
Abstract
Gouramanis, C. & McLoughlin, S., January 2016. Siluro-Devonian trace fossils from the Mereenie Sandstone, Kings Canyon, Watarrka National Park, Amadeus Basin, Northern Territory, Australia. Alcheringa 40, xxx–xxx. ISSN 0311-5518
Nine trackways referable to the ichnogenus Diplichnites are preserved in the upper Silurian to Lower Devonian Mereenie Sandstone at Kings Canyon, Watarrka National Park, Northern Territory, Australia. Eight trackways are consistent with earlier descriptions of D. gouldi, and one trackway could not be assigned to an ichnospecies. The trackways are co-preserved with a range of sub-horizontal burrows referable to Beaconites and Taenidium, and several vertical burrows, surficial circular traces and a horizontal trail of uncertain identities. The ichnofossil assemblage highlights the diversity of animals present in the late Silurian to Lower Devonian paralic to fluvial environments of central Australia at the time of early colonization of the land’s surface. The assemblage is similar to ichnofaunas from coeval strata elsewhere in Australia and throughout Gondwana, and it highlights the potential of this region for further ichnological studies to elucidate the early stages of terrestrialization in the palaeoequatorial belt.
Chris Gouramanis [[email protected]], Earth Observatory of Singapore, Nanyang Technological University, Singapore, 639798; Stephen McLoughlin [[email protected]], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, S-104 05 Stockholm, Sweden.
INVERTEBRATE trace fossils have been found in Australian continental settings from the Early Cambrian (e.g., Walter et al. Citation1989) and have provided important information on the ecological structure of early terrestrial communities. By the late Silurian and Early Devonian in Australia, a diverse terrestrial invertebrate fauna existed based on the presence of trace fossils described from the Tumblagooda Sandstone in Western Australia (Trewin & McNamara Citation1994), Major Mitchell Sandstone in western Victoria (Gouramanis et al. Citation2003), and Mt Daubeny Formation (Neef Citation2004a) and Ravendale Formation (Neef Citation2004b) in New South Wales. Encroachment of animals onto the land’s surface globally at this time was probably facilitated by the contemporaneous diversification of bryophytes and early vascular plants as evidenced by both macrofossil and palynological records (Gray et al. Citation1974, Hagström & Mehlqvist Citation2012, Mehlqvist et al. Citation2015, Edwards & Kenrick Citation2015).
The upper Silurian–Lower Devonian mixed eolian and fluvial strata forming the upper Mereenie Sandstone in the western section of the George Gill Range (Wells et al. Citation1970, Bagas Citation1988) were deposited in sedimentary environments similar to those of the Tumblagooda Sandstone and Grampians Group. During the Silurian and Early Devonian, central Australia was located within 15° of the equator (Pickett et al. Citation2000, Talent et al. Citation2000). Thus, the Mereenie Sandstone and its stratigraphic equivalents in western and southeastern Australia preserve the sedimentological and ichnological expression of tropical lowland environments that were being colonized by early terrestrial plant and invertebrate communities.
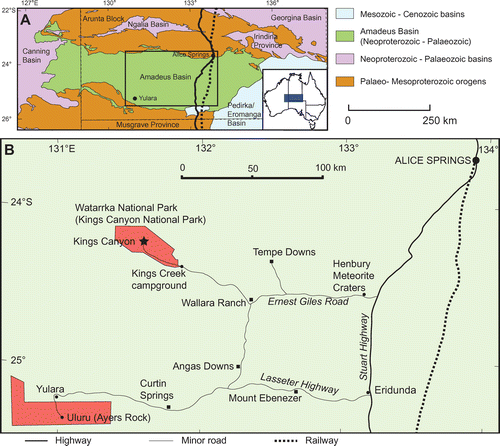
The Mereenie Sandstone crops out prominently at the iconic Kings Canyon within the Watarrka National Park, approximately 240 km west-southwest of Alice Springs within the Amadeus Basin, Northern Territory, Australia (Wells et al. Citation1970, Bagas Citation1988; Fig. ). The invertebrate trace fossil assemblages of the Mereenie Sandstone have not been described previously, although Wells et al. (Citation1970) recognized Skolithos piperock from the lower Mereenie Sandstone and Bagas (Citation1988, p. 14) noted “indeterminate trace fossils on bedding planes” near the ‘Garden of Eden’ staircase in the Watarrka National Park that resemble large interstitial burrows. Several other trace fossils occur within the Mereenie Sandstone at Kings Canyon, and some of the major forms are identified and described here for the first time.
Fig. 1. Locality details. A, Map of central Australia showing the position of the Amadeus Basin with respect to neighbouring geological provinces. B, Detailed map of the area delineated in A showing the location of Kings Canyon. Further geological details of the area were provided by Bagas (Citation1988).
Materials and methods
Several ichnogenera are preserved on bedding planes of the Mereenie Sandstone located on or near the Kings Canyon tourist walk, Watarrka National Park (Fig. ). Examples of these traces were photographed during two field campaigns in July 2012 and April 2015. In particular, nine large trackways (T1–T9) were recorded, with four located on the thin, horizontal to shallow-dipping beds forming the natural steps on the eroded ‘beehive’ structures at Cotterills Lookout (24°15′4″S, 131°34′57″E; Fig. A, B: T1–T3; Fig. F, G: T4), four from an extensive pavement near Kings Canyon Rim Walk navy blue marker 46 (KCRW46; 24°15′8.27″S, 131°34′58.7″E; Fig. C–E: T5–T8), and a single specimen from a cemented block forming part of the stairs near KCRW80 (24°15′20.35″S, 131°34′36.1″E; Fig. H: T9). Other traces described herein include a small trail near the top of the northern ascent to the canyon rim walk (24°14′59.96″S, 131°34′25.35″E; Fig. ); small circular traces exposed in the eastern cliff of the ‘Garden of Eden’ walk (24°14ʹ56.46″S, 131°34ʹ59.71″E; Fig. ); small backfilled burrows in blocks forming part of the stairs near 24°15ʹ23.11″S, 131°34ʹ33.43″E; Fig. ); large backfilled burrows on the canyon rim walk (24°14ʹ56.02″S, 131°34ʹ54.11″E; Fig. ); and a radial burrow-mouth trace near the eastern rim of the canyon (24°15ʹ00.08″S, 131°35ʹ01.08″E; Fig. ).
Fig. 2. Diplichnites gouldi trackways with arrows depicting the direction of movement. A, Photo, and B, interpretive diagram of trackways T1–T3 from Cotterills Lookout. C–E, Photos and interpretive diagram of trackways T5–T8 from KCRW46. F, Photo, and G, interpretive diagram of the trackway T4 from Cotterills Lookout. H, Diplichnites isp. (trackway T9) found on a boulder as float in cemented (arrows) stairs near KCRW80. Note sunglasses are 145 mm wide in each image and scale bars = 100 mm.

Fig. 3. Small, complex trail on a sandstone block near the top of the northern ascent to the Kings Canyon Rim Walk. A, Overview of the trail; B, Enlargement of the best-preserved end of the trail; C, Line-drawing of a portion of the trail showing individual elements. Scale bars = 10 mm.

Fig. 4. Surficial circular traces (?Laevicyclus isp.) recorded at the base of the staircase leading up from the eastern base of the ‘Garden of Eden’ walk. Scale bar = 100 mm.

Fig. 5. Horizontal to sub-horizontal Beaconites antarcticus (Vialov) emend. Keighley & Pickerill, Citation1994, lined, backfilled burrows in isolated blocks near the top of the stairs descending from the southern rim of the Kings Canyon trail. A, Burrows in fine-grained sandstone with iron-stained infill. B, Enlargement of selected burrows in A. C, Burrows with silty lining and diffuse meniscate laminae in fine-grained sandstone. D, Enlargement of one burrow in C. Scale bars = 10 mm.

Fig. 6. Large, horizontal to oblique Taenidium barretti (Bradshaw) emend. Keighley & Pickerill, Citation1994 burrows. A, Several irregularly orientated burrows associated with a ripple-marked bedding plane. B, Low-angle view of two burrows illustrated in A. C, Cluster of intersecting burrows, some with Cruziana-like surface striations. Circled area hosts numerous ?vertical Skolithos-like burrow openings. D, E, Horizontal burrows bearing complex surface striations. F, Horizontal burrow bearing fine transverse striations interpreted to represent current lineations on the partially exhumed burrow fill. G, I, Horizontal burrows bearing arcuate or en echelon markings along the margins (arrowed). H, Segmented burrow reminiscent of dorsal tergite impressions of a large arthropod. Scale bars = 10 mm except in A (=100 mm); coin in B and C = 25 mm diameter.

Fig. 7. Burrow entrance with radial feeding traces. A, Burrow opening exposed on the surface of a bed of fine-grained sandstone. B, Enlargement of A showing ridge and furrow traces radiating from a central featureless burrow mouth. Scale bars = 10 mm.

Ichnology
Diplichnites gouldi Bradshaw, Citation1981 (Fig. A–G)
Description and interpretation
Eight trackways of biserial epichnial imprints attributable to Diplichnites gouldi were identified (Table ). They have preserved lengths up to 0.61 m, internal widths of 10–117 mm, and external widths of 36–161 mm. Individual appendage imprints in the smallest trackway are ca 1–7 mm in diameter; those in the largest trackway are ca 19–24 mm in diameter.
Table 1. Summary of the dimensions of each Diplichnites trackway recorded from Kings Canyon.
Trackways T1–T3 (Fig. A, B), T5–T8 (Fig. C–E) and T4 (Fig. F, G) occur as biserial parallel sequences of circular to ovoid epichnial depressions reflecting individual or compound imprints (Table ). Epichnial mounds behind individual or grouped imprints are common and reflect sediment pushed backwards during forward propulsion, indicating the direction of movement. Trackways T6 and T7 transition from well-preserved epichnial depressions to parallel, continuous epichnial ridges, where overlying sediment has not been eroded (Fig. C–E). The transition from depressions to continuous ridges in T6 and T7 (Fig. C–E) is more likely due to preservation, i.e., where overburden has not been removed completely, rather than environmental conditions at the time of the trackway formation (cf. the animals walking from relatively dry to wet sediments as documented by Trewin & McNamara Citation1994).
The flanks of each trackway comprise sets of two to three closely spaced imprints that are either discrete or formed as superimposed clusters. Where the individual imprints are distinct, the orientation of the medial to lateral imprints are orientated forward (up to 20°), perpendicular or backwards (up to 26°) from the direction of movement. Typically, the distal and central imprints are larger and deeper and most likely to be preserved or identified, whereas the medial imprints are smaller and less commonly preserved. No medial ridges or grooves are present in any of these trackways.
Trackways T1 and T3 are straight and parallel; the animal was travelling eastwards. These trackways cross T2 but it is unclear as to which trackways were formed first (Fig. A, B). The T2 animal was travelling south-southeast before turning (29°) to the south. The animal that formed the short T4 was also heading east (Table , Fig. F, G), and part of the northern section of the trackway is obscured by a Beaconites antarcticus burrow (Fig. F). It is unclear from the preservation of the bedding plane how many tracks in a series are present (Fig. A, B).
The longer trackways (T5–T8, Fig. C–E) are all straight with T6 to T8 parallel, and T5 crossing T6 and T7 at angles of 20° and 29°, respectively. The push-back mounds observed behind the individual imprints of T5 indicate that the animal moved eastwards, whereas the animals producing T6 to T8 were moving in an east-northeasterly direction. It is assumed that T5 also crosses T8, but this is obscured by overburden. It is probable that T5 crossed T6 and T7 because T5 has distinct impressions between both T6 and T7, whereas the T6 and T7 imprints become obscure within T5. It is unclear whether T6 to T8 were formed at the same time, but the unidirectional movement and parallel nature of the trackways suggest that this is a possibility.
Remarks
There is great morphological and preservational variability both between and within individual Diplichnites gouldi trackways (e.g., Bradshaw Citation1981, Citation2010, Trewin & McNamara Citation1994). The forms preserved in the Mereenie Sandstone most closely resemble D. gouldi Form B from the Tumblagooda Sandstone, Western Australia (Trewin & McNamara Citation1994). Similar D. gouldi Form B trackways have been described from a range of sedimentary settings including coastal environments (Bradshaw Citation1981, Citation2010, Draganits et al. Citation2001), dry and damp aeolian surfaces (Trewin & McNamara Citation1994, Retallack Citation2009), subaqueous fluvial and damp subaerial surfaces (Woolfe Citation1993, Smith et al. Citation2003, Neef Citation2004a) and an emergent sheet-flooded fluvial environment (Neef Citation2004b). Diplichnites gouldi has been ascribed to myriapods (Briggs et al. Citation1979, Bradshaw Citation1981, Johnson et al. Citation1994, Trewin & McNamara Citation1994, Draganits et al. Citation2001, Smith et al. Citation2003, Neef Citation2004a), trilobites (Bradshaw Citation2010), euthycarcinoids (Trewin & McNamara Citation1994, Retallack Citation2009) and xiphosurans (Trewin & McNamara Citation1994). It is likely that a similar non-marine aqueous to terrestrial trackway-producing arthropod existed in the Amadeus Basin as the Mereenie Sandstone was deposited.
Diplichnites isp. (Fig. H)
Description
This trackway (T9) occurs on a bedding plane within a boulder preserved as cemented float; the original source of the block could not be identified. The trackway is preserved as nine large distinct depressions with no obvious push-back mounds (cf. D. gouldi; Table , Fig. H). The direction of travel is impossible to determine, and the trackway may be an undertrack in which smaller appendage impressions are not preserved (see Goldring & Seilacher Citation1971).
Remarks
This trackway has dimensions that are slightly larger than those described above for Diplichnites gouldi (Table ), and because it lacks smaller appendage imprints it is provisionally retained in a separate informal ichnospecies. However, owing to its different style of preservation (as a probable undertrack), the apparent differences may be taphonomic. An arthropod trackmaker is proposed, but a more precise biological affiliation is not possible based on the available evidence.
Small indeterminate trail (Fig. )
Description
This incomplete linear trail is 242 mm long, 23 mm in maximum width, is truncated by a rock fracture at one end and dissipates via weathering at the other (Fig. A). The ichnofossil appears to be preserved as an undertrack, with all components of the trace preserved in positive relief (Fig. B). The trail is complex, consisting of weakly repeating sets of individual elements whose centres are ca 30–38 mm apart (Fig. C). The centre of the trail is defined by discontinuous sinuous traces (mostly lenticular mounds) that are 2 mm wide and ca 9–11 mm long. This central trace is flanked closely on either side by one or two paired arcuate or reniform elements up to 2 mm wide and 10 mm long. Around 8 mm beyond the margins of these elements are sets of roughly circular mounds ca 2 mm in diameter and 11–17 mm apart longitudinally. These outermost elements are weakly preserved or absent on one side of the trail.
Remarks
This trail is notably smaller than the examples of Diplichnites described above. It also differs in the arrangement and shape of the individual trace components. The direction of movement of the trail producer is difficult to ascertain as there are no clear signs of ‘push-up’ mounds. However, in a few cases, individual imprints appear to be deeper on the right side (Fig. C) suggesting (if this is an undertrack) that movement was to the left of the illustrated block.
The relatively small size and multiple discrete elements making up the trail suggest an arthropod producer. However, the sinuous form of the central trace is atypical of arthropods. This small trail also lacks the clusters of multiple sharply discrete lateral appendage imprints of early insects (Minter & Braddy Citation2006) and most other arthropods. Nevertheless, the Kings Canyon trace are broadly similar in size and complexity to some specimens of Stiaria intermedia Smith, Citation1909 from the Mississippian of USA that consist of repeating sets of central linear, inner flanking lobate and marginal circular elements that have been tentatively attributed to apterygote insects, perhaps walking within a moving fluid (Fillmore et al. Citation2012).
We can not exclude a small fish as a producer of this ichnofossil, in which case the weakly sinuous to reniform character of the central and lateral traces might reflect fin-tip furrows (cf. Wisshak et al. Citation2004). Among roughly coeval ichnofossil assemblages from Australia, Siskemia sp. of Trewin & McNamara (Citation1994) has a straighter and more continuous central ridge and lacks the inner zone of arcuate structures, and the outer zone has smaller and more densely spaced appendage impressions. There are also some similarities in terms of repeated lobate elements with putative fish traces from the Lower Triassic of Australia (Chen et al. Citation2012).
?Laevicyclus isp. Weber & Braddy, Citation2003 (Fig. )
Description
These traces occur on the bedding plane at the foot of the metallic staircase leading up from the eastern cliff of the ‘Garden of Eden’ walk (24°14ʹ56.46″S, 131°34ʹ59.71″E) and were originally identified but not described by Bagas (Citation1988). These epichnial traces range from circular to D-shaped with diameters between 68 and 189 mm (Fig. ). The traces consist of a peripheral or coiled burrow 10–30 mm wide, surrounding a central bowl-shaped depression.
Remarks
The coiled burrow may represent a horizontal feeding trace and the depression may have formed by sediment collapse into a central cavity. Until vertical profiles of these burrows are examined, the three-dimensional structure, mode of formation and producer remain speculative. Weber & Braddy (Citation2003) identified similar but much smaller traces ascribed to ?Laevicyclus isp. from Lower Ordovician shallow marine strata of Antarctica.
Beaconites antarcticus (Vialov) emend. Keighley & Pickerill, Citation1994 (Fig. )
Description
Horizontal to sub-horizontal, straight or gently curved, parallel-sided, cylindrical burrows reaching >150 mm long and 5–13 mm in diameter (Fig. A, C). The burrows occur in fine- to medium-grained sandstone and have an infilling dominated by grains similar in size to the surrounding matrix but divided into weak compartments of unequal size by silty meniscate laminae (Fig. B, D). The burrows have a distinct lining, <1 mm thick, defined by a finer grainsize (silt) and/or different colour to the remainder of the fill (Fig. B, D). The burrows are variably orientated within the same exposure (Fig. A, C).
Remarks
Small, roughly horizontal, lined burrows of this type are relatively common in the Mereenie Sandstone and are attributed to Beaconites antarcticus (Vialov Citation1962) emend. Keighley & Pickerill, Citation1994. This ichnotaxon was originally described from Devonian fluvial strata of Antarctica (Vialov Citation1962) but, subsequently, has been reported from a broad range of ages and depositional environments (Keighley & Pickerill Citation1994 and references therein).
Burrows similar or equivalent to B. antarcticus are common in eolian sandsheet and pond facies of the Tumblagooda Sandstone, Carnarvon Basin, Western Australia (Trewin & McNamara Citation1994) and this taxon is a key element of Bradshaw’s (Citation1981) ‘Beaconites Ichnocoenosis’ recognized in Devonian fluvial to paralic deposits of Victoria Land, Antarctica. Beaconites antarcticus and Taenidium barretti are commonly associated in mid-Palaeozoic fluvial deposits globally (Bradshaw Citation1981, Allen & Williams Citation1981, O’Sullivan et al. Citation1986, Marriott et al. Citation2009), and both are represented in the Mereenie Sandstone. The identity of the B. antarcticus burrow maker is uncertain, but such traces have generally been attributed to small arthropods or, less commonly, to annelids and even holothurians (Bradshaw Citation1981, Weber & Braddy Citation2003).
Taenidium barretti (Bradshaw) emend. Keighley & Pickerill, Citation1994 (Fig. )
Description
Large, more or less straight, horizontal to oblique burrows filled by muddy sand within beds of fine-grained sandstone. The burrows are up to 470 mm long and 27–84 mm wide, irregularly orientated (Fig. A) and variably isolated (Fig. B) or clustered (Fig. C). The burrows are weakly septate, incorporating poorly developed and somewhat irregular meniscate backfill compartments that are ca 20–25 mm wide (Fig. D, E, H). Regular complex arcuate or en echelon markings are developed along the margins of several burrows repeating at intervals of 10–25 mm depending on the size of the burrow (Fig. G, I). A few burrows are devoid of surface markings (Fig. B), and one bears fine transverse striations (Fig. F). The burrows are preserved in the same beds as small vertical burrows (?Skolithos sp.; Fig. C) and are locally associated with extensive asymmetrical ripple fields (Fig. A).
Remarks
Originally placed in Beaconites Vialov, Citation1962 by Bradshaw (Citation1981), this ichnospecies was transferred to Taenidium by Keighley & Pickerill (Citation1994) on the basis that, although menisci merge at the burrow margins, no distinctive burrow lining is developed. Those burrows lacking obvious surface markings in the Mereenie Sandstone may have been subject to pedestrian attrition. The fine transverse striations on one specimen (Fig. F) are interpreted to represent current lineations on a partially exhumed burrow. Most specimens are marked by somewhat irregular meniscate features that are linked to repetitive arcuate marginal traces interpreted to represent sediment push-back traces produced by arthropod appendages. The Cruziana-like surface striations developed in a few cases (Fig. C) also favour burrow production by a large arthropod (cf. examples illustrated by CitationGibb et al. 2009). Indeed, the segmentation evident in some burrows (Fig. H) is reminiscent of the dorsal tergite impressions of a large arthropod (cf. Fayers et al. Citation2010, pl. 1).
The burrow illustrated by Bagas (Citation1988, pl. 12) from the Mereenie Sandstone appears to be referable to the same ichnotaxon as the examples described here. Similar burrow forms probably produced by large arthropods are relatively common in Siluro-Devonian strata of eastern Gondwana (Webby Citation1968, Bradshaw Citation1981, Trewin & McNamara Citation1994, Draganits et al. Citation2001, Gouramanis et al. Citation2003). As noted by Hocking (Citation1991) and Trewin & McNamara (Citation1994), morphologically similar traces described from diverse localities and ages may have been produced by a range of burrowing organisms. However, the similarity in size to Diplichnites gouldi and preservation in the same successions but within separate facies may reflect different behavioural aspects of the same large arthropod (Fayers et al. Citation2010).
Small burrow openings (Fig. C)
Remarks
Small openings to probable vertical burrows are present on the bedding surfaces where the examples of Taenidium barretti (e.g., Fig. C) are preserved. These traces occur in clusters of several closely spaced openings and commonly in pairs. Unfortunately, vertical profiles of these burrows were not observed, thus making ichnotaxonomic classification impossible, although ichnospecies of Arenicolites, Monocraterion or Skolithos can not be excluded.
Burrow entrance with radial feeding traces (Fig. )
Description
Circular trace fossil (in plan view) exposed on the upper surface of a sandstone bed (Fig. A). Trace having a total diameter of 37 mm, incorporating a central featureless zone (15 mm in diameter); outer zone ca 10–12 mm consisting of more or less regular and radially arranged alternating troughs and ridges with rounded crests (ca 20 ridges; Fig. B). Troughs broaden and deepen slightly with distance from the centre of the trace. The outer margin of the trace is entire or slightly lobate. The trace is preserved entirely in fine-grained sandstone with no obvious lithological difference between the central zone, ridges or troughs.
Remarks
This circular trace has a superficial resemblance to a medusoid cnidarian. However, most medusoid impression fossils are characterized by concentric contractional muscle or desiccation features. There are also superficial similarities to the putative Silurian vendobiont Rutgersella (Retallack Citation2015) in having a central disorganized zone surrounded by a broad rim of radial grooves and ridges. However, Rutgersella differs in being typically elliptical with finer radial components that bifurcate sporadically. The Kings Canyon trace best matches the general characters of the un-named stellate trace fossils illustrated by Collette et al. (Citation2011), although the specific dimensions of the new trace do not match previous examples. The Kings Canyon trace is also reminiscent of an ichnofossil illustrated by Shi et al. (Citation2009) and assigned to ?Lorenzinia isp., which consists of radially arranged epichnial ridges surrounding a featureless central zone. However, the latter has only five radial ridges and occurs in bathyal turbiditic deposits. There are also similarities with Dactyloidites Hall, Citation1886 in its radial form. However, Dactyloidites is generally interpreted to represent a vertical or oblique burrow with a subterranean (i.e., basal) rosette of feeding tunnels (Wilmsen & Niebuhr Citation2014). By contrast, the Kings Canyon trace is interpreted to represent a central vertical or oblique burrow with a rosette of radial feeding traces on the bedding surface around the burrow mouth. This is consistent with the interpretation by Collette et al. (Citation2011) of Early Jurassic stellate ichnofossils from ephemeral lacustrine deposits that are similar to the modern feeding traces of infaunal bivalves, such as Macoma tenta. Without additional specimens and vertical sections, we can not determine if there are multiple levels of radial features in this trace as represented in some examples of Dactyloidites (Fürsich & Bromley Citation1985).
Discussion and conclusions
The age of the Mereenie Sandstone is poorly constrained owing to a dearth of body fossils and the presence of unconformities at the base and top of the unit. The Mereenie Sandstone overlies the Carmichael Sandstone dated as Late Ordovician (Wells et al. Citation1970), and underlies the Park Siltstone of Middle to Late Devonian age (Young Citation1985). The similarities between the ichnofaunas of the Mereenie Sandstone and those of the Tumblagooda Sandstone (Trewin & McNamara Citation1994), Major Mitchell Sandstone (Gouramanis et al. Citation2003), Mt Daubeny Formation (Neef Citation2004a) and Ravendale Formation (Neef Citation2004b) elsewhere in Australia favour a roughly contemporaneous (late Silurian–early Devonian) age of deposition.
The Mereenie Sandstone ichnofauna occurs in a succession that hosts extensive shallow-water sedimentary structures (trough, tabular and low-angle cross-bedding, and wave- and current-ripple cross-laminations; Bagas Citation1988). Some trackways appear to be developed on emergent surfaces with thin silty coatings. Other traces include vertical or sub-horizontal burrows that were clearly produced by subsurface activities in unconsolidated sediment. A few traces appear to be influenced by active water currents during or after their formation. The co-occurrence of Diplichnites, Beaconites, Taenidium and Skolithos-like traces appears to represent a typical ichnofossil association of mid-Palaeozoic paralic to fluvial successions (Bradshaw Citation1981, Trewin & McNamara Citation1994, Gouramanis et al. Citation2003, Marriott et al. Citation2009), although some similar assemblages may be found in strata as old as the Early Ordovician (Weber & Braddy Citation2003).
The ichnofaunal assemblage of the Mereenie Sandstone represents an expression of the development of complex ecosystems at the interface of terrestrial and aquatic environments during the mid-Palaeozoic. The assemblage highlights the preliminary steps leading to the more extensive terrestrialization of animals and plants that developed later in the Devonian. Our reconnaissance study of the Mereenie Sandstone ichnofauna highlights the probable presence of several clades of organisms invading paralic to fluvial environments and engaging in a diverse array of behaviours. We contend that future systematic surveys of the extensive exposures of this rock unit will probably reveal more diverse ichnofossil suites that, in turn, will provide a clearer perspective of the early steps towards biotic occupation of freshwater ecosystems and the land surface in the mid-Palaeozoic. In particular, this formation has strong potential for elucidating the initial enchroachment of arthropods (MacNaughton et al. Citation2002) and even early tetrapodomorphs (e.g., Niedźwiedzki et al. Citation2010) into shallow river systems and onto land in the Siluro-Devonian palaeoequatorial realm.
Acknowledgements
We thank Shannon Carne of the Watarrka Ranger Station for providing GPS coordinates of the trackways and Becky Rush who helped photograph the trackways. We thank Vivi Vajda for providing several of the images used in Figs 6 and 7. We also thank two reviewers whose comments helped improve the quality of the manuscript. This paper is Earth Observatory of Singapore Contribution Number 92. SM acknowledges funding from the Swedish Research Council (VR grant 2014-5234).
Related Research Data
References
- Allen, R.J.L. & Williams, B.P.J., 1981. Beaconites antarcticus: a giant channel-associated trace fossil from the Lower Old Red Sandstone of South Wales and the Welsh Borders. Geological Journal 16, 255–269.
- Bagas, L., 1988. Geology of Kings Canyon National Park. Northern Territory Geological Survey, Darwin, Report 4, 1–21.
- Bradshaw, M.A., 1981. Paleoenvironmental interpretations and systematics of Devonian trace fossils from the Taylor Group (lower Beacon Supergroup), Antarctica. New Zealand Journal of Geology and Geophysics 24, 615–652.
- Bradshaw, M.A., 2010. Devonian trace fossils of the Horlick Formation, Ohio Range, Antarctica: systematic description and palaeoenvironmental interpretation. Ichnos 17, 58–114.
- Briggs, D.E.G., Rolfe, W.D.I. & Brannan, J., 1979. A giant myriapod trail from the Namurian of Arran, Scotland. Palaeontology 22, 273–291.
- Chen, Z.-Q., Fraiser, M.L. & Bolton, C., 2012. Early Triassic trace fossils from Gondwana Interior Sea: implication for ecosystem recovery following the end-Permian mass extinction in south high-latitude region. Gondwana Research 22, 238–255.
- Collette, J.H., Getty, P.R. & Hagadorn, J.W., 2011. Insights into an Early Jurassic dinosaur habitat: ichnofacies and enigmatic structures from the Portland Formation, Hoover Quarry, Massachusetts, U.S.A. Atlantic Geology 47, 81–98.
- Draganits, E., Braddy, S.J. & Briggs, D.E.G., 2001. A Gondwanan coastal arthropod ichnofauna from the Muth Formation (Lower Devonian, Northern India): paleoenvironment and tracemaker behavior. Palaios 16, 126–147.
- Edwards, D. & Kenrick, P., 2015. The early evolution of land plants, from fossils to genomics: a commentary on Lang (1937) ‘On the plant-remains from the Downtonian of England and Wales’. Philosophical Transactions of the Royal Society B 370, 20140343. doi:10.1098/rstb.2014.0343.
- Fayers, S.R., Trewin, N.H. & Morrissey, L., 2010. A large arthropod from the lower Old Red Sandstone (Early Devonian) of Tredomen Quarry, South Wales. Palaeontology 53, 627–664.
- Fillmore, D.L., Lucas, S.G. & Simpson, E.L., 2012. Ichnology of the Mississippian Mauch Chunk Formation, eastern Pennsylvania. New Mexico Museum of Natural History and Science Bulletin 54, 1–136.
- Fürsich, F.T. & Bromley, R.G., 1985. Behavioural interpretation of a rosetted spreite trace fossil: Dactyloidites ottoi (Geinitz). Lethaia 18, 199–207.
- Gibb, S., Chatterton, B.D.E. & Pemberton, S.G., 2009. Arthropod ichnofossils from the Ordovician Stairway Sandstone of central Australia. Memoirs of the Association of Australasian Palaeontologists 37, 695–716.
- Goldring, R. & Seilacher, A., 1971. Limulid tracks and their sedimentological implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 137, 422–442.
- Gouramanis, C., Webb, J.A. & Warren, A.A., 2003. Fluviodeltaic sedimentology and ichnology of part of the Silurian Grampians Group, western Victoria. Australian Journal of Earth Sciences 50, 811–825.
- Gray, J., Laufeld, S. & Boucot, A.J., 1974. Silurian trilete spores and spore tetrads from Gotland: their implications for land plant evolution. Science 185, 260–263.
- Hagström, J. & Mehlqvist, K., 2012. The dawn of terrestrial ecosystems on Baltica: First report on land plant remains and arthropod coprolites from the upper Silurian of Gotland, Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology 317–318, 162–170.
- Hall, J., 1886. Note on some obscure organisms in the roofing slate of Washington County, New York, NY. New York State Museum of Natural History, 39th Annual Report, 160.
- Hocking, R.M., 1991. The Silurian Tumblagooda Sandstone, Western Australia. Geological Survey of Western Australia Report 27, 1–124.
- Johnson, E.W., Briggs, D.E.G., Suthren, R.J., Wright, J.L. & Tunnicliff, S.P., 1994. Non-marine arthropod traces from the subaerial Ordovician Borrowdale Volcanic Group, English Lake District. Geological Magazine 131, 395–406.
- Keighley, D.G. & Pickerill, R.K., 1994. The ichnogenus Beaconites and its distinction from Ancorichnus and Taenidium. Palaeontology 37, 305–337.
- MacNaughton, R.B., Cole, J.M., Dalrymple, R.W., Braddy, S.J., Briggs, D.E.G. & Lukie, T.D., 2002. First steps on land: arthropod trackways in Cambrian-Ordovician eolian sandstone, southeastern Ontario, Canada. Geology 30, 391–394.
- Marriott, S.B., Morrissey, L.B. & Hillier, R.D., 2009. Trace fossil assemblages in upper Silurian tuff beds: evidence of biodiversity in the Old Red Sandstone of southwest Wales, UK. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 160–172.
- Mehlqvist, K., Wigforss-Lange, J. & Vajda, V., 2015. A palynological study from Sweden reveals stable terrestrial environments during Late Silurian extreme marine conditions. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 105, 149–158.
- Minter, N.J. & Braddy, S.J., 2006. Walking and jumping with Palaeozoic apterygote insects. Palaeontology 49, 827–835.
- Neef, G., 2004a. Non-marine ?Late Silurian-Early Devonian trace fossils, Darling Basin, western New South Wales. Alcheringa 28, 389–399.
- Neef, G., 2004b. Devonian arthropod trackways from the fluvial Ravendale Formation, western New South Wales. Alcheringa 28, 401–402.
- Niedźwiedzki, G., Szrek, P., Narkiewicz, K., Narkiewicz, M. & Ahlberg, P.E., 2010. Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43–48.
- O’Sullivan, M.J., Cooper, M.A., MacCarthy, I.A.J. & Forbes, W.H., 1986. The palaeoenvironment and deformation of Beaconites-like burrows in the Old Red Sandstone at Gortnabinna, SW Ireland. Journal of the Geological Society of London 143, 897–906.
- Pickett, J.W., Burrow, C.J., Holloway, D.J., Munson, T.J., Percival, I.G., Rickards, R.B., Sherwin, L., Simpson, A.J., Strusz, D.L., Turner, S. & Wright, A.J., 2000. Silurian palaeobiogeography of Australia. Memoirs of the Association of Australasian Palaeontologists 23, 127–165.
- Retallack, G.J., 2009. Cambrian-Ordovician non-marine fossils from South Australia. Alcheringa 33, 355–391.
- Retallack, G.J., 2015. Reassessment of the Silurian problematicum Rutgersella as another post-Ediacaran vendobiont. Alcheringa 39, 573–588.
- Shi, G.R., Gong, Y.-M. & Potter, A., 2009. Late Silurian trace fossils from the Melbourne Formation, Studley Park, Victoria, southeastern Australia. Alcheringa 33, 185–209.
- Smith, A., Braddy, S.J., Marriott, S.B. & Briggs, D.E.G., 2003. Arthropod trackways from the Early Devonian of South Wales: a functional analysis of producers and their behaviour. Geological Magazine 140, 63–72.
- Smith, J., 1909. Upland Fauna of the Old Red Sandstone Formation of Carrick, Ayrshire. A.W. Cross, Kilwinning, 60 pp.
- Talent, J.A., Mawson, R., Aitchison, J.C., Becker, R.T., Bell, K.N., Bradshaw, M.A., Burrow, C.J., Cook, A.G., Dargan, G.M., Douglas, J.G., Edgecombe, G.D., Feist, M., Jones, P.J., Long, J.A., Phillips-Ross, J.R., Pickett, J.W., Playford, G., Rickards, R.B., Webby, B.D., Winchester-Seeto, T., Wright, A.J., Young, G.C. & Shen, Y.-Y., 2000. Devonian palaeobiogeography of Australia and adjoining regions. Memoirs of the Association of Australasian Palaeontologists 23, 167–257.
- Trewin, N.H. & McNamara, K.J., 1994. Arthropods invade the land: trace fossils and palaeoenvironments of the Tumblagooda Sandstone (?late Silurian) of Kalbarri, Western Australia. Transactions of the Royal Society of Edinburgh: Earth Sciences 85, 177–210.
- Vialov, O.S., 1962. Problematica of the Beacon Sandstone at Beacon Height West, Antarctica. New Zealand Journal of Geology and Geophysics 5, 718–732.
- Walter, M.R., Elphinstone, R. & Heys, G.R., 1989. Proterozoic and Early Cambrian trace fossils from the Amadeus and Georgina Basins, central Australia. Alcheringa 13, 209–256.
- Webby, B.D., 1968. Devonian trace fossils from the Beacon Group of Antarctica. New Zealand Journal of Geology and Geophysics 11, 1001–1008.
- Weber, B. & Braddy, S.J., 2003. A marginal marine ichnofauna from the Blaiklock Glacier Group (?Lower Ordovician) of the Shackleton Range, Antarctica. Transactions of the Royal Society of Edinburgh: Earth Sciences 94, 1–20.
- Wells, A.T., Forman, D.J., Ranford, L.C. & Cook, P.J., 1970. The geology of the Amadeus Basin, central Australia. Bureau of Mineral Resources, Geology and Geophysics, Australia, Bulletin 100, 1–222.
- Wilmsen, M. & Niebuhr, B., 2014. The rosetted trace fossil Dactyloidites ottoi (Geinitz, 1849) from the Cenomanian (Upper Cretaceous) of Saxony and Bavaria (Germany): ichnotaxonomic remarks and palaeoenvironmental implications. Paläontologische Zeitschrifte 88, 123–138.
- Wisshak, M., Volohonsky, E. & Blomeier, D., 2004. Acanthodian fish trace fossils from the Early Devonian of Spitsbergen. Acta Palaeontologica Polonica 49, 629–634.
- Woolfe, K.J., 1993. Devonian depositional environments in the Darwin Mountains: marine or non-marine? Antarctic Science 5, 211–220.
- Young, G.C., 1985. New discoveries of Devonian vertebrates from the Amadeus Basin, central Australia. BMR Journal of Australian Geology and Geophysics 9, 239–254.