: 0.9–1.0 mm vs 1.1–1.5 mm).
Haddingograptus Maletz, 1997
Type species. Pseudoclimacograptus oliveri Bouček, 1973
Haddingograptus oliveri (Bouček, Citation1973) (, )
1973 Pseudoclimacograptus (Pseudoclimacograptus) oliveri Bouček, p. 121.
1997 Haddingograptus oliveri (Bouček, Citation1973); Maletz, pp. 64–65, pl. 4, figs A–S, pl. 7, figs I, J, text-figs 30A–S, 32A, B.
Material
Only one incomplete but partially pyritized specimen lacking the proximal end is available (, ), from the D. artus Biozone of the Shihtien Formation, Baoshan.
Description
Rhabdosome is slender, with the broken portion being 10.4 mm long and 1.4 mm wide. The proximal end is broken and not preserved. Thecae are of typical haddingograptid type, with prominent geniculae, collars, and convex supragenicular walls 0.7 mm long. Thecal apertures are introverted with U-shaped excavations that are 0.4 mm deep and 1.6 mm high. The apertural margin seems to be even and straight, and a prominent ventral notch is observed in the genicular collar. The median septum is undulate. There are five thecae per 5 mm (2TRD=1.9 mm).
Table 4. Stipe width (mm) measured in one specimen of D. murchisoni without a pakrianus-like proximal sheath. The position here refers to theca number
Acknowledgements
We are indebted to Luo Huilin of the Kunming Institute of Geology and Mineral Resources, and Xiao Yinwen of the Yunnan Geological Survey, and our colleague Wang Yi for providing information and assistance with the fieldwork. We thank Chuck Mitchell and Barrie Rickards for their careful and constructive reviews of the manuscript. The trilobites of the Shihtien Formation were identified by Zhou Zhiyi. Financial support from the Chinese Academy of Sciences (KZCX2-YW-122), Ministry of Science and Technology (2006CB806402, 2006FY1203000-4) and Natural Science Foundation of China (40532009) is acknowledged.
Related Research Data
References
- Armstrong , J. , Young , J. and Robertson , D. 1876 . “ Catalogue of the Western Scottish Fossils ” . In Proceedings of the British Association of the Advancement of Science 1876 164 Glasgow
- Beck , D. 1839 . The Silurian System , 786 London : R.I. Murchison, John Murray .
- Berry , W. B.N. 1964 . The Middle Ordovician of the Oslo Region, Norway . Norsk Geologisk Tidsskrift , 44 : 61 – 170 .
- Berry , W. B.N. 1966 . A discussion of some Victorian Ordovician graptolites . Proceedings of the Royal Society of Victoria , 79 : 415 – 448 .
- Bouček , B. 1944 . O nových názech graptolitů v českém ordoviku . Veda přírodní , 22 : 226 – 233 .
- Bouček , B. 1973 . Lower Ordovician Graptolites of Bohemia , 185 Prague : Publishing House of the Czechoslovak Academy of Sciences .
- Brongniart , A. 1828 . Histoire des végétaux fossiles Paris XII + 488 pp
- Brown , J. C. 1913 . Contributions to the geology of the Province of Yunnan in western China, (3) notes on the stratigraphy of the Ordovician and Silurian Beds of western Yunnan . Records of the Geological Survey of India , 1913 ( Part 3 ) : 327 – 334 .
- Brown , J. C. 1916 . Contributions to the geology of the Province of Yunnan in western China, (5) Geology of parts of the Salween and Mekong Valleys . Records of the Geological Survey of India , 47 : 205 – 266 .
- Brussa , E. D. , Mitchell , C. E. , Ortega , G. , Maletz , J. and Astini , R. A. . Middle Ordovician graptolite biostratigraphy from the Los Azules Formation at Los Gatos Creek, Central Precordillera, Argentina . Proceedings of the 7th International Graptolite Conference & Field Meeting of the International Subcommission on Silurian Stratigraphy . Edited by: Ortega , G. and Acenolaza , G. F. Instituto Superior de Correlacion Geologica (INSUGEO), Tucumán, Argentina, 21–25
- Brussa , E. D. , Toro , B. A. and Benedetto , J. L. 2003b . “ Biostratigraphy ” . In Ordovician Fossils of Argentina , Edited by: Benedetto , J. L. 75 – 90 . Cordoba : Universidad Nacionnal de Cordoba .
- Bulman , O. M.B. 1953 . Some graptolites from the Ogygiocaris Series (4aa) of the Oslo district . Arkiv för Zoologi , 1 : 509 – 517 .
- Bulman , O. M.B. 1963 . On Glyptograptus dentatus (Brongniart) and some allied species . Palaeontology , 6 : 665 – 689 .
- Chen , X. and Rong , J. Y. 1992 . “ Ordovician plate tectonics of China and its neighboring regions ” . In Global Perspectives on Ordovician Geology , Edited by: Webby , B. D. , Laurie , J. R. and Balkema , A. A. 277 – 291 . Rotterdam .
- Cocks , L. R.M. and Zhan , R.-B. 1998 . Caradoc brachiopods from the Shan States, Burma (Myanmar) . Bulletin of the Natural History Museum, (Geology), London , 54 : 109 – 130 .
- Cooper , A. H. , Rushton , A. W.A. , Molyneux , S. G. , Hughes , R. A. , Moore , R. M. and Webb , B. C. 1995 . The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the English Lake District . Geological Magazine , 132 : 185 – 211 .
- Dean , W. T. , Monod , O. , Rickards , R. B. , Demir , O. and Bultynck , P. 2000 . Lower Paleozoic stratigraphy and palaeontology, Karadere-Zirze area, Pontus Mountains, northern Turkey . Geological Magazine , 137 : 555 – 582 .
- Ekström , G. 1937 . Upper Didymograptus Shale in Scania . Sveriges Geologiska Undersökning, Ser. C. , 403 : 1 – 53 .
- Elles , G. L. and Wood , E. M.R. 1901 . A Monograph of British Graptolites, Part 1—Dichograptidae , 54 London : Palaeontographical Society .
- Elles , G. L. and Wood , E. M.R. 1907 . A Monograph of British Graptolites VI , London : Palaeotographical Society . xcvii–cxx + 217–272
- Fang , Z. J. and Cope , J. C.W. 2008 . Affinities and palaeobiogeographical significance of some Ordovician bivalves from east Yunnan, China . Alcheringa , 32 : 297 – 312 .
- Fang , Z. J. , Zhou , Z. C. and Lin , M. J. 1992 . On several questions concerning Changing-Menglian Suture from perspect of stratigraphy . Journal of Stratigraphy , 16 : 292 – 303 .
- Fortey , R. A. and Cocks , L. R.M. 1998 . “ Biogeography and palaeogeography of the Sibumasu terrane in the Ordovician: a review ” . In Biogeography and Geological Evolution of SE Asia , Edited by: Hall , R. and Holloway , J. D. 43 – 56 . Leiden : Backhuys Publishers .
- Fortey , R. A. and Cocks , L. R.M. 2003 . Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions . Earth-Science Reviews , 61 : 245 – 307 .
- Fortey , R. A. and Owens , R. M. 1987 . The Arenig Series in South Wales . Bulletin of the British Museum (Natural History), Geology , 41 : 69 – 307 .
- Fortey , R. A. , Harper , A. T. , Ingham , J. K. , Owen , A. W. and Rushton , A. W.A. 1995 . A revision of Ordovician series and stages from the historical type area . Geological Magazine , 132 : 15 – 30 .
- Fortey , R. A. , Harper , A. T. , Ingham , J. K. , Owen , A. W. , Parkes , M. A. , Rushton , A. W.A. and Woodcock , N. H. 2000 . A revised correlation of Ordovician Rocks in the British Isles . Geological Society Special Report , 24 : 1 – 83 .
- Frech , F. 1897 . Lethaea Geognostica; Theil 1, Lethaea Palaeontologica, 1. BD., Graptolithiden , 544 – 684 . Stuttgart : Schweizerbart .
- Geh , M. Y. 1963 . Graptolites from the Miaopo Shale (Middle Ordovician), W. Hubei (II) . Acta Palaeontologica Sinica , 11 : 240 – 261 .
- Hall , J. 1851 . New genera of fossil corals . American Journal of Science , 11 : 398 – 401 .
- Hall , J. 1858 . Descriptions of Canadian graptolites . Report of the Geological Survey of Canada , 1857 : 111 – 145 .
- Hall , J. 1865 . Figures and Descriptions of Canadian Organic Remains—Graptolites of the Quebec Group , 151 Montreal : Geological Survey of Canada .
- Harris , W. J. 1924 . Victorian graptolites (new series), Part I . Proceedings of the Royal Society of Victoria , 36 : 92 – 106 .
- Harris , W. J. and Keble , R. A. 1932 . Victorian graptolite zones, with correlations and description of species . Proceedings of the Royal Society of Victoria , 44 : 25 – 48 .
- Hisinger , W. 1840 . Lethaea Suecica seu Petrificata Suecica, supplementum 2 , 11 Stockholm : Holmiae .
- Holm , G. 1881 . Bidrag till kånnedomen om Skandinaviens Graptoliter I . Kunglia Svenska Vetenskapsakademi Förhandlingar , 38 : 71 – 84 .
- Hopkinson , J. and Lapworth , C. 1875 . Description of the graptolites of the Arenig and Llandeilo rocks of St. David's . Quarterly Journal of the Geological Society of London , 31 : 631 – 672 .
- Hughes , R. A. 1989 . Llandeilo and Caradoc graptolites of the Builth and Shelve Inliers . Monograph of the Palaeontographical Society , 141 ( 577 ) : 1 – 89 .
- Jaanusson , V. 1960 . Graptoloids from the Ontikan and Viruan (Ordov.) limestones of Estonia and Sweden . Bulletin of the Geological Institutions of the University of Uppsala , 38 : 289 – 365 .
- Jenkins , C. J. 1983 . Ordovician graptolites from the Great Paxton Borehole, Cambridgeshire . Palaeontology , 26 : 641 – 653 .
- Lapworth , C. 1873 . On an improved classification of the Rhabdopora, Part I . Geological Magazine , 10 : 500 – 504 .
- Lapworth , C. 1876 . On Scottish Monograptidae 4 . Geological Magazine , 13 : 544 – 552 .
- Liu , B. P. , Feng , Q. L. and Fang , N. Q. 1993 . Tectonic evolution of Palaeo-Tethys Poly-Island-Ocean in Changning-Menglian and Lancanjiang belts, southwestern Yunnan, China . Earth Science: Journal of China University of Geosciences , 18 : 529 – 539 .
- Loczy , L. V. 1893 . Die Wissenschaftlichen Ergebnisse der Reise des Grafen Béla Széchenyi in Ost-Asien , Vol. 1 , 767 Wien : Hölzel .
- Maletz , J. 1997 . Graptolites from the Nicholsonograptus fasciculatus and Pterograptus elegans Zones (Abereiddian, Ordovician) of the Oslo region, Norway . Greifswalder Geowissenschaftliche Beitraege , 4 : 5 – 98 .
- Maletz , J. 2005 . Early Middle Ordovician graptolite biostratigraphy of the Lovisefred and Albjära drill cores (Scania, southern Sweden) . Palaeontology , 48 : 763 – 780 .
- Metcalfe , I. 1998 . “ Palaeozoic and Mesozoic geological evolution of the SE Asian region: multidisciplinary constraints and implications for biogeography ” . In Biogeography and Geological Evolution of SE Asia , Edited by: Hall , R. and Holloway , J. D. 25 – 41 . Leiden : Backhuys .
- Miller , S. A. 1889 . North American Geology and Paleontology , 664 Cincinnati .
- Mitchell , C. E. 1987 . Evolution and phylogenetic classification of the Diplograptacea . Palaeontology , 30 : 353 – 405 .
- Mitchell , C. E. 1992 . “ Evolution of the Diplograptacea and the international correlation of the Arenig-Llanvirn boundary ” . In Global Perspectives on Ordovician Geology , Edited by: Webby , B. D. and Laurie , J. R. 171 – 183 . Rotterdam : Balkema .
- Mu , E. Z. , Ge , M. Y. , Chen , X. , Ni , Y. N. and Lin , Y. K. 1979 . Lower Ordovician graptolites of Southwest China . Palaeontologia Sinica , 156 ( New Series B, no. 13 ) : 1 – 192 .
- Murchison , R. I. 1839 . The Silurian System (2 volumes) , 786 London : John Murray .
- Nicholson , H. A. 1872 . A Monograph of the British Graptolitidae , London : Edinburgh . i–x, 133 pp
- Perner , J. 1895 . Studie o ceskych graptolitech II , 52 Prague : Leipzig .
- Popov , L. E. , Ghobadi Pour , M. and Hosseini , M. “ Early to Middle Ordovician lingulate brachiopods from the Lashkarak formation, Eastern Alborz Mountains, Iran ” . Alcheringa 32, 1–35
- Reed , F. R.C. 1917 . Ordovician and Silurian fossils from Yun-nan . Memoirs of the Geological Survey of India, Palaeontologica Indica (New Series) , 6 ( 3 ) : 1 – 69 .
- Rickards , R. B. and Khashogji , M. S. 2000 . The tuning fork graptoloid, Didymograptus murchisoni (Beck, 1839) . Transactions of the Royal Society of Edinburgh: Earth Sciences , 90 : 173 – 187 .
- Ruedemann , R. 1908 . Graptolites of New York (Part 2, Graptolites of the Higher Beds) . New York State Museum Memoir , 11 : 1 – 583 .
- Ruedemann , R. 1947 . Graptolites of North America . Geological Society of America Memoir , 19 : 1 – 652 .
- Sedgwick , A. and M'Coy , F. 1851 . A Synopsis of the Classification of the British Palaeozoic Rocks, with a Systematic Description of the British Palaeozoic Fossils Found in the Geological Museum of the University of Cambridge , 832 Cambridge : Cambridge University Press .
- Skevington , D. 1965 . Graptolites from the Ontikan Limestones (Ordovician) of Öland, Sweden. II. Graptoloidea and Graptovermida . Bulletin of the Geological Institutions of the University of Uppsala , 43 : 1 – 74 .
- Skevington , D. 1973 . Graptolite fauna of the Great Paxton Borehole, Huntingdonshire . Bulletin of the Geological Survey of Great Britain , 43 : 41 – 60 .
- Strachan , I. 1986 . The Ordovician graptolites of the Shelve District, Shropshire . Bulletin of the British Museum (Natural History), Geology , 40 ( 1 ) : 1 – 58 .
- Sun , Y. C. 1945 . The Sino-Burmese Geosyncline of early Palaeozoic time with special reference to its extent and character . Bulletin of the Geological Society of China , 25 : 1 – 7 .
- Sun , Y. C. and Szetu , H. T. 1947 . Stratigraphy and Structure of Baoshan, Western Yunnan . Proceedings of the Geology Department of Peking University , 32 : 1 – 15 .
- Toro , B. A. and Brussa , E. D. 2003 . “ Chapter 11: Graptolites ” . In Ordovician Fossils of Argentina , Edited by: Benedetto , J. L. 441 – 505 . Cordoba : Universidad Nacional de Cordoba .
- VandenBerg , A. H.M. and Cooper , R. A. 1992 . The Ordovician graptolite sequence of Australasia . Alcheringa , 16 : 33 – 85 .
- Whitfield , R. P. 1902 . Notice of a new genus of marine algae fossil in the Niagara Shale . Bulletin of the American Museum of Natural History , 16 : 399 – 400 .
- Wiman , C. 1895 . Über die Graptolithen . Bulletin of the Geological Institutions of the University of Uppsala , 2 : 239 – 316 .
- Yin , T. H. 1936–1937 . Brief description of the Ordovician and Silurian Fossils from Shihtien . Bulletin of the Geological Society of China 16 (special volume in honour of Ding Wenjiang) , : 281 – 298 .
- Yin , T. H. and Lu , C. H. 1936–1937 . On the Ordovician and Silurian beds of Shihtien, western Yunnan . Bulletin of the Geological Society of China 16 (special volume in honour of Ding Wenjiang) , : 41 – 56 .
- Yu , J. H. 1962 . Graptolites (Ptilograptidae) from West Yunnan . Bulletin of Nanjing University (Geology) , 2 : 47 – 54 .
- Yunnan Geological Survey . 1980 . Regional Geological Report (Baoshan, with 1/200,000 Geological Map) Ordovician System , 26 – 42 . Baoshan : Yunnan Geological Survey .
- Zalasiewicz , J. A. , Taylor , L. S. , Rushton , A. W.A. , Loydell , D. K. , Rickards , R. B. and Williams , M. in press . Graptolites in British Stratigraphy . Geological Magazine ,
- Zhan , R. B. and Cocks , L. R.M. 1998 . Late Ordovician brachiopods from the South China Plate and their palaeogeographical significance . Special Papers in Palaeontology , 59 : 1 – 70 .
- Zhang , Y. D. and Chen , X. 2003 . “ The Early-Middle Ordovician graptolite sequence of the Upper Yangtze region, South China ” . In Ordovician from the Andes , Edited by: Albanesi , G. L. , Beresi , M. S. and Peralta , S. H. 173 – 180 . Tucuman : Universidad Nacional de Tucuman .
- Zhang , Y. D. , Chen , X. and Goldman , D. 2007 . Diversification patterns of Early and Mid Ordovician graptolites in South China . Geological Journal , 42 : 315 – 337 .
- Zhou , Z. Y. and Zhen , Y. Y. 2008 . Trilobite-constrained Ordovician biogeography of China with reference to faunal connections with Australia . Proceedings of the Linnean Society of New South Wales , 129 : 183 – 195 .
- Zhou , Z. Y. , Dean , W. T. and Luo , H. L. 1998 . Early Ordovician trilobites from Dali, West Yunnan, China, and their palaeogeographical significance . Palaeontology , 41 : 429 – 460 .