
Abstract
The hyolith fauna of the middle Bystraya Formation (Cambrian Series 2) of eastern Transbaikalia (Zabaykalsky Krai), Siberia, includes at least 17 described taxa distributed between the orders Hyolithida and Orthothecida. Species of Microcornus, Parkula, Conotheca, Neogloborilus and Cupitheca are widely reported from Cambrian strata elsewhere. Triplicatella uslonica sp. nov. is known only from Transbaikalia. Dauritheca is proposed as a replacement generic name for the junior homonym Pachytheca, originally described from Xinjiang, China, and its range is extended to Transbaikalia. A conch with prominent comarginal ribs is referred to Salanytheca daurica sp. nov., a genus otherwise known from the early Cambrian of Mongolia and northern Siberia.
John S. Peel [[email protected]], Department of Earth Sciences (Palaeobiology), Uppsala University, Uppsala, Sweden; Alexander P. Gubanov [[email protected]], Museum of Evolution, Uppsala University, Uppsala, Sweden.
THE CAMBRIAN Bystraya Formation of eastern Transbaikalia (Zabaykalsky Krai) crops out in southeastern Siberia in a tectonically complex region near the common border with Mongolia and China (). It was originally established by Knyazev (Citation1962) but current usage follows Pakhomov (Citation1995). A rich fauna of small shelly fossils is known from the middle member of the formation, some of which has been described by Vologdin & Fonin (Citation1966), Melnikova (Citation1988), Ushatinskaya (Citation1988), Vasil’eva [Vassiljeva] & Sayutina (Citation1988) and Parkhaev (Citation2004). The present paper describes hyoliths from the middle member at the locality Uslon, near the village of Georgievka (), and briefly illustrates some accompanying fossils ().
Figure 1. Geological and geographical information. A, Location of eastern Transbaikalia (Zabaykalsky Krai) and the village Georgievka. B, Cambrian stratigraphy in the Georgievka area (after Pakhomov Citation1995).
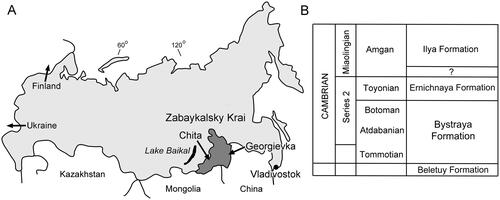
Figure 2. Small shelly fossils associated with hyoliths in sample 7000/68, Bystraya Formation, Uslon section, Georgievka, eastern Transbaikalia. A–D, Rhombocorniculum cancellatum (Cobbold, Citation1920). A, D, PMU 39243, with detail of tip (D). B, PMU 39241. C, PMU 39242. E, trilobite protaspid, PMU 39244. F, R, Monospinites Sayutina in Vasil’eva & Sayutina, Citation1988. F, PMU 39245. R, PMU 39246 with raised brim around basal perforation. G, Liangshanella sayutinae (Melnikova, Citation1988), PMU 392477. I, sponge spicule PMU 39248. J, V, Microdictyon sp., PMU 39249. H, Sphenothallus sp., PMU 39250. K, echinoderm plate PMU 39251. L, Pelagiella adunca Missarzhevsky in Rozanov & Missarzhevsky, Citation1966, PMU 39252, internal mould, lateral view with basal surface upwards. M, Parailsanella cf. lata Parkhaev in Gravestock et al., Citation2001, PMU 39253, internal mould. N, Chancelloria sp., PMU 39254. O, Yochelcionella crassa Zhegallo in Esakova & Zhegallo, Citation1996, PMU 39255, internal mould. P, Pojetaia runnegari Jell, Citation1980, PMU 39256, internal mould. Q, T, U, Archiasterella sp. Q, PMU 39257. T, PMU 39258. U, PMU 39259. S, Allonia sp., PMU 39260. Scale bars: 50 µm (V), 100 µm (A–D, H–J, L–N, R), 200 µm (F, G, K, O–Q, S–U).

Elucidation of the taxonomic relationships and classification of Cambrian hyoliths is complicated by the nature of their preservation. Most of the older literature and many contemporary studies are based on disarticulated macrofossils (cm size) preserved in calcareous or fine-grained siliceous strata, e.g., Holm (Citation1893), Syssoiev (Citation1958), Meshkova (Citation1974), Val’kov (Citation1975), Qian (Citation1978), Malinky (Citation1988, Citation1989, Citation2014), Berg-Madsen & Malinsky (Citation1999), Malinky & Berg-Madsen (Citation1999) and Malinky & Geyer (Citation2019). Classification of such material is often hindered by breakage or diagenetic flattening, but exquisitely preserved articulated specimens have also been described (Martí Mus & Bergström Citation2007, Martí Mus et al. Citation2014, Moysiuk et al. Citation2017, Peel & Willman Citation2018, Liu et al. Citation2021). Other, more recent studies focus mainly on microfossils (µm scale) that are usually recovered as disarticulated elements from residues of calcareous rocks after digestion in weak acids (e.g., Missarzhevsky in Rozanov et al. Citation1969, Qian Citation1989, Bengtson et al. Citation1990, Devaere et al. Citation2014, Pan et al. Citation2019). The general postmortal disarticulation of hyoliths has resulted in most taxa being described from isolated conchs or opercula, with the result that many historically described species based on conchs are poorly known (Malinky & Berg-Madsen Citation1999). However, the distinctive morphology of many isolated opercula from acid residues plays an increasingly important role in classification, especially within orthothecid hyoliths.
Concurrent descriptions of macro- and micro hyolith assemblages from the same formations are uncommon (Malinky & Skovsted Citation2004), but even here the ten-fold, or more, difference in size proves a difficult barrier to a common taxonomy. Peel et al. (Citation2016) traced the ontogeny of one Cambrian (Series 2, Stage 4) species from the Henson Gletscher Formation of North Greenland through a size range from 250 µm (acid residue) to 20 mm (knock out), but similar growth studies are rare. The extent to which individual assemblages from Cambrian acid residues comprise taxonomically distinct meiofaunas or microfaunal early ontogenetic stages of macrofauna (Waloszek Citation2003, Maas et al. Citation2006), and the influence of environmental and diagenetic variation generally remains to be established.
Background
Pakhomov (Citation1995) noted that the Bystraya Formation (, thickness 2620 m) consisted of three subformations and provided faunal lists. A lower subformation (620 m) is mainly composed of white and grey dolostone and has yielded fossil algae. The middle subformation (500 m), consisting of limestones with interbeds of shale and layers of phosphorite, yields a rich fauna of trilobites, archaeocyaths and small shelly fossils listed by Pakhomov (Citation1995). The upper subformation (1500 m) of dolostone with interbedded sandstone yields trilobites. Pakhomov (Citation1995) considered the Bystraya Formation to range in age from the Tommotian Stage to the Botoman Stage. It is evident from his faunal lists that fossils described by Melnikova (Citation1988), Ushatinskaya (Citation1988), Melnikova et al. Citation1997), Vasil’eva & Sayutina (Citation1988) and Parkhaev (Citation2004) were recovered from the middle subformation. Most were recovered from samples collected at Uslon, near Georgievka () by S.M. Sinitsa and T.A. Sayutina during 1982–1985, but some were derived from the nearby ‘archaeocyathan hill’ located midway between Uslon and Georgievka (Parkhaev Citation2004, fig. 1).
Vologdin & Fonin (Citation1966) described several endemic archaeocyaths from the Bystraya Formation in the Georgievka region. Debrenne et al. (Citation1989) noted that these archaeocyaths suggested an age of late Atdabanian Stage–Botoman Stage (Cambrian Series 2). Descriptions of obolellid brachiopods (Ushatinskaya Citation1988), bradoriid arthropods (Melnikova Citation1988, Melnikova et al. Citation1997) and chancelloriids (Vasil’eva & Sayutina Citation1988) were mainly based on samples collected at Uslon, considered to be from the lower Cambrian (Cambrian Series 2, Botoman Stage).
Parkhaev (Citation2004) gave full details of the geographical and geological derivation of the material collected by S.M. Sinitsa and T.A. Sayutina, recognizing seven members within the Bystraya Formation. Additionally, he also monographed the diverse mollusc fauna, with almost all fossils being derived from Members 5 and 6 (Parkhaev Citation2004, fig. 3). He followed Debrenne et al. (Citation1989) in suggesting an early Cambrian age (Cambrian Series 2, late Atdabanian Stage–Botoman Stage).
Materials and methods
Samples T2.6, 7000/44 and 7000/68 were collected by Alexander P. Gubanov during 2000 from the middle Bystraya Formation in the structurally inverted Uslon section (Parkhaev Citation2004, figs 1–3). The locality lies about 15 km west-southwest of the village of Georgievka (51°22′N, 119°27′E; ), which is situated about 70 km west of the River Argun marking the border between Transbaikalia and China (Parkhaev Citation2004, fig. 1). Sample T2.6 was collected from near the base of Member 3, close to sample 7000/28 of Parkhaev (Citation2004, figs 2, 3) from which he described just Pojetaia runnegari Jell, Citation1980 and Pelagiella adunca Missarzhevsky in Rozanov & Missarzhevsky, Citation1966. Parkhaev (Citation2004, fig. 3) placed sample 7000/44 near the base of Member 5 and considered both samples to be of Atdabanian Stage age. Sample 7000/68 was collected near the middle of Member 6 of the Bystraya Formation, just below the Atdabanian–Botoman boundary according to Parkhaev (Citation2004, figs 2, 3).
Residues of samples У1 and У2 (Cyrillic letters for U1 and U2) from Uslon were kindly donated by Elena A. Zhegallo during a visit to Uppsala in 1998. In terms of lithology and fauna, these two samples are closely similar to the residue of sample 7000/68, but the precise stratigraphic relationship of these three samples is not known.
Specimens were hand-picked from residues of limestone dissolved in weak acetic acid, prior to examination and imaging by scanning electron microscopy. Images were assembled in Adobe Photoshop CS4.
Abbreviations and repositories
PMU prefix indicates a specimen deposited in the palaeontological type collection of the Museum of Evolution, Uppsala University, Sweden. Other repositories are stated in the text.
Hyolith fauna
The described hyolith fauna of the middle Bystraya Formation contains 17 described taxa distributed between the orders Hyolithida and Orthothecida. Species of Microcornus Mambetov, Citation1972, Parkula Bengtson in Bengtson et al. Citation1990, Conotheca Missarzhevsky in Rozanov et al. Citation1969, Neogloborilus Qian & Zhang, Citation1983 and Cupitheca Duan, Citation1984 have been widely reported from Cambrian strata on several palaeocontinents (Qian Citation1989, Bengtson et al. Citation1990, Malinky & Berg-Madsen Citation1999, Malinky & Skovsted Citation2004, Devaere et al. Citation2014, Kouchinsky et al. Citation2015, Citation2017, Citation2022, Skovsted et al. Citation2016, Malinky & Geyer Citation2019, Pan et al. Citation2019). Triplicatella Conway Morris in Bengtson et al. Citation1990 was originally described from the early Cambrian Parara Limestone of Australia (Bengtson et al. Citation1990, Gravestock et al. Citation2001, Skovsted et al. Citation2014). Liu et al. (Citation2021) listed six species of Triplicatella, with notable occurrences in Laurentia (Skovsted et al. Citation2004, Skovsted & Peel Citation2007), the Siberian platform (Kouchinsky et al. Citation2015) and China (Pan et al. Citation2019, Liu et al. Citation2021). Triplicatella uslonica sp. nov. is currently known only from Transbaikalia but the triplicatellid Dauritheca (new name) is known elsewhere only from the original description from Xinjiang, China under the pre-occupied name Pachytheca Qian, Yin & Xiao (Citation2000). The genus Salanytheca Missarzhevsky, Citation1981, represented in the Uslon section by a new species Salanytheca daurica sp. nov., was originally described from Mongolia (Missarzhevsky Citation1981), but is also recognized in the fauna of the Emyaksin Formation of northern Siberia described by Kouchinsky et al. (Citation2015).
Accompanying fauna
Comments concerning the fauna associated with the Uslon hyoliths mainly relate to illustrations of specimens from sample 7000/68, the largest available residue ().
Parkhaev (Citation2004, fig. 3) described 10 species of molluscs from the Bystraya Formation, some of which were later discussed by Parkhaev (Citation2006, Citation2007) and Vendrasco et al. (Citation2011). Of these, only Pelagiella adunca was recorded from sample 7000/68. In addition to Pelagiella (), the currently available material from that sample includes Pojetaia runnegari (), Yochelcionella crassa Zhegallo in Esakova & Zhegallo, Citation1996 () and Parailsanella cf. lata Parkhaev in Gravestock et al. Citation2001 (), all of which occur in underlying and overlying samples in the Uslon section (Parkhaev Citation2004, fig. 3).
Chancelloriids are common in sample 7000/68. Vasil’eva & Sayutina (Citation1988) described several forms from Uslon, Georgievska. Most notable amongst these is Monospinites Sayutina in Vasil’eva & Sayutina, Citation1988, with type species Monospinites simplex Sayutina in Vasil’eva & Sayutina, Citation1988, which was considered to be a single, barrel-shaped sclerite tapering to a distal spine, with a broad basal foramen. Vasil’eva & Sayutina (Citation1988) noted that Monospinites simplex most commonly occurred as internal moulds but these lacked any indication of contact with surrounding rays. Their illustrations (Vasil’eva & Sayutina Citation1988, pl. 31, fig. 4a–c) indicate an elongate specimen similar to but fine details are lacking. Esakova & Zhegallo (Citation1996) described “Monospinites” sp. from Cambrian Series 2 (Botoman Stage) at Nuur-Mogoy, Mongolia. Bokova & Vasil’eva (Citation1990) named a second species, Monospinites piriformis Bokova & Vasil’eva, Citation1990 from the lower Cambrian of the Olenek Uplift (see also Novozhilova & Korovnikov, Citation2019), but their illustrations suggest the presence of facets around the base of the spine, indicating that it was the axial ray of a multi-rayed chancelloriid. Demidenko in Parkhaev & Demidenko (Citation2010, p. 903) considered that Monospinites simplex represented the central spine of a stellate chancelloriid and placed the genus in synonymy with Chancelloria Walcott, Citation1920. Moore et al. (Citation2014) noted Demidenko’s opinion but considered that other material placed in the genus by Vasil’eva (Citation1998) seemed to represent genuine one and two-rayed sclerites (Moore et al. Citation2010). Internal moulds of Monospinites simplex with spines of different lengths are common in sample 7000/68 () and many, but not all, do not show facets indicating contact with the radiating rays of Chancelloria (). However, such facets are not always present on internal moulds of the axial ray in articulated stellate chancelloriids. Less common specimens from sample 7000/68 preserve the outer wall, but flattened surfaces indicating contact with radial rays have not been observed in these (). Stellate Chancelloria sclerites with conjoined axial and radial rays () are infrequent in sample 7000/68 but their axial rays have essentially straight sides without the finely tapering spine of Monospinites simplex.
The chancelloriid Cambrothyra Moore, Porter, Steiner & Li, Citation2010 from the early Cambrian of China has variable, globose, sclerites that are usually isolated, or rarely paired, rather than grouped into rosettes, and in this feature are comparable to the concept of Monospinites advanced by Vasil’eva & Sayutina (Citation1988). They lack the pointed apex of Monospinites from Uslon () and generally develop a channel, the basal facet of Moore et al. (Citation2010, fig. 3), around the foramen in contrast to the ridge around the foramen often seen in many specimens of Monospinites () and other chancelloriids ().
Vasil’eva & Sayutina (Citation1988) also described Chancelloria tuberculata Sayutina sp. nov., Stellaria sp. and Archiasterella tetractina Vasil’eva & Sayutina sp. nov. from Uslon. Spicules of Archiasterella Sdzuy, Citation1969 are common in sample 7000/68 (), particularly a triradiate form with a globose upturned ray (). Specimens of Allonia Doré & Reid, Citation1965 with three or four upturned rays are also numerous ().
Melnikova (Citation1988) and Melnikova et al. (Citation1997) described three bradoriid species from samples from the Uslon section. The current sample contains rare specimens of Liangshanella sayutinae (Melnikova, Citation1988; ), a species also recorded from the Bastion Formation of North-East Greenland by Skovsted (Citation2006). Trilobites identified from the Bystraya Formation were listed by Melnikova (Citation1988) and Pakhomov (Citation1995). The residue of sample 7000/68 contains unassigned protaspids () and meraspid pygidia.
New records in sample 7000/68 from Uslon include Rhombocorniculum cancellatum (Cobbold, Citation1920), which is represented by several of the morphotypes illustrated by Kouchinsky et al. (Citation2015) from the Emyaksin Formation of northern Siberia in their revision of the genus (). Rare sclerites () are similar to Microdictyon cf. M. rhomboidale Bengtson, Matthew, & Missarzhevsky, Citation1986, as illustrated by Kouchinsky et al. (Citation2015, fig. 55) from the Emyaksin Formation of northern Siberia. Abundant holdfasts of Sphenothallus sp. () are similar to specimens described by Peel (Citation2021a) from the Silurian of North Greenland. Missarzhevsky & Mambetov (Citation1981) referred similar holdfasts from the lower Cambrian of Maly Karatau, but with a proportionately wider stem, to a new species, Torellella explicata Missarzhevsky & Mambetov, Citation1981.
Systematic palaeontology
Phylum uncertain
Class HYOLITHA Marek, Citation1963
Order HYOLITHIDA Syssoiev, Citation1957
Family UNDET.
Microcornus Mambetov, 1972
Type species.
Microcornus parvulus Mambetov, Citation1972 from the lower Cambrian of Maly Karatau.
Discussion.
Microcornus Mambetov, Citation1972 is globally distributed and widely reported from Cambrian Stages 3–5 (Pan et al. Citation2019). The conch is triangular in shape with a well-marked median dorsal ridge; the ventral surface is shallowly convex, with a semi-circular ligula. The operculum is not well known, but the convex conical shield is separated from the reduced and narrow, upturned cardinal shield by a weak concave furrow (Mambetov Citation1972, Yang et al. Citation2015, fig. 8K; Pan et al. Citation2019). An operculum with long clavicular ridges on the interior, attributed to Microcornus by (Yang et al. Citation2015, fig. 8M), was excluded from the genus by Pan et al. (Citation2019).
Microcornus eximius Duan, 1984
()
Figure 3. Hyolith conchs from sample 7000/68, Bystraya Formation, Uslon section, Georgievka, eastern Transbaikalia. A, C, E, G–I, K, N, O, Q, R, Microcornus eximius Duan, Citation1984. A, Q, PMU 39261. C, PMU 39262. E, PMU 39263. G, PMU 39264. H, PMU 39265. I, PMU 39266. K, R, PMU 39267. N, PMU 392658. O, PMU 39269. B, ?J, Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969. B, PMU 39270. ?J, PMU 39271. D, Microcornus? cf. delicatus He & Pei in He et al., Citation1984, PMU 39272. F, ‘Conotheca’ tenuis Missarzhevsky, Citation1989, PMU 39273. M, Parkula bounites Bengtson in Bengtson et al., Citation1990, PMU 39274, arrows indicate clavicle pits. L, P, Neogloborilus applanatus Qian & Zhang, Citation1983. L, PMU 39275. P, PMU 39276. Scale bars: 100 µm (A–D, H, J, K, M, N, Q), 200 µm (E–G, I, L, O, P, R).
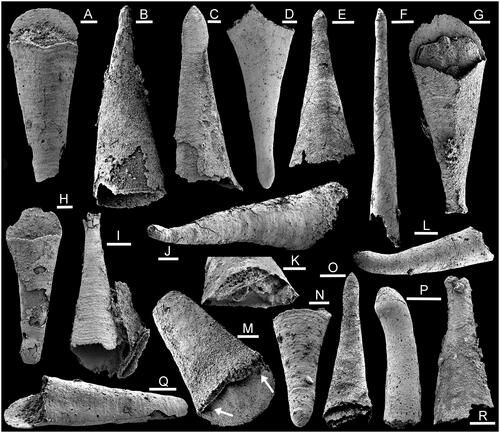
Figure 4. Hyoliths from the Bystraya Formation, Uslon section, Georgievka, eastern Transbaikalia. A, D, G, Microcornus eximius Duan, Citation1984. A, D, PMU 39292, from sample У1. G, PMU 39293, from sample 7000/44. B, Majatheca sp., PMU 39294, from sample У1. C, Operculum 2, PMU 39295, from sample У1. E, H, I, J, Triplicatella uslonica sp. nov., PMU 39296, holotype, from sample T2.6-1. F, Parkula bounites Bengtson in Bengtson et al., Citation1990, PMU 39297, from sample 7000/44. Scale bars: 100 µm, except C (200 µm).

1984 Microcornus eximius Duan, p. 153, pl. 1, fig. 5.
1990 Microcornus eximius; Bengtson in Bengtson et al., p. 221, fig. 148.
2019 Microcornus eximius; Pan et al., p. 370, fig. 9A–D.
(includes additional references)
Figured material.
PMU 39261–PMU 39269 from sample 7000/68; PMU 39292 from sample У1; PMU 39293 from sample 7000/44. Bystraya Formation, Uslon, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Full descriptions of this globally distributed and widely reported species from Cambrian Stages 3–5 were given by Bengtson et al. (Citation1990) and Pan et al. (Citation2019). Numerous conchs in samples 7000/68 and У2 agree closely with the illustrations given by Duan (Citation1984) from the Xihaoping Formation of Hubei Province, China and by Bengtson et al. (Citation1990) from the Parara Limestone of South Australia. In distinguishing conchs of Microcornus eximius from conchs of Parkula Bengtson in Bengtson et al., Citation1990, which also occurs in the Parara Limestone, Bengtson in Bengtson et al. (Citation1990) noted the more strongly developed median dorsal angulation in Microcornus, often culminating in a shallow sinus (), and triangular cross-section of the former. Additionally, the ligula of Microcornus eximius is semi-circular in plan view () but shorter with an elliptical margin in Parkula, although some specimens from Uslon have an intermediate form.
The degree of expression of the median dorsal angulation is variable in material from Uslon. The dorsal profile may be almost uniformly convex without the median sinus (), and the ventral surface may be almost flat (). However, the distinctive semi-circular ligula is retained. The apex is often absent due to crushing in larger specimens (), which may be a diagenetic effect of incomplete postmortal mineralization or reflect the presence of a septum preventing apical mineralization, although a septum has not been observed. Rare specimens preserve a shallowly concave longitudinal profile to the ventral surface () rather than the usual shallowly convex or flat form.
A malformed specimen from sample У1 shows repetition of shell form of the protoconch ().
Microcornus? cf. delicatus He & Pei in He et al., 1984
()
Figure 5. Hyolith conchs from the Bystraya Formation, sample У2, Uslon section, Georgievka, eastern Transbaikalia. A, D, E, Neogloborilus applanatus Qian & Zhang, Citation1983. A, E, PMU 39298, lateral and dorsal views. D, PMU 39299, lateral view. B, C, Microcornus? delicatus He & Pei in He et al., Citation1984, PMU 39300. F, Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969, PMU 39240. G, I, ‘Conotheca’ tenuis Missarzhevsky, Citation1989, PMU 39301. H, J, Neogloborilus cf. spinatus (Qian & Xiao, Citation1984). H, PMU 39302. J, PMU 39303. Scale bars: 100 µm, except F (200 µm).

Figured material.
PMU 39272 from sample 7000/68. PMU 39300 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
In terms of the increasing rate of expansion of the conch to about 40° near the aperture, and resultant concave lateral margins in ventral view (), these specimens are compared to Parakorilithes delicatus He & Pei in He et al. (Citation1984, pl. 1, fig. 24) from the Xinji Formation of Henan Province, China, although this poorly known species is tentatively assigned to Microcornus. The ligula appears to be more elliptical in dorsal view () when compared to the semi-circular shape in Microcornus eximius, but this difference is emphasized by breakage. Similarly shaped conchs are developed in Paracornus poulseni Malinky & Skovsted, Citation2004 from the Bastion Formation of North-East Greenland, but the apex in that species is not known. Paracornus poulseni is distinguished by rows of comarginal tubercles.
Li et al. (Citation2019, fig. 2) assigned a large (length 1.2 mm) internal mould from the Xinji Formation of China to Microcornus eximius Duan, Citation1984. The early growth stages are typical for the species. The conch expands rapidly (25–30° in dorsal view) in later growth but without the concave sides of Microcornus? cf. delicatus.
Parkula Bengtson in Bengtson et al., Citation1990
Type species.
Parkula bounites Bengtson in Bengtson et al., Citation1990, from the lower Cambrian Parara Limestone of South Australia.
Parkula bounites Bengtson in Bengtson et al., 1990
()
Figure 6. Hyolith opercula from the Bystraya Formation, sample У2, Uslon section, Georgievka, eastern Transbaikalia. A–D, Parkula bounites Bengtson in Bengtson et al., Citation1990. A, B, PMU 39304. C, D, PMU 39305. E, Neogloborilus cf. spinatus (Qian & Xiao, Citation1984), PMU 39306. F, I, Neogloborilus applanatus Qian & Zhang, Citation1983, PMU 39307. G, H, J, K, N, Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969. G, H, J, PMU 39308. J, detail of apex. K, N, PMU 39309. L, M, Operculum 3, PMU 39310. Scale bars: 100 µm, except G (50 µm) and J (20 µm).

1990 Parkula bounites Bengtson in Bengtson et al., p. 223, figs 149–151.
2019 Parkula bounites; Pan, Skovsted, Sun, & Li, p. 371, figs 10A, B, 11A–J.
(includes additional references).
Figured material.
PMU 39274 from sample 7000/68, PMU 39297 from sample 7000/44; PMU 39304 and PMU 39305 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Full descriptions were given by Bengtson et al. (Citation1990) and Pan et al. (Citation2019). A broken conch with the operculum in place, as a mould of the interior surface, is assigned to Parkula (). The upturned margin of the cardinal shield is almost straight, as in , but shows a slight dorsal deflection around the summit. Clavicles are preserved as round pits near the lateral margins (, arrows).
The exterior surface of the conical shield is convex, although appearing flattened in the illustration (). In plan view, the boundary between the conical shield and the upturned margin of the cardinal shield is almost straight, whereas it is strongly V-shaped, pointed towards the dorsal margin, with passage around the dorsal edge of the summit, in specimens from the Bastion Formation of North-East Greenland described by Malinky & Skovsted (Citation2004) or from the Aftenstjernesø Formation of North Greenland by Peel (Citation2021b). In this respect, the Uslon specimens more closely resemble type material of Parkula bounites from South Australia (Bengtson et al. Citation1990). In part, the difference between the specimens illustrated from Uslon and those from the Bastion Formation may be ontogenetically controlled, since the former are only about one fifth of the width of the latter.
The operculum of Parkula esmeraldina Skovsted, Citation2006 from the lower Cambrian Emigrant Formation of Nevada is distinguished by prominent, rounded, comarginal ribs along the ventral margin and more strongly protruding cardinal processes. Strongly protruding cardinal processes are also present in material from North-East Greenland and northern Siberia (Cambrian Stage 4) discussed by Pan et al. (Citation2019, p. 369).
Order ORTHOTHECIDA Marek, Citation1966
Family CIRCOTHECIDAE Missarzhevsky in Rozanov et al., Citation1969
Conotheca Missarzhevsky in Rozanov et al., Citation1969
Type species.
Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969, lower Cambrian (Tommotian Stage), Siberia.
Conotheca mammilata Missarzhevsky in Rozanov et al., 1969
(; ; ; )
Figure 7. Hyolith opercula from sample 7000/68, Bystraya Formation, Uslon section, Georgievka, eastern Transbaikalia. A–D, Dauritheca triangula (Qian, Yin & Xiao, Citation2000). A, PMU 39277. B, C, PMU 39278. D, PMU 39279. G, PMU 39280. E, F, Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969. E, PMU 39281. F, PMU 39282. H, Operculum 1, PMU 39283. I, ‘Conotheca’ tenuis Missarzhevsky, Citation1989, PMU 39284. Scale bars: 100 µm (D–H), 200 µm (B, C, I), 1 mm (A).

1969 Conotheca mammilata Missarzhevsky in Rozanov et al., p. 135, pl. 8, figs 2, 4, 7–8.
1989 Conotheca mammilata; Missarzhevsky, p. 157, pl. 4, figs 6, 10.
Figured material.
Conchs: PMU 39270 from sample 7000/68; PMU 39240 from sample У2. Opercula: PMU 39281 and PMU 39282 from sample 7000/68; PMU 39308 and PMU 38309 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Missarzhevsky in Rozanov et al. (Citation1969) commented on the variety of shell form in Conotheca mammilata Missarzhevsky in Rozanov et al., Citation1969, with slender specimens being more common than the shorter, more rapidly expanding specimens that are dominant from Uslon (). A specimen tentatively placed here displays a sigmoidal early growth stage ().
Conotheca mammilata and Conotheca australiensis Bengtson in Bengtson et al., Citation1990 are essentially distinguished on the state of knowledge concerning their operculum, which is well described in material assigned to the latter species (Bengtson et al. Citation1990, Malinky & Skovsted, Citation2004, Pan et al. Citation2019) but not in the former. The operculum of Conotheca mammilata illustrated by Missarzhevsky (Citation1989, pl. 4, fig. 6) has a pair of prong-like cardinal processes and a prominent comarginal clavicular ring that appears to be extended into narrow, blade-like clavicles in lateral areas. The operculum of Conotheca australiensis as illustrated by Bengtson et al. (Citation1990, fig. 144) has clavicles consisting of tubes or rods that slope at a low angle in towards the centre of the operculum from the clavicular ring. The clavicular rods are not visible in specimens from North-East Greenland that Malinky & Skovsted (Citation2004, fig. 14D) referred to Conotheca australiensis, although the clavicular ring is raised laterally and thus resembles Conotheca mammilata. Rods are present in Conotheca laurentiensis Landing & Bartowski, Citation1996, which Malinky & Skovsted (Citation2004) and Pan et al. (Citation2019) placed as a junior synonym of Conotheca australiensis. Peel (Citation2021b) recognized Conotheca laurentiensis from North and North-East Greenland, but on the basis of its unusually long cardinal processes.
Several types of isolated orthothecid opercula are present in the Uslon samples and a circular form is tentatively placed together with conchs referred to Conotheca mammilata. The outer surface is shallowly convex, with a sub-central apex that is located closer to the presumed dorsal margin (). The dorsal sector is slightly depressed relative to the ventrolateral flanks and the surface is ornamented with comarginal growth lines that are periodically prominent. The apical area is raised and smoothly convex, but with a median depression containing small pits penetrating the surface (). Details of the inner surface are partially obscured by diagenetic encrustation, but a flattened brim passes around a weak clavicular ring/angulation into a smooth central area (). A pair of closely spaced, short (possibly broken) cardinal processes rises perpendicular to the clavicular ridge near to the dorsal margin.
Kouchinsky et al. (Citation2015, fig. 28A, B) illustrated similar opercula from the Emyaksin Formation of northern Siberia that differ in having a flat outer surface and longer cardinal processes. Opercula of Conotheca australiensis from the Bastion Formation of North-East Greenland (Malinky & Skovsted Citation2004, fig. 14B, D, E) have a flat outer surface, more prominent clavicular ring and diverging cardinal processes. Type material of the same species from South Australia (Bengtson et al. Citation1990, fig. 144) has a shallowly convex outer surface and short cardinal processes, as does the Uslon material, but the better preserved type material displays clavicular rods extending inwards from the relatively subdued clavicular ridges.
‘Conotheca’ tenuis Missarzhevsky, 1989
()
1989 Conotheca tenuis Missarzhevsky, pl. 1, fig. 4 (non fig. 8).
Figured material.
PMU 39273 and PMU 39284 from sample 7000/68, PMU 39301 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Missarzhevsky (Citation1989) figured two Tommotian specimens of Conotheca tenuis Missarzhevsky, Citation1989 that do not appear to be conspecific. The designated holotype (Missarzhevsky, Citation1989, pl. 1, fig. 4) shows the same slightly sigmoidal form at the apex as a specimen from Uslon (), whereas the accompanying specimen has a dorsally curved tip (Missarzhevsky, Citation1989 pl. 1, fig. 8). The slender form and sigmoidal apex of the holotype does not compare well with the more rapidly expanding, straight to strongly curved conchs of other species of Conotheca () and the tentative reference of tenuis to that genus is therefore arbitrary.
The slender conchs from Uslon have an oval cross-section in contrast to the usually circular cross-section of Conotheca. In terms of their low rate of whorl expansion and straight to slightly sigmoidal form (in lateral profile), they can be compared with the early growth stages of Tulenicornus? frykmani Peel, Streng, Geyer, Kouchinsky & Skovsted, Citation2016 from the Henson Gletscher Formation (Cambrian Stage 4) of North Greenland. The hyolithid Microcornus petilus Bengtson in Bengtson et al., Citation1990 from the Parara Limestone of South Australia has a similar slender conch, but with a distinct ligula, and is ornamented with a distinctive lamellose transverse ornamentation (Bengtson et al. Citation1990, fig. 145).
Isolated specimens of an elliptical operculum () may belong to ‘Conotheca’ tenuis Missarzhevsky, Citation1989.
Family GLOBORILIDAE Syssoiev, Citation1957
Neogloborilus Qian & Zhang, 1983
Type species.
Neogloborilus applanatus Qian & Zhang, Citation1983, lower Cambrian of South China.
Neogloborilus applanatus Qian & Zhang, 1983
()
1983 Neogloborilus applanatus Qian & Zhang, p. 84, pl. 4, figs 10, 11.
2019 Neogloborilus applanatus; Pan et al., p. 359, figs 3A–J, 4A.
(includes additional references).
Figured material.
PMU 39275 and PMU 392756 from sample 7000/68; PMU 39298, PMU 39299 and PMU 39307 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Full descriptions of Neogloborilus applanatus Qian & Zhang, Citation1983 were given by Qian (Citation1989) and Pan et al. (Citation2019). The slender conchs from sample У2 at Uslon possess a sub-circular cross-section and an upturned, rounded, protoconch (). The poorly preserved inner surface of an operculum from sample У2 has an elliptical form with a prominent pair of divergent cardinal processes and narrow, acute, clavicles (); it is similar to specimens from the Houjiashan Formation of North China illustrated by Pan et al. (Citation2019, fig. 3I).
Wrona (Citation2003) illustrated conchs from the early Cambrian of King George Land, West Antarctica, that display a similar upturned and rounded apex to the Uslon specimens, but are slightly more robust. Wrona (Citation2003) referred the specimens and accompanying opercula to Conotheca but Pan et al. (Citation2019) considered that they were closer to Neogloborilus.
Globorilus globiger (Saito, Citation1936) from the Miaolingian Series (Wuliuan Stage) of Korea, has a similar globose protoconch but the conch quickly increases its diameter with growth, unlike the slender specimens from Uslon (Saito Citation1936).
Neogloborilus cf. spinatus (Qian & Xiao, 1984)
()
Figured material.
Conchs: PMU 39302 and PMU 39303 from sample У2. Operculum: PMU 39306 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
Neogloborilus spinatus (Qian & Xiao, Citation1984) and Neogloborilus applanatus from the lower Cambrian of China were recently revised by Pan et al. (Citation2019). The slowly expanding conch of the present specimens from Uslon has a circular cross-section and a shallowly sigmoidal apex, with the tip curved dorsally and bearing a short spine from its centre (). The conch surface is heavily etched but the orthocline apertural margin is evident from the transverse fibrous structure (). Specimens from the same sample assigned to Neogloborilus applanatus have a slender, less robust, conch with a rounded apex.
The single operculum tentatively placed here () is sub-circular, compared to the elliptical form of Neogloborilus applanatus (). The cardinal processes are closer together and less divergent than in Neogloborilus applanatus. The clavicles extend almost horizontally towards the ventral margin (), in which respect they resemble those of a specimen from the Aftenstjernesø Formation of Lauge Koch Land, North Greenland, illustrated by Peel (Citation2021b, fig. 9.1–9.3). The latter specimen, however, differs in the wider separation of the cardinal processes and the conical form of the internal surface of the operculum, equivalent to a deeply concave outer surface. The Uslon specimen has a shallowly concave inner surface and a flat or shallowly convex outer surface ().
Family TRIPLICATELLIDAE Liu, Skovsted, Topper & Zhang, 2021
Triplicatella Conway Morris in Bengtson et al., Citation1990.
Type species.
Triplicatella disdoma Conway Morris in Bengtson et al., Citation1990 from the Parara Limestone of South Australia (Cambrian Series 2).
Discussion.
Skovsted et al. (Citation2014) described a conjoined operculum and conch in the type species Triplicatella disdoma Conway Morris in Bengtson et al., Citation1990 from the Flinders Ranges of South Australia (Cambrian Series 2). They noted that helens were absent, which prompts placement of Triplicatellidae within Order Orthothecida. The conch displayed slight concave curvature along its dorsal surface, in lateral view, and was sub-circular in cross-section, as was the operculum.
Liu et al. (Citation2021) gave a thorough description of the morphology of Triplicatella, including soft anatomy, and listed six constituent named species. Additional species in open nomenclature were described by Skovsted et al. (Citation2004).
Triplicatella uslonica sp. nov.
()
Figure 8. Hyolith opercula from the Bystraya Formation, sample У2, Uslon section, Georgievka, eastern Transbaikalia. A–D, H, Dauritheca triangula (Qian, Yin & Xiao, Citation2000). A–D, PMU 39311. H, PMU 39312. E–G, Operculum sp. 4, PMU 39313. I, J, Triplicatella sp., PMU 39314. K, L, M, Triplicatella uslonica sp. nov., PMU 39315. Scale bars: 100 µm.

Holotype.
PMU 39296 from sample T2.6-1, Bystraya Formation, Uslon, near Georgievka, eastern Transbaikalia (Zabaykalsky Krai).
Other figured material.
PMU 39315 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Etymology.
From its occurrence at Uslon, near Georgievka, eastern Transbaikalia.
Diagnosis.
A species of Triplicatella with a sub-circular to dorsoventrally oval operculum with an excentric, obscure to round protoconch raised above the shorter, concave, marginal field, which lacks folds. A broad, rounded median fold with flanking shallow concavities extends from the apex to the opposite margin. Ornamentation of comarginal growth lines. Internal surface of operculum and conch not known.
Description.
Length is slightly greater than transverse width, with an excentric, smooth, protoconch () that is obscure in the holotype (). A prominent fold, producing a broad, deep sinus in the dorsal margin (), expands along the median plane of symmetry from the protoconch to the margin. The protoconch is raised high above the ventral area and the dorsolateral areas fall away rapidly from the crest of the dorsal fold towards the operculum margin (). Ornamentation consists of comarginal growth lines.
Discussion.
The sinuate margin is interpreted as dorsal by comparison with other opercula of Triplicatella and the co-occurring Dauritheca new name, which places the protoconch closest to the ventral margin (, ventral is left in the figure).
The operculum of Triplicatella disdoma, the type species of Triplicatella, has a prominent median fold extending from the summit to the dorsal margin but differs on account of three folds extending towards the ventral margin. Triplicatella sinuosa Skovsted, Peel & Atkins, Citation2004, originally described from the Bastion Formation of North-East Greenland and the Aftenstjernesø Formation of North Greenland by Skovsted et al. (Citation2004), is distinguished by four prominent folds in the dorsal margin.
Triplicatella peltata Skovsted, Peel & Atkins, Citation2004 from North and North-East Greenland usually has a transversely oval operculum and weak ventral folds (Skovsted et al. Citation2004, Skovsted & Peel Citation2007). Its apex is located closer to the dorsal margin in contrast with Triplicatella uslonica.
The operculum of Triplicatella xinjia, originally described by Pan et al. (Citation2019) from the Xinji Formation (Cambrian Series 2) of North China, differs in having a more transversely oblong shape, a central summit and prominent folds in the dorsal and the ventral margins (Pan et al. Citation2019, Peel Citation2021b). Triplicatella papilo Kouchinsky in Kouchinsky et al., Citation2015, from the upper Emyaksin Formation (Cambrian Series 2, Botoman Stage) of northern Siberia, differs in possessing two prominent folds in the dorsal margin.
Triplicatella sp.
()
Figured material.
PMU 39314 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
In plan view, this single operculum is wider than long, with a flattened ventral margin and an arched dorsal margin (). The smooth, round protoconch is located slightly closer to the dorsal margin, and is raised above the shallowly convex flanks, in contrast to Triplicatella uslonica where it is closer to the ventral margin. In lateral view, the margin is uniformly convex, indicating a concave apertural margin of the conch between the mid-dorsum and the venter, as described by Skovsted et al. (Citation2014, fig. 3) in Triplicatella disdoma. Three shallow folds, of which the central one is most strongly developed, extend from the protoconch to the dorsal margin. The outer surface is ornamented with comarginal growth lines. The inner surface and the associated conch are not known.
In plan view, Triplicatella sp. resembles Triplicatella sp. A of Skovsted et al. (Citation2004) from the Bastion Formation of Albert Heim Bjerge, North-East Greenland, although the protoconch in that species lies closer to the ventral margin. Triplicatella peltata usually has a transversely oval operculum and weak ventral folds (Skovsted et al. Citation2004, Skovsted & Peel Citation2007).
Dauritheca new name
Type species.
Pachytheca triangula Qian, Yin & Xiao, Citation2000 from the lower Cambrian Yuertus Formation of Xinjiang, China.
Etymology.
From Dauria, the region of the east Asian steppes extending from Tranbaikalia into eastern Mongolia and northern China.
Diagnosis.
Operculum sub-triangular in plan view, with the summit located between one third and half of the distance from the angular dorsal margin to the shallowly curved ventral margin; summit overhanging the shallowly concave dorsal field, which is crossed by a low median fold that expands towards the dorsal margin. External ornamentation of radiating ribs, which are coarser and more widely spaced in the dorsal field. Internal surface with raised, transverse, dorsal bar passing abruptly into shallowly concave median and ventral area. Conch not known.
Discussion.
Pachytheca Qian, Yin & Xiao, Citation2000, described as an orthothecid hyolith from the lower Cambrian Yuertus Formation of Xinjiang, China (Qian et al. Citation2000), is a junior homonym of Pachytheca Schlüter, Citation1885, a Devonian tabulocospongiid poriferan, as also are the bryozoan Pachytheca Canu, Citation1913 and the extant spider Pachytheca Schmidt, Citation2012. The only described species, Pachytheca triangula Qian, Yin & Xiao, Citation2000, is here transferred to Dauritheca, established as a new name to replace the homonym.
The opercula of most species of Triplicatella are distinguished by prominent folds that extend from the summit to the dorsal and ventral margins. Only a single, relatively shallow fold extending radially to the dorsal margin is present in Dauritheca, a situation that is reminiscent of Triplicatella sp. and Triplicatella uslonica. However, Dauritheca is delimited from these, and other species of Triplicatella, by its pronounced radial ornamentation. The ornamentation of Triplicatella species is composed of comarginal growth lines, sometimes with obscure radial ribs. Internally, the operculum of Dauritheca is closely similar to the interior of Triplicatella xinjia Pan, Skovsted, Sun & Li, Citation2019, which bears a robust transverse bar close to the dorsal margin (Pan et al. Citation2019, fig. 6L).
Dauritheca triangula (Qian, Yin & Xiao, 2000)
()
2000 Pachytheca triangula Qian, Yin & Xiao, pp. 409, 414–415, pl. 3, figs 1–4.
2010 Pachytheca triangula; Parkhaev & Demidenko, pp. 1132, 1143.
(non Pachytheca Schlüter, Citation1885).
Lectotype.
Here designated as specimen number 124974 (Nanjing Institute of Geology and Palaeontology), illustrated by Qian et al. (Citation2000, pl. 3, fig. 1). Paralectotypes: specimens numbered 124975–124977, illustrated by Qian et al. (Citation2000, pl. 3, figs 2–4).
Figured material.
PMU 39277–PMU 39280 from sample 7000/68. PMU 39311 and PMU 39312 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Description.
The operculum is sub-triangular in shape with a broad, shallowly convex ventral margin and a round-pointed dorsal apex. The most complete specimen has a width of about 1.3 mm and its length along the plane of symmetry is about 60% of the width (); maximum width is reached at about one quarter of the length, measured from the ventral margin. Smaller specimens are proportionately taller and more transversely oval (). The summit lies at between half and one third of the distance from the dorsal margin to the ventral margin and slightly overhangs the dorsal area (). The opercular margins are planar in larger specimens, with the exception of a shallow median fold that broadens from the summit to the dorsal margin and is bounded by shallow depressions (). The margins are convex in lateral perspective in smaller specimens, and the operculum is proportionately higher, with a steeper ad-dorsal field and a relatively more prominent median fold (). The internal surface is shallowly concave but traversed by a broad, robust platform dorsally. Ornamentation consists of numerous fine radial ribs crossed by comarginal growth lines; the ribs become coarser and more widely spaced near the dorsal apex. The conch has not been recognized.
Discussion.
The operculum of Triplicatella disdoma, the type species of Triplicatella, is distinguished by the narrow median fold extending from the summit to the dorsal margin and the three folds extending towards the ventral margin. Its ornamentation consists of comarginal growth lines and obscure radial ribs in contrast to the prominent radial ornamentation of Dauritheca. Triplicatella peltata differs from Dauritheca triangula in possessing a pair of shallow ventral folds and its more elliptical shape.
In addition to its prominent radial ornamentation, Dauritheca triangula is readily delimited from Triplicatella sinuosa by the four prominent folds in the dorsal margin of the latter (Skovsted et al. Citation2004). Only a single, shallow dorsal fold is present in Dauritheca triangula. Triplicatella xinjia has prominent folds in the dorsal and ventral margins (Pan et al. Citation2019, Peel Citation2021b), while Triplicatella papilo from northern Siberia differs from Dauritheca triangula is possessing two prominent folds in the dorsal margin.
One operculum of Dauritheca triangula shows a deep invagination in the ventral margin () that is interpreted as the result of a probable predatory attack (Ebbestad & Peel Citation1997, Alexander & Dietl Citation2003, Lindström & Peel Citation2005). A closely similar scar transgressing the hinge zone is known in an unfigured specimen of the bradoriid Liangshanella sayutinae (Melnikova, Citation1988).
Salanytheca Missarzhevsky, 1981
Type species.
Salanytheca papillaris Missarzhevsky, Citation1981, lower Cambrian Bayan Gol Formation of Mongolia.
Discussion.
The generic and specific names were inadvertently referred to Missarzhevsky (Citation1977) by Missarzhevsky (Citation1989, p. 159). Qian & Bengtson (Citation1989) placed Salanytheca as a junior synonym of Paragloborilus Qian, Citation1977, but Kouchinsky et al. (Citation2017, p. 389) noted that Missarzhevsky’s (Citation1981) species was distinguished from other species assigned to Paragloborilus by its prominent comarginal ribs and late early Cambrian age. Salanytheca is re-established here on the basis of material from Uslon and specimens from the Emyaksin Formation (Atdabanian Stage) of northern Siberia referred by Kouchinsky et al. (Citation2015, fig. 22F–I) to Allatheca? cf. corrugata Missarzhevsky in Rozanov et al., Citation1969.
Khetatheca Missarzhevsky, Citation1989, from the lower Cambrian of Siberia, is similar to Salanytheca in developing prominent comarginal ribs. However, Rozanov et al. (Citation1969) and Kouchinsky et al. (Citation2017) noted that the type species, Khetatheca cotuiensis (Syssoiev, Citation1959), is characterized by slight helical twisting that is not evident in Salanytheca.
Cupitheca holocyclata (Bengtson in Bengtson et al., Citation1990), originally described from the lower Cambrian Parara Limestone of South Australia, shows prominent comarginal ribs, although these are usually sharply delimited and more closely spaced than in Salanytheca. Cupitheca Duan, Citation1984 is characterized by truncation of the apical portion of the conch at a transverse septum (Bengtson et al Citation1990, Skovsted et al. Citation2016, Sun et al. Citation2018, Vendrasco et al. Citation2017), usually evident in the form of a smooth apical termination of the internal mould (). No evidence of septation or truncation has been observed in the Uslon material referred to Salanytheca ().
Figure 9. Hyolith conchs from the Bystraya Formation, Uslon section, Georgievka, eastern Transbaikalia. A–H, Salanytheca daurica sp. nov. A, PMU 39285, holotype, from sample У2. B, F, G, PMU 39286, sample 7000/68, with apex enlarged (F). G, detail of surface of F showing stacked shell lamellae. C, H, PMU 39287, from sample У2. D, PMU 39288 from sample У1. E, PMU 39289 from sample У2. I–K, Cupitheca holocyclata (Bengtson in Bengtson et al., Citation1990) from sample У2. K, PMU 39290, with reticulate ornamentation. I, J, PMU 39291, exfoliated specimen with terminal septa, and trace of comarginal ribs (arrow in J). Scale bars: 100 µm, except G (50 µm).

Salanytheca daurica sp. nov.
()
2015 Allatheca? cf. corrugata; Kouchinsky et al., p. 443, fig. 22F–I.
Holotype.
PMU 39285 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Additional figured material.
PMU 39286 from sample 7000/68; PMU 39288 from sample У1, PMU 39287 and PMU 39289 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Diagnosis.
Shallowly curved, slowly expanding (about 5°) conch with circular cross-section. Prominent comarginal ribs are separated by concave interareas, which vary from twice to about five times the width of the ribs. Apex and operculum not known.
Etymology.
From Dauria, the region of the east Asian steppes extending from Transbaikalia into eastern Mongolia and northern China.
Description.
Broken specimens from samples У1 and У2 expand at a greater rate () than the single specimen from sample 7000/68 () and are shallowly concave along the length of the dorsal surface (). Their comarginal ribs are slightly coarser than in the specimen from sample 7000/68 (), and may fade to some extent on the ventral surface (). Ornamentation consists of comarginal ribs and separating concave areas which are two to five times broader than the ribs themselves. The upper surface of the ribs varies from rounded to flattened (); fine comarginal growth lines may occur in the concave areas between the ribs. The aperture is sub-circular in cross-section; internal septation and the nature of the apex have not been observed. Conch surface, as preserved, shows imbrication of shell foliate lamellae (). The operculum is not known.
Discussion.
In terms of shape and the comarginal ribbing, the specimen from sample 7000/68 () is similar to some specimens of Hyolithellus Billings, Citation1871, Pseudorthotheca Cobbold, Citation1935 and Rugatotheca He in Yin et al., Citation1980, such as those illustrated by Qian (Citation1989, pls 1, 10). However, the preserved impression of an imbricated foliate shell structure () indicates that the specimen is an internal mould of a calcareous tube. Structures of this kind are not recorded in the organophosphatic tubes of Hyolithellus (Skovsted & Peel Citation2011, Demidenko Citation2021), but imprints of inner shell layers of calcareous shells in association with phosphatized internal moulds in Cambrian molluscs and hyoliths have been described by Runnegar (Citation1985), Kouchinsky (Citation2000a, Citation2000b, Citation2007), Vendrasco et al. (Citation2010, Citation2011, Citation2017), Kouchinsky et al. (Citation2015), Moore & Porter (Citation2018) and Li et al. (Citation2019, Citation2020). Li et al. (Citation2019) clearly demonstrated aragonite foliation in Cambrian hyoliths from North China, with the growing edge of the foliae perpendicular to the shell margin. In contrast, the growing edge of the foliation is oblique in the figured Uslon specimen, is inclined at a low angle to the comarginal angulations ().
Kouchinsky et al. (Citation2015, fig. 22F–I) assigned to Allatheca? cf. corrugata Missarzhevsky in Rozanov et al., Citation1969 specimens from the Emyaksin Formation (Atdabanian Stage) of northern Siberia that have a flattened ventral surface, whereas the Uslon material appears to be generally circular in cross-section. However, the Emyaksin specimens are also referred to Salanytheca duarica. The Uslon specimens and material described by Kouchinsky et al. (Citation2015) have more regular and clearly defined comarginal ribs than the type material of Allatheca corrugata illustrated by Rozanov et al. (Citation1969, pl. 1, fig. 6; pl. 12, figs 10–11), where the transverse ornamentation is less regular and periodically lamellose.
Salanytheca papillaris differs from Salanytheca duarica in terms of its coarser, more closely spaced, ribbing. The holotype retains a globose protoconch (Missarzhevsky Citation1989, pl. 4, fig. 1) but the protoconch is not known in Salanytheca duarica.
In addition to its reported slight helical twisting, Khetatheca cotuiensis, as illustrated by Kouchinsky et al. (Citation2017, fig. 50A–C, F–H) from the early Cambrian Medvezhya Formation of Siberia, displays a range in shape comparable to that of the Uslon material, but most specimens have broader comarginal ribs that are more widely spaced. Paragloborilus songbaiensis Duan, Citation1984 from the lower Cambrian Xihaoping Formation of Hubei Province, China, shows similar well-defined ornamentation but has a strongly curved shell in lateral profile (Qian Citation1989, pl. 7, fig. 5–8).
Family CUPITHECIDAE Duan, Citation1984
Discussion.
Pan et al. (Citation2019) emended the diagnosis of Cupithecidae Duan, Citation1984 to take account of the apical decollation during ontogeny and the configuration of the operculum, including only the eponymous genus.
Cupitheca Duan, 1984
Type species.
Paragloborilus mirus He in Qian, Citation1977, lower Cambrian (Fortunian Stage), South China.
Cupitheca holocyclata (Bengtson in Bengtson et al., Citation1990)
()
1990 Actinotheca holocyclata Bengtson in Bengtson et al., p. 204, figs 134–136.
1990 Actinotheca clathrata Bengtson in Bengtson et al., p. 204, figs 141, 142.
2016 Cupitheca holocyclata; Skovsted et al., pp. 123–130, figs 2–4.
2019 Cupitheca holocyclata; Pan et al., p. 356, fig. 2f–k.
(includes additional references).
Figured material.
PMU 39290 and PMU 39291 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
This exfoliated, straight conch preserves the aperture and terminates apically in a convex septum, therefore apparently representing the body chamber. Traces of the closely spaced comarginal ribs (, arrow) indicate similar ornamentation to that seen in material illustrated by Bengtson et al. (Citation1990, fig. 134A, B) from the Parara Limestone of South Australia. This outer shell layer preserves a comarginal fibrous structure (), but an inner layer surrounding the aperture has longitudinal fibres. Detailed discussions of shell structure in Cupitheca were given by Vendrasco et al. (Citation2017) and Sun et al. (Citation2020).
A fragment of a larger specimen displays fine longitudinal striations crossing comarginal ribs (), producing the reticulate ornamentation upon which Bengtson et al. (Citation1990) established Cupitheca clathrata (Bengtson in Bengtson et al., Citation1990). The species was placed into synonymy with Cupitheca holocyclata by Skovsted et al. (Citation2016).
Family UNCERTAIN
Majatheca Missarzhevsky in Rozanov et al., 1969
Type species.
Majatheca tumefacta Missarzhevsky in Rozanov et al., Citation1969, Cambrian, upper Tommotian Stage, middle Lena River region, Siberia.
Majatheca sp.
()
Figured material.
PMU 39294 from sample У1. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
This single operculum is more transversely oval in shape than the specimen of Majatheca tumefacta Missarzhevsky in Rozanov et al., Citation1969 illustrated by Kouchinsky et al. (Citation2015, fig. 24) from the uppermost Emyaksin Formation (Botoman Stage) of the Anabar uplift in northern Siberia. Peel (Citation2021b) illustrated as allathecid sp. 1 a specimen from the basal Aftenstjernesø Formation (Cambrian Stage 4) of northern Lauge Koch Land, North Greenland, which is sub-circular in shape. It differs from Majatheca sp. in terms of its robust clavicular ring that is more strongly developed and of uneven height.
A similar operculum is present in Tegminites Duan & Xiao, Citation1992 but it is distinguished from the current specimen by developing small but distinct folds along the dorsal margin of the clavicular ring (Pan et al. Citation2019, fig. 5H, H1).
Operculum 1
()
Figured material.
PMU 39283 from sample 7000/68. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
This operculum is similar to Operculum type 3 of Kouchinsky et al. (Citation2015) from the uppermost Emyaksin Formation (Cambrian Series 2, lower Botoman Stage) of northern Siberia. It differs in being elliptical in form, rather than equidimensional; the ventrolateral margins are broken. The cardinal process is broken and encrusted with diagenetic phosphate, but comprises a short tooth located on each side of a narrow median groove. The clavicles, at mid-height (), are composed of five or six radial rods that slope in towards the median line. The median area of the inner surface preserves three sub-circular depressions, with a broad, shallow Y-shaped ridge connecting the dorsal extremities of the clavicles to the median ventral margin. In addition to the figured example, a crushed specimen is known from sample У1.
Operculum 2
()
Figured material.
PMU 39295 from sample У1. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
The inner surface of the only available specimen is mainly smooth but the dorsal quarter is traversed by a bar-like platform with a shallow median groove (). The cardinal processes are broken but appear to be short and divergent, and continue laterally into a ridge (, left side of apex). Distal to this ridge, the surface slopes outwards to the opercular margin.
Operculum 3
()
Figured material.
PMU 39310 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
The single specimen is transversely oval in shape with a shallowly concave upper surface and an upturned, presumed dorsal, margin. The protoconch is swollen, dorsoventrally elongate and lies closer to the dorsal margin. Apart from the smooth protoconch, ornamentation consists of comarginal growth of variable relief. The inner surface is not known.
Operculum 2 differs from Operculum 3 in the much greater concavity of its upper surface.
Operculum 4
()
Figured material.
PMU 39313 from sample У2. Bystraya Formation, eastern Transbaikalia (Zabaykalsky Krai).
Discussion.
The outer surface is strongly concave, with the ventral and dorsal fields inclined to each other but not delimited by folds or grooves. The protoconch is swollen and smooth () and similar to that seen in some specimens of Triplicatella uslonica (). Ornamentation consists of comarginal growth lines that are more strongly defined on the dorsal field ().
Acknowledgements
Alexander Gubanov’s field work in Transbaikalia during 2000 was supported by the Swedish Research Council through grants to J.S. Peel. We thank S.M. Sinitsa (Chita) and her colleagues for assistance during 2000. Residues of samples У1 and У2 were kindly donated by Elena A. Zhegallo (Moscow). Comments from two anonymous reviewers and the editorial staff are gratefully acknowledged.
Disclosure statement
No potential conflict was reported by the authors.
Data archiving statement
This published work and the nomenclatural acts it contains have been registered in ZooBank: zoobank.org:pub:C39B5277-F926-4267-95AE-92C1212D6B58
References
- Alexander, R.R. & Dietl, G.P., 2003. The fossil record of shell-breaking predation on marine bivalves and gastropods. In Predator–Prey Interactions in the Fossil Record. Kelley, P.H., Kowalewski, M. & Hansen, T.A., eds. Kluwer Academic/Plenum Publishers, New York, 141–176, 464 pp.
- Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A. & Runnegar, B.N., 1990. Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9, 1–364.
- Bengtson, S., Matthew, S.C. & Missarzhevsky, V.V., 1986. The Cambrian net-like fossil Microdictyon. In Problematic Fossil Taxa, Hoffman, A. & Nitecki, M.H., eds, Oxford Monographs on Geology and Geophysics 5. Oxford University Press, New York, 97–115.
- Berg-Madsen, V. & Malinsky, J.M., 1999. A revision of Holm’s Late Mid and Middle Cambrian hyoliths of Sweden. Palaeontology 42, 841–885.
- Billings, E., 1871. On some new species of Palaeozoic fossils. Canadian Naturalist 6, 213–233, 240.
- Bokova, A.R. & Vasil’eva, N.I., 1990. Some new species of problematic skeletal fossils from the Lower Cambrian Olenek-uplift. In Fossil Problems of the USSR. Sokolov, B.S., ed., Trudy Instituta Geologii i Geofiziki, Sibirskoe Otdelenie, Akademiya Nauk SSSR 783, Russian, 28–30. [in Russian].
- Canu, F., 1913. Études morphologiques sur trois nouvelles familles de Bryozoaires. Bulletin de la Société Géologique de France, Séries 4, 13, 132–147.
- Cobbold, E.S., 1920. The Cambrian horizons of Comley and their Brachiopoda, Pteropoda, Gastropoda, etc. Quarterly Journal of the Geological Society of London 76, 325–386.
- Cobbold, E.S., 1935. Lower Cambrian Faunas from Hérault, France. Annals and Magazine of Natural History, Series 10, 16, 25–48.
- Debrenne, F., Zhuravlev, A. & Rozanov, A.Y., 1989. Pravil’nye arkheotsiaty. Trudy Paleontologicheskiy Institut, Akademiya Nauk SSSR 233, 199.
- Demidenko, Y.U., 2021. Microstructure of tubes of the Cambrian zooproblematics of the Family Hyolithellidae. Paleontologicheskiy Zhurnal 2021, 10–22. [in Russian].
- Devaere, L., Clausen, S., Alvaro, J.J., Peel, J.S. & Vachard, D., 2014. Terreneuvian orthothecid (Hyolitha) digestive tracts from northern Montagne Noire, France; taphonomic, ontogenetic and phylogenetic implications. PLoS One 9, e88583.
- Doré, F. & Reid, R.E., 1965. Allonnia tripodophora nov. gen., nov. sp., nouvelle Éponge du Cambrien inférieur de Carteret (Manche). Compte Rendu Sommaire des Séances de la Société Géologique de France 1965, 20–21.
- Duan, C.H., 1984. Small shelly fossils from the lower Cambrian Xihaoping Formation in the Shennongjia district, Hubei Province—Hyoliths and fossil skeletons of unknown affinities. Bulletin of the Tianjin Institute of Geology and Mineral Resources 7, 143–183. [Chinese with English abstract].
- Duan, C.H. & Xiao, B., 1992. Restudy on the stratotype of Yurtus Formation. Bulletin Tianjin Institute of Geology and Mineral Resources 26, 325–355. [Chinese with English abstract].
- Ebbestad, J.O.R. & Peel, J.S., 1997. Attempted predation and shell repair in Middle and Upper Ordovician gastropods from Sweden. Journal of Paleontology 71, 1007–1019.
- Esakova, N.V. & Zhegallo, E.A., 1996. Biostratigrafiya i fauna nizhnego kembriya Mongolii. Trudy Sovmestnaya Rossiysko-Mongol’skaya Paleontologicheskaya Ekspeditsiya 46, 216 pp.
- Gravestock, D.I., Alexander, M.I., Demidenko, E.Y., Esakova, N.V., Holmer, L.E., Jago, J.B., Melnikova, L.M., Parkhaev, P.Y., Rozanov, A.Y., Ushatinskaya, G.T., Zang, W., Zhegallo, E.A. & Zhuravlev, A.Y., 2001. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Russian Academy of Sciences Transactions of the Palaeontological Institute 282, 344 pp.
- He, T.G., Pei, F. & Fu, G.H., 1984. Small shelly fossils from the lower Cambrian Xinji Formation in Fangcheng County, Henan Province. Acta Palaeontologica Sinica 23, 350–357. [Chinese with English abstract]
- Holm, H., 1893. Sveriges Kambrisk-Siluriska Hyolithidae och Conularidae. Sveriges Geologiska Undersökning Serie C 112, 1–173.
- Jell, P.A., 1980. Earliest known bivalve on Earth—A new early Cambrian genus from South Australia. Alcheringa 4, 233–239.
- Knyazev, G.I., 1962. Stratigrafiya nizhnego paleozoya i dokembriya Priargun’ya. Chitinskoe Otdelenie Tsentral’nogo Nauchno-Issledovatel’skogo Gornorazvedochnogo Instituta, Chita, 124 pp.
- Kouchinsky, A., 2000a. Shell microstructures in Early Cambrian molluscs. Acta Palaeontologica Polonica 45, 119–150.
- Kouchinsky, A., 2000b. Skeletal microstructures of hyoliths from the Early Cambrian of Siberia. Alcheringa 24, 65–81.
- Kouchinsky, A., 2007. Shell microstructures of the Early Cambrian Anabarella and Watsonella as new evidence on the origin of the Rostroconchia. Lethaia 32, 173–180.
- Kouchinsky, A., Alexander, R., Bengtson, S., Bowyer, F., Clausen, S., Holmer, L.E., Kolesnikov, K.A., Korovnikov, I.V., Pavlov, V., Skovsted, C.B., Ushatinskaya, G., Wood, R. & Zhuravlev, A.Y., 2022. Early–middle Cambrian stratigraphy and faunas from northern Siberia. Acta Palaeontologica Polonica 67, 341–464.
- Kouchinsky, A., Bengtson, S., Clausen, S. & Vendrasco, M., 2015. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontographica Polonica 60, 421–512.
- Kouchinsky, A., Bengtson, S., Landing, E., Steiner, M., Vendrasco, M. & Ziegler, K., 2017. Terreneuvian stratigraphy and faunas from the Anabar Uplift. Acta Palaeontologica Polonica 62, 331–340.
- Landing, E. & Bartowski, K.E., 1996. Oldest shelly fossils from the Taconic allochthon and late Early Cambrian sea − levels in eastern Laurentia. Journal of Paleontology 70, 741–761.
- Li, L., Skovsted, C.B., Yun, H., Betts, M.J. & Zhang, X., 2020. New insight into the soft anatomy and shell microstructures of early Cambrian orthothecids (Hyolitha). Proceedings of the Royal Society, Biological Sciences 287, 20201467.
- Li, L., Zhang, X., Skovsted, C.B., Yun, H., Pan, B. & Li, G., 2019. Homologous shell microstructures in Cambrian hyoliths and molluscs. Palaeontology 62, 515–532.
- Lindström, A. & Peel, J.S., 2005. Repaired injuries and shell form in some Palaeozoic pleurotomarioid gastropods. Acta Palaeontologica Polonica 50, 697–704.
- Liu, F., Skovsted, C.B., Topper, T.P. & Zhang, Z., 2021. Revision of Triplicatella (Orthothecida, Hyolitha) with preserved digestive tracts from the early Cambrian Chengjiang Lagerstätte, South China. Historical Biology 33, 1857–1871.
- Maas, A., Braun, B., Dong, X., Donoghue, P.C.J., Müller, K.J., Olempska, E., Repetski, J.E., Siveter, D.J., Stein, M. & Waloszek, D., 2006. The ‘Orsten’—more than a Cambrian Konservat-Lagerstätte yielding exceptional preservation. Palaeoworld 15, 266–282.
- Malinky, J.M., 1988. Early Paleozoic Hyolitha from North America: reexamination of Walcott’s and Resser’s type specimens. Journal of Paleontology 62, 218–233.
- Malinky, J.M., 1989. New Early Paleozoic Hyolithida and Orthothecida (Hyolitha) from North America. Journal of Paleontology 63, 302–319.
- Malinky, J.M., 2014. Cambrian Hyolitha and problematica from West Laurentian North America: taxonomy and palaeobiogeography. Alcheringa 38, 338–363.
- Malinky, J.M. & Berg-Madsen, V., 1999. A revision of Holm’s Early and early Mid Cambrian hyoliths of Sweden. Palaeontology 42, 25–65.
- Malinky, J.M. & Geyer, G., 2019. Cambrian Hyolitha of Siberian, Baltican and Avalonian aspect in east Laurentian North America: taxonomy and palaeobiogeography. Alcheringa 43, 171–203.
- Malinky, J.M. & Skovsted, C.B., 2004. Hyoliths and small shelly fossils from the Lower Cambrian of North-East Greenland. Acta Paleontologica Polonica 49, 551–578.
- Mambetov, A.M., 1972. A new lower Cambrian hyolithid genus from Malyy Karatau (northwestern Tien Shan). Paleontological Journal 6, 268–269.
- Marek, L., 1963. New knowledge on the morphology of Hyolithes. Sborník Geologických Ve˘d, řada Paleontologie 1, 53–72.
- Marek, L., 1966. New hyolithid genera from the Ordovician of Bohemia. Časopis Národního Muzea. Oddil Přirodove˘dny 135, 89–92.
- Martí Mus, M. & Bergström, J., 2007. Skeletal microstructure of helens, lateral appendages of hyolithids. Palaeontology 50, 1231–1243.
- Martí Mus, M., Jeppsson, L. & Malinky, J.M., 2014. A complete reconstruction of the hyolithid skeleton. Journal of Paleontology 88, 160–170.
- Melnikova, L.M., 1988. Some bradoriids (Crustacea) from the Botomian Stage of the East Trans-Baikal Region. Paleontological Journal 1998, 113–116. [=Paleontologicheskiy Zhurnal 1988, 114–117]. Note incorrect title of eastern Kazakhstan on English reprint
- Melnikova, L.M., Siveter, D.J. & Williams, M., 1997. Cambrian Bradoriida and Phosphatocopida (Arthropoda) of the former Soviet Union. Journal of Micropalaeontology 16, 179–191.
- Meshkova, N.P., 1974. Lower Cambrian hyoliths of the Siberian Platform. Trudy Instituta Geologii i Geofiziki, Sibirskoe Otdelenie Akademii Nauk SSSR 97, 110 pp. [in Russian].
- Missarzhevsky, V.V., 1977. Conodonts (?) and phosphatic problematica from the Cambrian of Mongolia and Siberia. In Bespozvonochnye paleozoya Mongolii. Tatarinov, L.P., ed.,. Nauka, Moskva, 10–19. [in Russian].
- Missarzhevsky, V.V., 1981. Early Cambrian hyoliths and gastropods from Mongolia [in Russian]. Paleontologicheskiy zhurna1 1981, 21–28.
- Missarzhevsky, V.V., 1989. Oldest skeletal fossils and stratigraphy of Precambrian and Cambrian boundary beds. Trudy Geologicheskogo Instituta, Akademiya Nauk SSSR 443, 237 pp. [in Russian].
- Missarzhevsky, V.V. & Mambetov, A.M., 1981. Stratigraphy and fauna of the Cambrian and Precambrian boundary beds of the Maly Karatau Range. Trudy Geologicheskogo Instituta Akademiya Nauk SSSR 326, 92 pp. [in Russian].
- Moore, J.L., Li, G. & Porter, S.M., 2014. Chancelloriid sclerites from the lower Cambrian (Meishucunian) of eastern Yunnan, China, and the early history of the group. Palaeontology 57, 833–878.
- Moore, J.L. & Porter, S.M., 2018. Plywood-like shell microstructures in hyoliths from the Middle Cambrian (Drumian) Gowers Formation, Georgina Basin, Australia. Palaeontology 61, 441–467.
- Moore, J.L., Porter, S.M., Steiner, M. & Li, G., 2010. Cambrothyra ampulliformis, an unusual coeloscleritophoran from the lower Cambrian of Shaanxi Province. Journal of Paleontology 84, 1040–1060.
- Moysiuk, J., Smith, M.R. & Caron, J.-B., 2017. Hyoliths are Palaeozoic lophophorates. Nature 541, 394–397.
- Novozhilova, N.V. & Korovnikov, I.V., 2019. Small shelly fossils in the Cambrian basement of the West Siberian Geosyneclise. Stratigrafiya, Geologicheskaya korrelyatsiya 27, 3–11.
- Pakhomov, N.N., 1995. Eastern Transbaikalia, 73–75. In The Cambrian System of the foldbelts of Russia and Mongolia Ashtaskin, V.A., Pegel, T.V., Repina, L.N., Belyaeva, G.V., Esakova, N.V., Rozanov, A.Yu., Zhuravlev, A.Yu., Osadchaya, D.V. & Pakhomov, N.N. International Union of Geological Sciences 32, Trondheim, Norway, 132 pp.
- Pan, B., Skovsted, C.B., Sun, H.J. & Li, G.X., 2019. Biostratigraphical and palaeogeographical implications of early Cambrian hyoliths from the North China Platform. Alcheringa 43, 351–380.
- Parkhaev, P.Y., 2004. Malacofauna of the Lower Cambrian Bystraya Formation of Eastern Transbaikalia. Paleontological Journal 38, 9–25.
- Parkhaev, P.Y., 2006. On the genus Auricullina Vassiljeva, 1998 and shell pores of the Cambrian helcionelloid mollusks. Paleontological Journal 40, 20–33.
- Parkhaev, P.Y., 2007. Shell chirality in Cambrian gastropods and sinistral members of the Genus Aldanella Vostokova, 1962. Paleontological Journal 41, 233–240.
- Parkhaev, P.Y. & Demidenko, Y., 2010. Zooproblematica and Mollusca from the lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontological Journal 44, 883–1161.
- Peel, J.S., 2021a. Holdfasts of Sphenothallus (Cnidaria) from the early Silurian of western North Greenland (Laurentia). GFF 143, 384–389.
- Peel, J.S., 2021b. An outer shelf shelly fauna from Cambrian Series 2 (Stage 4) of North Greenland (Laurentia). Journal of Paleontology 95, 1–41.
- Peel, J.S., Streng, M., Geyer, G., Kouchinsky, A. & Skovsted, C.B., 2016. Ovatoryctocara granulata assemblage (Cambrian Series 2–Series 3 boundary) of Løndal, North Greenland. Australasian Palaeontological Memoirs 49, 241–282.
- Peel, J.S. & Willman, S., 2018. The Buen Formation (Cambrian Series 2) biota of North Greenland. Papers in Palaeontology 4, 381–432.
- Qian, Y., 1977. Hyolitha and some problematica from the Lower Cambrian Meishucun Stage in central and south western China. Acta Palaeontologica Sinica 16, 255–278. (Chinese with English abstract)
- Qian, Y., 1978. The Early Cambrian hyolithids in Central and Southwest China and their stratigraphicalal significance. Memoirs of the Nanjing Institute of Geology and Palaeontology. Academia Sinica 11, 1–38. (in Chinese with English summary)
- Qian, Y., 1989. Early Cambrian small Shelly Fossils of China with Special Reference to the Precambrian–Cambrian boundary. Stratigraphy and Palaeontology of Systemic Boundaries in China Precambrian—Cambrian Boundary (2). Nanjing University Publishing House, Nanjing, 341 pp.
- Qian, Y. & Bengtson, S., 1989. Palaeontology and biostratigraphy of the Early Cambrian Meishucunian Stage in Yunnan Province, South China. Fossils and Strata 24, 156.
- Qian, J.X. & Xiao, B., 1984. An Early Cambrian small shelly fauna from Aksu–Wushi Region, Xinjiang. Professional Papers on Stratigraphy and Palaeontology 13, 65–90. Chinese with English abstract).
- Qian, Y., Yin, G.Z. & Xiao, B., 2000. Opercula of hyoliths and operculum-like fossils from the Lower Cambrian Yuertus Formation, Xinjiang. Acta Micropalaeontologica Sinica 17, 404–415.
- Qian, Y. & Zhang, S.B., 1983. Small shelly fossils from the Xihaoping Member of the Tongying Formation in Fangxian County of Hubei Province and their stratigraphical significance. Acta Palaeontologica Sinica 22, 82–94. [Chinese with English abstract]
- Rozanov, A.Y. & Missarzhevsky, V.V., 1966. Biostratigrafiya i fauna nizhnikh gorizontov kembriya. Trudy Geologicheskogo Instituta Akademiya Nauk SSSR 148, 125.
- Rozanov, A.Yu., Missarzhevsky, V.V., Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller, B.M., Korolyuk, I.K., Lendzion, K., Michniak, R., Pykhova, N.G. & Sidorov, A.D., 1969. The Tommotian Stage and the Cambrian lower boundary problem. Trudy Geologicheskogo Instituta Akademiya Nauk SSSR 296, 380 pp. [in Russian. English translation published as Raaben, M.E., ed., 1981. Amerind Publishing Company, New Delhi, 359 pp.]
- Runnegar, B., 1985. Shell microstructures of Cambrian molluscs replicated by phosphate. Alcheringa 9, 245–257.
- Saito, K., 1936. Older Cambrian Brachiopoda, Gastropoda, etc., from Northwestern Korea. Journal of the Faculty of Science, the Imperial University of Tokyo, Section 2, 4, 345–367.
- SchlÜter, C., 1885. Über einige neue Anthozoen aus dem Devon. Naturhistorischen Vereins Preussischen Rheinlande und Westfalens. Verhandlungen, Sitzungsberichte der Niederrheinische Gesellschaft für Natur- und Heilkunde, Jahrgang 42, 144–151.
- Schmidt, G., 2012. Bonnetina (Pachytheca) tanzeri subgen. et sp. n., eine bisher unbeschriebene Vogelspinnenart aus Mexico (Araneae: Theraphosidae: Theraphosinae). Arthropoda Scientia 2-2012, 21–28.
- Sdzuy, K., 1969. Unter- und mittelkambrische Porifera (Chancellorida und Hexactinellida.): Paläontologische Zeitschrift 43, 115–147.
- Skovsted, C.B., 2006. Small shelly fossils from the upper Lower Cambrian Bastion and Ella Island Formations, North-East Greenland. Journal of Paleontology 80, 1087–1112.2.0.CO;2]
- Skovsted, C.B., Pan, B., Topper, T.P., Betts, M.J., Li, G. & Brock, G.A., 2016. The operculum and mode of life of the lower Cambrian hyolith Cupitheca from South Australia and North China. Palaeogeography, Palaeoclimatology, Palaeoecology 443, 123–130.
- Skovsted, C.B. & Peel, J.S., 2007. Small shelly fossils from the argillaceous facies of the Lower Cambrian Forteau Formation of western Newfoundland. Acta Palaeontologica Polonica 52, 729–748.
- Skovsted, C.B. & Peel, J.S., 2011. Hyolithellus in life position from the lower Cambrian of North Greenland. Journal of Paleontology 85, 37–47.
- Skovsted, C.B., Peel, J.S. & Atkins, C.J., 2004. The problematic fossil Triplicatella from the Early Cambrian of Greenland, Canada and Siberia. Canadian Journal of Earth Sciences 41, 1273–1283.
- Skovsted, C.B., Topper, T.P., Betts, M.J. & Brock, G.A., 2014. Associated conchs and opercula of Triplicatella disdoma (Hyolitha) from the early Cambrian of South Australia. Alcheringa 38, 148–153.
- Sun, H., Yin, Z., Li, G., Zhao, F., Zeng, H. & Zhu, M., 2020. Periodic shell decollation as an ecology-driven strategy in the early Cambrian Cupitheca. Palaeontology 63, 431–442.
- Sun, H.J., Malinky, J.M., Zhu, M.Y. & Huang, D.Y., 2018. Palaeobiology of orthothecid hyoliths from the Cambrian Manto Formation of Hebei Province, North China. Acta Palaeontologica Polonica 63, 87–101.
- Syssoiev, V.A., [Sysoev, V.A.], 1957. To the morphology, systematics and systematic position of the hyoliths. Doklady Akademiya Nauk SSSR 116, 304–307. [in Russian].
- Syssoiev, V.A., [Sysoev, V.A.], 1958. Superorder Hyolithoidea [in Russian]. In Orlov, U.A., Luppov, N.P. & Druŝic, V.V. (eds.), Osnovy paleontologii. Molluski − Golovonogie. 2, 184–190. Akademiya Nauk SSSR, Moskva. [in Russian].
- Syssoiev, V.A. [Sysoev, V.A.], 1959. Hyoliths of the genus Circotheca from the Lower Cambrian of the Tajmyr National District. Paleontologicheskiy Zhurnal 1959, 84–94. [in Russian].
- Ushatinskaya, G.T., 1988. Obolellids (brachiopods) with hinged articulation of valves from the Lower Cambrian of Transbaikalia. Paleontological Journal 1988 (1), 32–38 [=Paleontologicheskiy Zhurnal 1988 (1), 34–39].
- Val’kov, A.K., 1975. Biostratigrafiya i khiolity kembriya severo-vostoka Sibirskoy platformy. Instituta Geolgia, Moscow, Yakutskiy filial’ Sibirskoe otdelenia, Nauk, Moscow. Akademiya Nauk SSSR, 136 pp.
- Vasil’eva, N.I., 1998. Melkaya rakovinnaya fauna i biostratigrafiya nizhnego kembriya Sibirskoj platformy. VNIGRI, St. Petersburg, 139. pp.
- Vasil’eva [Vassiljeva], N.I. & Sayutina, T.A., 1988. Morfologicheskoe raznoobrazie skleritov khantselloriy. 190–198. In Kembriy Sibiri i Sredney Azii. Zhuravleva, I.T. & Repina, L.N., eds., Trudy Instituta Geologii i Geofiziki, Sibirskoe Otdelenie, Akademiya Nauk SSSR, Russian,720, 215. pp.
- Vendrasco, M.J., Checa, A.G. & Porter, S.M., 2017. Periostracum and fibrous shell microstructure in the unusual Cambrian hyolith Cupitheca. Spanish Journal of Palaeontology 32, 95–108.
- Vendrasco, M.J., Kouchinsky, A.V., Porter, S.M. & Fernandez, C.Z., 2011. Phylogeny and escalation in Mellopegma and other Cambrian molluscs. Palaeontologica Electronica 14, 2, 11A. palaeo-electronica.org/2011_2/221/index.html
- Vendrasco, M.J., Porter, S.M., Kouchinsky, A., Li, G. & Fernandez, C.Z., 2010. New data on molluscs and their shell microstructures from the middle Cambrian Gowers Formation, Australia. Palaeontology 53, 97–135.
- Vologdin, A.G. & Fonin, V.D., 1966. Novye odnostennye tenial’nye arkheotsiaty Priargun’ya. Doklady Akademiya Nauk SSSR 167, 187–190.
- Walcott, C.D., 1920. Cambrian geology and paleontology IV: 6—Middle Cambrian Spongiae. Smithsonian Miscellaneous Collections 67, 261–364.
- Waloszek, D., 2003. The ‘Orsten’ Window—A three-dimensionally preserved Upper Cambrian meiofauna and its contribution to our understanding of the evolution of Arthropoda. Paleontological Research 7, 71–88.
- Wrona, R., 2003. Early Cambrian molluscs from glacial erratics of King George Island, West Antarctica. Polish Polar Research 24, 181–216.
- Yang, B., Steiner, M. & Keupp, H., 2015. Early Cambrian palaeobiogeography of the Zhenba–Fangxian Block (South China): independent terrane or part of the Yangtze Platform? Gondwana Research 28, 1543–1565.
- Yin, J., Ding, L., He, T., Li, S. & Shen, L., 1980. The palaeontology and sedimentary environment of the Sinian system in Emei − Ganuluo area, Sichuan. Geological College of Chengdu, Chengdu. 231. pp. [in Chinese].