
Abstract
The oribatid mite genus Sacculogalumna (Oribatida: Galumnidae) is recorded in the Philippines for the first time. A new species, S. samarensis n. sp., is described from secondary forest litter in the Western and Eastern Samar Provinces in Samar Island. It differs from S. saccularis and S. suiyangensis in having pointed rostrum, setiform bothridial setae, reticulate pteromorphs, long and thickened adanal setae ad1 and ad2 and one seta on anterior edges of genital plates, by the location of lamellar setae between lamellar lines, and the absence of postanal porose area, and by the medially interrupted dorsosejugal furrow. The inclusion of the new species in Sacculogalumna is proved, and an identification key to known species of the genus is given.
http://zoobank.org/urn:lsid:zoobank.org:pub:4D40D41A-EF5C-423D-AAB3-857C16B10BE8
Introduction
This work is a part of our continuing study of the Philippine oribatid mites (Acari: Oribatida) of the superfamily Galumnoidea (Ermilov et al. Citation2014a, 2014b; Ermilov and Corpuz-Raros Citation2015a, Citation2015b, Citation2015c). During taxonomic studies of new materials from the collections from the Museum of Natural History (University of the Philippines Los Baños), we discovered a new species of Sacculogalumna Engelbrecht, Citation1973 (Galumnidae). It is the first record of the genus in the Philippines. The main goal of the paper is to describe and illustrate this species under the name S. samarensis n. sp. Also, an identification key to known species of Sacculogalumna is given.
Sacculogalumna was proposed by Engelbrecht (Citation1973) with Sacculogalumna saccularis Engelbrecht Citation1973 from South Africa as type species. Later, Liang et al. (Citation2012) described a second species of the genus, S. suiyangensis Liang, Yang & Huang, Citation2012 from southern China.
Material and methods
The collection locality and habitat for new species are given in the “Material examined” section.
Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. The body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the ventral plate. Notogastral width refers to the maximum width in dorsal aspect. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence: trochanter–femur–genu–tibia–tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence: genu–tibia–tarsus. Morphological terminology used in this paper follows that of F. Grandjean: see Travé and Vachon (Citation1975) for general references, Norton (Citation1977) for leg setal nomenclature, and Norton and Behan-Pelletier (Citation2009), for overview. Drawings were made with a camera lucida using a Carl Zeiss transmission light microscope “Axioskop-2 Plus”.
Sacculogalumna samarensis n. sp.
(Figures , , , )
Figure 1–2. Sacculogalumna samarensis n. sp. Male: 1, dorsal view; 2 – anterior part of prodorsum, frontal view. S
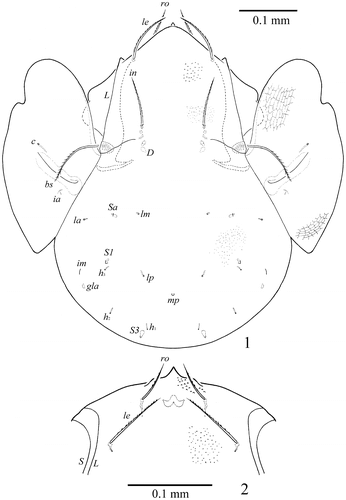
Figure 3–4. S. samarensis n. sp. Male: 3, ventral view (gnathosoma and legs not illustrated); Female: 4, ano-adanal region.
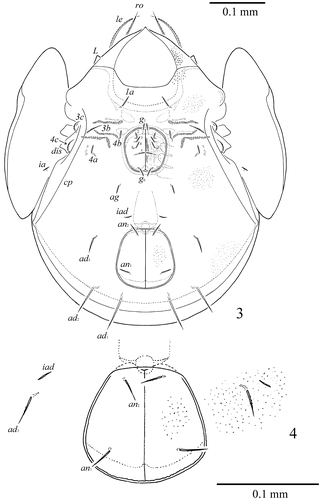
Figure 5–6. S. samarensis n. sp. Male: 5, lateral view (gnathosoma and legs not illustrated); 6, posterior view (legs not illustrated).
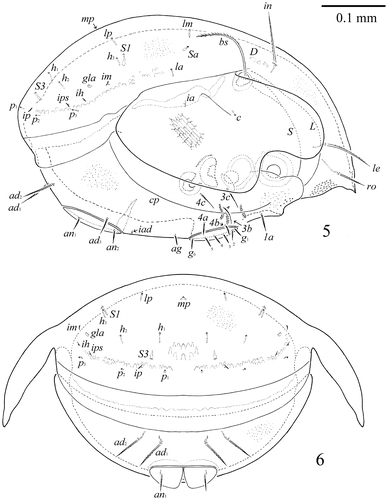
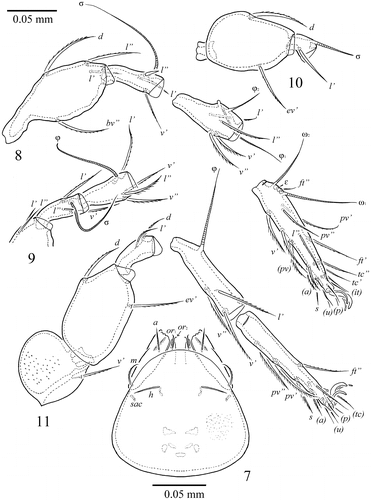
Figure 7–11. S. samarensis n. sp. Male: 7, subcapitulum, ventral view; 8, leg I, without trochanter, right, antiaxial view; 9, tibia, genu and anterior part of femur of leg II, right, antiaxial view; 10, genu and femur of leg III, left, antiaxial view; 11, leg IV, left, antiaxial view.
Diagnosis
Body size: 531–614 × 415–448. Body surface heavily granulate, anterior part of prodorsum and often antero-lateral parts of ventral plate tuberculate. Pteromorphs reticulate. Rostrum pointed. Lamellar and sublamellar lines parallel. Rostral, lamellar and interlamellar setae long, setiform, barbed. Bothridial setae setiform, ciliate. Dorsosejugal furrow interrupted medially. Notogastral setae represented by alveoli. Three pairs of saccules, Sa located between la and lm. Median pore absent in female and present in males. Epimeral setae 1a, 4a and 4b, genital and aggenital setae thickened, slightly dilated in median part, barbed, epimeral setae 3b, 3c and 4c setiform, barbed, anal and adanal setae slightly thickened, erect, ad1 and ad2 long. Circumpedal carinae directed to pedotecta I. One seta inserted on anterior edges of genital plates. Adanal lyrifissures located in preanal position in males and in inverse apoanal position in female. Postanal porose area absent.
Description
Measurements. Body length: 597 (holotype: ♂), 531–614 (7 paratypes: ♂♂); 597 (1 paratype: ♀); notogaster width: 448 (holotype: ♂), 415–448 (7 paratypes: ♂♂); 448 (1 paratype: ♀). Integument. Body colour brown to dark brown. Body surface, pteromorphs, genital and anal plates, subcapitular mentum and legs heavily and densely granulate (diameter of granules up to 1). Anterior part of prodorsum and often antero-lateral parts of ventral plate tuberculate (diameter of tubercles up to 4). Pteromorphs distinctly reticulate. Genital plates slightly longitudinally striate. Prodorsum. Rostrum pointed. Lamellar (L) and sublamellar (S) lines distinct, parallel. Rostral (ro, 57–61), lamellar (le, 86–94) and interlamellar setae (in, 94–98) setae setiform, barbed. Bothridial setae (bs, 136–145) setiform, ciliate medio-distally. Exobothridial setae and sejugal porose areas absent. Notogaster. Dorsosejugal furrow interrupted medially. Dorsophragmata (D) elongated longitudinally. Notogastral setae represented by 10 pairs of alveoli. Three pairs of saccules (S2 absent) with elongate channels and small openings: Sa located between la and lm and equally distanced from them, S1 – anteriorly and nearly to h3, S3 – posteriorly to h1. Median pore (mp) absent in female and present in males, located posteriorly to the virtual lines connecting lp. All lyrifissures (ia, im, ip, ih, ips) distinct, im located laterally to h3 and distanced from them. Opisthonotal gland openings located posteriorly to im. Gnathosoma. Generally, morphology of subcapitulum, palps and chelicerae typical for Galumnidae (see Engelbrecht Citation1969; Ermilov and Anichkin Citation2010, Citation2011). Subcapitulum size: 141–145 × 123–127. Subcapitular setae (a, m, h) similar in length (28–32) setiform, slightly barbed. Adoral setae (or1, or2, 16–20) setiform, curved distally, barbed. Palps (118–123) with typical setation: 0–2–1–3–9(+ω). Axillary saccules (sac) slightly elongated. Chelicerae (192–196) with two setiform, barbed setae, cha (53–57) longer than chb (36–41). Trägårdh’s organ long, tapered. Epimeral and lateral podosomal regions. Anterior margin of ventral plate smooth. Apodemes 2, 3 and sejugal apodemes well visible, similar in length. Epimeral setal formula: 1–0–2–3. Setae 1a (28–36) and 4a, 4b (24–28) thickened, slightly dilated in median part, barbed, setae 3b, 3c and 4c (36–41) setiform, barbed. Pedotecta I concave, pedotecta II trapezoid in ventral view. Discidia (dis) triangular, slightly blunted distally. Circumpedal carinae (cp) distinct, directed anteriorly to pedotecta I. Anogenital region. Six pairs of genital (g1–g6) and one pair of aggenital (ag) setae similar in length (20–24), thickened, slightly dilated in median part, barbed. Genital setae on each genital plate inserted in one longitudinal row with one seta on anterior edge. Two pairs of anal (an1, an2, 28–36) and three pairs of adanal (ad1, 57–65; ad2, 53–61; ad3, 32–41) setae slightly thickened, erect, barbed. Adanal setae ad3 inserted laterally to anal aperture. Adanal lyrifissures (iad) located in preanal position in males and in inverse apoanal position in female. Postanal porose area absent. Legs. Median claw thicker than laterals, all indistinctly barbed on dorsal sides. Formulas of leg setation and solenidia: I (1–4–3–4–20) [1–2–2], II (1–4–3–4–15) [1–1–2], III (1–2–1–3–15) [1–1–0], IV (1–2–2–3–12) [0–1–0]; homologies of setae and solenidia indicated in Table . Many setae well barbed, some ventral setae ciliate. Solenidia ω1 and ω2 on tarsi II and σ on genua III thickened, blunt-ended, other solenidia setiform. Solenidia σ of genua II inserted dorso-antiaxially, specifically curved. Solenidia φ of tibiae IV inserted dorsally in posterior part of segment.
Table 1. Leg setation and solenidia of adult Sacculogalumna samarensis n. sp.
Type material examined
Holotype (♂): Mt. Huraw, San Jose de Buan municipality, Western Samar Province, Samar Island, Philippines, 25.X.2002, collected by William Sm. Gruezo, extracted with Berlese funnel from sample of secondary forest litter. Three paratypes (3♂♂): same data as holotype. Four paratypes (3♂♂, 1♀): Sitio Napatuo, Barangay Osmena, Dolores municipality, Eastern Samar Province, Samar Island, Philippines, 21.X.2003, collected by William Sm. Gruezo, extracted with Berlese funnel from sample of secondary forest litter. One paratype (1♂): Barangay Bagong Silang, Basey municipality, Western Samar province, Samar Island, Philippines, 7.X.2003, collected by William Sm. Gruezo, extracted with Berlese funnel from sample of secondary forest litter over limestone.
Depository
The holotype (♂; alcohol) and two paratypes (2♂♂; alcohol): Senckenberg Institute, Görlitz, Germany; six paratypes (5♂♂, 1♀; alcohol): Tyumen State University Museum of Zoology, Tyumen, Russia.
Etymology
The specific name samarensis refers to the Philippine Island, Samar, where the holotype and paratypes were collected.
Remarks
All known species of Sacculogalumna can be distinguished by the key presented below.
Differential diagnosis
Sacculogalumna samarensis n. sp. differs from both known species of the genus, S. saccularis and S. suiyangensis, by two important morphological characters: (1) lamellar setae inserted between lamellar lines (vs. between lamellar and sublamellar lines); (2) adanal lyrifissures clearly distanced from anal plates (vs. located near anal plates). These characters are basic and usually constant in genera and subgenera of Galumnidae (Grandjean Citation1936; Balogh and Balogh Citation1990; Ermilov and Anichkin Citation2014). Hence, theoretically, we could offer a new genus or subgenus, because there are no superspecies among galumnid taxa with the combination of listed above characters as in the new species. Reasons of our refusal of this idea in the following:
| (1) | In spite of the fact that the position of lamellar setae has been used often as diagnostic generic character earlier, there are several genera and subgenera in Galumnidae, which include species with a different position of these setae (between lamellar lines and between lamellar and sublamellar lines), for example, Galumna (Cosmogalumna) Aoki, Citation1988 (see revised diagnosis in Ermilov and Corpuz-Raros Citation2015c) and Notogalumna Sellnick, Citation1959 (see revised diagnosis in Ermilov and Corpuz-Raros Citation2015a). Besides, in some cases lamellar setae are located very near to lamellar lines and their position can be changeable in relation to these lines (Ermilov et al. Citation2013). | ||||
| (2) | The localization of adanal lyrifissures (distanced from anal plates or near to them) is an important character in support of some galumnid taxa, for example, Allogalumna (Globogalumna) Balogh & Balogh, Citation1990; Indogalumna Balakrishnan, Citation1985; Galumna (Neogalumna) Hammer, Citation1973. However, the problem is that some genera include species with variable positions of adanal lyrifissures. For example, the adanal lyrifissures of many representatives of Pilogalumna Grandjean, Citation1956 and Trichogalumna Balogh, Citation1960 are located near anal plates, but they clearly distanced from anal plates in Pilogalumna bloemfonteinensis Engelbrecht, Citation1972a and Trichogalumna punctata Engelbrecht, Citation1972b. | ||||
Hence, based on localization of lamellar setae and adanal lyrifissures which can vary within one genus/subgenus, we consider it impossible to offer a new superspecies taxon and include it in Sacculogalumna.
In addition, the new species has genital setae inserted in one row on each plate with one seta on anterior edge, and has reticulate pteromorphs. These characters are very seldom in Galumnidae, however, they are specific and noted in selected species in different genera: for example, similar localization of genital setae present in Allogalumna superporosa Mahunka Citation1996 Dimidiogalumna comoroensis Mahunka, Citation1994 Galumna (Galumna) armatifera Mahunka, Citation1996 reticulate pteromorphs – in Stictozetes bisulcatus (Balogh, Citation1960), some Galumna (Galumna) Heyden, Citation1826 (G. (Galumna) reticulata Hammer, Citation1958) and Pergalumna (Pergalumna) Grandjean, Citation1936 (P. (Pergalumna) striata (Pérez-Íñigo & Baggio, Citation1980), some populations of P. (Pergalumna) emarginata (Banks, Citation1895) and P. (Pergalumna) nervosa (Berlese, Citation1914)), and many representatives of the subgenus Galumna (Cosmogalumna).
Key to known species of Sacculogalumna
| 1 | Rostrum pointed; lamellar setae inserted between lamellar lines; bothridial setae setiform; dorsosejugal furrow interrupted medially; pteromorphs reticulate; adanal setae ad1 and ad2 long, thickened; anterior edges of genital plates with one genital seta; adanal lyrifissures in preanal or inverse apoanal position, distanced from anal plates; postanal porose area absent; body size: 531–614 × 415–448 … S. samarensis n. sp. Distribution: Philippines. | ||||
| - | Rostrum rounded; lamellar setae inserted between lamellar and sublamellar lines; bothridial setae with developed heads; dorsosejugal furrow complete; pteromorphs not reticulate; anterior edges of genital plates with 2 or 3 genital setae; adanal setae ad1 and ad2 short, simple; adanal lyrifissures in paraanal position, near to anal plates; postanal porose area present......................................................................................................2 | ||||
| 2 | Notogastral setae absent, represented by alveoli; interlamellar setae long, longer than lamellar and rostral setae; bothridial setae widely lanceolate, with well-developed heads, barbed; saccules Sa located near to pteromorphal hinges, anteriorly to setal alveoli la; median pore present; adanal setae ad3 inserted laterally to adanal lyrifissures; postanal porose area elongate oval; body size: 438–493 × 315–339 … S. saccularis Engelbrecht, Citation1973. Distribution: South Africa. | ||||
| - | Notogastral setae present, minute; interlamellar setae short, shorter than lamellar and rostral setae; bothridial setae narrowly lanceolate, with slightly developed heads, smooth; saccules Sa located distanced from pteromorphal hinges, located between setae la and lm; median pore absent; adanal setae ad3 inserted anteriorly to adanal lyrifissures; postanal porose area rounded; body size: 410–440 × 320–335 .…… S. suiyangensis Liang, Yang & Huang, Citation2012. Distribution: Southern China. | ||||
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
The taxonomic study on Galumnoidea was supported by the Russian Foundation for Basic Research [project number 15-04-02706 A].
Acknowledgements
We cordially thank two anonymous reviewers for the valuable comments; Dr William Sm. Gruezo (College of Arts and Sciences and Museum of Natural History, University of the Philippines Los Baños, College, Laguna, Philippines) for his extensive litter samples in Samar Island including material used in this study; and Mr Jeremy Naredo (Museum of Natural History, University of the Philippines Los Baños, College, Laguna, Philippines) for facilitating transmittal of the specimens, together with other oribatids, to Russia.
References
- Aoki J. 1988. Oribatid mites (Acari: Oribatida) from the Tokara Islands, Southern Japan. II. Bulletin of the Biogeographical Society of Japan. 43(6):31–33.
- Balakrishnan MM. 1985. A new genus of Galumnidae (Acari: Oribatei) from India. Oriental Insects. 19:155–161.
- Balogh J. 1960. Oribates (Acari) nouveaux d’Angola et du Congo Belge (2ème série). Companhia de Diamantes de Angola, Lisboa. 51:15–40.
- Balogh J, Balogh P. 1990. Identification key to the genera of the Galumnidae Jacot, 1925 (Acari: Oribatei). Acta Zoologica Hungarica. 36(1–2):321–328.
- Banks N. 1895. On the Oribatoidea of the United States. American Entomological Society. 22:1–16.
- Berlese A. 1914. Acari nuovi Manipulus IX. Redia. 10:113–150.
- Engelbrecht CM. 1969. Some South African species of the genus Galumna von Heyden, 1826 (Acari: Galumnidae). Journal of the Entomological Society of Southern Africa. 32(1):99–122.
- Engelbrecht CM. 1972a. Ctenogalumna moresonensis sp. n. and Pilogalumna bloemfonteinensis sp. n., two new South African species of the subfamily Allogalumninae Balogh. 1960 Galumnidae: Oribatei. Acarologia. 14(3):497–510.
- Engelbrecht CM. 1972b. South African species of the genus Trichogalumna Balogh, 1961 (Galumnidae: Oribatei). Acarologia. 14(2):268–280.
- Engelbrecht CM. 1973. South African Galumnoidea (Oribatei: Acari): new taxa and records. Navorsinge van die Nasionale Museum, Bloemfontein. 2(2):394–415.
- Ermilov SG, Anichkin AE. 2010. Three new species of Galumnidae (Acari: Oribatida) from Cat Tien National Park, southern Vietnam. Zootaxa. 2681:20–34.
- Ermilov SG, Anichkin AE. 2011. New oribatid mites of the genera Pergalumna and Galumnella (Acari, Oribatida, Galumnoidea) from Vietnam. Acarina. 19(2):242–251.
- Ermilov SG, Anichkin AE. 2014. Taxonomic study of oribatid mites (Acari, Oribatida) of Bi Dup – Nui Ba National Park (southern Vietnam). Zootaxa. 3834(1):1–86.
- Ermilov SG, Corpuz-Raros LA. 2015a. A new species of Notogalumna (Acari, Oribatida, Galumnidae) from the oriental region with a key to known species. Acarologia. 55(3):267–276.
- Ermilov SG, Corpuz-Raros LA. 2015b. New species of galumnid oribatid mites of the genera Mirogalumna and Pergalumna from the Philippines (Acari, Oribatida, Galumnidae). Systematic and Applied Acarology. 20(5):556–566.
- Ermilov SG, Corpuz-Raros L. 2015c. New species of oribatid mites with auriculate pteromorphs (Acari, Oribatida, Galumnidae) from the Philippines. Zootaxa. 3905(4):511–528.
- Ermilov SG, Corpuz-Raros L, Shimano S. 2014a. First reports of Galumnellidae (Acari, Oribatida) from the Philippines, with description of Galumnella junichiaokii sp. n. Acarina. 22(2):85–91.
- Ermilov SG, Corpuz-Raros L, Tolstikov AV. 2014b. The oribatid mite subgenus Galumna (Galumna) (Acari, Oribatida, Galumnidae) in the Philippines. ZooKeys. 452:1–13.
- Ermilov SG, Weigmann G, Tolstikov AV. 2013. Morphology of adult and juvenile instars of Galumna obvia (Acari, Oribatida, Galumnidae), with discussion of its taxonomic status. ZooKeys. 357:11–28.
- Grandjean F. 1936. Les Oribates de Jean Frédéric Hermann et de son père. Annales de la Société Entomologique de France. 105:27–110.
- Grandjean F. 1956. Galumnidae sans carènes lamellaires (Acariens, Oribates). 1e série. Bulletin de la Société Zoologique de France. 81(2–3):134–150.
- Hammer M. 1958. Investigations on the oribatid fauna of the Andes Mountains. I. The Argentine and Bolivia. Det Kongelige Danske Videnskabernes Selskab Biologiske Skrifter. 10((1)):1–129.
- Hammer M. 1973. Oribatids from Tongatapu and Eua, the Tonga Islands, and from Upolu, Western Samoa. Det Kongelige Danske Videnskabernes Selskab Biologiske Skrifter. 20(3):1–70.
- Heyden C. 1826. Versuch einer systematischen Eintheilung der Acariden. Isis, Oken. 1(4):607–613.
- Liang W, Yang M, Huang R. 2012. First record of the genus Sacculogalumna Engelbrecht (Oribatida: Galumnidae) in China. Entomotaxonomia. 34(2):489–493.
- Mahunka S. 1994. Further oribatid species from the Comoro Islands (Acari: Oribatida). Acta Zoologica Academiae Scientiarum Hungaricae. 40(3):273–288.
- Mahunka S. 1996. Galumnoid taxa (Acari: Oribatida) from Madagascar (Part I). Acta Zoologica Academiae Scientiarum Hungaricae. 42(2):163–181.
- Norton RA. 1977. A review of F. Grandjean’s system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal DL, editor. Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. p. 33–61.
- Norton RA, Behan-Pelletier VM. 2009. Oribatida. In: Krantz GW, Walter DE, editors. A manual of acarology. Lubbock: Texas Tech University Press; Chapter 15; p. 430–564.
- Pérez-Íñigo C, Baggio D. 1980. Oribátidos edáficos do Brasil. I. Boletin de Zoologia da Universidade de São Paulo. 5:111–147.
- Sellnick M. 1959. Acarina from Southeastern Polynesia–II (Oribatidae). Occasional papers of Bernice P Bishop Museum. 22(9):109–152.
- Travé J, Vachon M. 1975. François Grandjean. 1882–1975 (Notice biographique et bibliographique). Acarologia. 17(1):1–19.