
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Following floods experienced in southwestern Alberta in June 2013, 36 palaeoenvironmental sediment samples were collected from the banks of four affected streams: the Kananaskis River, Highwood River, Tongue Creek and Jumpingpound Creek. The sampled layers were deposited during the Holocene, and provide evidence of riparian ecology in the region since the end of the last glacial interval. The samples were processed to extract sub-fossil macroremains (including seeds, fruits, and aquatic and terrestrial mollusc shells). Macrofossils are particularly useful for creating fine-resolution (site-specific) reconstructions. An innovative aspect of this study was the derivation of regional bioclimatic trends through time. This was achieved by applying a dual-layered weighted calibration function which incorporated the relative productivity of indicator taxa, their modelled climatic optima, and the influence of climate (temperature and precipitation) on their ecological niche. Optima and climatic influence values were calculated using the machine-learning maximum entropy environmental niche modelling tool, MAXENT, trained on global occurrence records for the taxa. This study aimed to address gaps in prior palaeoenvironmental research, allowing for a spatially explicit quantitative reconstruction of climate in the transitional foothill region near Calgary. Results indicated warm conditions immediately following glacial retreat, followed by an early to mid-Holocene cooling which coincided with the driest interval. Temperature and moisture regimes oscillated in the mid-Holocene, exhibiting the highest values for both parameters. Increased moisture was at least in part prompted by increased input from summer glacial meltwater (the result of higher summer temperatures) which added to the riparian water budget. The mid- to late Holocene, and up to modern times, showed a gradual decrease in both moisture and temperature, until stabilizing near the Holocene average.
À la suite des inondations subies dans le sud-ouest de l’Alberta en juin 2013, 36 échantillons de sédiments paléoenvironnaires ont été prélevés sur les rives de quatre cours d’eau touchés: la rivière Kananaskis, la rivière Highwood, le ruisseau Tongue et le ruisseau Jumpingpound. Les couches échantillonnées ont été déposées pendant l’Holocène et fournissent des preuves d’écologie riveraine dans la région depuis la fin du dernier intervalle glaciaire. Les échantillons ont été traités pour extraire les macrorémains sous-fossiles (y compris les graines, les fruits et les coquilles de mollusques aquatiques et terrestres). Les macrofossiles sont particulièrement utiles pour créer des reconstructions à résolution fine (site spécifique). Un aspect novateur de cette étude a été la dérivation des tendances bioclimatiques régionales à travers le temps. Ceci a été réalisé en appliquant une fonction d’étalonnage pondérée à deux couches qui a incorporé la productivité relative des taxons indicateurs, leur optimum climatique modélisé et l’influence du climat (température et précipitations) sur leur niche écologique. Les valeurs de l’optima et de l’influence climatique ont été calculées à l’aide de l’outil de modélisation de la niche environnementale d’entropie maximale, Maxent, formé sur les enregistrements d’occurrence globale des taxons. Cette étude visait à combler les lacunes de la recherche paléoenvironnementale antérieure, permettant une reconstruction quantitative spatialement explicite du climat dans la région des contreforts de transition, près de Calgary. Les résultats indiquaient des conditions chaudes immédiatement après la retraite glaciaire, suivies d’un refroidissement du début à la mi-Holocène qui coïncidait avec l’intervalle le plus sec. Les régimes de température et d’humidité ont oscillé au milieu de l’Holocène, affichant les valeurs les plus élevées pour les deux paramètres. L’augmentation de l’humidité a été au moins en partie motivée par une augmentation de l’apport de l’eau de fonte glaciaire d’été (résultat des températures estivales plus élevées) qui s’est ajoutée au budget riverain de l’eau. L’Holocène moyen à tardif, et jusqu’aux temps modernes, a montré une diminution progressive de l’humidité et de la température, jusqu’à stabilisation près de la moyenne Holocène.
Introduction
Floods influence the geomorphology of rivers and adjacent landscapes while exerting unique selective pressures on riparian biota (Gottesfeld Citation1996; Hupp and Osterkamp Citation1996; Kozlowski Citation2002; Katz et al. Citation2005). In 2013, several watersheds in southwestern Alberta were impacted by a flood event that modified channels and damaged infrastructure (Calgary Herald Citation2013; Milrad et al. Citation2015; Pomeroy et al. Citation2015). Following the flood, the Archaeological Survey of Alberta (ASA) implemented a multi-year flood impact assessment and subsequent mitigation project. The project was focused primarily on heritage resources – that is, assessment of the impact on archaeological and historic sites. As part of this project, samples of organic-rich sediments were collected from the banks of several of the most impacted streams. This collecting was in part to provide contextual information for the postglacial human history of this region. Samples were processed to extract macroremains, with the objective of developing local and regional riparian palaeoenvironmental reconstructions. Macrofossil data from the Highwood River have been presented in Bolton and Beaudoin (Citation2016); these and other samples are documented in reports to the ASA (Bolton Citation2016, Citation2017). The macrofossil data were used to construct a regional representation of riparian climatic trends (moisture and temperature) since the end of the Pleistocene, using a novel application of maximum entropy ecological niche modelling.
Resolution of palaeoenvironmental analysis
Biological remains have been used as proxies for palaeoclimatic reconstructions at site, regional, and continental scales (e.g. Dyke Citation2005; Lorenz et al. Citation2016). These reconstructions, a type of climatic model, are predominantly based on pollen and tree-ring data (e.g. Luckman et al. Citation1997; Davis et al. Citation2003), and, to a lesser extent, macrofossils (Bartlein et al. Citation2011). Biological proxy data are also used to calibrate and validate meteorological models that predict future climate (e.g. global circulation models, GCMs) via back-testing, as in the Paleoclimate Modelling Intercomparison Project (PMIP), now moving into its fourth phase (https://pmip4.lsce.ipsl.fr/doku.php). However, for both modelling approaches (meteorological and biological) the scale of the output determines the applicability of the model (Jost et al. Citation2005). Restrictions on the spatial resolution of models are particularly pertinent in the case of riparian environments, because river valleys are frequently smaller than the resultant gridded output cells from climate models.
Additionally, with a few exceptions, many palaeoclimatic GCMs and biological proxy models lack precise temporal resolution. Many proxy models are restricted to only one or a few sites (e.g. Vance et al. Citation1995), and most available pre-historic GCM results present only one interval, the mid-Holocene (ca. 6000 years before present [yr BP]), with only a few providing a second period, the last glacial maximum (LGM; ca. 22,000 yr BP). Among the exceptions are Lorenz et al. (Citation2016) and Viau and Gajewski (Citation2009), who report reconstructed climate throughout the Holocene at sub-millennial resolution, although even these reconstructions have limitations. For example, the data of Lorenz et al. (Citation2016) are downscaled from large-scale process-based models, each with its own limitations, and inherently pass on the bias or inaccuracy of the primary models to their results (Trzaska and Schnarr Citation2014). In the case of Viau and Gajewski’s (Citation2009) work, the outputs provide results whose geographic scope represents only a highly simplified version of inherently spatially heterogeneous phenomena; they reduced all of boreal Canada to just four representative zones.
Macrofossil analyses are well positioned to reduce the temporal and spatial scales of palaeoclimatic reconstructions, especially because their preservation is usually representative of a unique and distinct site (unlike pollen analysis, which can incorporate signals from plants sometimes kilometers distant from their source, obscuring local patterns) and remains from flood deposits characterize biota from fixed time-slices. Moreover, the presence of delicate remains indicates minimal transport or reworking (Burnham Citation1993; Work et al. Citation2005). Where biologically or ecologically associated remains are preserved together it suggests that the material (including the fossil material and the sediment matrix) is largely in situ (Trewin et al. Citation2004). In an alluvial context, Thomasson (Citation1991) showed that modern seeds trapped in riparian sediments were accurate predictors of the local extant vegetation. However, taphonomic effects in fluvial settings may be complex. Remains may have originated elsewhere in the up-slope catchment area; further, evidence of previous burial and redeposition, particularly from similar communities or of very robust taxa, is not always discernable. Aarnes et al. (Citation2012) have recently applied macrofossil assemblage data to generate quantitative climate reconstructions; they also outline the advantages (and limitations) of this type of analysis.
To date, there are remarkably few late Quaternary palaeoenvironmental reconstructions focussed on riparian conditions. A few riverine projects are largely from localities outside of North America, including South Africa (Baxter Citation1997), Australia (Gell et al. Citation2009; Gell Citation2010), France (Ejarque et al. Citation2015) and Turkey (Alçiçek et al. Citation2017), or focus on older intervals, such as Marmi et al.’s (Citation2015) study from the Maastrichtian of Spain. The few North American examples are from areas distant from Alberta, such as California (Brown and Pasternack Citation2005), Iowa (Work et al. Citation2005), New York (Rippke et al. Citation2010) and Ontario (Thornbush and Desloges Citation2011). Aside from the present study, no similar reconstructions have been conducted in the Canadian Prairies or adjacent Rocky Mountains. This study therefore helps address spatial and temporal gaps in available coverage.
Methods
Study area
This project focuses on four catchment basins in southwestern Alberta: the Highwood River, Kananaskis River, Tongue Creek and Jumpingpound Creek (Figure ). Bankfull flow return intervals, i.e. how often streams have sufficient flow to reach their bank limits, are often estimated at about every 1.5 to 2 years (Leopold Citation1994). However, much research suggests that this value may be inappropriate for estimating bankfull and greater events (i.e. floods) in Alberta. Albertan bankfull return is generally interpreted as occurring at intervals of no less than 2.4 years (Smith Citation1979), and in some cases, considerably more rarely (e.g. 40 years or greater per event; Smith Citation1979; Ward et al. Citation2010). Using modern regional bankfull return intervals as a guide, it is reasonable to assume that floods (and their associated overbank deposition) occurred perhaps every 10 or 15 years in this region during the Holocene. However, stratigraphic evidence from this study’s sample sites suggests that major (accretional) events were considerably rarer (i.e. only a few each millennium or more). It is this sort of substantial flood deposit that capped most of the palaeoenvironmental samples examined in this study, preserving them for hundreds to thousands of years.
Figure 1. Map of study area, surrounding region, and drainage area for the downstream-most location sampled on each stream.
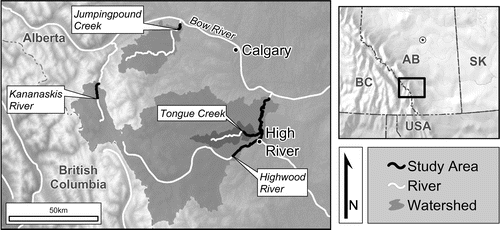
The region’s modern climate is a mixture of humid continental and continental subarctic types (respectively, Dfb and Dfc, Köppen–Geiger–Pohl climate classifications), with the highest regions of the nearby Canadian Rockies presenting an alpine tundra climate (ET; Kottek et al. Citation2006). Annually the watersheds of the study area receive ~600 mm of precipitation (water equivalent) on average, with ~250 cm falling as snow. This is ≈250 mm snow water equivalent (SWE) assuming a 10:1 relationship between snow depth and SWE. This is a generally accepted estimate of average SWE, as used at Environment and Climate Change Canada (Citation2016). However, studies in alpine areas (e.g. Sevruk Citation1986), including the Kananaskis Valley (Musselman et al. Citation2015), indicate that snow density is decidedly heterogeneous, and can often be denser than this general rule indicates. The SWE input should therefore be taken as a minimum estimate. The annual mean temperature of the basins is 1.4°C (Figure ). Not surprisingly, the local annual climatic parameters show marked variation with elevation. Elevation is a consistently implemented covariate for predicting climate at fine scales (Goodale et al. Citation1998; Hijmans et al. Citation2005; Simpson et al. Citation2005) and summer orographic lapse rates are well known in the region (e.g. Berg et al. Citation2007), although winter inversions often complicate conditions, particularly in topographically varied terrain (Daly et al. Citation2008).
Figure 2. Mapped climatic parameters of study watersheds, modelled using geographically weighted regression (GWR). (a) Total annual precipitation (mm of water equivalent); (b) total annual snowfall (cm of fresh snow); (c) mean annual temperature (°C). Data source: Canadian Climate Normals 1981–2010, climate normal and averages (Environment and Climate Change Canada Citation2016).
Sampling and chronology
In October 2014, sections of the Highwood River were surveyed for exposures of strata rich in palaeoenvironmental remains (i.e. those which contained visible mollusc shells or high organic matter concentrations; Bohach and Frampton Citation2015). At each site the estimated landscape age for the palaeoenvironmental layer was assigned based on the terrace height relative to the modern stream level and other indicators (L. Bohach, Stantec Consulting Ltd., pers. comm. 2017). Low terraces (up to 2 m) were considered very young (e.g. late Holocene, or were classified as ‘modern’ if they contained seeds from introduced weeds); medium terraces (2 to 5 m) were deposited earlier (e.g. mid-to late Holocene); high terraces (5 to 25 m) represented even older sediments, sometimes from the earliest major fluvial incision following glaciation (e.g. early to mid-Holocene). Stream meander dynamics were also used to inform age interpretation; the meander interiors (e.g. slip-off slopes) were more recently deposited than the exteriors of bends (e.g. along cut banks). In the area surrounding the study area, including along the Kananaskis, Highwood, and Bow Rivers, the river terraces have been well studied and age relationships are reasonably well supported by radiocarbon and tephra dating (Wilson Citation1974, Citation1993; Stene Citation1980; Pennock Citation1984).
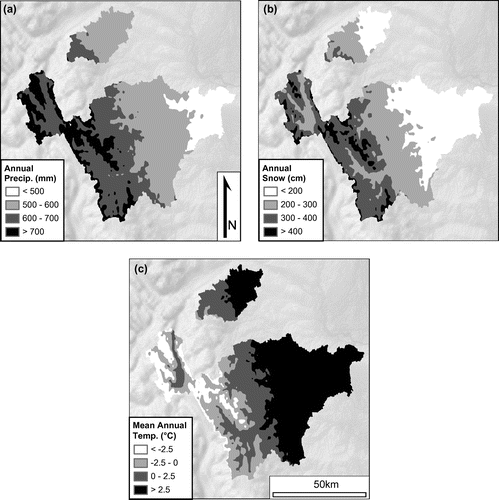
Stratigraphic information, especially the relationship of samples to the regionally deposited Mazama tephra (Beaudoin and Oetelaar Citation2006), provided additional chronologic control. The Mazama tephra is dated at about 7600 cal. yr BP (Zdanowicz et al. Citation1999) and provides a region-wide chronostratigraphic marker delineating early from mid-Holocene sediments. Varved glaciolacustrine sediments in the terraces represent late Pleistocene sediments. Relative positions of beds within a single exposure were then assessed to provide a gauge of deposition age for the palaeoenvironmental samples, keeping in mind the often-episodic aggradational (and degradational) regimes near streams.
The age classes defined in the field, their relative (estimated) chronologic extents, and relationship to the Mazama tephra layer are shown in Figure . The ages therein are presented in calendrical equivalent years before present (cal. yr BP; where ‘present’ datum = 1950). The calendrical equivalent dates were calibrated from conventional radiocarbon dates (14C yr BP) using OxCal software (v. 4.3; Bronk Ramsey Citation2009) and the IntCal13 calibration curve (Reimer et al. Citation2013). All further dates in this paper are relayed in calendrical equivalent form; in cases where only conventional 14C dates were reported, both are given.
Figure 3. Estimated chronological extent of field-determined intervals used in this study. Circles represent the mean values of the upper and lower limits (vertical bars) per class. The tephra chronostratigraphic layer, the Mazama ash, is indicated by the dashed line, dated to ~7600 cal. yr BP (Zdanowicz et al. Citation1999; Egan et al. Citation2015).
Where possible, radiocarbon dating of wood and terrestrial macrofossils (e.g. Picea needles) was used to validate the general interpretations of the field-determined age classes (Table ). Most recovered macrofossils were from aquatic taxa and were not appropriate for radiocarbon dating (Oswald et al. Citation2005). In addition, much of the bedrock in the area is carbonate rich; problems of dating aquatic macroremains or bulk sediments in this region are well documented (MacDonald et al. Citation1987; Wilson Citation1993). Nevertheless, the radiocarbon dates, though few, lend support to the age classes based on stratigraphic and landscape position (Table ; Figure ).
Table 1. 14C dates, calibration, and comparative radiocarbon and field-determined age classes. Near-modern samples (denoted by *) were calibrated with the Bomb13NH1 curve (Hua et al. Citation2013), all others used the IntCal13 curve (Reimer et al. Citation2013). Samples processed at the André E. Lalonde Accelerator Mass Spectrometry Laboratory (Ottawa, Ontario) and the Keck Carbon Cycle Accelerator Mass Spectrometry facility (Irvine, California).
Sample processing and analysis
Bulk sediment samples of at least 1 kg in weight were collected from the palaeoenvironmental layers and allowed to air-dry. This process was repeated along sections of the Kananaskis River, Tongue Creek and Jumpingpound Creek during October 2015. This work was undertaken by consultants as part of the flood assessment project directed by the Archaeological Survey (Bohach and Frampton Citation2015; Bohach Citation2016). In total, 36 samples were collected from 25 localities. The numerical distributions and spatial proximities of samples and localities for each stream are shown in Table .
Table 2. Number of localities and samples, and maximum stream-course distance between sites per stream.
The samples were processed at the Royal Alberta Museum’s Quaternary Environments laboratory. From each sample, 100 mL of material was wet-screened to consolidate macrofossils. Initially the samples were soaked and stored at 3°C for at least 12 h. Very strongly aggregated (e.g. highly clay/organic-rich) samples were soaked for longer, in some cases up to several weeks. After soaking, the material was pre-screened on a 90-μm sieve to remove the majority of clay and silt particles. The remaining material was fractionated by wet-screening following Beaudoin (Citation2007) on a series of nested sieves (3.35 mm to 90 μm).
The fractionated material was sorted to remove macrofossils from the residual matrix. Sorting was conducted under a dissecting microscope (7.5 to 112.5× magnification) with an auxiliary ‘cool light’ illuminator. Trays of material were continuously scanned while removing macrofossil material with fine forceps until readily identifiable remains were no longer detected.
Macrofossils were identified using published sources and comparative material. Resources of particular note included Montgomery (Citation1977) for seeds and fruits, Clarke (Citation1981) for freshwater molluscs and Forsyth (Citation2004) for terrestrial molluscs. Whenever possible the Royal Alberta Museum’s seed reference collection was used to verify identifications. All macroremains are accessioned and archived at the Royal Alberta Museum.
Climate reconstruction
Predicting optimum climatic parameters
After identification, the results of all the samples were compiled and sorted according to the age class of their deposition. Samples were ‘binned’ into relatively broad temporal categories (as per Figure , plus ‘modern’ where they included recently introduced weed taxa). Data representing entities which occurred in multiple age classes were separated for further analysis. These taxa, henceforth ‘indicator taxa’, were used in the climate modelling. The relative abundance of these indicator taxa (which varied over time) was used to derive the climate signal. As such, taxa that occurred during only one interval were excluded from the analysis. Taxa with globally rare georeferenced records (arbitrarily < 50, as per the Global Biodiversity Information Facility ([GBIF Citation2016; www.GBIF.org]; i.e. with poorly known geographic extents) were removed from the pool of eligible indicators. The final list of selected indicator taxa is displayed in Table ; their general organismal group and habitat are also included.
Table 3. List of Linnaean and common names, authorities, and organismal groups of indicator taxa used for climate reconstruction.
The GBIF database was queried for records of those taxa. For taxa identified at the genus level or higher (including multiple-species complexes or indistinguishable species), all records for the constituent species were compiled (e.g. all records of Gyraulus parvus and G. circumstriatus were retrieved). These records were assigned an aggregate identifier as well, such that all groups could be filtered for or analyzed simultaneously. In the case of Zonitoides spp. land snails, only the species Z. arboreus and Z. nitidia were combined because these are the only two species of this genus from the region (Forsyth Citation2004, Citation2006). Although the aquatic mollusc Physa skinneri occurred in samples from multiple ages, it was not included in the climate reconstruction because it was only rarely recorded in GBIF data (93 total records, only 19 of which were georeferenced), and, as such, contributed an unrepresentative climate response profile. Record data was parsed to remove fossil occurrences, non-georeferenced records, and those points whose recorded coordinates failed to align with the documented country of origin. These data, along with modern annual average temperature and total annual precipitation grids (10 arc-minute resolution) from the WorldClim v.1.4 data set (www.WorldClim.org; originally in Hijmans et al. Citation2005) were used as input data to the MAXENT model (v. 3.3.3 k; described in Phillips et al. Citation2004, Citation2006). MAXENT employs a ‘maximum entropy’ method to model the geographic distribution/environmental niches of taxa (Phillips et al. Citation2006; Merow et al. Citation2013). In addition to developing spatial models of probable habitat suitability (the normal published output), the software also develops species response curves for environmental variables (covariates) and can estimate the relative importance of the covariates on the modelled suitability (Phillips et al. Citation2006; Elith et al. Citation2011). It is these two capacities which were used in this study to model biotic optima; response curves and variable importance values for each indicator taxon relative to the two climatic parameters were calculated.
To aid in counteracting observation bias, grid-duplicate presence records were removed from the analysis, resulting in only one record per 10-arc-minute cell. Density related sub-sampling of this nature has been recommended by Fourcade et al. (Citation2014), among others, as a simple and effective bias reduction method. Up to 1000 model iterations were conducted, or until the drop in log-loss per iteration fell below 0.00001; this convergence threshold is recommended as a conservative value by the model originators (Phillips et al. Citation2006) and has been used in numerous studies to produce robust model results (e.g. Phillips et al. Citation2006; Bond Citation2012; Servat et al. Citation2013). The model was run with 10-fold cross-validation replicates for each of the 15 indicator taxa (Table ). Of the response curves generated (considering each of the climatic variables independently), the average values from all of the replicates were recorded. The maximum predicted suitability for temperature and precipitation (modelled using only the corresponding variable) were extracted from the average of each taxon; these values represent approximations of the actual (global) climatic optima for the taxa based on their known occurrences. The relative importance of the two climatic variables in determining the complete suitability model (i.e. the percent contribution of each variable) for each taxon was also calculated by averaging the covariate contribution values from each replicate.
Modelling regional climate
The modelling approach adopted in this study is that of a weighted average calibration function, fundamentally as described by Birks et al. (Citation2010), but instead of considering only climatic optima and a single weighting array based on taxon abundance, a second weight array was incorporated to reflect the relative influence that the climatic variable in question has on each taxon’s modelled niche. This method was adopted because it does not require analogous environments and relies on the simple, yet generally robust, ecological principal of unimodal species response to environmental gradients (see Ter Braak and Looman Citation1986; Rydgren et al. Citation2003; Horsák Citation2006; though Bongers et al. Citation1999 and Austin Citation1987, Citation2002 showed species response curves are not always bell shaped). Various other strengths and weaknesses of this and other quantitative climate reconstruction methods are described in Birks et al. (Citation2010).
The average number of entities per sample per age class was calculated for each of the taxa. Then the relative macrofossil yield, or ‘assemblage value’ (AV), of each taxon (taxon) per age class (age), AV(taxon;age), was calculated by dividing each age-taxon’s value (n(taxon;age)) by the maximum value of that taxon in any age (nmax(taxon)), yielding values in the range [0, 1]:(1)
(1)
A second set of weights were calculated for each taxon for each of the two climatic variables (denoted by clim) (Weight(clim;taxon)). Seen in Equation (Equation2)(2)
(2) , this calculation relativized the influence of a particular taxon–climate variable combination (influence(clim;taxon)) by the maximum value for that parameter on any taxon (influencemax(clim)):
(2)
(2)
For each of the two climatic parameters, a composite weight (C(taxon;age)) was calculated which combined the relative productivity of that taxon during an age class and the contribution of the climatic variable towards the ecological niche model (Equation Equation3(3)
(3) ). A unique composite weight for each taxon per age class was generated by multiplying the percent-of-most AV value by the relative influence of each variable, also relativized by the largest value for that parameter.
(3)
(3)
The contribution of each taxon to a particular age class’ modelled temperature or precipitation was calculated by multiplying the optimum for that species and parameter (Optimum(clim;taxon)) by the composite weight for that age/taxon and then dividing by the sum of all composite weights for that age:(4)
(4)
The sum of all the taxa’s contributions in an age class is the predicted temperature or precipitation for that age class. However, because the 15 taxa selected as indicators (Table ) do not relay the complete assemblages (e.g. communities) present in their habitats, calibration was required to align the values with other observed or modelled conditions. Temperature was scaled by a fixed amount (Z(temp)), such that the modelled modern temperature (Temp(modern)) equalled 1.4°C:(5)
(5)
Accordingly, temperatures for all age classes (CalTemp(age)) were calculated by subtracting the fixed value from the uncalibrated temperature for that period (Temp(age)):(6)
(6)
This additive method was adopted instead of multiplicative proration because multiplication-related scaling errors become significant when values close to 0°C are considered.
Precipitation was scaled to equal modern values as well, though because precipitation is an absolute measurement (i.e. 0 mm equals the complete lack of precipitation) it was scaled multiplicatively (e.g. a uniform scaling factor [Z(temp)] was multiplied by the values to calibrate them, such that the uncalibrated modern value [Precip(modern)] equals the known modern level):(7)
(7)
Equation Equation(8)(8)
(8) shows the relationship between the precipitation scaling factor and uncalibrated values from all ages (Precip(age)) to produce calibrated precipitation values for each period (CalPrecip(age)):
(8)
(8)
Particularly because there is no way to validate the calibration beyond the modern period, the resultant values should be considered estimated indicators of relative increase or decrease in climatic conditions between periods only. Considering this, the relative divergence from the mean (of the uncalibrated values) in percent was also calculated as a more representative assessment of climate change over time.
Results and discussion
Ecological optima
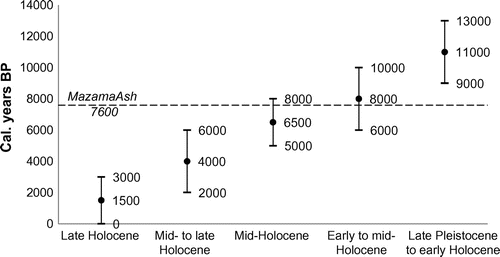
After parsing for multi-aged entities and limiting the analysis to those taxa with sufficient occurrence data to generate reliable statistical profiles, only 15 taxa were selected as bioclimatic indicators. These included 11 mollusc taxa and four plant taxa (Table ). The modelled optimum climatic preferences (from MAXENT) for the 15 indicator taxa are presented in a simplified climate space diagram (Figure ). Considering the niche characteristics of these taxa, there is a weak positive correlation (r2 = 0.20) between the two climatic variables. This indicates that when reconstructions based on these data are conducted both variables will co-vary rather similarly. However, because the majority of variability in these parameters is not explained by the linear relationship alone, this parameter similarity is considered immaterial. There is sufficient divergence of temperature and precipitation values such that the modelled results are expected to be representative of the actual environmental conditions. Cropper (Citation1984) also remarked on the co-linearity between temperature and precipitation; he concluded that at an annual scale the bias introduced is negligible.
Figure 4. Climate space (annual precipitation and mean annual temperature) diagram of 15 indicator taxa. Discus whitneyi = Dw, Eleocharis palustris = Ep, Euconulus fulvus = Ef, Galba modicella = Gm, Gyraulus parvus/circumstriatus = Gp/c, Helisoma anceps = Ha, Physa gyrina = Pg, Pisidium spp. = Psdm, Potamogeton spp. = Pgtn, Scirpus microcarpus = Sm, Stagnicola elodes = Se, Succineidae = S, Vallonia gracilicosta = Vg, Zannichellia palustris = Zp, and Zonitoides spp. = Za/n. Note: some labels have been slightly moved from their centroids to aid legibility.
Temporal regional results
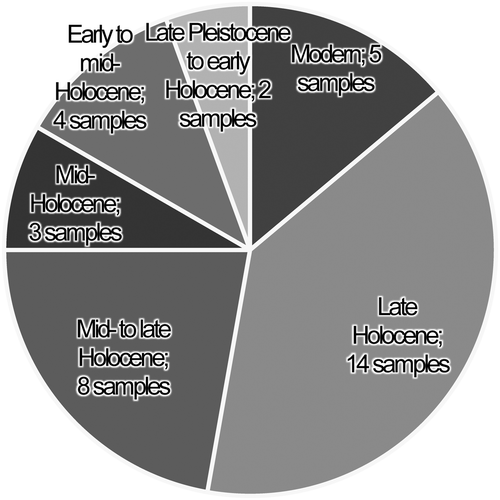
Because the ages of sampled localities are relatively evenly distributed across the study region (Figure ; Table ), an aggregate temporal reading of climate can be made by combining the average biological data per age class. Considerably more samples are available for analysis from more recent intervals (Figure ). Indeed, more late Holocene and modern samples were collected than all samples assigned to the mid-Holocene and earlier combined. Further, only two samples represent the late Pleistocene/early Holocene class. Owing to the unequal availability of various aged samples, there is more certainty about inferences based on the modern and late Holocene samples. Other aged samples presumably remain representative of the conditions in the region, though the amount of possible bias increases as the number of samples decrease.
Figure 5. Pie chart depicting the number of samples per age class.
Generally, more recent samples were more productive (i.e. yielded more taxa). This may be explained by climatic/ecological variables. However, a substantial amount of the observed difference probably arises from preservation bias, i.e. the loss of potentially identifiable remains due to taphonomic processes. These effects are known to particularly impact small and carbonaceous (e.g. shelly) macrofossils (Kidwell and Bosence Citation1991; Cooper et al. Citation2006). By using only those indicator taxa that were recorded in multiple age classes (and excluding potentially fragile or rarely preserved taxa) this bias over time was somewhat negated, allowing for a more realistic representation of the community productivity over time. By comparing relative abundances of the indicator taxa by age (as in Equation Equation4(4)
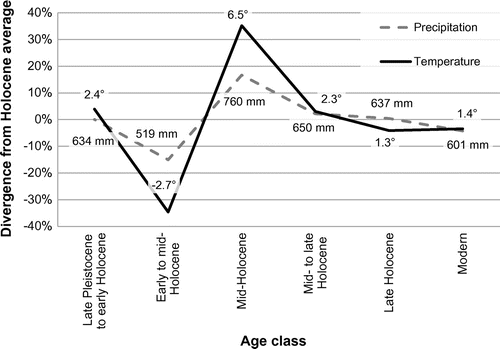
(4) ), this bias is further reduced (Table ). This approach does, however, assume that all indicators are affected by taphonomy equally over time, which is probably an oversimplification (Cooper et al. Citation2006). The subset of indicator taxa (Table ) was then used to model annual precipitation and mean annual temperature for the discrete age classes (Figure ). These results are the chief outcome of this study. Values are plotted relative to their divergence from the modelled mean over the entire Holocene for each of the two climate variables and annotated with the calibrated absolute values calculated per age class.
Table 4. Average number of indicators (by taxon) per age class and per sample. The initial value is the count of a taxon in all samples of an age class; the value in parentheses is the average value per sample; values with * are maximum values (e.g. AV = 1) for that taxon.
Figure 6. Modelled annual precipitation and mean annual temperature for the study region; plotted values represent uncalibrated percent divergence from mean; data labels indicate calibrated values (precipitation is in mm; temperature is in °C).
Late Pleistocene to early Holocene
The late Pleistocene to early Holocene interval (ca. 13,000 to 9000 cal. yr BP) appears to have been closely aligned with the modelled Holocene average, though it was probably about 4% warmer than the epoch average (Figure ). Rapid late Pleistocene warming has been identified in many studies. Viau and Gajewski (Citation2009) noted a similar increase in early Holocene temperature. Burn (Citation1997) also found the early Holocene to be warmer than present day in northwestern Canada. Likewise, Cole (Citation2010) remarked on the sudden shift from cool to warm temperatures in the early Holocene and the sometimes-dramatic changes in vegetation that accompanied the modulation. Additionally, the end of the Pleistocene and the early Holocene are known to have exhibited significant temperature oscillations (e.g. the Bølling and Allerød warm intervals and the Older and Younger Dryas cold intervals; Dyke Citation2004).
Unfortunately, little comparative information is available within this period (only two sites contribute to the regional reconstruction), though the Holocene maximum of both Gyraulus and Pisidium occurred at this time. These molluscs are ranked intermediately in terms of both precipitation and temperature regimes, explaining the medial interpretation during this time. Succineid snails were also present in these samples, though only at 4% of their Holocene maximum AV. Elsewhere in the Bow and Kananaskis watersheds, Beierle and Smith (Citation1998) also noted warm and arid conditions at terminal Pleistocene based on the widespread evidence of marl layers (indicating CaCO3 enrichment under evaporative conditions) and the conversion of some lakes to peat-developing systems.
Early to mid-Holocene
Declines in both temperature and precipitation were modelled for the early to mid-Holocene (ca. 10,000 to 6000 cal. yr BP; Figure ). This interval may have been the Holocene low for both parameters, though it was probably less dry than it was cool (15% below average precipitation, but 35% below average temperature). Although the absolute value of the modelled temperature during this time seems unexpectedly low (below freezing; Figure ), a temperature decline meshes well with continental-scale cooling documented around 8200 cal. yr BP (variously attributed to the effect of a massive influx of meltwater on the heat-transferring North Atlantic Deep Water formation ([Rohling and Pälike Citation2005] or changes in solar output [Bond et al. Citation2001]). Although there is much large-scale evidence (particularly from ice cores) to support the cooling interpretation, it is also clear that competing climate signals (i.e. those suggesting warming, or, at least, only restricted cooling), particularly in the northern Rocky Mountains, are present in the various proxy records from this time as well (Grimm et al. Citation2001; Menounos et al. Citation2009). An increase of both chronological resolution and the number of sample sites representative of this interval would lend greater certainty to the interpretation.
The decreased thermal trend modelled is largely due to the signal derived from the abundance of cool-tolerant land snails (Discus whitneyi and Vallonia gracilicosta), both of which were very successful (with 80% and 83% of their maximum AV, respectively). These were the two most cool-preferring taxa assessed. However, because these species are on opposite ends of the precipitation spectrum, the moisture regime predicted by the snails is somewhat less clear. For each taxon assessed, the climate niche is mostly dictated by temperature; on the average niche calculations for all taxa were 69% established by temperature. Only 31% of the niche assignments were attributed to changes in precipitation. This uneven weighting is particularly evident in the case of Discus and Vallonia snails; temperature contributed about 80% and 70%, respectively. Other studies, particularly of gastropods, also suggest the increased weight of thermal variables on ecological niche development (e.g. Byers et al. Citation2013; Mumladze Citation2014; Tytar and Makarova Citation2015; although in cases where gastropod water demand is high, precipitation is increasingly important [Sarma et al. Citation2015]). Similarly, studies based on plant macrofossils have often been successful at predicting temperature, though the weak relationship to moisture and precipitation has presented a confounding variable to other researchers (e.g. Conolly and Dahl Citation1970; Thompson et al. Citation2008).
Given the weak ecological relationship of the taxa to precipitation, confidence in modelled hydrology is less than that of temperature. Despite this, the net precipitation signal, indicating an arid early to mid-Holocene period, generally agrees with reconstructions from the area. Lofty Lake (near Athabasca, Alberta, about 450 km northeast of the study area) was mostly dry and highly saline (resulting from a lower precipitation-to-evaporation balance) during the early Holocene (Schweger and Hickman Citation1989; Vance et al. Citation1995). Increased salinity was also evident by saline-indicating pollen and diatoms during this period at Goldeye Lake in the central Alberta foothills (Hickman and Schweger Citation1993). Toboggan Lake (located between the Tongue Creek and Jumpingpound Creek watersheds) showed increases in dry-condition-indicating pine pollen and charcoal accumulation; this indicates that this dry period also coincides with the local maximum Holocene fire frequency (Macdonald Citation1989). However, the calibrated precipitation data from the present study indicates that the decline in precipitation relative to the present was even greater in the study area than at Lofty Lake. Here, annual precipitation was about 82 mm less than the modern value, whereas Vance et al. (Citation1995), modelling climate from pollen data, indicated the difference near Athabasca was about 50 mm less between 8000 and 6000 14C years ago (ca. 9000 to 7000 cal. yr BP). It is possible that the severity of aridity during this time was more influential in determining ecological niches than it is today, thereby favoring xerophilic taxa, despite their temperature preferences. This may help explain the apparent temperature divergence of this study from the other local studies and accord better with the fire regime Macdonald (Citation1989) noted, assuming increased fire frequency results from concurrent decrease in precipitation and increase in temperature.
Mid-Holocene
The mid-Holocene interval (ca. 8000 to 5000 cal. yr BP) spans the later part of the Holocene Thermal Maximum (HTM; Renssen et al. Citation2012). The warming noted in the mid-Holocene (limited to the Northern Hemisphere) was probably driven by orbital forcing (Lorenz and Lohmann Citation2004; Renssen et al. Citation2012). However, Mount Mazama erupted about 7600 cal. yr BP (Zdanowicz et al. Citation1999; Egan et al. Citation2015), and spread ash across northwestern North America, including much of Alberta (Beaudoin and Oetelaar Citation2006). The introduction of large volumes of ash, particulates and aerosols into the atmosphere probably resulted in a cooling effect, at least in the short term (Zdanowicz et al. Citation1999; Beaudoin and Oetelaar Citation2006).
The effects of these influences are hard to separate, particularly from aggregated mid-Holocene-aged samples (collected from above and below the ash layer). In the study region, this period appears to have been substantially warmer and wetter than the Holocene average (Figure ). The increased moisture modelled largely agrees with the large-scale reconstruction Viau and Gajewski (Citation2009) developed from pollen data. Likewise, this is consistent with Jungerius’ (Citation1969) suggestion that the Cypress Hills region in southeast Alberta, based on vegetative cues in soil, probably experienced maximum Holocene humidity starting around the end of the mid-Holocene, 4500 14C yr BP (ca. 5100 cal. yr BP). However, the modelled increase in moisture contrasts with Macdonald’s (Citation1989) findings, which, based on continued occurrence of halophytic and marginal wet area plants, suggest continuing reductions of regional water availability and fluctuating water levels from 8400 years BP on into the mid-Holocene. Despite this incongruence, MacDonald’s (Citation1989) interpretation, which principally hinges on large accumulations of Chenopodiaceae and cattail (Typha latifolia) pollen, can also be ascribed to increased open areas following the prior fire regime and expanded (rather than contracted) lake fringes (i.e. providing increased lake perimeter habitat for the cattails). Following from the drier early- to mid-Holocene, most palaeoecological records from this region indicate the early mid-Holocene as an interval of prolonged aridity (e.g. Sauchyn and Sauchyn Citation1991). Marked increases in precipitation probably did not occur until the later part of this period.
The apparent moisture variability during the mid-Holocene is probably not only the effect of local reductions in precipitation noted in large areas of the Canadian prairies (Zoltai and Vitt Citation1990) and inherent spatial heterogeneity in meteorological patterns but also the confounding effect of increased meltwater contribution to montane watersheds. Considering the relationship between temperature and glacial melting and the abundance of alpine glaciers and icefields in the upper drainage basins of the study area, increased temperatures, particularly evident in the mid-Holocene, may have resulted in increased seasonal flow and, therefore, net riparian moisture (e.g. the combination of local precipitation and more distantly sourced streamflow). In this way, the modelled precipitation values may actually be more representative of ‘local moisture’, i.e. the net riparian water condition, instead of resulting from meteoric input to the site alone.
Streamflow and, by extension, flood frequency are conditioned by climate (Fleming et al. Citation2007). However, at least in the upper Bow River watershed, flood regime does not appear significantly correlated with changes in climate (Whitfield and Pomeroy Citation2016). In fact, instead of a uniform increase in flood severity with warming conditions, as is often thought to occur, Rood et al. (Citation2016) found a generally moderating effect of warming on flood regimes in the northern Rocky Mountains at large. Nevertheless, climatic changes and periodic meteorological cycles (e.g. the Pacific Decadal Oscillation, PDO) certainly impart noticeable influence on both flood frequency and severity (Gurrapu et al. Citation2016; Rood et al. Citation2016), though streamflow response to climate-derived changes in glacial sources is commonly parabolic in nature (Brahney et al. Citation2017). Short-term climatic fluctuations (such as the PDO) are probably averaged out in the detectible signal in this study. Despite potential changes to total stream flow in the mid-Holocene, the effect of variable flood frequency (and the associated burial of palaeoenvironmental layers) may have been negligible.
Though the absolute difference in temperature from the early Holocene to the mid-Holocene is difficult to assess, an increase was present (Figure ). The modern temperature of the study region (Figure ) is coldest in the western high-elevation areas. Gravity is a substantial driver of transport, particularly for taxa with small ranges (e.g. molluscs) or that are passively transported (e.g. seeds). As such, it would probably be easier for taxa to change their range in response to cooling temperatures (resulting in a down-slope shift) than when warming occurred (requiring up-slope migration), though the distance required to reach a desired temperature would be less in the latter case. Elevation-related range shifts (driven by temperature) have been documented throughout the Holocene (Beaudoin Citation1986; Macdonald Citation1989; Davis and Shaw Citation2001) and are a major component of faunal and floral responses to modern climate change (Walther et al. Citation2002; Lenoir et al. Citation2008).
Mid- to late Holocene
The mid- to late Holocene interval (ca. 6000 to 2000 cal. yr BP) showed a return to moisture and temperature regimes similar to modern ones (Figure ). Temperature appears to have stayed slightly higher than precipitation (relative to the modelled average) during this time, though both parameters decreased substantially from the mid-Holocene. The decline in moisture was most likely related to simultaneous reductions in summer meltwater contributions, declining glacial reserves (hydraulic mass preserved in ice), as well as changes in seasonal precipitation.
In terms of the indicator taxa analyzed, this was the most productive period, with seven of the maximum taxon AV values. These taxa included Helisoma anceps, the second-warmest-preferring taxon analyzed. MacDonald (Citation1982) also noted H. anceps as a ‘south-ranging’ species and defined it as an indicator of warm temperatures in the Kananaskis Valley, just north of the study area. Helisoma anceps is the only taxon in the assemblage whose niche is defined more by moisture regime (52%) than by temperature (48%).
This period also exhibited a proliferation of succineid snails, yielding the maximum apparent AV for this taxon. Though these snails are often considered ‘wetland taxa’ (Burch Citation1962; Forsyth Citation2004), their ranking in terms of modelled species preference reveals an inclination to rather drier conditions, as well as moderately warm temperatures. Galba modicella, the most thermophilic taxon, was also most productive at this time. The abundance of these snails may indicate a residual lag in response to the warm middle Holocene thermal regime. Additionally, the aquatic snails (Galba and Helisoma) may have benefitted marginally from thermal lagging of shallow groundwater inputs (e.g. advective flux) into their surface water habitats, where contributing groundwater temperature more closely reflects the previous – in this case, warmer – climate (see Caissie Citation2006; Kurylyk et al. Citation2015), though this effect was probably minimal over the course of several hundred years.
The biota from this time presents an even more complex image of the moisture regime. The macrofossils that are most prolific during this time are split approximately equally into dry- and wet-preferring groups. Considering the disparity of indicators, it is easy to imagine a regional biota in flux, under pressure to respond to changes in climate and associated hydrology. Additionally, substantial community lag may be present following climate change; biological changes are not always synchronous with climatic or other environmental changes (Cole Citation2010). Cole noted that in many cases, the response (particularly of vegetation) is lagged relative to the environment, such that communities are often more representative of the conditions prior to their lifetime than the conditions within which they lived. Nonetheless, the ecological responses of these taxa to hydrological conditions are far weaker than their relationships with temperature. Some of the apparent ‘noise’ detected from this signal may simply be plasticity within the taxa’s realized ecological niches.
Late Holocene and modern
The late Holocene, ca. 3000 cal. yr BP to present, saw regional climate even closer to the Holocene average (Figure ). Precipitation values were roughly equivalent to the mean, and temperature was slightly cooler than average (4% below). This temperature decline probably reflects the onset of Neoglacial conditions, typified by low summer temperatures (Denton and Karlén Citation1973). In the Alberta Rockies, the onset of Neoglacial conditions is marked by glacier advances around 4000 14C yr BP (Luckman et al. Citation1993), ca. 4500 cal. yr BP. Regionally, late Holocene climate is modelled as rather stable, although other studies reveal notable temperature variance within this period (Gajewski and Viau Citation2011). Major late Holocene fluctuations include intervals of warmer (e.g. Medieval Warm Period) and cooler (e.g. Little Ice Age) climate (Edwards et al. Citation2008). Considering these oscillations in the light of the smoothing effects of biotic response lag, the rather uniform late Holocene temperature signal (Figure ) is not unexpected. That is, short-term climate/biota equilibrium was not reached during this period; the biological response to variation was too slow to be noticeable when integrated at this coarse time scale.
Late Holocene samples exhibited considerable riparian biodiversity, though this may be influenced by sampling or preservation bias. Indicator taxa were nearly half as abundant per sample as they were in either the preceding mid- to late Holocene or the subsequent modern periods. The most numerous taxa included Discus whitneyi, the marginal aquatic plant Eleocharis palustris, wet-condition-obligate aquatic snail Helisoma anceps, and the aquatic plant genera Potamogeton and Zannichellia. Despite the apparent preference towards wetter conditions, these plants are actually rather tolerant of dry climates so long as their local habitat remains moist. In fact, Eleocharis palustris and Zannichellia palustris have two of the lowest-precipitation climatic optima of the indicators assessed.
The shift into modern times (with samples yielding seeds from recently introduced plants) brought with it a return to high indicator taxon AV values. Only two species, Euconulus fulvus and Helisoma anceps, were absent from this age class. In these most recent samples, taphonomic processes of degradation have not been able to act extensively on the samples, and as such they probably provide a more complete view of the climate and local environments. Six of the 15 indicators reached their maximum AV during this time as well. These included taxa which exhibited a multitude of precipitation preferences (from the dry-tolerant land snail Vallonia gracilicosta to the high-moisture-preferring plant Scirpus microcarpus).
The modern period had temperature and precipitation values which were about 4% less than the Holocene average (Figure ). This agrees with the regional model produced by Viau and Gajewski (Citation2009), which showed a decline in both parameters during the past thousand years. However, their results indicate a return to wetter conditions in central Canada more recently. This was not detected in the present study, perhaps because of ecological lags in biotic adjustment. In order to understand the present effects of recent climate change, scientists may need to wait for a century or more to fully discern the effect on biological communities in the area.
Conclusions
Collecting newly exposed sediments following major flooding near Calgary, Alberta, provided a unique opportunity to understand the responses of riparian and aquatic communities to changes in climate. This study has leveraged the faunal and floral macrofossil remains from 36 palaeoenvironmental samples collected from the banks of four streams in southwestern Alberta to generate a new postglacial regional interpretation of annual precipitation and temperature. The modelling approach used here is novel and has not hitherto been applied to macrofossil samples in western Canada.
Following the warming which signalled the onset of the Holocene, temperature and precipitation are modelled to have fallen again in the early to mid-Holocene. Of particular note was the subsequent apparent increase in riparian moisture which probably accompanied (or followed) Holocene-maximum temperatures. From the mid-Holocene until modern times, both temperature and moisture regimes are modelled as having stabilized at a lower level.
In a broad sense, the results of this study agree well with previous work conducted in the area. The results suggest an overarching regional signal, overprinted with local variation derived from the particular landscape setting investigated. The study highlights the fact that different proxies encode different aspects of the climate system. In addition, the riparian and fluvial focus of this study reveals a unique subset of regional climatic conditions particular to the near-stream environment. The challenge of understanding bioclimatic responses to temperature and moisture, particularly in a fluvial context, is also highlighted by this work. This study is the first formal step towards understanding the relationships of riparian flora and fauna with the environment at a regional scale on the post-glacial Alberta landscape.
Acknowledgements
We thank Darryl Bereziuk, Director, and Wendy Unfreed, Regional Archaeologist, Archaeological Survey of Alberta, for support and encouragement during this study. We also thank the consultants involved in the river surveys and in collecting the samples, especially Dr. Lisa Bohach and Emily Frampton (Stantec Consulting Ltd.). Thanks also to three anonymous reviewers whose input greatly improved the tone and clarity of this paper. This project was funded in part by the Southern Alberta Flood Investigation Program, administered by the Historic Resources Management Branch of Alberta Culture and Tourism, under the direction of Matthew Wangler, Executive Director.
References
- Aarnes, I., N. Kühl, and H. H. Birks. 2012. Quantitative climate reconstruction from late-glacial and early Holocene plant macrofossils in western Norway using the probability density function approach. Review of Palaeobotany and Palynology 170: 27–39. doi:10.1016/j.revpalbo.2011.11.001.
- Alçiçek, H., F. P. Wesselingh, M. C. Alçiçek, G. Jiménez-Moreno, F. J. Feijen, L. W. H. Ostende, S. Mayda, and A. S. Tesakov. 2017. A multiproxy study of the early Pleistocene palaeoenvironmental and palaeoclimatic conditions of an anastomosed fluvial sequence from the Çameli Basin (SW Anatolia, Turkey). Palaeogeography, Palaeoclimatology, Palaeoecology 467: 232–252.
- Austin, M. P. 1987. Models for the analysis of species’ response to environmental gradients. Vegetatio 69 (1–3): 35–45.10.1007/BF00038685
- Austin, M. P. 2002. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecological Modelling 157 (2–3): 101–118.10.1016/S0304-3800(02)00205-3
- Bartlein, P. J., S. P. Harrison, S. Brewer, S. Connor, B. A. S. Davis, K. Gajewski, J. Guiot, et al. 2011. Pollen-based continental climate reconstructions at 6 and 21 ka: A global synthesis. Climate Dynamics 37 (3): 775–802. doi:10.1007/s00382-010-0904-1.
- Baxter, A. J. 1997. Late Quaternary palaeoenvironments of the Sandveld, Western Cape Province, South Africa. PhD thesis, Department of Environmental and Geographical Science, University of Cape Town, 355 pp.
- Beaudoin, A. B. 1986. Using Picea/Pinus ratios from the Wilcox Pass core, Jasper National Park, Alberta, to investigate Holocene timberline fluctuations. Géographie physique et Quaternaire 40: 145–152.10.7202/032634ar
- Beaudoin, A. B. 2007. On the laboratory procedure for processing unconsolidated sediment samples to concentrate subfossil seed and other plant macroremains. Journal of Paleolimnology 37 : 301–308.10.1007/s10933-006-9031-8
- Beaudoin, A. B., and G. A. Oetelaar. 2006. The day the dry snow fell: The record of a 7627-year-old disaster. In Alberta Formed Alberta Transformed, Volume 1, eds. Michael Payne, Don Wetherell and Catherine Kavanaugh, 36–53. Alberta: University of Alberta Press and University of Calgary Press.
- Berg, K. J., G. M. Samuelson, C. R. Willms, D. W. Pearce, and S. B. Rood. 2007. Consistent growth of black cottonwoods despite temperature variation across elevational ecoregions in the Rocky Mountains. Trees 21 (2): 161–169.
- Beierle, B. D., and D. G. Smith. 1998. Severe drought in the early Holocene (10,000-6800 BP) interpreted from lake sediment cores, southwestern Alberta, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 75–83.10.1016/S0031-0182(98)00044-3
- Birks, H. J. B., O. Heiri, H. Seppä, and A. E. Bjune. 2010. Strengths and weaknesses of quantitative climate reconstructions based on Late-Quaternary biological proxies. The Open Ecology Journal 3: 68–110.
- Bohach, L. L. 2016. Historical resources impact assessment for Palaeontology: Flood impact assessment program 2015, Kananaskis River, Jumpingpound Creek and Tongue Creek. Stantec Consulting Ltd., Calgary, Alberta. Edmonton, Alberta, Canada: Archaeological Survey of Alberta.
- Bohach, L. L., and E. Frampton. 2015. Historical resources impact assessment for palaeontology, flood impact assessment program 2014, Highwood River. Stantec Consulting Ltd., Calgary, Alberta. Edmonton, Alberta, Canada: Archaeological Survey of Alberta.
- Bolton, M. S. 2016. Palaeoenvironmental analysis of flood samples collected from historical resources impact assessment for palaeontology, Highwood River, 2014. Royal Alberta Museum, Edmonton, Alberta. Edmonton: Archaeological Survey of Alberta and Royal Alberta Museum, 67 pp.
- Bolton, M. S. 2017. Palaeoenvironmental analysis of Jumpingpound Creek, Kananaskis River, and Tongue Creek post-flood microfaunal samples. Royal Alberta Museum, Edmonton, Alberta. Edmonton: Archaeological Survey of Alberta and Royal Alberta Museum, 70 pp.
- Bolton, M. S., and A. B. Beaudoin. 2016. Reconstructing the Highwood River’s Holocene palaeoecology: Assessment of organic-rich alluvial samples collected following the 2013 flood event. In Back on the horse: Recent developments in archaeological and palaeontological research in Alberta, ed. R. Woywitka, 161–168. Edmonton: Archaeological Survey of Alberta.
- Bond, J. E. 2012. Phylogenetic treatment and taxonomic revision of the trapdoor spider genus Aptostichus Simon (Araneae, Mygalomorphae, Euctenizidae). Zoo Keys 252: 1–209. http://zookeys.pensoft.net/articles.php?id=3454 (accessed December 2016).
- Bond, G., B. Kromer, J. Beer, R. Muscheler, M. N. Evans, W. Showers, S. Hoffmann, R. Lotti-Bond, I. Hajdas, and G. Bonani. 2001. Persistent solar influence on North Atlantic climate during the Holocene. Science 294 (5549): 2130–2136.
- Bongers, F., L. Poorter, R. S. A. R. Rompaey, and M. P. E. Parren. 1999. Distribution of twelve moist forest canopy tree species in Liberia and Cote d’Ivoire: Response curves to a climatic gradient. Journal of Vegetation Science 10 (3): 371–382.10.2307/3237066
- Brahney, J., F. Weber, V. Foord, J. Janmaat, and P. J. Curtis. 2017. Evidence for a climate-driven hydrologic regime shift in the Canadian Columbia Basin. Canadian Water Resources Journal: 1–14. (accessed March 2017. doi:10.1080/07011784.2016.1268933.
- Bronk Ramsey, C. 2009. Bayesian analysis of radiocarbon dates. Radiocarbon 51 (1): 337–360.
- Brown, K. J., and G. B. Pasternack. 2005. A palaeoenvironmental reconstruction to aid in the restoration of floodplain and wetland habitat on an upper deltaic plain, California, USA. Environmental Conservation 32 (2): 103–166.10.1017/S037689290500216X
- Burch, J. B. 1962. How to know the eastern land snails. Pictured-key nature series. Dubuque: William C. Brown, 214 pp.
- Burn, C. R. 1997. Cryostratigraphy, paleogeography, and climate change during the early Holocene warm interval, western Arctic coast, Canada. Canadian Journal of Earth Sciences 34 (7): 912–925.10.1139/e17-076
- Burnham, R. J. 1993. Time resolution in terrestrial macrofloras: Guidelines from modern accumulations. In Taphonomic approaches to time resolution in the fossil record, eds. S. M. Kidwell and A. K. Behrensmeyer, 57–78. Knoxville, TN: Paleontological Society.
- Byers, J. E., W. G. McDowell, S. R. Dodd, R. S. Haynie, L. M. Pintor, and S. B. Wilde. 2013. Climate and pH predict the potential range of the invasive apple snail (Pomacea insularum) in the southeastern United States. PLoS ONE 8 (2): e56812.10.1371/journal.pone.0056812
- Caissie, D. 2006. The thermal regime of rivers: A review. Freshwater Biology 51 (8): 1389–1406.10.1111/fwb.2006.51.issue-8
- Clarke, A. H. 1981. The freshwater molluscs of Canada. Ottawa: National Museum of Natural Sciences, National Museums of Canada, 446 pp.
- Cole, K. L. 2010. Vegetation response to early Holocene warming as an analog for current and future changes. Conservation Biology 24 (1): 29–37.10.1111/cbi.2010.24.issue-1
- Conolly, A. P., and E. Dahl. 1970. Maximum summer temperature in relation to the modern and Quaternary distributions of certain arctic-montane species in the British Isles. In Studies in the vegetational history of the British Isles: Essays in honour of Harry Godwin, eds. D. Walker and R. G. West, 159–223. Cambridge: Cambridge University Press.
- Cooper, R. A., P. A. Maxwell, J. S. Crampton, A. G. Beu, C. M. Jones, and B. A. Marshall. 2006. Completeness of the fossil record: Estimating losses due to small body size. Geology 34 (4): 241–244.
- Cropper, J. P. 1984. Multicollinearity within selected western North American temperature and precipitation data sets. Tree-Ring Bulletin 44: 29–37.
- Daly, C., M. Halbleib, J. I. Smith, W. P. Gibson, M. K. Doggett, G. H. Taylor, J. Curtis, and P. P. Pasteris. 2008. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. International Journal of Climatology 28 (15): 2031–2064.
- Davis, B. A. S., S. Brewer, A. C. Stevenson, and J. Guiot. 2003. The temperature of Europe during the Holocene reconstructed from pollen data. Quaternary Science Reviews 11 (15): 1701–1716.
- Davis, M. B., and R. G. Shaw. 2001. Range shifts and adaptive responses to Quaternary climate change. Science 292 (5517): 673–679.10.1126/science.292.5517.673
- Denton, G. H., and W. Karlén. 1973. Holocene climatic variations – their pattern and possible cause. Quaternary Research 3 (2): 155–205.10.1016/0033-5894(73)90040-9
- Dyke, A. S. 2004. An outline of North American deglaciation with emphasis on central and northern Canada. In Quaternary glaciations—extent and chronology: Part II: North America, eds. J. Ehlers and P. L. Gibbard, 371–406. Amsterdam: Elsevier.
- Dyke, A. S. 2005. Late Quaternary vegetation history of northern North America based on pollen, macrofossil, and faunal remains. Géographie physique et Quaternaire 59 (2–3): 211–262.10.7202/014755ar
- Edwards, T., S. J. Birks, B. H. Luckman, and G. M. MacDonald. 2008. Climatic and hydrologic variability during the past millennium in the eastern Rocky Mountains and northern Great Plains of western Canada. Quaternary Research 70 (2): 188–197.10.1016/j.yqres.2008.04.013
- Egan, J., R. Staff, and J. Blackford. 2015. A revised age estimate of the Holocene Plinian eruption of Mount Mazama, Oregon using Bayesian statistical modelling. The Holocene 25 (7): 1054–1067.10.1177/0959683615576230
- Ejarque, A., A. Beauger, Y. Miras, J. -L. Peiry, O. Voldoire, F. Vautier, M. Benbakkar, and J. Steiger. 2015. Historical fluvial palaeodynamics and multi-proxy palaeoenvironmental analyses of a palaeochannel, Allier River, France. Geodinamica Acta 27 (1): 25–47.10.1080/09853111.2013.877232
- Elith, J., S. J. Phillips, T. Hastie, M. Dudík, Y. E. Chee, and C. J. Yates. 2011. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions 17 (1): 43–57.10.1111/ddi.2010.17.issue-1
- Environment and Climate Change Canada. 2016. Canadian Climate Normals: 1981-2010 Climate Normals and Averages. http://climate.weather.gc.ca/climate_normals/index_e.html (accessed December 2016).
- Fleming, S. W., P. H. Whitfield, R. D. Moore, and E. J. Quilty. 2007. Regime‐dependent streamflow sensitivities to Pacific climate modes cross the Georgia–Puget transboundary ecoregion. Hydrological Processes 21 (24): 3264–3287.
- Forsyth, Robert G. 2004. Land snails of British Columbia. Victoria: Royal British Columbia Museum, 188 pp.
- Forsyth, Robert G. 2006. An annotated checklist (based mostly on literature records) and bibliography of the recent terrestrial Mollusca of Alberta. http://www.mollus.ca/canada/checklists/Alberta_Checklist.pdf (accessed December 2016).
- Fourcade, Y., J. O. Engler, D. Rödder, and J. Secondi. 2014. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 9 (5): e97122.10.1371/journal.pone.0097122
- Gajewski, Konrad, and Andre E. Viau. 2011. Abrupt climate changes during the Holocene across North America from pollen and paleolimnological records. In Abrupt climate change: Mechanisms, patterns, and impacts, eds. Harunur Rashid, Leonid Polyak, and Ellen Mosley-Thompson, 173–183. Washington, DC: American Geophysical Union.
- GBIF. 2016. Global biodiversity information facility. http://www.gbif.org/ (accessed December 2016).
- Gell, P. 2010. With the benefit of hindsight: The utility of palaeoecology in wetland condition assessment and identification of restoration targets. In Ecology of industrial pollution, eds. L. C. Batty and K. B. Hallberg, 162–188. Cambridge, England, UK: Cambridge University Press.10.1017/CBO9780511805561
- Gell, P., J. Fluin, J. Tibby, G. Hancock, J. Harrison, A. Zawadzki, D. Hynes, S. Khanum, F. Little, and B. Walsh. 2009. Anthropogenic acceleration of sediment accretion in lowland floodplain wetlands, Murray-Darling Basin, Australia. Geomorphology 108 (1–2): 122–126.10.1016/j.geomorph.2007.12.020
- Goodale, C. L., J. D. Aber, and S. V. Ollinger. 1998. Mapping monthly precipitation, temperature, and solar radiation for Ireland with polynomial regression and a digital elevation model. Climate Research 10 (1): 35–49.
- Gottesfeld, A. S. 1996. British Columbia flood scars: Maximum flood-stage indicators. Geomorphology 14 (4): 319–325.10.1016/0169-555X(95)00045-7
- Grimm, E. C., S. Lozano-Garciá, H. Behling, and V. Markgraf. 2001. Holocene vegetation and climate variability in the Americas. In Interhemispheric climate linkages, ed. V. Markgraf, 325–370. San Diego, CA: Academic Press.10.1016/B978-012472670-3/50022-7
- Gurrapu, S., J. St-Jacques, D. J. Sauchyn, and K. R. Hodder. 2016. The influence of the Pacific Decadal Oscillation on annual floods in the rivers of Western Canada. JAWRA Journal of the American Water Resources Association 52 (5): 1031–1045.10.1111/jawr.2016.52.issue-5
- Herald, Calgary. 2013. The flood of 2013: A summer of angry rivers in Southern Alberta. Vancouver: Greystone Books, 136 pp.
- Hickman, M., and C. E. Schweger. 1993. Late glacial-early Holocene paleosalinity in Alberta, Canada - climate implications. Journal of Paleolimnology 8: 149–161.
- Hijmans, R. J., S. E. Cameron, J. L. Parra, P. G. Jones, and A. Jarvis. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25 (15): 1965–1978.10.1002/(ISSN)1097-0088
- Horsák, M. 2006. Mollusc community patterns and species response curves along a mineral richness gradient: A case study in fens. Journal of Biogeography 33 (1): 98–107.10.1111/jbi.2006.33.issue-1
- Hua, Q., M. Barbetti, and A. J. Rakowski. 2013. Atmospheric radiocarbon for the period 1950–2010. Radiocarbon 55 (4): 2059–2072.
- Hupp, C. R., and W. R. Osterkamp. 1996. Riparian vegetation and fluvial geomorphic processes. Geomorphology 14 (4): 277–295.
- Jost, A., D. Lunt, M. Kageyama, A. Abe-Ouchi, O. Peyron, P. J. Valdes, and G. Ramstein. 2005. High-resolution simulations of the last glacial maximum climate over Europe: A solution to discrepancies with continental palaeoclimatic reconstructions? Climate Dynamics 24 (6): 577–590. doi:10.1007/s00382-005-0009-4.
- Jungerius, P. D. 1969. Soil evidence of postglacial tree line fluctuations in the Cypress Hills area, Alberta, Canada. Arctic and Alpine Research 1 (4): 235–245.10.2307/1550112
- Katz, G. L., J. M. Friedman, and S. W. Beatty. 2005. Delayed effects of flood control on a flood-dependent riparian forest. Ecological Applications 15 (3): 1019–1035.10.1890/04-0076
- Kidwell, S. M., and D. W. J. Bosence. 1991. Taphonomy and time-averaging of marine shelly faunas. In Taphonomy: releasing the data locked in the fossil record: Topics in geobiology. vol. 9, eds. P.A. Allison and D.E.G. Briggs, 115–209. New York: Plenum Press.
- Kottek, M., J. Grieser, C. Beck, B. Rudolf, and F. Rubel. 2006. World map of the Köppen-Geiger climate classification updated. Meteorologische Zeitschrift 15 (3): 596–263.
- Kozlowski, T. T. 2002. Physiological-ecological impacts of flooding on riparian forest ecosystems. Wetlands 22 (3): 550–561.10.1672/0277-5212(2002)022[0550:PEIOFO]2.0.CO;2
- Kurylyk, B. L., K. T. B. MacQuarrie, D. Caissie, and J. M. McKenzie. 2015. Shallow groundwater thermal sensitivity to climate change and land cover disturbances: Derivation of analytical expressions and implications for stream temperature modeling. Hydrology and Earth System Sciences 19 (5): 2469–2489.10.5194/hess-19-2469-2015
- Lenoir, J., J. Gégout, P. A. Marquet, P. De Ruffray, and H. Brisse. 2008. A significant upward shift in plant species optimum elevation during the 20th century. Science 320 (5884): 1768–1771.10.1126/science.1156831
- Leopold, L. B. 1994. A view of the river. Cambridge, Massachusetts: Harvard University Press, 320 pp.
- Lorenz, S. J., and G. Lohmann. 2004. Acceleration technique for Milankovitch type forcing in a coupled atmosphere-ocean circulation model: Method and application for the Holocene. Climate Dynamics 23 (7–8): 727–743.10.1007/s00382-004-0469-y
- Lorenz, D. J., D. Nieto-Lugilde, J. L. Blois, M. C. Fitzpatrick, and J. W. Williams. 2016. Downscaled and debiased climate simulations for North America from 21,000 years ago to 2100AD. Scientific Data 3: 160048. http://www.nature.com/articles/sdata201648 (accessed December 2016).10.1038/sdata.2016.48
- Luckman, B. H., G. Holdsworth, and G. D. Osborn. 1993. Neoglacial glacial fluctuations in the Canadian Rockies. Quaternary Research 39 (2): 144–153.10.1006/qres.1993.1018
- Luckman, B. H., K. R. Briffa, P. D. Jones, and F. H. Schweingruber. 1997. Tree-ring based reconstruction of summer temperatures at the Columbia Icefield, Alberta, Canada, A.D. 1073–1983. The Holocene 7 (4): 375–389.
- MacDonald, G. M. 1982. Late Quaternary paleoenvironments of the Morley flats and Kananaskis valley of southwestern Alberta. Canadian Journal of Earth Sciences 19: 23–35.10.1139/e82-003
- MacDonald, G. M. 1989. Postglacial palaeoecology of the subalpine forest — grassland ecotone of southwestern Alberta: New insights on vegetation and climate change in the Canadian rocky mountains and adjacent foothills. Palaeogeography, Palaeoclimatology, Palaeoecology 73: 155–173.10.1016/0031-0182(89)90001-1
- MacDonald, G. M., R. P. Beukens, W. E. Kieser, and D. H. Vitt. 1987. Comparative radiocarbon dating of terrestrial plant macrofossils and aquatic moss from the "ice-free corridor" of western Canada. Geology 15: 837–840.
- Marmi, J., C. Martín-Closas, M. T. Fernández-Marrón, V. Fondevilla, and B. Gomez. 2015. A riparian plant community from the upper Maastrichtian of the Pyrenees (Catalonia, NE Spain). Cretaceous Research 56: 510–529.10.1016/j.cretres.2015.06.004
- Menounos, B., G. Osborn, J. J. Clague, and B. H. Luckman. 2009. Latest Pleistocene and Holocene glacier fluctuations in western Canada. Quaternary Science Reviews 28 (21–22): 2049–2074.10.1016/j.quascirev.2008.10.018
- Merow, C., M. J. Smith, and J. A. Silander. 2013. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 36 (10): 1058–1069.10.1111/ecog.2013.36.issue-10
- Milrad, S. M., J. R. Gyakum, and E. H. Atallah. 2015. A meteorological analysis of the 2013 Alberta flood: Antecedent large-scale flow pattern and synoptic-dynamic characteristics. Monthly Weather Review 143 (7): 2817–2841.10.1175/MWR-D-14-00236.1
- Montgomery, F. H. 1977. Seeds and fruits of plants of eastern Canada and northeastern United States Toronto: University of Toronto Press, 232 pp.
- Mumladze, L. 2014. Sympatry without co-occurrence: Exploring the pattern of distribution of two Helix species in Georgia using an ecological niche modelling approach. Journal of Molluscan Studies 80 (3): 249–255.10.1093/mollus/eyu045
- Musselman, K. N., J. W. Pomeroy, R. L. H. Essery, and N. Leroux. 2015. Impact of windflow calculations on simulations of alpine snow accumulation, redistribution and ablation. Hydrological Processes 29: 3983–3999.
- Oswald, W. W., P. M. Anderson, T. A. Brown, L. B. Brubaker, F. S. Hu, A. V. Lozhkin, W. Tinner, and P. Kaltenrieder. 2005. Effects of sample mass and macrofossil type on radiocarbon dating of arctic and boreal lake sediments. The Holocene 15 (5): 758–767.
- Pennock, D. J. 1984. Soil landscape evolution in the Highwood River basin, Southern Alberta. PhD thesis, Queen's University.
- Phillips, S. J., M. Dudík, and R. E. Schapire. 2004. A maximum entropy approach to species distribution modeling. In Proceedings of the twenty-first international conference on machine learning. Banff, Alberta, July 4-8, 2004, p. 83.
- Phillips, S. J., R. P. Anderson, and R. E. Schapire. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190 (3–4): 231–259.10.1016/j.ecolmodel.2005.03.026
- Pomeroy, J. W., R. E. Stewart, and P. H. Whitfield. 2015. The 2013 flood event in the South Saskatchewan and Elk River basins: Causes, assessment and damages. Canadian Water Resources Journal 41 (1–2): 105–117.
- Reimer, P. J., E. Bard, A. Bayliss, J. W. Beck, P. G. Blackwell, C. Bronk Ramsey, C. E. Buck, et al. 2013. INTCAL13 and MARINE13 radiocarbon calibration curves 0–50,000 years cal BP. Radiocarbon 55 (4): 1869–1887.
- Renssen, H., H. Seppä, X. Crosta, Hugues Goosse, and D. M. Roche. 2012. Global characterization of the Holocene thermal maximum. Quaternary Science Reviews 48: 7–19.10.1016/j.quascirev.2012.05.022
- Rippke, M. B., M. T. Distler, and J. M. Farrell. 2010. Holocene vegetation dynamics of an upper St. Lawrence River wetland: Paleoecological evidence for a recent increase in cattail (Typha). Wetlands 30 (4): 805–816.
- Rohling, E. J., and H. Pälike. 2005. Centennial-scale climate cooling with a sudden cold event around 8,200 years ago. Nature 434 (7036): 975–979.10.1038/nature03421
- Rood, S. B., S. G. Foster, E. J. Hillman, A. Luek, and K. P. Zanewich. 2016. Flood moderation: Declining peak flows along some Rocky Mountain rivers and the underlying mechanism. Journal of Hydrology 536: 174–182.10.1016/j.jhydrol.2016.02.043
- Rydgren, K., R.Halvorsen Økland, and T. Økland. 2003. Species response curves along environmental gradients: A case study from SE Norwegian swamp forests. Journal of Vegetation Science 14 (6): 869–880.10.1111/j.1654-1103.2003.tb02220.x
- Sarma, R. R., M. Munsi, and A. N. Ananthram. 2015. Effect of climate change on invasion risk of giant African snail (Achatina fulica Ferussac, 1821: Achatinidae) in India. PLoS ONE 10 (11): e0143724.10.1371/journal.pone.0143724
- Sauchyn, M. A., and D. J. Sauchyn. 1991. A continuous record of Holocene pollen from Harris Lake, southwestern Saskatchewan, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology 88: 13–23.10.1016/0031-0182(91)90012-G
- Schweger, C. E., and M. Hickman. 1989. Holocene paleohydrology of central Alberta: Testing the general-circulation-model climate simulations. Canadian Journal of Earth Sciences 26 (9): 1826–1833.10.1139/e89-155
- Servat, G. P., T. P. Feria, N. Hurtado, W. Mendoza, and R. F. Alcocer. 2013. Potential distribution and habitat characterization of Atlapetes melanopsis (Aves: Emberizidae) in a montane forest ecotone of the Apurímac river valley. In Monitoring biodiversity: Lessons from a Trans-Andean megaproject, eds. A. Alonso, F. Dallmeier and G. P. Servat, 141–153. Washington, DC, USA: Smithsonian Institution Scholarly Press.
- Sevruk, B. 1986. Correction of precipitation measurements: Swiss experience. Zürcher Geographische Schriften 23: 187–196.
- Simpson, J. J., G. L. Hufford, C. Daly, J. S. Berg, and M. D. Fleming. 2005. Comparing maps of mean monthly surface temperature and precipitation for Alaska and adjacent areas of Canada produced by two different methods. Arctic 58 (2): 137–161.
- Smith, D. G. 1979. Effects of channel enlargement by river ice processes on bankfull discharge in Alberta, Canada. Water Resources Research 15 (2): 469–475.
- Stene, L. P. 1980. Observations on lateral and overbank deposition—evidence from Holocene terraces, southwestern Alberta. Geology 8 (7): 314–317.
- Ter Braak, C. J. F., and C. W. N. Looman. 1986. Weighted averaging, logistic regression and the Gaussian response model. Vegetatio 65 (1): 3–11.10.1007/BF00032121
- Thomasson, J. R. 1991. Sediment-borne “seeds” from Sand Creek, northwestern Kansas: Taphonomic significance and paleoecological and paleoenvironmental implications. Palaeogeography, Palaeoclimatology, Palaeoecology 85 (3–4): 213–225.10.1016/0031-0182(91)90161-J
- Thompson, R. S., K. H. Anderson, and P. J. Bartlein. 2008. Quantitative estimation of bioclimatic parameters from presence/absence vegetation data in North America by the modern analog technique. Quaternary Science Reviews 27 (11–12): 1234–1254.10.1016/j.quascirev.2008.02.014
- Thornbush, M. J., and J. R. Desloges. 2011. Palaeoenvironmental reconstruction at an oxbow lake situated at the lower Nottawasaga River, southern Ontario, Canada. Environmental Archaeology 16 (1): 1–15.10.1179/146141010X12640787648216
- Trewin, N., S. R. Fayers, L. I. Anderson. 2004. Charophytes. The biota of early terrestrial ecosystems: The Rhynie Chert. University of Aberdeen, Aberdeen. https://www.abdn.ac.uk/rhynie/charo.htm (accessed December 2016).
- Trzaska, S., and E. Schnarr. 2014. A review of downscaling methods for climate change projections. Report for United States Agency for International Development by Tetra Tech ARD. https://www.climatelinks.org/resources/review-downscaling-methods-climate-change-projections (accessed May 2017).
- Tytar, V., and N. Makarova. 2015. Distribution of the freshwater snail species Fagotia (Gastropoda, Melanopsidae) in Ukraine according to climatic factors. I. Fagotia esperi. Vestnik zoologii 49 (4): 341–350.
- Vance, R. E., A. B. Beaudoin, and B. H. Luckman. 1995. The paleoecological record of 6 ka BP climate in the Canadian Prairie Provinces. Géographie physique et Quaternaire 49: 81–98.10.7202/033031ar
- Viau, A. E., and K. Gajewski. 2009. Reconstructing millennial-scale, regional paleoclimates of boreal Canada during the Holocene. Journal of Climate 22 (2): 316–330.10.1175/2008JCLI2342.1
- Walther, G., E. Post, P. Convey, A. Menzel, C. Parmesan, T. J. C. Beebee, J. Fromentin, O. Hoegh-Guldberg, and F. Bairlein. 2002. Ecological responses to recent climate change. Nature 416 (6879): 389–395.10.1038/416389a
- Ward, P., R. B. Hassen, A. Yassien, and S. J. Burges. 2010. Flood flows and channel conveyance in the Athabasca River Delta, Canada. International Journal of River Basin Management 8 (2): 161–172.
- Whitfield, P. H., and J. W. Pomeroy. 2016. Changes to flood peaks of a mountain river: Implications for analysis of the 2013 flood in the Upper Bow River, Canada. Hydrological Processes 30 (25): 4657–4673.10.1002/hyp.v30.25
- Wilson, M. C. 1974. Fossil bison and artifacts from the Mona Lisa site, Calgary, Alberta. Part I: Stratigraphy and artifacts. Plains Anthropologist 19: 34–45.
- Wilson, M. C. 1993. Radiocarbon dating in the Ice-free Corridor: Problems and implications. In The palliser triangle: A region in time and space, eds. R.W. Barendregt, M.C. Wilson and F.J. Jankunis, 165–206. Lethbridge: University of Lethbridge.
- Work, P. T., H. A. Semken, and R. G. Baker. 2005. Pollen, plant macrofossils and microvertebrates from mid-Holocene alluvium in east-central Iowa, USA: Comparative taphonomy and paleoecology. Palaeogeography, Palaeoclimatology, Palaeoecology 223 (3–4): 204–221.10.1016/j.palaeo.2005.04.005
- WorldClim. n.d. Global climate data version 1.4. WorldClim – Global Climate Data. http://www.worldclim.org/current (accessed December 2016).
- Zdanowicz, C. M., G. A. Zielinski, and M. S. Germani. 1999. Mount Mazama eruption: Calendrical age verified and atmospheric impact assessed. Geology 27: 621–624.10.1130/0091-7613(1999)027<0621:MMECAV>2.3.CO;2
- Zoltai, S. C., and D. H. Vitt. 1990. Holocene climatic change and the distribution of peatlands in western interior Canada. Quaternary Research 33: 231–240.10.1016/0033-5894(90)90021-C