Abstract
Mangrove habitats are host to many waterbirds around the globe. However, the roles of mangrove habitat structures in association with variation in individuals and their community of waterbirds have not been examined in detail. The objective of this study was to determine the abundance, diversity and evenness of waterbirds in an Avicennia mangrove habitat. The study was conducted using a point count method at Hara Protected Area (HPA) in Iran for 1 year. A total of 59,387 waterbirds observations, belonging to 56 species, were recorded and categorized based on different seasons, where 13,825 (49 spp.), 34,267 (54 spp.), 6939 (46 spp.) and 4356 (35 spp.) observations were recorded in the different seasons of fall, winter, spring and summer respectively. In total, 80.98% of individual observations had occurred during the migratory season. Dromas ardeola (11,329 observation; 19.08%) and Calidris alpina (5261 observation; 8.86%) were the most abundant, while three species including Phalacrocorax nigrogularis (4 observations), Vanellus indicus (2 observations) and Chlidonias leucopterus (2 observations) were the least abundant waders and seabird species. There were significant differences between the number of waders and seabirds observed in fall (t = 1.22, P < 0.05), winter (t = 1.27, P < 0.05), spring (t = − 0.78, P < 0.05) and summer (t = − 1.55, P < 0.05). The results also showed that there was a significant difference between the number of individuals observed in different seasons (F3,232 = 7.4, P < 0.01). The abundance of waterbirds at HPA was highest in winter while the diversity and evenness were highest in spring. The HPA is important because of the presence of exotic species such as Numenius arquata that is listed as near threatened in the red list of the International Union for Conservation of Nature. HPA mangrove forest could be one of the “megadiversity” places in the Middle East. Thus, preventing further losses of the mangrove and reducing the impacts of disturbance on birds and their habitats should be prioritized.
Introduction
The “Hara Forests” is the common name for mangrove forests on the southern coast of Iran (Safa Citation2006), in the Persian Gulf, spreading over several locations between the 25°11′ and 27°52′ parallels (Ghasemi et al. Citation2010). The word “mangrove” has been used to refer either to the constituent plants of tropical intertidal forest communities or the community itself (Ellison Citation2008). Mangroves, which are one of the most disturbed habitats mostly due to intense human activities (Walton et al. Citation2007), are characterized by important concentrations of bird species at both breeding colonies and roosting sites (Colombini et al. Citation1995; Alves et al. Citation1997), and tend to show higher waterbird abundances and species richness due to high nutrient levels.
The Hara Protected Area (HPA) in southern Hormozgan province protects the largest mangrove forests remaining in Iran (Danehkar Citation2001). As a suitable feeding and roosting habitat for a diverse range of waterbirds (Behrouzi-Rad Citation1991), HPA bears the largest number of overwintering migrant waterbirds protected by the Department of Environment (DoE) of Iran in 1975. Danehkar (Citation2006) noted that this location in the East Asian–Australasian flyway of migratory birds contributes to the importance of HPA as staging sites for migrants.
Iran has about 105 Important Bird Areas (IBA) hosting about 500 species (Firouz Citation1974; Behrouzi-Rad Citation2006). Given their geographical and ecological situation, these wetlands are used as overwintering sites for many predatory birds, landbirds, wader and waterfowl populations. Some early reports on wintering birds in Iran have been provided in the Iranian bird species lists in these areas (Firouz Citation2005; Scott and Adhami Citation2006; Lepage Citation2010). Research on wintering birds has also focused mainly on census counts by the Environmental Department.
The mangroves of Hormozgan province are historically a suitable feeding and roosting habitat for a diverse range of waterbirds among the IBA in Iran (Danehkar Citation2001). Despite the long-term presence of mangrove forests in Iranian coasts of Persian Gulf, there is a lack of planned management for these rare ecosystems (Danehkar Citation2001). At present, there is no basic information on waterbird assemblage annual structure not only in mangroves of Hormozgan province but also all over Iran. This is a cause for concern. The main objective of this paper was to analyse the seasonal variability in the structure of waterbird assemblages at HPA mangrove site.
Material and methods
Study area
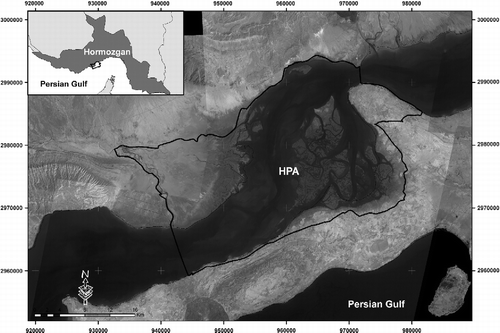
HPA or “Khouran Straits” is located in southern Persian Gulf between the region of the Mehran river and Kol river deltas and the island of Qeshm, within quadrant of 26°23′–26°59 ′ N and 55°32′–55°48′ E (Figure ). The annual mean, minimum and maximum temperatures were 27.6, 2 and 48 °C in a 30-year period (1975–2005) respectively. The mean annual rainfall is about 80.3 mm that mainly occurs in winter. The mean monthly relative humidity is 83.4% and the range of high tide is 4.33 m from the Port of Shahid Rajaee, nearest to the study site. Avicennia marina forms a pure mangrove stand in HPA.
Figure 1 Hara Protected Area, Hormozgan, Iran.
Survey design
The study was conducted biweekly over four seasons including fall (22 September–21 December), winter (21 December–20 March), spring (21 March–20 June) and summer (21 June–21 September) for a period of 1 year starting from 21 September 2008, using the point count sampling method. The fall and winter are migratory seasons while spring and summer are non-migratory seasons.
Mangrove site was divided by many intertidal channels. A total of three transects, with 1000–3000 m length, were established on the map randomly within three main channels in HPA. Transects were run parallel to creek at the predecided locations distributed in HPA. A total of 35 count point were established within transect 1, 30 points in transect 2 and 32 points in transect 3, randomly. The points were selected at least 300 m apart from each other based on the characteristics of the habitat and availability of time and facilities (Figure ).
Figure 2 Randomization of sampling points among transects.
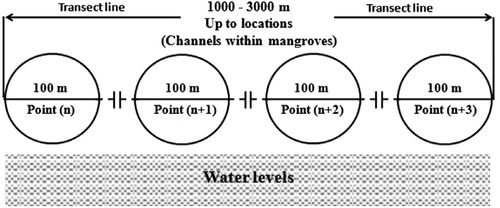
Some points inside the dense mangroves or deep water could not be used by the waterbirds and were not available for the researchers. To this end, the total numbers of points was reduced to 16 points. This method has been also reported to be suitable for study in wetland areas (Bokach Citation2005; Luo et al. Citation2010). A small outboard engine boat was used to transport us to each point that was selected to count the waterbirds. In some areas the points were extremely soft, which made it difficult for the researchers to move around with the telescope even over short distances; thus, another counting method was used. To this end, the strategy was to approach the coast with the boat as close as possible and then to start counting the birds while sitting on the boat. The methodology was followed as described by Van der Winden et al. (Citation2007).
The survey was conducted from 08:00 to 14:00 hours during low tides. Low tides expose the mudflats (the preferred feeding area for most waders) and provide an opportunity for waders to forage and easy access to conduct bird surveys. Each point was surveyed two consecutive times at monthly intervals (n = 2 × 12 = 24 times) to achieve reliable diversity estimates because the replication of point count stations increases precision and provides reliable results.
The number of birds was recorded for 10 minutes within 100 m radius at 360° for each point. A 10-minute count allows sampling of high number of individuals with minimum effort and disturbance (Lee and Marsden Citation2008; Zakaria et al. Citation2009). A pair of binoculars (10x) and spotting scopes (30x and 60x) were used for detecting and identifying birds. Species and number of individuals detected by sight were recorded in each point count survey.
To avoid overlapping or double counting of the same individual birds, some steps were taken as follows: (1) to follow the movements of the birds, all point stations were established 300 m apart from each other. (2) Only visible bird species within 100 m radius at 360° for each point were counted by fully trained observers. All observers had >80 hours of experience waterbirds estimating at the study areas. (3) The movement and number of disturbed birds were tracked, and the birds that were recorded at overlapping places were counted only once.
Data analysis
The relative abundance (%) of bird species was determined using the expression: n/N × 100 (where n is the number of the recorded bird species and N the total number of observations recorded) (Zakaria et al. Citation2009). Each observation represents one bird. All statistical tests were conducted by using SPSS 16 software.
The similarity percentages (SIMPER) and other ecological structures of the communities were tested using the community analysis package software (CAP, version 4.0) by Seaby and Henderson (Citation2007), to compare the conventional diversity indices of waterbird's community. As noted by Josens et al. (Citation2009), SIMPER uses the standard deviation of the Bray–Curtis dissimilarity matrix, attributed to a species, for all species pairs and compares it with average contribution of a species to the dissimilarity. Furthermore, when numbers of habitats or seasons are to be compared, the similarity measures can form the basis of cluster analysis, which aims at identifying groups of sites, stations or seasons that are similar in their species composition. Thus, a cluster analysis (using Bray–Curtis distances on standardized data) was performed to better visualize species composition across seasons.
Results
On the basis of the observations by the point count method in the 1-year period starting from 22 September 2008, a total of 59,387 waterbird observations, belonging to 56 species, representing 14 families and 4 orders, were recorded at HPA. Out of the total number, 1893 sandpipers, 1098 gulls and 517 terns were identified (Table ). Four orders of waterbirds including Charadriiformes (51,176 observations, 86.17%), Ciconiformes (4604 observations, 7.75%), Pelecaniformes (3495 observations, 5.89%) and Anseriformes (12 observations, 0.02%) were recorded at HPA. Furthermore, 56 bird species observations were recorded where Charadriiformes (44 spp., 78.57%) and Ciconiformes (8 spp., 14.29%) were the most abundant. Only one species of Anseriformes was observed (1.78%) during the study period. Dromas ardeola Paykull, 1805 (11,329 observations; 19.08%), Calidris alpina Linnaeus, 1758 (5261 observations; 8.86%) and Numenius arquata Linnaeus, 1758 (3839 observations; 6.46%) were the most abundant, while Phalacrocorax nigrogularis Ogilvie-Grant and Forbes, 1899, Vanellus indicus Boddaert, 1783 and Chlidonias leucopterus Temminck, 1815 (4, 2 and 2 observations respectively) were the least abundant bird species. The total observations for these three species did not exceed 0.01% of the total observations.
Table 1 Relative abundance of waterbird observations recorded at HPA, Iran.
The observations were categorized under different seasons, where a total of 49 (13,825 observations; 23.28%), 54 (34,267 observations; 57.70%), 46 (6939 observations; 11.68%) and 35 (4356 observations; 7.33%) species were recorded for fall, winter, spring and summer respectively. A total of 80.98% of individual observations occurred during the migratory seasons.
In fall, Chlidonias leucopterus, Tringa nebularia Gunnerus, 1767, Calidris ferruginea Pontoppidan, 1763 and Larus canus Linnaeus, 1758 were the most abundant (with 100%, 61.32%, 61.11% and 60% observations respectively), Sterna repressa Hartert, 1916, Ardeola grayii Sykes, 1832 and Sterna saundersi Hume, 1877 were the least abundant bird species (4.43%, 3.23% and 1.42% observations respectively). Moreover, in this season, Phalacrocorax nigrogularis, Ardea purpurea Linnaeus, 1766, Nycticorax nycticorax Linnaeus, 1758, Anas penelope Linnaeus, 1758, Vanellus indicus, Charadrius dubius Scopoli, 1786, Limosa lapponica Linnaeus, 1758 and Sterna sandvicensis Latham, 1787 were absent. However, in winter, all the 54 species, except for Vanellus indicus and Chlidonias leucopterus were observed. As the results of the observations showed, Phalacrocorax nigrogularis, Anas penelope, Sterna sandvicensis (100% each), Sterna saundersi (94.33%) and Calidris alba Pallas, 1764 (94.03%) were dominant. Vanellus indicus, Nycticorax nycticorax and Charadrius dubius were of dominant observations in spring, accounting for 100%, 92.86% and 84.93% of the total bird numbers respectively. Pelecanus crispus Bruch, 1832 (0.86%), Charadrius alexandrinus Linnaeus, 1758 (0.74%) and Limosa lapponica Linnaeus, 1758 (0.55%) were the least abundant waterbirds during spring visits. In addition, Phalacrocorax nigrogularis, Anas penelope, Tringa stagnatilis Bechstein, 1803, Calidris alba, Calidris minuta Leisler, 1812, Calidris ferruginea, Philomachus pugnax Linnaeus, 1758, Larus canus, Sterna sandvicensis and Chlidonias leucopterus were totally absent in spring. Thirty-five species were observed during summer in HPA, where Sterna repressa, Sterna anaethetus Scopoli, 1786 and Sterna bengalensis Lesson, 1831 were the dominant species and were the most abundant of the bird community in summer, accounting for 69.79%, 67.46% and 57.14% of the total bird numbers respectively.
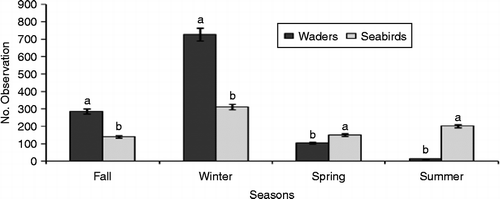
The mean values of waders during fall, winter, spring and summer were estimated at 285.9 ± 79.4, 726.3 ± 222.4, 104.6 ± 34.9 and 13.5 ± 4.2 observations per season respectively, whereas the mean values of seabird observations for the same seasons were estimated at 140.8 ± 47.9, 312.2 ± 98.2, 150.5 ± 45.1 and 201.6 ± 121.5 observations per season respectively. Comparing mean values of waders and seabirds among seasons, t-test results indicated that there was a significant difference between the number of waders and seabirds observed in fall (t = 1.22, P < 0.05), winter (t = 1.27, P < 0.05), spring (t = − 0.78, P < 0.05) and summer (t = − 1.55, P < 0.05) (Figure ). The results also showed that there was a significant difference between the number of individuals observed in different seasons (F3,232 = 7.4, P < 0.01). In addition, the post hoc multiple comparisons by Tukey's honestly significant difference test clearly indicated that more individuals were recorded in winter than in fall (P < 0.05), spring (P < 0.05) and summer (P < 0.01).
Figure 3 Comparing mean values of waders and seabirds among seasons at HPA. Note: Different letters represent statistical differences (P < 0.05).
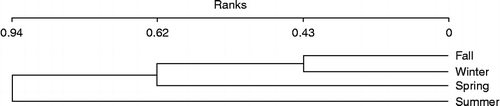
The results from a cluster analysis of waterbird assemblages across seasonal changes illustrated that waterbirds of fall and winter seasons fell into group with 0.43 similarity rank and spring season represented 0.62 similarity ranks with sum of fall and winter, whereas waterbird assemblages in the summer have 0.94 similarity with total groups of three other seasons (Figure ). It means that birds in summer are common with other seasons and new species were not seen.
Figure 4 Grouping of waterbird assemblages as defined by cluster analysis across seasonal changes at HPA, Iran.
A SIMPER analysis was performed to test the contribution of waterbirds at the mangrove habitat. The results indicated that total average similarity of waterbird species in HPA was 32.90%. The HPA was dominated by Dromas ardeola, Larus genei Brème, 1839 and Sterna anaethetus (SIMPER, percentage of contribution to similarity of 17.3%, 10.75% and 9.14% respectively). Seven species, namely Dromas ardeola, Larus genei, Sterna anaethetus, Larus heuglini Bree, 1876, Numenius arquata, Calidris alpina, and Larus ichthyaetus Pallas, 1773, represented more than 55% of the observations at HPA. Ninety per cent of the total population of birds were also represented by twenty species of birds only (Table ).
Table 2 Contribution of waterbird species similarity at HPA mangrove habitat (SIMPER analysis).
Comparison of seasonal richness, diversity and evenness among seasons indicated that the abundance of waterbirds at HPA was highest in winter than that in other seasons (54 vs. 49, 46 and 33 in fall, spring and summer respectively). In this habitat, the diversity was also highest in spring than in other seasons (Shannon-Wiener's D were 3.05, 3.01, 3.1 and 1.94 in the fall, winter, spring and summer respectively and Simpson's D were 13.98, 11.88, 14.5 and 3.25 in the same seasons respectively. Moreover, the evenness McIntosh's E value was 0.85, 0.82, 0.86 and 0.53 and Pielou J was estimated to be 0.74, 0.73, 0.76 and 0.47 in the same seasons respectively).
Discussion
Mangroves are situated within the tidal limits on alluvial flats in the delta (Benfield Citation2002), on sheltered muddy coastal areas (Hogarth Citation1999; FAO Citation2004), in estuaries and on marine shorelines (Benfield Citation2002), where other plants cannot grow (Maguire et al. Citation2000). Several authors have reported contradictory findings about number of waterbirds in mangrove forests. For instance, Nisbet (Citation1968) reported 135 bird species in the mangrove habitats of Peninsular Malaysia. Likewise, Altenburg and van Spanje (Citation1989) counted 125 species in Guinea-Bissau, West Africa. Murphy and Sigurdsson (Citation1990) identified sandpipers, plovers, herons and egrets in Singapore mangrove habitats. Hussain and Acharya (Citation1994) reported 315 species of birds from the Sunderbans, Bangladesh. Noske (Citation1996) identified 104 species in mangroves of north-western Australia, and Alves et al. (Citation1997) counted 32 bird species (2 marine species, 18 terrestrial species and 12 waterfowl) in the mangroves of Jequiaman, Brazil. Zou et al. (Citation2008) reported 22 shorebird species on mangrove wetland surrounding Leizhou Peninsula, China. In several reports, the importance of mangrove areas for waterbirds, especially some threatened species, has been pointed out (Yorke Citation1984; Brown et al. Citation2001; Gawlik Citation2002; Kasawani et al. Citation2007; Fernández et al. Citation2008; Fujita et al. Citation2009). However, it is extremely hard to find the comparison of waterbird diversity among the mangroves, because of differences in the link to the nutrient cycles (De Graaf et al. Citation1985) or physical structure of the environment (Josens et al. Citation2009). Apart from structural complexity, the patterns of resource availability have an effect on diversity of waterbird species in mangroves (Orians Citation1969). Nonetheless, far too little attention has been paid to the mangroves of Iran (Ghasemi and Zakaria Citation2011). At present, DoE of Iran is counting the wintering bird communities every year, but there have been no controlled studies that compare the differences in reference to seasons.
This study was designed to compare the seasonal waterbirds community composition, richness and abundance at A. marina mangrove habitat in the Hormozgan province, Iran. Low densities of waterfowl were expected in all seasons based on water salinity. Moreover, the highest observations were expected to record in the migratory season.
The season of winter was expected to have a higher ratio of waterbird composition, richness and abundance due to its microclimate and microhabitat structures (Andrei et al. Citation2009), but the results of observation in HPA indicated that waterbird diversity and evenness were highest in spring than in other seasons. Although it is unclear what processes were involved, one likely explanation is that 2009 was an unusually cool spring, especially in the first months, so many migrants delayed their emigration time that may have positively influenced the presence of wintering waterbird populations till early spring.
Furthermore, it is well known that environmental quality may affect the equilibrium of wildlife population size (Varasteh Moradi and Zakaria Citation2009), richness and abundance of species (Zou et al. Citation2008); therefore, the waterbirds would respond quickly to any change in their mangrove habitat.
More than 80% of waders at both mangrove habitats in the study sites were recorded on intertidal flats, which proved to be the most important habitat for waders. This means that the locations of proposed mangrove plantations must be considered with respect to conservation of intertidal flats (Zou et al. Citation2008). Field observations showed that human presence at the mangroves in study sites was frequent and often intense, and appeared to be an important factor affecting waterbirds' distribution, especially during the breeding season for six species, namely Casmerodius albus Linnaeus, 1758, Ardeola grayii, Platalea leucorodia and Egretta gularis Bosc, 1792, in spring. Moreover, many mangrove forests have been destroyed and fragmented due to economic development. Therefore, to help secure the conservation of waterbirds, the DoE of Hormozgan province must strive to minimize human (and sometimes governmental conflict) activities that disturb waders and seabirds' feeding/shelter areas to enhance the birds' fitness.
Abundance of waterbirds at the study sites fluctuated seasonally depending on the interaction between species and habitat. Thus, they can be used as a key tool for monitoring the status of changes in mangrove degradation in this area. Prior studies have noted the importance of environmental condition on feeding activities (Liang et al. Citation2002). For example, water salinity may change waterbirds' surface-feeding (Zou et al. Citation2008) because different species of aquatic invertebrates demonstrate differences in salinity tolerance (Liang et al. Citation2002). The surface water of mangrove habitat is also affected by soil character, which is affected by the bedrock, glacial activity and the plants growing on the soil (Badola and Hussain Citation2005).
This study supported previous researches, which have noted the importance of mangrove wetlands for diverse wintering migrant birds (Colombini et al. Citation1995; Alves et al. Citation1997). The highest number of waders was present during migratory seasons (fall and winter). One likely explanation is that most of the waders are winter migrants and that they form important links in the mangrove food chain during migratory season. Thus, it is very clear that wading birds such as Dromas ardeola, Charadrius alexandrinus, Charadrius mongolus Pallas, 1776, Limosa lapponica, Numenius phaeopus Linnaeus, 1758, Numenius arquata, Tringa tetanus Linnaeus, 1758, Tringa cinerea Güldenstädt, 1775, Calidris alpina, and Limicola falcinellus Pontoppidan, 1763 were more abundant than seabirds. On the other hand, the population trends of birds indicated that during non-migratory seasons (spring and summer), seabirds such as gulls and terns as well as herons were the dominant groups. The reason for this is not clear but it may have something to do with resident status of gulls or summer migration of terns in Hormozgan province. Four species of terns are known as summer migrants and breeding birds, which can be observed in this season in Persian Gulf islands such as Sheedvar island near HPA. Also, the numerous observations of Pelecanus crispus, Phalacrocorax nigrogularis and Tringa cinerea use HPA. Waterbird reproduction along the HPA mangrove forest was between 24 March 2009 and 13 May 2009. Six species, namely Egretta gularis Bosc, 1792, Egretta alba Linnaeus, 1758, Ardeola grayii, Platalea leucorodia, Esacus recurvirostris Curvier, 1829, and Charadrius alexandrinus, breed in A. marina habitats or sandy ridges around mangroves in HPA. Previous study also showed that HPA hosts one of the largest breeding colonies of Cosmerodius albus that resides at mangroves in Persian Gulf and Oman Sea limits (Evans Citation1994). The findings related to six breeding species including four species of Ciconiformes indicated that HPA is probably a suitable habitat for these waterbirds due to its availability of best nursery habitat, accessibility of food, and density of gastropods and other organisms, and availability of shelter. Some of these waterbird species are listed as vulnerable or near-threatened birds in the endangered red list of the International Union for Conservation of Nature (IUCN) (IUCN Citation2008; Table ). HPA is the critical habitat for Numenius arquata that is listed as near threatened in the endangered red list of IUCN (IUCN Citation2008). This is largely because of infrastructure development and disturbance at its nesting colonies, possibly exacerbated by marine oil pollution (Birdlife International Citation2010). Therefore, the importance of the HPA becomes much more apparent by looking at the numbers of important and exotic species. As a result, HPA mangrove forest area could be considered one of the “megadiversity” places not only in Iran but also in the Middle East.
Table 3 World population estimates of selected species compared with the total number of individuals recorded in HPA during the period of study.
The two main hypotheses proposed to explain why HPA may attract waterbird species among other animals are:
(a) The predator refuge hypothesis, which states that these species can avoid predators by migrating into HPA mangroves ecosystem when the trees are inundated by tides (Laegdsgaard and Johnson Citation2001). This could be because the structural complexity of mangroves above ground may impede predator movement and vision and thereby reduce their efficiency. Similarly, high turbidity levels within HPA mangroves may also decrease the foraging efficiency of predators (Abrahams and Kattenfeld Citation1997). | |||||
(b) The feeding hypothesis, which suggests a greater abundance of food due to the claimed high mangrove productivity and the presence of benthic fauna, and hence, greater abundance and diversity of aquatic tides (Laegdsgaard and Johnson Citation2001). | |||||
Ultimately, it is expected that the findings of this study contribute to an in-depth knowledge of mangrove waterbirds and that emphasis is placed on the importance of conservation of habitat at HPA to ensure the maintenance of a biologically diverse ecosystem. It should not be neglected that Iran has a long and well-established history of business related to mangrove (Ghasemi et al. Citation2010). The relevant agencies in Iran have been either slow or reluctant to recognize the value of the mangrove existence. There must be a willingness to change old perceptions of this natural resource and create a readiness to accept the new and different requirements offered by mangrove resources. The new management challenges of the mangrove reserve should openly be accepted. Moreover, the occurrence of globally threatened species such as pelicans suggests the urgent need to recognize and protect this site. Regular wader and seabird population counts on a monthly basis with particular focus on the fall and winter months are necessary. It would be interesting to incorporate methodological proposals on long-term survey designs of HPA waterbird community and abundance, as well as proposals for clear management actions of the HPA mangrove habitats, including threatened waterbird colonies and foraging grounds. Public awareness is the best instrument to conservation and to change human attitudes as well as behaviour patterns towards mangrove forests (Ghasemi et al. Citation2010). Environmental NGOs have long recognized that the public can be a powerful ally for them and a formidable foe (Dahdouh-Guebas et al. Citation2004) for those who harm the environment.
Acknowledgements
This work is part of a project funded by the Department of Environment (DoE) in Hormozgan Province. Thanks to Dr Dehghani in the Islamic Azad University, Bandar Abbas Branch, who provided us with all the necessary facilities and great thanks to Mr Meisam Ghasemi for his field assistance.
References
- Abrahams , M and Kattenfeld , M. 1997 . The role of turbidity as a constraint on predator-prey interactions in aquatic environments . Behavioral Ecology and Sociobiology , 40 ( 3 ) : 169 – 174 .
- Altenburg , W and van Spanje , T. 1989 . Utilization of mangroves by birds in Guinea-Bissau . Ardea , 77 : 57 – 74 .
- Alves , VS , Soares , AA and Ribeiro , AB. 1997 . “ Birds of the Jequia mangrove system, Ilha do Governador, Baía de Guanabara, and Rio de Janeiro, Brazil. p. 163–170 ” . In Mangrove ecosystem studies in Latin America and Africa , Edited by: Kjerfve , B , Lacerda , LD and Diop , S . 349 Paris : UNESCO/ISME/USDA .
- Andrei , A , Smith , L , Haukos , D , Surles , J and Johnson , W. 2009 . Foraging ecology of migrant shorebirds in Saline Lakes of the Southern Great Plains . Waterbirds , 32 ( 1 ) : 138 – 148 .
- Badola , R and Hussain , SA. 2005 . Valuing ecosystem functions: an empirical study on the storm protection functions of Bhitarkanika mangrove ecosystem, India . Environmental Conservation , 32 ( 1 ) : 85 – 92 .
- Behrouzi-Rad , B. 1991 . “ Protected areas with mangrove habitat ” . In Draft Report World Conservation Centre , Edited by: Fisher , P and Spalding , MD . 49 – 50 . Cambridge (UK) : World Conservation Centre .
- Behrouzi-Rad , B. 2006 . Avifauna of Gori Gol, East Azerbaijan Province, Northwest Iran . Podoces , 1 ( 1–2 ) : 53 – 60 .
- Benfield , S. 2002 . An assessment of mangrove cover and forest structure in Punta Mala Bay, Panama city, Panama by means of field study and GIS analysis of aerial photographs [Master's Thesis] , [Edinburgh (Scotland)] : Heriot Watt University .
- BirdLife International. 2010. The birdLife checklist of the birds of the world with conservation status and taxonomic sources [Internet]. [Retrieved 18 February 2010]. Available from http://www.birdlife.org/datazone/search/species_search.html
- Bokach , M . 2005 . The relative influences of predation and pray availability on ardeid breeding colony site selection [Master's Thesis] , 48 [Gainesville (FL)] : University of Florida .
- Brown , S , Hickey , C , Harrington , B and Gill , R . 2001 . The United States shorebird conservation plan , 2nd ed. , 61 Manomet (MA) : Manomet Center for Conservation Sciences .
- Colombini , I , Berti , R , Ercolini , A , Nocita , A and Chelazzi , L. 1995 . Environmental factors influencing the zonation and activity patterns of a population of Periophthalmus sobrinus Eggert in a Kenyan mangrove . Journal of Experimental Marine Biology and Ecology , 190 ( 1 ) : 135 – 149 .
- Dahdouh-Guebas , F , Koedam , N , Godefroid , S , Kairo , JG , Sivcev , V , Verheyden , A , Bosire , JO , Dubois , T and Obade , PT. 2004 . “ Mangroves, p. 61–62 ” . In Scientific tools for biodiversity conservation: monitoring, modelling and experiments , Proceedings of the 5th meeting of the European Platform for Biodiversity Research Strategy; 2001 Dec 2–4; Belgian EU Presidency Edited by: Segers , H , Branquart , E , Caudron , A and Tack , J . 225 Brussels : Belgian Biodiversity Platform .
- Danehkar A. 2001. Intraction of Avicennia marina trees and related animals (underscoring of gastropoda fauna) in mangrove forest of Qeshm and Khamir region (Harra Biospheric Reserve) [Ph.D. Thesis in Forestry]. [Tehran (Iran)]: Faculty of Natural Resources, University of Tarbiat Modares. 131 p
- Danehkar , A. 2006 . Extension and management plan of mangrove forest in Hormozgan province , Bandar Abbas (Iran) : Natural Resource Dept. of Hormozgan .
- De Graaf , R , Tilghman , N and Anderson , S. 1985 . Foraging guilds of North American birds . Environmental Management , 9 ( 6 ) : 493 – 536 .
- Ellison , JC. 2008 . Long-term retrospection on mangrove development using sediment cores and pollen analysis: a review . Aquatic Botany , 89 ( 2 ) : 93 – 104 .
- Evans , MI . 1994 . Important Bird Areas in the Middle East , 410 Cambridge (UK) : Birdlife International .
- FAO . 2004 . The state of world fisheries and aquaculture , 155 Rome (Italy) : Food and Agriculture Organization of the United Nations .
- Fernández , G , Buchanan , J , Gill , R. Jr , Lanctot , R and Warnock , N . 2008 . Conservation plan for Dunlin with breeding populations in North America (Calidris alpina arcticola, C.a. pacifica) , Version 1.0. Manomet (MA) : Manomet Center for Conservation Sciences .
- Firouz , E . 1974 . Environment Iran, natural society for the conservation of natural resources and human environment , 51 Tehran (Iran) : Department of Environment .
- Firouz , E . 2005 . The complete fauna of Iran , 322 New York : I.B. Tauris and Co Ltd .
- Fujita , G , Totsu , K , Shibata , E , Matsuoka , Y , Morita , H , Kitamura , W , Kuramoto , N , Masuda , N and Higuchi , H. 2009 . Habitat management of little terns in Japan's highly developed landscape . Biological Conservation , 142 ( 9 ) : 1891 – 1898 .
- Gawlik , DE. 2002 . The effects of prey availability on the numerical response of wading birds . Ecological Monographs , 73 : 329 – 346 .
- Ghasemi , S , Zakaria , M , Hazandy , AH , Yusof , E , Danehkar , A and Rajpar , MN. 2010 . A review of mangrove value and conservation strategy by local communities in Hormozgan province, Iran . Journal of American Science , 6 ( 10 ) : 329 – 338 .
- Ghasemi , S and Zakaria , M. 2011 . Relative abundance and diversity of waterbirds in Rhizophora Mangrove Forest in Iran . Asia Life Science , 20 ( 1 ) : 210 – 238 .
- Hogarth , PJ . 1999 . The biology of mangroves , 229 Oxford (UK) : Oxford University Press .
- Hussain , Z and Acharya , G . 1994 . Mangroves of the Sundarbans , 257 Bangladesh : IUCN-The World Conservation Union . Vol. 2
- [IUCN] International Union for Conservation of Nature. 2008. The IUCN red list of threatened species. [Retrieved 8 February 2008]. Available from http://www.iucnredlist.org
- Josens , M , Haydee , E and Favero , M. 2009 . Seasonal variability of waterbird assemblages in relationship to habitat characteristics in a Pampas wetland . Waterbirds , 32 ( 4 ) : 523 – 530 .
- Kasawani , I , Kamaruzaman , J and Nurun-Nadhirah , MI. 2007 . Biological diversity assessment of Tok Bali mangrove forest, Kelantan, Malaysia . WSEAS Transactions on Environment and Development , 3 ( 2 ) : 37 – 44 .
- Laegdsgaard , P and Johnson , C. 2001 . Why do juvenile fish utilise mangrove habitats? . Experimental Marine Biology and Ecology , 257 : 229 – 253 .
- Lee , DC and Marsden , SJ . 2008 . Adjusting count period strategies to improve the accuracy of forest bird abundance estimates from point transect distance sampling surveys . Ibis , 150 ( 2 ) : 315 – 325 .
- Lepage D. 2010. Checklist of birds of Iran. [Retrieved 2 March 2010]. Available from http://avibase.bsc-eoc.org/checklist.jsp?lang = ENand region = irand list = clements (http://avibase.bsc-eoc.org/checklist.jsp?lang=enandregion=irandlist=clements)
- Liang , SH , Shieh , BS and Fu , YS. 2002 . A structural equation model for physiochemical variable of water, benthic invertebrates, and feeding activity of waterbirds in the Sitsao wetlands of Southern Taiwan . Zoology Study , 41 : 441 – 451 .
- Luo , Z , Sun , O and Xu , H. 2010 . A comparison of species composition and stand structure between planted and natural mangrove forests in Shenzhen Bay, South China . Journal of Plant Ecology , 3 ( 3 ) : 165 – 174 .
- Maguire , T , Saenger , P , Baverstock , P and Henry , R. 2000 . Microsatellite analysis of genetic structure in the mangrove species Avicennia marina (Forsk.) Vierh. (Avicenniaceae) . Molecular Ecology , 9 ( 11 ) : 1853 – 1862 .
- Murphy D, Sigurdsson J. 1990. Birds, mangroves and man: prospects and promise of the new Sungei Buloh Bird Reserve. In: Chou LM and Ng PKL, editors. Essays in Zoology. Singapore: Singapore University Press. p. 233–243
- Nisbet , ICT. 1968 . The utilization of mangroves by Malayan birds . Ibis , 110 ( 3 ) : 348 – 352 .
- Noske , R. 1996 . Abundance, zonation and foraging ecology of birds in mangroves of Darwin Harbour, Northern Territory . Wildlife Research , 23 ( 4 ) : 443 – 474 .
- Orians , GH. 1969 . The number of bird species in some tropical forests . Ecology , 50 ( 5 ) : 793 – 801 .
- Safa , H. 2006 . A survay of Mangrove Forest structure at Tiab and Kolahi (In Persian) [Thesis] , 90 [Bandar Abbas (Iran)] : Islamic Azad University, Branch of Bandar Abbas .
- Scott , D and Adhami , A. 2006 . An updated checklist of the birds of Iran . Podoces , 1 ( 1/2 ) : 1 – 16 .
- Seaby , R and Henderson , P . 2007 . Community analysis (package 4.0). Searching for structure in community data , Lymington (England) : PISCES Conservation Ltd .
- Van der Winden J, Siaka A, Dirksen S, Poot M. 2007. Waterbirds in coastal wetlands of Sierre Leone, January-February 2005. WIWO-report no. 84. Beek-Ubbergen (The Netherlands): Foundation WIWO. 98 p
- Varasteh Moradi , H and Zakaria , M . 2009 . Insectivorous bird community change along an edge-interior gradient in an isolated tropical rainforest in peninsular Malaysia . Malaysian Nature Journal , 61 ( 1 ) : 48 – 66 .
- Walton , ME , Vay , LL , Lebata , JH , Binas , J and Primavera , JH. 2007 . Assessment of the effectiveness of mangrove rehabilitation using exploited and non-exploited indicator species . Biological Conservation , 138 ( 1–2 ) : 180 – 188 .
- Yorke , CD. 1984 . Avian community structure in two modified Malaysian habitats . Biological Conservation , 29 ( 4 ) : 345 – 362 .
- Zakaria , M , Rajpar , MN and Sajap , SA. 2009 . Species diversity and feeding guilds of birds in Paya Indah Wetland Reserve, Peninsular Malaysia . International Journal of Zoological Research , 5 ( 3 ) : 86 – 100 .
- Zou , F , Zhang , H , Dahmer , T , Yang , Q , Cai , J , Zhang , W and Liang , C. 2008 . The effects of benthos and wetland area on shorebird abundance and species richness in coastal mangrove wetlands of Leizhou Peninsula, China . Forest Ecology and Management , 255 : 3813 – 3818 .