
Abstract
A review of the Southern Hemisphere Mesozoic tribosphenic mammal fossil record supports the hypothesis that Tribosphenida arose in the Southern Hemisphere during the Early Jurassic, around 50 million years prior to the clade’s reliably dated first appearance in the Northern Hemisphere. Mesozoic Southern Hemisphere tribosphenic mammals are known from Australia, Madagascar, South America and the Indian subcontinent, and are classified into three families: Bishopidae (fam. nov.), Ausktribosphenidae and Henosferidae. These are stem therians, and considerable morphological evolution occurred within the lineage between the Jurassic and late Early Cretaceous. Important dental modifications include a graduated transition between premolars and molars, development of molar wear facets V and VI, loss of facets for postdentary bones, reduction in the Meckelian groove and development of a true dentary angle. Previous classifications of Southern Hemisphere tribosphenic mammals are ambiguous because information from the upper dentition has been lacking. Upper molars attributed to the late Early Cretaceous (Albian) Southern Hemisphere group Bishopidae fam. nov. are now known to possess a prominent protocone and stylar cusp C. We thus consider bishopids to be the sister group to Theria.
Timothy F. Flannery [[email protected]], Kristofer M. Helgen [[email protected]], Australian Museum, 1 William St Sydney 2000, Australia; Thomas H. Rich [[email protected]], Museums Victoria, PO Box 666, Q28 Melbourne, Victoria 3001, Australia; Patricia Vickers-Rich [[email protected]; [email protected]], School of Earth, Atmosphere and Environment, Monash University, Victoria 3800, Australia; Swinburne University of Science and Technology, Department of Chemistry and Biotechnology, Hawthorn, Victoria 3122, Australia; Elizabeth Grace Veatch [[email protected]], National Museum of Natural History, Smithsonian Institution, Washington, DC 20013, USA.
THEORIES about the origins of therians (= Metatheria and Eutheria) can be traced back to Charles Darwin, who wrote to Lyell in 1860 outlining two competing hypotheses: (1) either that marsupials were ancestral to both later marsupials and placentals; or (2) that both groups were derived from a non-marsupial, non-placental ancestor (Archibald Citation2012). Cope (Citation1883) identified the central importance of the ‘trituberculate’ molar to therian evolution, and Simpson (Citation1936) coined the term ‘tribosphenic’ for molars of the type described by Cope (Citation1883). Simpson (Citation1937) subsequently expanded on Cope’s (Citation1883) view that this ‘primitive molar type’ is critical for clarifying the origins of therians. Patterson (Citation1956) recognized that the protocone (the key defining feature of the Tribosphenida) is, despite its name, a neomorphic structure, as was first noted by Butler (Citation1939). Crompton (Citation1971) described the form and function of the tribosphenic molar in detail and reconstructed the molar morphology in the ancestral therian. Tribosphenic molars are functionally complex, involving precise interactions of six matching shearing surfaces (three of which are not present in non-tribosphenic mammals) along with crushing and puncturing components present on the upper and lower teeth (see Crompton Citation1971, Davis Citation2011).
These seminal studies were based exclusively on the Northern Hemisphere mammal fossil record. While a relatively rich record of Northern Hemisphere Mesozoic tribosphenic mammals has been accumulating since the nineteenth century (e.g., Cope Citation1883, Simpson Citation1937), it was not until 1997 that a Mesozoic mammal with tribosphenic molars was reported from the Southern Hemisphere (Rich et al. Citation1997). The discovery of Ausktribosphenos nyktos Rich, Vickers-Rich, Constantine, Flannery, Kool & van Klaveren, Citation1997, from the uppermost Barremian ‘Wonthaggi Formation’ outcrops at the Flat Rocks site on the Bass Coast of southern Victoria in Australia (Poropat et al. Citation2018, Wagstaff et al. Citation2020), was the first of several finds which, over the past quarter of a century have built up a small and still highly incomplete fossil record of Mesozoic mammals with tribosphenic molars from the Southern Hemisphere.
Six genera are now named, and at least two remain unnamed: Ausktribosphenos Rich, Vickers-Rich, Constantine, Flannery, Kool and van Klaveren, Citation1997, Bishops Rich, Flannery, Trusler, Kool, van Klaveren & Vickers-Rich, Citation2001 and Kryoparvus Rich, Trusler, Kool, Pickering, Evans, Siu, Maksomenko, Kundrat, Gostling, Morton & Vickers-Rich, Citation2020 from Australia; Ambondro Flynn, Parrish, Rakotosamimanana, Simpson & Wyss, Citation1999 from Madagascar; and Asfaltomylos Rauhut, Martin, Ortiz-Jaureguizar & Puerta, Citation2002, Henosferus Rougier, Martinelli, Forasiepi & Novacek, Citation2007, unnamed genus (‘aff. Bishops’: Martin et al. Citation2022) from South America; and an unnamed taxon from the Indian subcontinent (Prasad et al. Citation2015). In addition, several pre-tribosphenic, peramuran-like fossil mammals from the Jurassic and Early Cretaceous of Africa display some tribosphenic characteristics (Davis Citation2012). These include three species of Hypomylos Sigogneau-Russell, Citation1992 from the Jurassic and Early Cretaceous of Morocco, and Brancatherulum tendagurense Dietrich, Citation1927 and Tendagurutherium dietrichi Heinrich, Citation1998 from the Jurassic of Tanzania.
The phylogenetic relationships of Mesozoic Southern Hemisphere tribosphenic taxa are highly contentious. In describing A. nyktos, Rich et al. (Citation1997) noted that this species possessed an unusual mosaic of character states: three tribosphenic molars per jaw quadrant; a graduated transition from molar to premolar morphology (reflecting the presence of a molariform posterior-most premolar); a dentary with a distinct Meckelian groove and facets for surangular and coronoid bones; and no inflection of the angle of the dentary. As such, Rich et al. (Citation1997) proposed the alternative hypotheses that: (1) A. nyktos might be a eutherian; (2) A. nyktos might belonged in a sister group to Eutheria; or (3) A. nyktos could be a tribosphenic mammal unrelated to Eutheria.
Kielan-Jaworowska et al. (Citation1998, p. 31) disputed this hypothesis (1) arguing that the plesiomorphic dentary of A. nyktos indicated that it was possibly a ‘relic of a more ancient radiation’ that had derived from primitive symmetrodonts. Rich et al. (Citation1998) accordingly responded with a suggestion that the stem placental mammals Prokennalestes trofimovi Kielan-Jaworowska & Dashzeveg, Citation1989 and Prokennalestes minor Kielan-Jaworowska & Dashzeveg, Citation1989 (whose affinities have never been contested) also possess a Meckelian groove and facet on the dentary for a coronoid bone. Moreover, the species of Prokennalestes Kielan-Jaworowska & Dashzeveg, Citation1989 are considered stem placentals based on their dental formula and the gradual morphological change between the molars and premolars (Kielan-Jaworowska & Dashzeveg, Citation1989); both characteristics shared with A. nyktos (Rich et al. Citation1998).
In describing Ambondro mahabo Flynn, Parrish, Rakotosamimanana, Simpson & Wyss, Citation1999, from the middle Jurassic of Madagascar, Flynn et al. (Citation1999) hypothesised that tribosphenic mammals could have originated in Gondwana before spreading to the northern continents during the mid-Cretaceous. The discovery of Mesozoic tribosphenic mammals from India (Prasad et al. Citation2015) and South America (Rauhut et al. Citation2002, Rougier et al. Citation2007) subsequently demonstrated that Mesozoic, tribosphenic mammals were widely distributed in Gondwana ().
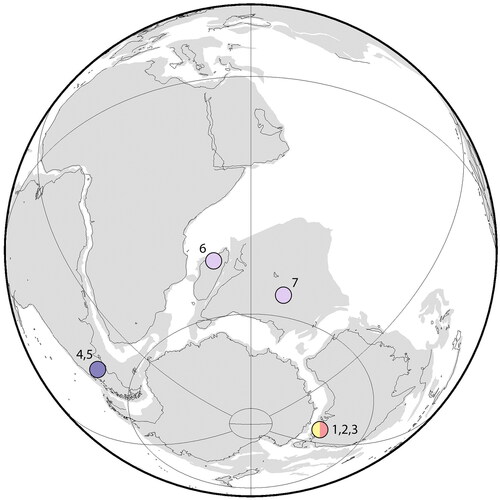
Figure 1. Palaeogeographical reconstruction of the southern landmasses at ca 125 Ma showing the locations of Mesozoic Southern Hemisphere tribosphenidan mammals. Australia: (1) Bishops whitmorei Barremian–Albian, Flat Rocks and Eric the Red West sites; (2) Ausktribosphenos nyktos Barremian, Flat Rocks site; (3) Kryoparvus gerriti Barremian, Flat Rocks site (Rich et al. Citation1997, Citation2001, Citation2020). Argentina: (4) Asfaltomylos patagonicus Callovian, Cañadón Asfalto Basin; (5) Henosferus molus Toarcian, Queso Rallado locality (Rauhut et al. Citation2002, Rougier et al. Citation2007). Madagascar: (6) Ambondro mahabo Bathonian, Mahajanga Basin (Flynn et al. Citation1999). India: (7) unnamed taxon, Lower–Middle Jurassic, Pranhita Godvari valley (Prasad et al. Citation2015). Map created using the ODSN (Ocean Drilling Stratigraphic Network) Plate Tectonic Reconstruction Service (https://www.odsn.de/odsn/services/paleomap/paleomap.html).
Luo, Cifelli & Kielan-Jaworowska (Citation2001) advanced a higher-level classification of mammals that included the Mesozoic Southern Hemisphere tribosphenidans. Luo et al. (Citation2001) discarded the infraclass Tribosphenida (McKenna, Citation1975) and instead advocated a division of the subclass Holotheria (sensu Wible et al. Citation1995) into Boreosphenida (for the metatherians and eutherians originating in the Northern Hemisphere) and Australosphenida (for the Southern Hemisphere Mesozoic tribosphenidans and monotremes). To demonstrate this hypothesis, Luo et al. (Citation2001) presented a parsimony-based phylogenetic analysis of morphological data. At that time, only dentaries and lower dentitions were available for Southern Hemisphere tribosphenic Mesozoic mammals (Rich et al. Citation2020). Luo et al. (Citation2001), therefore, carried out two parallel analyses: one restricted to 55 dentary and lower dental characters; and a second integrating 70 additional cranial and postcranial characters. Sigogneau-Russell et al. (Citation2001) re-examined the two dental characters that served to define Australosphenida following Luo et al. (Citation2001): presence of a continuous mesial and lingual cingulid, which was originally considered plesiomorphic within Tribosphenida and variably expressed in other taxa; and a fully triangulated trigonid on the posterior premolar, which is actually unknown in many australosphenidan taxa and is morphologically contrasting in A. mahabo and A. nyktos. Accordingly, Sigogneau-Russell et al. (Citation2001) concluded that these dental states were incongruous with the Boreosphenida versus Australosphenida subdivision, and therefore considered it likely that the tribosphenic condition arose in clades that were distributed across the Southern Hemisphere. Other studies relying on the Luo et al. (Citation2001) dataset as a framework have since expectedly produced the same topology that has come to entrench the Australosphenidan hypothesis (Luo et al. Citation2002, Kielan-Jaworwska et al. Citation2004, Rougier et al. Citation2007, Wible et al. Citation2009, Pian et al. Citation2016, Luo et al. Citation2017, Celik & Phillips Citation2020, King & Beck Citation2020).
Rich et al. (Citation1997) and Woodburne et al. (Citation2003) put forward dissenting views that Southern Hemisphere tribosphenidans were eutherians or basal eutherians. Rougier et al. (Citation2007, p. 25) alternatively concluded that ‘Australosphenidans are bracketed between “symmetrodonts” (Tinodon Marsh, Citation1879; Zhangheotherium Hu, Wang, Luo & Li, Citation1997) and basal cladotheres (Amphitherium de Blainville, Citation1837, dryolestoids)’, but noted that ‘better specimens of australosphenidans have the potential to drastically change the topology of the [mammalian evolutionary] tree and the conclusions that can be drawn from it’ (Rougier et al. (Citation2007, p. 31). Hunter (Citation2004) subsequently regarded A. nyktos to be non-tribosphenic based on a re-evaluation of cusp homologies and wear facets (this reinterpreted facets V and VI of Crompton Citation1971 as expanded wear facets I and III), thereby positing that the upper molar of A. nyktos had an enlarged paracone that occluded with the hypoconid (renamed the labial cingular cusp in Hunter Citation2004). By contrast, Davis (Citation2011) mentioned that the holotype (NMV P208090) of A. nyktos lacked corresponding areas of the molar crowns, and thus found no evidence for a wear facet III in this taxon. Davis (Citation2011) further identified Kuehneotherium Kermack, Kermack & Musett, Citation1968 from the Late Triassic–Early Jurassic of Europe and Greenland as being a structural precursor to the tribosphenic condition. The earliest tribosphenidan according to Davis (Citation2011) is Tribactonodon Sigogneau-Russell, Hooker & Ensom, Citation2001 from the Berriasian (lowermost Cretaceous) of Britain, which is known from a single lower molar with almost no wear, making it impossible to reconstruct the occluding upper molar morphology. Davis (Citation2011), nevertheless, concurred with Luo et al. (Citation2001) in recognizing Australosphenida, but proposed that Ambondro and henosferids should be removed from the ancestral lineage with A. nyktos and Bishops whitmorei Rich, Flannery, Trusler, Kool, Van Klaveren, & Vickers-Rich, Citation2001, and monotremes on the grounds that Ambondro and henosferids probably had a protocone on the upper molar, while A. nyktos and B. whitmorei presumably possessed upper molars comparable to the stem ornithorhynchid Monotrematum Pascual, Archer, Jaureguizar, Prado, Godthelp & Hand, Citation1992.
Rougier et al. (Citation2021) re-examined Australosphenida within the context of Gondwanan Mesozoic mammal radiations, determining that previous studies placing australosphenidans at the base of the Theria (Rougier et al. Citation2007, Krause et al. Citation2014) were equally as parsimonious as the widely accepted Australosphenida versus Boreosphenida split. The accompanying suggestion that monotremes were derived from Mesozoic Southern Hemisphere tribosphenic mammals has also influenced these debates because the dental and mandibular morphology of monotremes has been used to interpret the highly incomplete Mesozoic Southern Hemisphere tribosphenic mammal record (e.g., Rougier et al. Citation2007, Davis Citation2011). However, most recently Flannery et al. (Citation2022) contended that monotremes are unrelated to Southern Hemisphere tribosphenidans because they fundamentally lack tribosphenic molars, and that the most plesiomorphic monotreme, Teinolophos trusleri Rich, Vickers-Rich, Constantine, Flannery, Kool & van Klaveren, Citation1999, has five molars per jaw quadrant rather than three as in all Mesozoic Southern Hemisphere tribosphenidans. Fossil monotremes that stratigraphically predate Cenomanian strata have four or more molars (Flannery et al. Citation2022). Yet, from the Late Cretaceous onwards most monotreme lineages appear to have standardized with three or less molars, prompting the Flannery et al. (Citation2022) conclusion that early monotremes could not have reverted from a tribosphenic molar condition, as well as independently decreasing from a higher molar number, but instead retain the plesiomorphic, non-therian condition for these states.
Finally, recent discoveries of another new Southern Hemisphere tribosphenic mammal, Kryoparvus gerriti Rich, Trusler, Kool, Pickering, Evans, Siu, Maksimenko, Kundrat, Gostling, Morton, & Vickers-Rich, Citation2020 from the ‘Wonthaggi Formation’ strata at the Flat Rocks site in Australia, as well as the first upper molars of Mesozoic Southern Hemisphere tribosphenidans from both Australia and South America (Rich et al. Citation2020), have provided impetus for the review of Mesozoic Southern Hemisphere tribosphenic mammal classification that we present herein, along with a discussion of their significance for on-going studies of early mammalian evolution.
Institutional abbreviations
GI PST, Mongolian Academy of Sciences, Section of Palaeontology and Stratigraphy, Ulan Bator, Mongolia; MEPF, Museo Paleontólogico Egidio Feruglio, Trelew, Argentina; MPM PV, Vertebrate Paleontology Collection, Museo Regional Provincial Padre Jesús Molina, Río Gallegos, Argentina; NMV, Melbourne Museum, Museums Victoria, Melbourne, Australia; QM, Queensland Museum, Brisbane, Australia; STM, Tianyu Museum of Nature, Linyi, Shandong Province, China; UA, University of Antananarivo, Antananarivo, Madagascar.
Methodological rationale
Because our aim is to present a classification synthesis, we have not undertaken a computational phylogenetic analysis for this study. Additionally, our dataset comprises only six terminal taxa, which is disproportionately influenced by missing character state information (Kearney & Clark Citation2003). There is also considerable disagreement about state interpretations and polarities (see the molar facet descriptions in Crompton Citation1971). Moreover, Rougier et al. (Citation2021) noted that algorithmic phylogenies can support either the australosphenidan or tribosphenidan hypotheses based on preferred priors. We therefore present our review from the perspectives of geochronology, zoogeography and comparative morphology focused on a few unambiguous functional–adaptive complexes including the presence of a talonid basin and its inferred occluding protocone, stylar cusp morphology, and the progressive loss of plesiomorphic features of the dentary. We anticipate that access to new fossils and larger, more complete datasets will phylogenetically resolve these contentions in the future.
Systematic palaeontology
Infraclass TRIBOSPHENIDA McKenna Citation1975
Remarks
We use the infraclass Tribosphenida based on a recent assessment of Southern Hemisphere Mesozoic tribosphenidans upper molars (Rich et al. Citation2020), together with geochronological, zoogeographical and morphological information. While Australosphenida, sensu Luo et al. (Citation2001), was based solely on lower teeth and dentaries, McKenna (Citation1975) defined Tribosphenida on the basis of a single character—the presence of a protocone on the upper molars. The discovery of upper molars with distinct protocones and wear facets V and VI (sensu Crompton Citation1971) attributed to the genus Bishops (Rich et al. Citation2020) unequivocally demonstrates that tribosphenidans (sensu McKenna Citation1975) were present during the Mesozoic in the Southern Hemisphere.
All Southern Hemisphere Mesozoic tribosphenidans (most of which are represented only by lower molar specimens) possessed a talonid basin, which often preserves wear facets. While there is no consensus regarding the number and/or nature of these facets, it is clear that a protocone or protocone-like structure occluded in the talonid basin. Luo et al. (Citation2001) argued that a protocone must have appeared convergently in Southern Hemisphere versus Northern Hemisphere lineages; however, there is only weak topological support for this hypothesis (Rougier et al. Citation2021).
Zoogeography also supports a single monophyletic origin for the protocone in Southern Hemisphere Mesozoic tribosphenidans ca 50 million years (Ma) before its appearance in Northern Hemisphere taxa. Over that time, considerable evolutionary change must have occurred in the dentition and dentary of these Southern Hemisphere lineages, with Jurassic forms retaining an extremely plesiomorphic dentary and premolar morphology, and molars with narrow talonid basins. Cretaceous Southern Hemisphere tribosphenidans have premolars, molars and dentaries that more closely resemble those of Northern Hemisphere representatives.
An alternative hypothesis is that Early Cretaceous peramurids are ancestral to Tribosphenida (McKenna & Bell Citation1997). Peramurids lack both post dentary bones and protocones on their molars, yet possess features present in Tribosphenida including a large stylar cusp C (Kraus Citation1979, Davis Citation2012). Early Cretaceous European peramurids displayed a highly autapomorphic enlarged masseteric foramen on the dentary, which is absent in most Tribosphenida, and therefore likely precludes them from any direct ancestry of Tribosphenida (Davis Citation2012). The oldest probable peramurans were present in the Late Jurassic of Tanzania, hinting at a possible Gondwanan origin for the group (Davis Citation2012). We subsequently consider it possible that Tribosphenida could have arisen from a peramuran ancestor in the Early Jurassic of Gondwana, and that the absence of earlier annectant forms likely reflects incompleteness of the fossil record. The sudden appearance of advanced tribosphenidans in the Northern Hemisphere accordingly suggests that the Boreal distribution of Tribosphenida initiated with migration from the Austral regions.
Members of the family Shuotheriidae possessed pseudo-tribosphenic lower molars with a talonid-like basin on the trigonid and a ‘pseudo-protocone’ on the upper molars (Luo et al. Citation2007). The pseudo-protocone of shuotheriids is independently derived since it occludes with the pseudo-talonid on the mesial side of the trigonid rather than the talonid. The existence of a pseudo-tribosphenic mammalian dentition implies that the tribosphenic condition could feasibly have arisen more than once; however, it does not necessarily require that the tribosphenic condition developed convergently multiple times within Tribosphenida.
Shuotheriids were designated as the sister group of Southern Hemisphere Mesozoic tribosphenic mammals by Kielan-Jaworowska et al. (Citation2004) on the basis of a shared trigonid-like posterior-most lower premolar, slender dentaries, and the presence of a postdentary groove. We concur with Kielan-Jaworowska et al. (Citation2004) that these features are similar, but note that henosferids possessed much simpler posterior-most premolar morphologies than shuotheriids, which infers convergence between shuotheriids and Ausktribosphenos nyktos. Importantly, shuotheriids are also stratigraphically restricted to the mid-Jurassic in the Northern Hemisphere (England and China: Flynn et al. Citation1999, Rougier et al. Citation2007), a time when tribosphenidans were already widespread throughout the southern Gondwanan landmasses.
Family HENOSFERIDAE Rougier, Martinelli, Forasiepi & Novacek, Citation2007
Diagnosis
Molars with a trigonid and three cusps arranged in a triangle, a talonid basin without occlusive wear, and an absence of facets V and VI (sensu Crompton Citation1971). The premolars are simple with an abrupt transition in morphology across the molar–premolar boundary. The posterior-most premolar has a small and shallow posterior cingulum. The dentary has a strongly developed medial flange preserving evidence of articulation with the postdentary bones. A postdentary groove, a deep dentary trough, and a transversely wide, spoon-like angular process are also present.
Remarks
The henosferid Asfaltomylos patagonicus Rauhut, Martin, Ortiz-Jaureguizar & Puerta, Citation2002 is known from a single crushed, partial dentary (MEPF-PV 1671) found in the Toarchian (Early Jurassic) Cañadón Asfalto Formation of Chubut Province in Argentina (; Martin & Rauhut Citation2005). The dentary hosts partial crowns of three molars and roots for three teeth interpreted as premolars. The molars exhibit highly specialized apical wear on the hypoconid, hypoconulid and the hypocristid (Martin & Rauhut Citation2005). Their talonid basins are also wider than long and lack Crompton’s (Citation1971) wear facets V and VI (Martin & Rauhut Citation2005). All of the presumed premolar teeth are double-rooted, with the posterior-most reconstructed with a simple triangular profile. The transition from molars to premolars is therefore likely to have been abrupt. The dentary has a postdentary groove, and the mandibular foramen is anteriorly positioned.
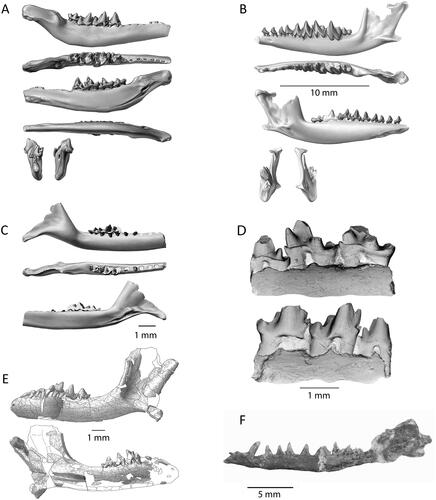
Figure 2. Mesozoic Southern Hemisphere tribosphenidan mammal dentaries. A, Ausktribosphenos nyktos (NMV P208090) in buccal, occlusal, lingual, ventral, anterior and posterior views, modified from Rich et al. (Citation1997). B, Bishops whitmorei (NMV P 210075) in buccal, occlusal, lingual, anterior and posterior views, modified from Rich et al. (Citation2001). C, Kryoparvus gerriti (NMV P210087) in buccal, occlusal and lingual views, modified from Rich et al. (Citation2020). D, Ambondro mahabo (UA 10602) in lingual and buccal views, modified from Flynn et al. (Citation1999). E, Asfaltomylos patagonicus (MEPF-PV 1671) in buccal and lingual views, modified from Martin & Rauhut (Citation2005). F, Henosferus molus (MEFP 2354) in lingual stereophotograph view, modified from Rougier et al. (Citation2007).
Henosferos molus Rougier, Martinelli, Forasiepi & Novacek, Citation2007 is known from two near-complete dentaries and an isolated upper premolar (MPEF 2353, MPEF 2354, MPEF 2355) from the Toarcian (Early Jurassic) Cañadón Asfalto Formation in Argentina—making them and A. patagonicus the oldest known tribosphenids (Cùneo et al. Citation2013). Henosferos molus is similar to A. patagonicus, but is about twice the size, and exhibits distinct diastemata between the premolars. The dentary has a prominent medial flange evidencing an articulation with postdentary bones, a deep dentary trough, a Meckelian groove, small and deep coronoid facet, and a transversely broad angular process. The dental formula of H. molus is i4, c1, p5, m3, and there is an abrupt transition between the premolars and molars. The molars are tribosphenic, with a trigonid and fully basined talonid. The m2 of MPEF 2354 shows the main cusps of the trigonid forming an angle of 85°; 25% greater than on the m1. It is uncertain whether wear facets IV and V (sensu Crompton Citation1971) are present, and wear facet VI is absent. Similar to A. patagonicus, no wear facet is discernible in the talonid basin (Rougier et al. Citation2007). The hypoconulid is very small and closely adpressed against the entoconid, which is distinct. In this regard, H. molus closely resembles Ausktribosphenos nyktos and Bishops whitmorei (Rich et al. Citation1997, Rich et al. Citation2001, Rougier et al. Citation2007).
Henosferids are stratigraphically restricted to the Early to Middle Jurassic, and share various plesiomorphic features with non-therian mammals (Rougier et al. Citation2007). Henosferids also possessed plesiomorphic states that differentiate them from A. nyktos and Bishops spp.: the possession of a dentary shelf with evidence for articulation with postdentary bones (), the transversely broad spoon-like dentary angle, an abrupt transition between premolars and molars, narrow molars with narrow talonid basins () and the lack of wear facet VI (and possibly wear facet V: sensu Crompton Citation1971) on the molars (Rich et al. Citation1997, Rich et al. Citation2001).
Figure 3. Mesozoic tribosphenidan mammal Meckelian grooves (arrows). A, Ausktribosphenos nyktos (NMV P208090), modified from Rich et al. (Citation1997). B, Bishops whitmorei (NMV P 210075), mirrored from Rich et al. (Citation2001). C, Henosferus molus (MEFP 2353) stereophotograph, modified from Rougier et al. (Citation2007). D, Asfaltomylos patagonicus (MEPF-PV 1671), mirrored from Rauhut et al. (Citation2002). E, Prokennalestes minor (GI PST 10-7a) SEM micrograph in posterior aspect, modified from Rich et al. (Citation1998) F, Ambolestes zhoui (STM33-5), modified from Bi et al. (Citation2018).
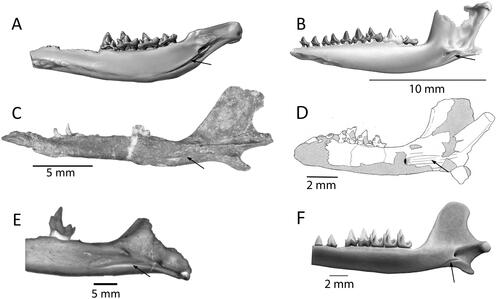
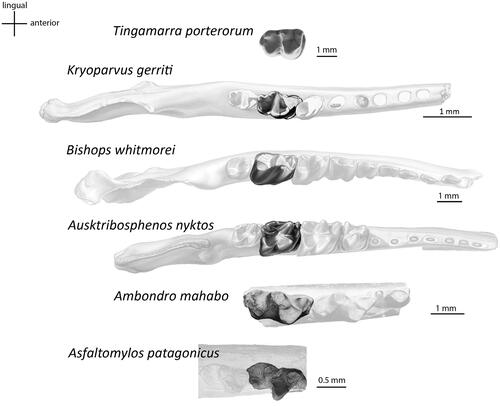
Figure 4. Occlusal views of lower second molars. Arrangement (top to bottom): Tingamarra porterorum (QMF20564), modified from Godthelp et al. (Citation1992); Kryoparvus gerriti (NMV P210087), modified from Rich et al. (Citation2020); Bishops whitmorei (NMV P210075), modified from Rich et al. (Citation2001); Ausktribosphenos nyktos (NMV P208090), modified from Rich et al. (Citation1997); Ambondro mahabo (UA-10602), modified from Flynn et al. (Citation1999); Asfaltomylos patagonicus (MEPF PV 1671), modified from Martin & Rauhut (Citation2005).
Family ?HENOSFERIDAE
Remarks
Prasad et al. (Citation2015) informally reported a tribosphenic lower molar from the Early–Middle Jurassic Kota Formation near Telengana in India. This tooth apparently has a trigonid with three cusps arranged in an obtuse triangle, and a talonid that is wider than long (though not as wide as that in ausktribosphenids: see Rich et al. Citation1997). It closely resembles the lower molars of Asfeltomylos patagonicus and Henosferos molus; therefore, we tentatively classify it as ?Henosferidae.
Family INDETERMINATE
Remarks
Ambondro mahabo is based on a single dentary fragment (UA 10602) containing three teeth, interpreted as the last premolar and first two molars, recovered from the Middle Jurassic Isalo III Formation of Madagascar, which has been radiometrically dated to the Bathonian 168.3 ± 1.3 Ma (Flynn et al. Citation1999). The molars are tribosphenic, and the trigonid is ‘open’ with crests from the metaconid and paraconid that form a right angle with the protoconid. There is a well-defined talonid basin with two wear facets (V and VI: sensu Crompton Citation1971) for occlusion with a protocone. These molars closely resemble those of henosferids. The dentary formula cannot be ascertained; however, the posterior-most premolar has a broad and deep posterior cingulum enclosing a prominent cuspule (). This is unique among Southern Hemisphere Mesozoic tribosphenic mammals, making the posterior moiety of the premolar morphologically similar to the talonid region on the molars. Moralization of the posterior-most premolar is also present in shuotheriids Ausktribosphenos nyktos and Bishops whitmorei (Rich et al. Citation1997, Rich et al. Citation2001).
Martin & Rauhut (Citation2005) challenged the interpretation that wear facets V and VI (sensu Crompton Citation1971) are present in A. mahabo. Rougier et al. (Citation2007) was likewise uncertain about their presence, and suggested that Henosferidae could be the sister lineage to A. mahabo. Davis (Citation2011) considered the molars of A. mahabo to be most similar to those of henosferids. Woodburne et al. (Citation2003) otherwise disputed the concept of Australosphenida and regarded A. mahabo to be a basal eutherian. Unfortunately, UA 10602 is so incomplete that these key character states remain ambiguous, making it impossible to adequately classify A. mahabo to family-level.
Family AUSKTRIBOSPHENIDAE Rich, Vickers-Rich, Constantine, Flannery, Kool & van Klaveren, Citation1997
Remarks
Ausktribosphenidae includes Ausktribosphenos nyktos and Kryoparvus gerriti. All exemplars of this family derive from the uppermost Barremian ‘Wonthaggi Formation’ of Australia (Rich et al. Citation2020). Ausktribosphenids differ from Bishops spp. in possessing a much more pronounced Meckelian groove, subangular and coronoid facets on the dentary, and in lacking a talonid-like structure on the rear of the posterior-most premolar. Compared to henosferids, ausktribosphenids lack a medial flange on the dentary (Rich et al. Citation1997) and possess a true dentary angle rather than a transversely broad spoon-shaped process (Lopatin & Averianov Citation2017). The posterior-most premolar also resembles the trigonid region of the molars.
Kryoparvus gerritti is known from two partial dentaries with fragmentary molars (NMV P210087, NMV P212490). Ausktribosphenos nyktos is represented by six dentaries (NMV P208090, NMV P 208228, NMV P 208482, NMV P216670, NMV P 232567) from the Flat Rocks site, and referred dentary NMV P257142 from the recently discovered Honey Locality, which is approximately 450 m south of Flat Rocks (Rich et al. Citation2022).
Kryoparvus gerritti is arguably the smallest-bodied Mesozoic mammal (Rich et al. Citation2020). It possesses a Meckelian groove on the dentary similar to that of A. nyktos and has prominent basal cingula on the anterobuccal region of the lower molars. Its molars, however, differ from those of both A. nyktos and Bishops spp. in that the trigonid is wider and more open, and there is no evidence of the ‘unnamed cristid 2 or 3’ described by Rich et al. (Citation1997) on the partially preserved crowns. While these are incomplete, crests in the area where the ‘unnamed cristid 2 or 3’ should occur were absent ().
Ausktribosphenos nyktos and Bishops spp. uniquely share: (1) the presence of an ‘unnamed cristid 2’ on the talonid crest of the lower molars; (2) an entocristid (reduced or absent on the molars of Bishops spp.). These features are absent in all Northern Hemisphere Mesozoic tribosphenidans. Ausktribosphenidans and Bishops spp. additionally possessed an angular process on the dentary comparable to that of therians (Averianov & Lopatin Citation2014).
Family BISHOPIDAE fam. nov.
Diagnosis
The molars of Bishops whitmorei differ from those of Ausktribosphenos nyktos in the m2 being the largest molar and the m3 being relatively small in size and in being more exaenodont and having a less extensive precingulid. The basal cingula on the lower molars are also reduced relative to those on A. nyktos (Rich et al. Citation2001). The posterior region of the posterior-most premolar consists of a broad talonid-like shelf that imparts a bilobed tooth profile in occlusal view. Bishops whitmorei specifically possesses six premolars, compared to five in A. nyktos. The dentary of B. whitmorei differs from that of henosferids and ausktribosphenids in having a greatly reduced Meckelian groove and no surangular or coronoid facet. The weak Meckelian groove is well separated from the mandibular foramen; this, together with location of the hypoconulid near the midline of the tooth and separated from the entoconid, renders B. whitmorei ‘more like the vast majority of placental mammals’ than A. nyktos (Rich et al. Citation2001, p. 7), or any other Mesozoic Southern Hemisphere tribosphenidan.
Remarks
Bishops whitmorei is represented by numerous dentaries and isolated teeth (NMV P210075, NMV P208484, NMV P209975, NMV P210070, NMV P212810, NMV P212950, NMV P221043, NMV P221157, NMV P229649, NMV P216578, NMV P232892) from the uppermost Barremian ‘Wonthaggi Formation’ at the Flat Rocks site. Other dentary fragments and upper molars (NMV P228848, NMV P231328, NMV P252207: see Rich et al. Citation2009, Citation2020) have also been recovered from the lower Albian Eumeralla Formation at the Eric the Red West site in the Otway Basin of Victoria (see Poropat et al. Citation2018 for summary). Lastly, an isolated upper molar fragment (MPM-PV-22879) attributed to ‘aff. Bishops’ has been reported from the Cenomanian Mata Amarilla Formation in southern Patagonia, Argentina (Martin et al. Citation2022).
The holotype left dentary of B. whitmorei (NMV P210075) is by far the most complete specimen yet recovered. It includes the articular process with most of the angle preserved, part of the ascending ramus, and the anterior ramus to the base of the incisor alveoli. There are three molars and six premolars. The dentary tapers anteriorly to an extremely shallow depth under the incisors. Judging from its alveolus, the canine must have been subequal in size to the anterior-most premolar. The m3 is greatly reduced in size relative to the other molars, and the Meckelian groove is reduced to a shallow depression immediately posteroventral to the mandibular foramen. The Meckelian groove and mandibular foramen are separated by a ridge ().
In comparison with the incomplete upper molars (NMV P231328) of Bishops sp. from the lower Albian Eumeralla Formation (Rich et al. Citation2020), the upper molar fragment (MPM-PV-22879) attributed to ‘aff. Bishops’ from the Cenomanian Mata Amarilla Formation of Patagonia () displays sufficiently different morphology to be placed in a separate genus, currrently unnamed, which we also consider to belong within the family Bishopidae. NMV P231328 consists of two heavily abraded and broken upper molars, both with a prominent protocone and a prominent basal cingulum. The basis for inferring an abraded protocone on the molars was discussed at length by Rich et al. (Citation2020). The more complete of the two (possibly M3), hosts a well-preserved stylar cusp C and a crushed and distorted stylar cusp B. Conversely, MPM-PV-22879 lacks its posterior and lingual margins, so that the protocone and metacone, if ever present, are not preserved. Nonetheless, the crown was considerably more buccolingually compressed than NMV P231328, and the twinned postparacrista evident on NMV P231328 are represented by a simple crest in MPM-PV-22879.
Figure 5. Occlusal views of upper molars assigned to Bishopidae. A, MPM-PV22879, modified from Martin et al. (Citation2022). B, NMV P231328, modified from Rich et al. (Citation2020). Reconstruction of MPM-PV22879 homologous to NMV P231328 (grey shading) modified from Martin et al. (Citation2022).
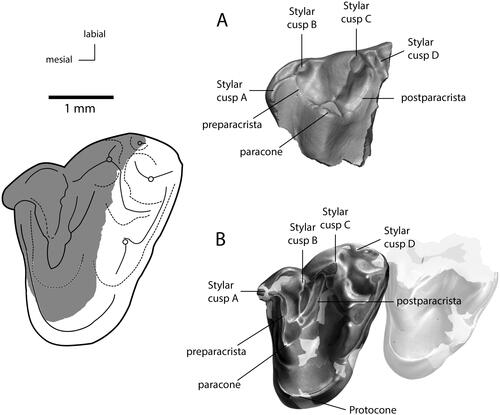
NMV P231328 and MPM-PV-22879 are strikingly similar in possessing four stylar cusps (stylar cusps A–D), the largest of which is stylar cusp C, followed in size by cusp B and stylar cusps D and A. Stylar cusp A may have been incorporated into a basal cingulum in both specimens. NMV P231328 also preserves evidence for stylar cusp E (the area of the crown where this cusp might have occurred is not preserved on MPM-PV22879). We are uncertain whether stylar cusp D is homologous in NMV P231328 and MPM-PV-22879, as it is slightly differently positioned. These teeth all have a breached paracone.
The presence of stylar cusps, particularly a large central stylar cusp (stylar cusp C), has been considered a key character state of metatherians (Jacobs et al. Citation1989), including Holoclemensia texana Slaughter, Citation1968 from the Aptian–Albian Trinity Formation of Texas and Oklahoma, USA. This taxon is known only from isolated teeth but has long been considered a metatherian based on its possession of four stylar cusps, including a prominent stylar cusp C (Davis & Cifelli Citation2011). The reconstructed dental formula and size of the stylocone and wide, flange-like parastylar lobe prompted Davis & Cifelli (Citation2011, p. 458) to reassign H. texana to Eutheria and interpret its stylar cusps as ‘a curious apomorphy … immaterial to its broader relationships’. An alternate interpretation is that H. texana is indeed a eutherian, but one which retains a plesiomorphic stylar cusp configuration shared with many metatherians and with bishopids.
We propose that the upper molars of NMV P231328 are consistent with the lower molars of Bishops on the basis of geographical, stratigraphical and morphological proximity. All were recovered over multiple years from the highly localized ca 75 × 10 m2 Eric the Red West site fossil excavation horizon within the Eumeralla Formation (Rich et al. Citation2009, Citation2020). The teeth also represent similar-sized animals (Rich et al. Citation2020) and share the diagnostic presence of basal cingula. Furthermore, the twinned postparacrista and premetacrista match an infolding of enamel on the talonids of the lower molars (). This is a highly unusual feature that is unknown in therians and other Southern Hemisphere tribosphenidans (Rich et al. Citation2020). Moreover, the holotype m1 of B. whitmorei (NMV P210075) can be reconstructed to precisely occlude with the anterior molar shear facets V and VI (sensu Crompton Citation1971) of NMV P231328 (Rich et al. Citation2020). Given this evidence, the hypothetical upper molar morphology of Bishops spp., which was predicted to be Monotrematum-like by Davis (Citation2011) or bearing an enlarged paracone occluding with the hypoconid (Hunter Citation2004), is in fact tribosphenic.
Figure 6. Stratigraphical ranges of Pangean mammaliaforms (left), Southern Hemisphere Gondwanian mammals (Middle), and Northern Hemisphere Laurasian mammals (right) from the Late Triassic to Cretaceous/Tertiary boundary (ca 66 Ma). Possible phylogenetic relationships (dashed lines) and hypotheses (solid lines) are discussed in the text.
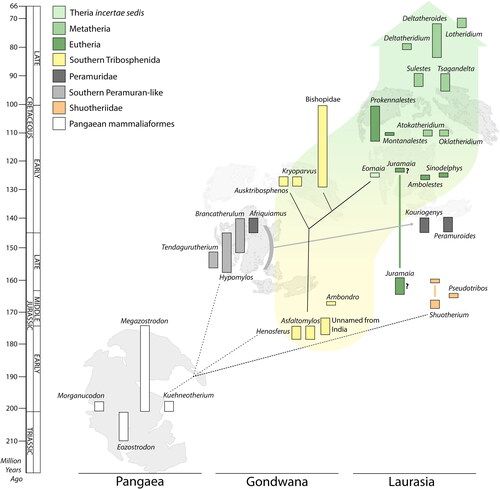
In summary, bishopids possessed a unique combination of character states. (1) Like eutherians and metatherians they have no dentary groove or postdentary bones. (2) The presence of three molars per jaw quadrant is a stem therian trait shared with shuotheriids. (3) The presence of large stylar cusps (especially stylar cusp C) is compatible with basal metatherians (Jacobs et al. Citation1989) and some basal therians (Davis & Cifelli Citation2011). Thus, bishopids appear to be the sister group to therians (eutherians and metatherians) whose evolution subsequently involved an increase in molar number through suppression of dental replacement in metatherians (as suggested by Williamson et al. Citation2016), and a loss of the stylar cusps in eutherians.
Discussion
Morphological evolution within Mesozoic Southern Hemisphere Tribosphenida
Considerable morphological change is evident between members of the Jurassic Henosferidae and the Cretaceous Ausktribosphenidae and Bishopidae. Henosferidans retain a plesiomorphic dentary structure, with an extensive dentary shelf and articular facets for the postdentary bones. In this, Henosferus molus is strikingly similar to species of the non-tribosphenic Late Triassic-earliest Jurassic mammaliform genus Morganucodon Kühne, Citation1949 (Rougier et al. Citation2007). These features are lost to varying degrees in ausktribosphenids and bishopids, leading to the development of a dentary in Bishops whitmorei that lacks postdentary bones and facets, thus being very similar to that of most therians (Rich et al. Citation2001).
Changes are also observable in the dentition. For example, the lower molars of henosferids differ from those of ausktribosphenids and bishopids in being elongate, with relatively narrow talonid basins, while in ausktribosphenids and bishopids the lower molars are broad with wide talonid basins (). In henosferids the five premolars are simple, laterally compressed cones, sometimes with small anterior and posterior cuspules (Rougier et al. Citation2007). By contrast, the posterior-most premolars of ausktribosphenids and bishopids are somewhat molariform, with that of A. nyktos resembling a molar trigonid, versus that of B. whitmorei which has an additional posterior talonid-like structure, giving the tooth an even more molariform shape ().
Stratigraphical range of Jurassic and Cretaceous tribosphenids
Southern Hemisphere tribosphenidans are known from the Mesozoic of Gondwana (Australia, India, Madagascar and South America). They persisted from the Early Jurassic (Toarcian, 182.7–174.1 Ma) until the Late Cretaceous (Cenomanian, 100.5–93.9 Ma), giving the lineage a duration of between ca 73.6-88.1 Ma ().
The Jurassic record of Northern Hemisphere tribosphenidans is controversial. Juramaia sinensis Luo, Yuan, Meng & Ji, Citation2011, was originally reported from the mid-Late Jurassic (ca 160 Ma) Daxigou site in the Tiaojishan Formation of China. The Daxigou site has also yielded Rugosodon eurasiaticus Yuan, Meng, Ji, Tabrum & Luo, Citation2013, which is purported to be the oldest known paulchoffatoid multituberculate. Bi et al. (Citation2018) and Meng (Citation2014), however, questioned the age estimate of the site based on the geological complexity and geographically widespread nature of the Tiaojishan Formation outcrops. Furthermore, a tip-dated analysis (King & Beck Citation2020, p. 1) suggested that J. sinensis is probably not contemporanenous with other Jurassic age fossils from the same biota, recovering ‘an Early Cretaceous age for J. sinensis, but similar analyses on 12 other mammaliaformes from the Yanliao Biota return the correct, Jurassic age’.
The oldest uncontested Northern Hemisphere tribosphenidan is Eomaia scansoria Ji, Luo, Yuan, Wible, Zhang, & Georgi, Citation2002 from the upper Barremian (ca 129.4–125 Ma) Dawangzhangzi locality of the Yixian Formation in Liaoning Province, northeastern China. Although E. scansoria was originally described as a basal eutherian, it lacks some diagnostic eutherian hind limb characteristics, has five upper and four lower incisors typical of metatherians but not eutherians, and five premolars (similar to Southern Hemishere tribosphenidans), leading O’Leary et al. (Citation2013, p. 665) to suggest that it ‘falls outside of Eutheria as a stem taxon of Theria’.
The eutherian Ambolestes zhoui Bi, Zheng, Wang, Cignetti, Wang & Wible, Citation2018 from the Lower Cretaceous Yixian Formation at Xisanjia in Inner Mongolia, China has been dated to ca 126 Ma, and is thus approximately coeval with E. scansoria, along with the basal eutherian Acristatherium yanensis Hu, Meng, Li & Yang, Citation2010 from the Lujiatun bed of the Yixian Formation, which has been dated at 123.2 ± 1.0 Ma.
The earliest metatherian occurrences in the fossil record are likewise debated. Sinodelphys szalayi Luo, Ji, Wible & Yuan, Citation2003, from upper Barremian (125 Ma) deposits of the Yixian Formation near Liaoning in China, is provisionally accepted by Williamson et al. (Citation2016) as the oldest recorded metatherian. However, Bi et al. (Citation2018) convincingly recovered it as a basal eutherian. The oldest undisputed metatherians are members of the order Deltatheroida, which have their geologically oldest records (Atokatheridium Kielan-Jaworowska & Cifelli Citation2001, and Oklatheridium Davis, Cifelli & Kielan-Jaworowska, Citation2008) in the Aptian or Albian (ca 115–106 Ma) of North America (Williamson et al. Citation2016, Eldridge et al. Citation2019). The presently limited fossil evidence therefore points to a divergence timeframe of metatherians versus eutherians to no later than the late Barremian–late Albian (129.4 ± 1.5–100.5 ± 1.0 Ma). Molecular divergence dates produced by Álvarez-Carretero et al. (Citation2022) have alternatively pushed this back to ca 166–123 Ma (early Callovian–early Aptian), thereby providing contextual support for our view that bishopids may be early therians or the sister group to Theria.
Zoogeography
Shuotheriids are known only from the Jurassic of Asia and Europe, while tribosphenidans occur in the Jurassic of Madagascar and South America. It seems possible that this distribution resulted from a vicariance event in the Late Triassic–Early Jurassic, when Pangaea split into Gondwana and Laurasia. The Jurassic and Early Cretaceous of Africa has not yet yielded undoubted evidence of tribosphenic mammals. Instead, the pre-tribosphenic, peramuran-like genera Brancatherulum Dietrich, Citation1927, Tendagurutherium Heinrich, Citation1998 and Hypomylos are present. This implies isolation of the African mammal fauna over this extended interval.
The abrupt appearance of tribosphenidans in Asia during the Barremian is puzzling. The oldest firmly dated tribosphenic mammal reported from North America is Montanalestes keebleri Cifelli, Citation1999, from the Cloverly Formation (Albian, ca 110 Ma) of Montana, and the oldest tribosphenidan from Europe is early Cenomanian (100.5–95 Ma) in age (Vullo et al. Citation2009). The fossil record of Mesozoic mammals is notoriously incomplete, thus it is possible that earlier tribosphenidans will be found in Europe and North America in the future. However, if tribosphenidans did in fact arrive in Asia prior to their appearance in North America and Europe, then the most likely dispersal route is directly from Australia. Palaegeographical reconstructions for the Barremian locate Australia south of Asia, with the expansive Ontong Java Nui igneous province potentially being subaerially exposed in the intervening seaway, thereby providing an island ‘stepping-stone’ for migrating species (Hochmuth et al. Citation2015).
The plesiomorphic morphology of many Cretaceous Northern Hemisphere tribosphenidans suggests that several Southern Hemisphere tribosphenidan lineages may have colonized the Northern Hemisphere. Prokennalestes retains a Meckelian groove, while Eomaia retains both a Meckelian groove and simple, conical lower premolars with anterior and posterior cuspules that are reminiscent of henosferids. Bishops whitmorei, which lacks a Meckelian groove and possesses a molariform posterior-most lower premolar, resembles later Northern Hemisphere tribosphenidans.
A vexing question is why did the Southern Hemisphere Mesozoic tribosphenidan lineage become extinct in Australia? There is a gap of ca 50 Ma between the mid-Cenomanian and early Eocene (Godthelp et al. Citation1992, Flannery et al. Citation2022) in the Australian mammal fossil record that complicates interpretations. The Cenomanian of Australia is dominated by monotremes (Flannery et al. Citation2022), all of which are of relatively large (≥4 kg body mass) and have been recovered from commercial opal mines and spoil heaps, a method that is unlikely to yield the expectedly very small dentigerous remains of tribosphenidans. Thus, aside from biased sampling we are unable to adequately explain the current absence of Mesozoic tribosphenic mammals after the Early Cretaceous in Australia.
The Tingamarra local fauna from stratigraphically undifferentiated lower Eocene strata near Murgon in southeastern Queensland has a minimum radiometric date of 54.6 ± 0.5 Ma, and is dominated by metatherians, many of which resemble South American taxa; this suggests that the assemblage may reflect faunas immediately postdating the arrival of metatherians in Australia (Godthelp et al. Citation1992). The Tingamarra local fauna also includes the possible basal condylarth Tingamarra porterorum Godthelp, Archer, Cifelli, Hand & Gilkeson, Citation1992, whose dental microstructure is unlike that of any metatherian but otherwise resembles archaic eutherians (Godthelp et al. Citation1992). Strikingly, the holotype molar (QM F20564) also possesses basal cingula, a feature seen in many Australian Mesozoic tribosphenic mammals, such as Ausktribosphenos nyktos ().
Ultimately, several hypotheses exist as to the extinction time of Southern Hemisphere Mesozoic tribosphenidans in Australia. It is possible that this group declined shortly after the Albian (their last recorded occurrence), perhaps as a result of competition with the rapidly diversifying monotremes (Flannery et al. Citation2022). Alternatively, Australian tribosphenidans might have disappeared during the end-Cretaceous mass extinction event. A third possibility is that they survived into the early Eocene, and that T. porterorum is a terminal member of this lineage, in which case Australian tribosphenidans could have terminally dwindled as a result of competition with the newly arrived metatherians.
Conclusions
Tribosphenic mammals evolved in the Southern Hemisphere in the Early Jurassic, ca 50 Ma prior to their earliest securely dated occurrence (late Barremian) in the Northern Hemisphere.
Considerable evolutionary change occurred in Southern Hemisphere tribosphenic mammals between the Jurassic and Early Cretaceous, including loss of primitive jaw structures and the development of fully tribosphenic molars bearing wear facets V and VI (sensu Crompton Citation1971).
Bishopids fam. nov. are the immediate sister taxon to therian mammals.
Acknowledgements
The excavation of all Cretaceous mammal fossils from the Victorian deposits was undertaken with the ParksVic permit 10009432, and funded by The National Geographic Society supported by the efforts of +700 volunteers in the field. Steve Morton and Lesley Kool (Monash University) contributed photography, fossil preparation and field logistics. Peter Trusler (Melbourne) provided illustrations. We thank the various curators and collection managers for access to specimens, and the Australian Museum Research Institute for financial support. The Chief Editor and Editorial Board of Alcheringa generously assisted with the compilation, submission and processing of our manuscript.
Disclosure Statement
No potential conflict of interest was reported by the author(s).
References
- Álvarez-Carretero, S., Tamuri, A.U., Battini, M., Nascimento, F.F., Carlisle, E., Asher, R.J., Yang, Z., Donoghue, P.C.J. & Dos Reis, M., 2022. A species level timeline of mammal evolution integrating phylogenomic data. Nature 602, 263–267.
- Archibald, D.J., 2012. Darwin’s two competing phylogenetic trees: marsupials as ancestors or sister taxa? Archives of Natural History 39, 217–233.
- Averianov, A.O. & Lopatin, A.V., 2014. On the phylogenetic position of monotremes (Mammalia, Monotremata). Paleontological Journal 48, 426–446.
- Bi, S., Zheng, X., Wang, X., Cignetti, N.E., Yang, S. & Wible, J.R., 2018. An early Cretaceous eutherian and the placental-marsupial dichotomy. Nature 558, 390–395.
- Butler, P.M., 1939. The teeth of the Jurassic mammals. Proceedings of the Zoological Society of London B109, 329–356.
- Celik, M.A. & Phillips, M.J., 2020. Conflict resolution for Mesozoic mammals: reconciling phylogenetic incongruence among anatomical regions. Frontiers in Genetics 11, 0651.
- Cifelli, R.L., 1999. Tribosphenic mammal from the North American Early Cretaceous. Nature 401, 363–366.
- Cope, E.D., 1883. Note on the trituberculate type of superior molar and the origin of the quadrituberculate. American Naturalist 17, 407–408.
- Crompton, A.W., 1971. The origin of the tribosphenic molar. Zoological Journal of the Linnean Society 50, 65–87.
- Cùneo, R., Ramezani, J., Scasso, R., Pol, D., Escapa, I., Zavattieri, A.M. & Bowring, S.A., 2013. High-precision U-Pb geochronology and a new chronostratigraphy for the Cañadón Asfalto Basin, Chubut, central Patagonia: implications for terrestrial faunal and floral evolution in Jurassic. Gondwana Research 24, 1267–1275.
- Davis, B.M., 2011. Evolution of the tribosphenic molar pattern in early mammals, with comments on the “Dual-Origin” Hypothesis. Journal of Mammalian Evolution 18, 227–244.
- Davis, B.M., 2012. Micro-computed tomography reveals a diversity of Peramuran mammals from the Purbeck Group (Berriasian) of England. Palaeontology 55, 789–817.
- Davis, B.M. & Cifelli, R.L., 2011. Reappraisal of the tribosphenidan mammals from the Trinity Group (Aptian–Albian) of Texas and Oklahoma. Acta Palaeontologica Polonica 56, 441–462.
- Davis, B.M., Cifelli, R.L. & Kielan-Jaworowska, Z., 2008. 1. Earliest evidence of Deltatheroida (Mammalia: Metatheria) from the Early Cretaceous of North America. In Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay. Sargis, E.J. & Dagosto, M. eds. Springer, Dordrect, NL. pp. 3–24.
- De Blainville, H.M.D., 1837. Doutes sur le prétendu Didelphe fossile de Stonefield [sic]. On à quelle Classe, àqelle Famille, à quel Genre doit-on rapporter l'animal auquel out apparttenu les ossements fossiles, à Stonefield, designès sous les noms de Did, pp. 402–418.
- Dietrich, W.O., 1927. Brancatherulum n. g., ein Proplacentalier aus dem obersten Jura des Tendaguru in Deutsch-Ostafrika. Centralblatt fur Mineralogie, Geologie, Palaontologie, Abt B 10, 423–426.
- Eldridge, M.D.B., Beck, R.M.D., Croft, D.A., Travouillon, K.J. & Fox, B.J., 2019. An emerging consensus in the evolution, phylogeny, and systematics of marsupials and their fossil relatives (Metatheria). Journal of Mammalogy 100, 802–837.
- Flannery, T.F., Rich, T.H., Vickers-Rich, P., Ziegler, T., Veatch, E.G. & Helgen, K.M., 2022. A review of monotreme (Monotremata) evolution. Alcheringa 45, 1–18.
- Flynn, J.J., Parrish, J.M., Rakotosamimanana, B., Simpson, W.F. & Wyss, A.R., 1999. A middle Jurassic mammal from Madagascar. Nature 401, 57–60.
- Godthelp, H., Archer, M., Cifelli, R., Hand, S.J. & Gilkeson, C.F., 1992. Earliest known Australian Tertiary mammal fauna. Nature 356, 514–516.
- Heinrich, W.-D., 1998. Late Jurassic mammals from Tendaguru. Journal of Mammalian Evolution 5, 269–290.
- Hochmuth, K., Gohl, K. & Uenzelmann-Neben, G., 2015. Playing jigsaw with Large Igneous Provinces—a plate tectonic reconstruction of Ontong Java Nui, West Pacific. Geochemistry, Geophysics, Geosystems 16, 3789–3807.
- Hu, Y., Wang, Y., Luo, Z. & Li, C., 1997. A new symmetrodont mammal from China and its implications for mammalian evolution. Nature 390, 137–142.
- Hu, Y., Meng, J., Li, C. & Wang, Y., 2010. New basal eutherian mammal from the Early Cretaceous Jehol biota, Liaoning, China. Proceedings of the Royal Society, London. Proceedings. Biological Sciences 277, 229–236.
- Hunter, J.P., 2004. Alternative interpretation of molar morphology and wear in the Early Cretaceous mammal Ausktribosphenos. Journal of Vertebrate Palaeontology 24, 73A.
- Jacobs, L.L., Winkler, D.A. & Murry, P.A., 1989. Modern mammal origins: evolutionary grades in the Early Cretaceous of North America. Proceedings of the National Academy of Sciences of the United States of America 86, 4992–4995.
- Ji, Q., Luo, Z.-X., Yuan, C.-X., Wible, J.R., Zhang, J.-P. & Georgi, J.A., 2002. The earliest known eutherian mammal. Nature 416, 816–822.
- Kearney, M. & Clark, J.M., 2003. Problems due to missing data in phylogenetic analyses including fossils: a critical review. Journal of Vertebrate Palaeontology 23, 263–274.
- Kermack, D.M., Kermack, K.A. & Mussett, F., 1968. The Welsh pantothere Kuehneotherium praecursoris. Journal of the Linnean Society of London, Zoology 47, 407–423.
- Kielan-Jaworowska, Z. & Cifelli, R.L., 2001. Primitive boreosphenidan mammal (?Deltatheroida) from the Early Cretaceous of Oklahoma. Acta Palaeontologica Polonica 46, 377–391.
- Kielan-Jaworowska, Z., Cifelli, R.L. & Luo, Z., 1998. Alleged Cretaceous placental from down under. Lethaia 31, 267–268.
- Kielan-Jaworowska, Z., Cifelli, R.L. & Luo, Z., 2004. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press, New York, 700 pp.
- Kielan‐Jaworowska, Z. & Dashzeveg, D., 1989. Eutherian mammals from the Early Cretaceous of Mongolia. Zoologica Scripta 18, 347–355.
- King, B. & Beck, R.M., 2020. Tip dating supports novel resolutions of controversial relationships among early mammals. Proceedings of the Royal Society B 287, 1–7.
- Kraus, M.J., 1979. Eupantotheria in Mesozoic Mammals.In The First Two-Thirds of Mammalian History. Lillegraven, J., Kielan-Jaworowska, Z., & Clemens, W.A., eds. University of California Press, Berkeley, pp. 162–171.
- Krause, D.W., Hoffman, S., Hu, Y., Wible, J.R., Rougier, G.W., Kirk, E.C., Groenke, J.R., Rogers, R.R., Rossie, J.B., Schultz, J.A., Evans, A.R., VON Koenigswald, W. & Rahantarisoa, L.J., 2014. Skeleton of Cretaceous mammal from Madagascar reflects long term insularity. Nature 581, 421–427.
- Kühne, W.G., 1949. On a Triconodont tooth of a new pattern from a fissure-filling in South Glamorgan. Proceedings of the Zoological Society of London 119, 345–350.
- Lopatin, A.V. & Averianov, A.O., 2017. The stem placental mammal Prokennalestes from the Early Cretaceous of Mongolia. Paleontological Journal 51, 1293–1374.
- Luo, Z., Ji, Q. & Yuan, C.X., 2007. Convergent dental adaptations in pseudo-tribosphenic and tribosphenic mammals. Nature 450, 93–97.
- Luo, Z.-X., Chen, P., Li, G. & Chen, M., 2007. A new eutriconodont mammal and evolutionary development in early mammals. Nature 446, 288–293.
- Luo, Z., Cifelli, R.L. & Kielan-Jaworowska, Z., 2001. Dual origin of tribosphenic mammals. Nature 409, 53–57.
- Luo, Z.-X., Kielan-Jaworowska, Z. & Cifelli, R.L., 2002. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47, 1–78.
- Luo, Z.-X., Ji, Q., Wible, J.R. & Yuan, C.-X., 2003. An Early Cretaceous tribosphenic mammal and metatherian evolution. Science 302, 1934–1940.
- Luo, Z.-X., Meng, Q.-J., Grossnickle, D.M., Liu, D., Neander, A.I., Zhang, Y.-G. & Ji, Q., 2017. New evidence for mammaliaform ear evolution and feeding adaptation in a Jurassic ecosystem. Nature 548, 326–329.
- Luo, Z., Yuan, C.-X., Meng, Q.-J. & Ji, Q., 2011. A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 476, 442–445.
- Marsh, O.C., 1879. Notice of new Jurassic reptiles. American Journal of Science s3-18, 501–505.
- Martin, T. & Rauhut, O.W.M., 2005. Mandible and dentition of Asfaltomylos patagonicus (Australosphenida, Mammalia) and the evolution of tribosphenic teeth. Journal of Vertebrate Palaeontology 25, 414–425.
- Martin, T., Goin, F.J., Schultz, J.A. & Gelfo, J.N., 2022. Early Late Cretaceous mammals from southern Patagonia, Santa Cruz Province, Argentina. Cretaceous Research 133, 105127–105127.
- Mckenna, M., 1975. Toward a phylogenetic classification of the Mammalia. In Phylogeny of the Primates., Luckett, W.P. & Szalay, F.S. eds. Plenum Publishing, New York. pp. 21–47.
- Mckenna, M. & Bell, S.K., 1997. Classification of Mammals above the Species Level. Columbia University Press, N.Y.
- Meng, J., 2014. Mesozoic mammals of China: implications for phylogeny and early evolution of mammals. National Science Review 1, 521–542.
- O’Leary, M.A., Bloch, J.I., Flynn, J.J., Gaudin, T.J., Giallombardo, A., Giannini, N.P., Goldberg, S.L., Kraatz, B.P., Luo, Z.-X., Meng, J., Ni, X., Novacek, M.J., Perini, F.A., Randall, Z., Rougier, G.W., Sargis, E.J., Silcox, M.T., Simmons, N.B., Spaulding, M., Velazco, P.M., Weksler Wible, M.J.R. & Cirranello, A.L., 2013. The placental mammal ancestor and the post-K-Pg radiation of placentals. Science 332, 662–667.
- Pascual, R., Archer, M., Jaureguizar, E.O., Prado, J.L., Godthelp, H. & Hand, S.J., 1992. First discovery of monotremes in South America. Nature 356, 704–706.
- Patterson, B., 1956. Early Cretaceous mammals and the evolution of mammalian molar teeth. Fieldiana Geology 13, 1–105.
- Pian, R., Archer, M., Hand, S.J., Beck, R.M. & Cody, A., 2016. The upper dentition and relationships of the enigmatic Australian Cretaceous mammal Kollikodon ritchiei. Memoirs of Museum Victoria 74, 97–105.
- Poropat, S.F., Martin, S.K., Tosolini, A.-M.P., Wagstaff, B.E., Bean, L.B., Kear, B.P., Vickers-Rich, P. & Rich, T.H., 2018. Early Cretaceous polar biotas of Victoria, southeastern Australia—an overview of research to date. Alcheringa 42, 157–229.
- Prasad, G.V.R., Parmar, V. & Kumar, D., 2015. A Jurassic australosphenidan mammal from India: Implications for mammalian evolution and distribution in former Gondwanaland. In XII International Symposium on Antarctic Earth Sciences, Goa (India). Abstracts, pp. 461–462.
- Rauhut, O.W.M., Martin, T., Ortiz-Jaureguizar, E. & Puerta, P., 2002. A Jurassic mammal from South America. Nature 416, 165–168.
- Rich, T.H., Flannery, T.F., & Vickers-Rich, P., 1998. Alleged Cretaceous placental from down under: reply. Lethaia 31, 346–348.
- Rich, T.H., Flannery, T.F., Evans, A.R., White, M., Ziegler, T., Maguire, A., Poropat, S., Trusler, P. & Vickers-Rich, P., 2020. Multiple hypotheses about two mammalian upper dentitions from the Early Cretaceous of Australia. Alcheringa 44, 528–536.
- Rich, T.H., Flannery, T.F., Trusler, P., Kool, L., VAN Klaveren, N.A. & Vickers-Rich, P., 2001. A second tribosphenic mammal from the Mesozoic of Australia. Records of the Queen Victoria Museum 110, 1–9.
- Rich, T.H., Lowery, M., Hall, M., Kool, L., White, M. & Vickers-Rich, P., 2022. A new Cretaceous fossil mammal locality from the Bass Coast of southeastern Australia. Alcheringa 46. In press.
- Rich, T.H., Trusler, P., Kool, L., Pickering, D., Evans, A., Siu, K., Maksimenko, A., Kundrat, M., Gostling, N.J., Morton, S. & Vickers-Rich, P., 2020. A third, remarkably small, tribosphenic mammal from the Mesozoic of Australia. In Biological Consequences of Plate Tectonics: New Perspectives on Post-Gondwana and Break-up—a Tribute to Ashok Sahni, Vertebrate Paleobiology and Paleoanthropology. Prasad, G.V. & Patnaik, R. eds. Springer International, Springer, Cham. pp. 67–76.
- Rich, T.H., Vickers-Rich, P., Constantine, A., Flannery, T.F., Kool, L. & VAN Klaveren, N., 1997. A tribosphenic mammal from the Mesozoic of Australia. Science 278, 1438–1442.
- Rich, T.H., Vickers-Rich, P., Constantine, A., Flannery, T.F., Kool, L. & VAN Klaveren, N., 1999. Early Cretaceous mammals from Flat Rocks, Victoria, Australia. Records of the Queen Victoria Museum. 106, 1–34.
- Rich, T.H., Vickers-Rich, P., Flannery, T.F., Pickering, D., Kool, L., Tait, A.M. & Fitzgerald, E.M.G., 2009. A fourth Australian Mesozoic mammal locality. Bulletin of the Museum of Northern Arizona 65, 677–681.
- Rougier, G.W., Martinelli, A.G. & Foraseipi, A.M., 2021. The fossil record of South American Mesozoic mammals and their close relatives. In Mesozoic Mammals from South America and Their Forerunners. Rougier, G.W., Martinelli, A.G., & Foraseipi, A.M. eds, Springer Earth System Science, Springer.
- Rougier, G.W., Martinelli, A.G., Forasiepi A, M. & Novacek, M.J., 2007. New Jurassic mammals from Patagonia, Argentina: a reappraisal of Australosphenidan morphology and interrelationships. American Museum Novitates 3566, 1.
- Sigogneau-Russell, D., 1992. Hypomylos phelizoni nov. gen. nov. sp., une étape précoce de l‘évolution de la molaire tribosphénique (Crétacé basal du Maroc). Geobios 25, 389–393.
- Sigogneau-Russell, D., Hooker, J.J. & Ensom, P.C., 2001. The oldest tribosphenic mammal from Laurasia (Purbeck Limestone Group, Berriasian, Cretaceous, UK) and its bearing on the ‘dual origin’of Tribosphenida. Comptes Rendus de l'Académie des Sciences – Series Iia – Earth and Planetary Science 333, 141–147.
- Simpson, G.G., 1936. Studies of the earliest mammalian dentitions. Dental Cosmos 78, 791–800–940–953.
- Simpson, G.G., 1937. The beginning of the Age of Mammals. Biological Reviews 12, 1–46.
- Slaughter, B.H., 1968. Earliest known marsupials. Science 162, 254–255.
- Vullo, R., Gheerbrant, E., de Muizon, C. & Néraudeau, D., 2009. The oldest modern therian mammal from Europe and its bearing on stem marsupial biogeography. Proceedings of the National Academy of Sciences of the United States of America 106, 19910–19915.
- Wagstaff, B.E., Gallagher, S.J., Hall, W.M., Korasidis, V.A., Rich, T.H., Seegets-Villiers, D.E. & Vickers-Rich, P.A., 2020. Palynological-age determination of Early Cretaceous vertebrate-bearing beds along the south Victorian coast of Australia, with implications for the spore-pollen biostratigraphy of the region. Alcheringa 44, 460–474.
- Wible, J.R., Rougier, G.W., Novacek, M.J., Mckenna, M.C. & Dashzeveg, D.D., 1995. A mammalian petrosal from the Early Cretaceous of Mongolia: implications for the evolution of the ear region and mammaliamorph relationships. American Museum Novitates 3149, 1–19.
- Wible, J.R., Rougier, G.W., Novacek, M.J. & Asher, R.J., 2009. The eutherian mammal Maelestes gobiensis from the Late Cretaceous of Mongolia and the phylogeny of Cretaceous Eutheria. Bulletin of the American Museum of Natural History 2009, 1–123.
- Williamson, T.E., Brusatte, S.L. & Wilson, P.G., 2016. The origin and early evolution of metatherian mammals. The Cretaceous record. ZooKeys 165, 1–76.
- Woodburne, M.O., Rich, T.H. & Springer, M.S., 2003. The evolution of tribospheny and the antiquity of mammalian clades. Molecular Phylogenetics and Evolution 28, 360–385.
- Yuan, C.X., Ji, Q., Meng, Q.J., Tabrum, A.R. & Luo, Z.X., 2013. Earliest evolution of multituberculate mammals revealed by a new Jurassic fossil. Science 341(6147), 779–783.