Abstract
Unfavourable environmental conditions for crown rust [Puccinia coronata f. sp. avenae] during 2007–2009 resulted in light incidence of crown rust on oat (Avena sativa) in Manitoba and eastern Saskatchewan. The first appearance of crown rust on 11 August in 2008 and on 15 August in 2009 was the latest ever seen in this region over the past three decades. Using 19 oat crown rust differentials, a large number of races were identified from isolates from wild oat each year, and a large proportion of the races were represented by a single isolate. There were significant differences between isolates from wild oat and cultivated oat in frequency of virulence to several genes in some years. Virulence frequency to Pc48, a gene in ‘Triple Crown’, was between 8.2–14.1% in isolates from wild oat and between 11.5–18.2% in isolates from cultivated oat during 2007–2009. Frequency of virulence to Pc68 was between 42.3–45.9% and 70.8–81.8% in isolates from wild oat and cultivated oat, respectively. As cultivars with the Pc38, 39, 68-gene combination were still commonly grown during these years, races with virulence to this gene combination were abundant. These cultivars were gradually replaced by new cultivars with different resistance genes. By 2009, ‘Leggett’ (Pc68, 94) accounted for 17.9% of the total oat hectarage, and ‘HiFi’ (Pc91) accounted for 3.0%. During 2007–2009, virulence to Pc94 was low (≤ 1.8%) or not detected, virulence to Pc91 was found in a single isolate, and virulence to gene temp_pc97 or temp_Pc98 was either low or not detected. The huge increase in frequency of virulence to Pc45 in 2008 and 2009 was most likely a result of isolates with this virulence migrating into the prairie region from the USA.
Résumé
De 2007 à 2009, des conditions adverses à la rouille couronnée (Puccinia coronata f. sp. avenae) ont fait que l'incidence de cette maladie sur l'avoine (Avena sativa) a été plutôt faible au Manitoba dans l'est de la Saskatchewan. Les premières occurrences de la rouille couronnée, le 11 août 2008 et le 15 août 2009, étaient les plus tardives jamais enregistrées dans cette région en 30 ans. À partir de 19 lignées différentielles de rouille couronnée, plusieurs races ont été identifiées chaque année grâce à des isolats de folle avoine, et une forte proportion des races était représentée par un isolat unique. Selon les années, il y avait des différences significatives entre les isolats de folle avoine et d'avoine quant à la fréquence de la virulence à l'égard de plusieurs gènes. De 2007 à 2009, la fréquence de la virulence à l'égard du gène Pc48, un gène du cultivar ‘Triple Crown’, variait de 8.2 % à 14.1 % chez des isolats de folle avoine, et de 11.5 % à 18.2 % chez des isolats d'avoine. La fréquence de la virulence à l’égard du gène Pc68 variait de 42.3 % à 45.9 % et de 70.8 % à 81.8 % chez les isolats de folle avoine et d'avoine, respectivement. Étant donné que les cultivars possédant la combinaison génique Pc38, 39, 68 étaient encore utilisés couramment durant ces années, les races virulentes à l’égard de cette combinaison étaient nombreuses. Ces cultivars ont été remplacés graduellement par de nouveaux qui possédaient des gènes de résistance différents. En 2009, le cultivar ‘Legget’ (Pc68, 94) était semé sur 17.9 % de la superficie consacrée à l'avoine et ‘HiFi’ (Pc91), sur 3 %. De 2007 à 2009, la virulence à l’égard du gène Pc94 était faible (≤ 1.8 %) ou non décelée; la virulence à l’égard du gène Pc91 a été détectée dans un seul isolat; et la virulence à l’égard du gène temp_pc97 ou temp_Pc98 était soit faible ou non décelée. L'immense hausse qu'a connue la fréquence de la virulence à l’égard du gène Pc45, en 2008 et 2009, découlait d'isolats qui possédaient cette virulence et qui migraient des États-Unis vers les Prairies canadiennes.
Introduction
Oat (Avena sativa L.) is an important cereal crop in Canada, ranking behind wheat, barley and canola. Approximately 2.1 Mha were seeded to oat in Canada during 2003–2007 (Statistics Canada, 2003–2007), 90% of which were located in the eastern prairie region (Manitoba and eastern Saskatchewan). Oat grown in the prairie region is highly vulnerable to crown rust [Puccinia coronata Corda f. sp. avenae Eriks.]. While estimated yield losses averaged 5.1% annually during the five-year period from 2001 to 2005, losses in individual years could be higher. For instance, losses in 2001 were 11.2% and losses in 2005 were 8.8% (McCallum et al., Citation2007). In both years, conditions were favourable for early outbreak of crown rust and subsequent development of the disease. In addition, isolates with virulence to overcome resistance in the commonly grown cultivars were abundant (Chong & Zegeye, Citation2004; Chong et al., Citation2008).
Host genetic resistance has long been recognized as the most effective and economical method to control crown rust. In North America, early breeding efforts to develop resistant oat cultivars relied solely on resistance obtained from cultivated hexaploid oat (Martens & Dyck, Citation1989). When all deployable sources of resistance from this host were exhausted in the late 1950s, a new source of resistance to crown rust was found in Avena sterilis L., a wild hexaploid oat native to the Mediterranean regions. Over 40 resistance genes have since been described from this host (Cereal Disease Laboratory, St. Paul, Minnesota: http://www.ars.usda.gov/SP2UserFiles/ad_hoc/36400500Resistancegenes/ocr.xls). However, not all of them were deployable. To some of the genes moderate levels of virulence already existed, even though these genes had never been exposed to the North American P. coronata f. sp. avenae populations (Chong & Kolmer, Citation1993). Additionally, some genes were tightly linked or allelic, and virulence to some of the genes was associated (Chong & Brown, Citation1996; Leonard et al., Citation2005).
At least 16 genes from A. sterilis have been deployed at various times in North America (Leonard, Citation2003). Pc38, Pc39 and Pc68, and to a lesser extent, Pc48, have been extensively deployed in the eastern prairie region of Canada. The gene Pc39 was the first to be deployed with the release of ‘Fidler’ in 1980. This was followed by the release of a series of cultivars (such as ‘Dumont’, ‘Robert’ and ‘Riel’) with both Pc38 and Pc39 combined during the period from 1982 to 1993. When Pc38 and Pc39 were no longer effective in the eastern prairie region in the mid 1990s due to a major shift in virulence to these two genes in the prairie rust population (Chong & Seaman, Citation1997), a second series of cultivars with the Pc38, 39, 68-gene combination were released starting with ‘AC Assiniboia’ in 1995, followed by six other cultivars with the same gene combination. Since Pc38 and Pc39 were no longer effective in the 1990s, these cultivars basically were protected only by Pc68. In 1998, ‘Triple Crown’ was released and its resistance was based on a single gene, Pc48. In 2001, late-planted fields of this cultivar were severely damaged by crown rust (Chong & Zegeye, Citation2004). In 2005, cultivars with the Pc38, 39, 68-gene combination (‘Assiniboia’ and ‘Ronald’) suffered the same fate, and were severely damaged by crown rust. (Chong et al., Citation2008)
In anticipation of race changes in the eastern prairie region, cultivars with different resistance gene combinations were developed. ‘Leggett’ was released in 2004 and has Pc68 and Pc94. ‘HiFi’ with Pc91 and unidentified genes (McMullen et al., Citation2005) was first released to North Dakota and Minnesota in 2001, and then released to the eastern prairie region in 2006. Pc91 is a highly effective gene derived from a cross between tetraploid A. magna Murphy and Terrell and diploid A. longiglumus Durr. (Rooney et al., Citation1994) with A. magna being the donor. Pc94 was derived from diploid oat, Avena strigosa Shreb. (Aung et al., Citation1996), and was still highly effective during 2002–2006 (Chong et al., Citation2008).
Annual oat crown rust surveys have been conducted at the Agricultural & Agri-Food Canada Cereal Research Centre, Winnipeg, since 1929, and the main objectives were to monitor incidence and severity of crown rust and to identify physiologic races. In the early years, common oat cultivars were used as the standard set of differentials, and changes in races in North America have made it necessary to change differential cultivars from time to time (Simons & Murphy, Citation1955). When most of the cultivar differentials became highly susceptible by 1970 (Fleischmann & Baker, Citation1971), the standard set of 10 common oat cultivars was replaced by a new set of 12 single-gene lines each with a newly derived A. sterilis resistance gene as differentials (Harder, Citation1975). The main advantage of using single-gene lines as differentials is that frequencies of virulence to individual genes are more definitive than using common oat cultivars with varying numbers of resistance genes. Several changes were made in subsequent years. This involved replacing several A. sterilis-derived resistance genes from the original set with other genes derived from A. sterilis (Chong & Kolmer, Citation1993). The standard set of differentials currently used for race nomenclature in North America was based on 16 of these single-gene lines as primary differentials (Chong et al., Citation2000). These differentials were selected on the basis of their differential response to the crown rust populations in North America, importance in breeding for resistance or in studies of virulence, and clear distinction of low and high infection types. In addition to the 16 primary differentials, Pc91, Pc94 and Pc96 were added to the standard set as supplemental differentials. Pc91 and Pc94 are present in cultivars ‘HiFi’ and ‘Leggett’, respectively. Pc96 was derived from A. sativa (Chong & Brown, Citation1996), and is a potential candidate for combining with other effective Pc genes.
The present study is part of the continuing effort to monitor the incidence and severity of oat crown rust in the eastern prairie region. The main objectives were to characterize the virulence of the prairie P. coronata f. sp. avenae population using the North American standard differential set (Chong et al., Citation2000), to determine the effectiveness of known Pc genes in cultivars currently grown in the eastern prairie region, and to evaluate the effectiveness of new Pc genes for their potential to be deployed in oat breeding programmes.
Materials and methods
In 2007, surveys for crown rust in Manitoba were conducted from 28 June to 23 August, and in eastern Saskatchewan on 18 and 25 July, and between the second and third week of August. In 2008, the surveys in Manitoba were conducted from 17 July to 29 August, and in eastern Saskatchewan on 26 and 27 August. In 2009, oat crown rust surveys in Manitoba were conducted from early August until 17 September, and in eastern Saskatchewan on 10 and 11 September. All locations surveyed were recorded using a handheld global-positioning device. The Modified Cobb scale (Peterson et al. Citation1948) was used to estimate crown rust severity.
Wild oat (Avena fatua L.) was commonly found in commercial farm fields of all agricultural crops across the eastern prairie region. To avoid over-sampling in any location, no samples were collected from wild oat or cultivated oat in commercial farm fields in locations less than 10 km apart. Samples were also collected from selected oat lines and cultivars in uniform rust nurseries (URN) located in Brandon, Emerson, and Morden, MB; and Regina and Indian Head, SK. The cultivars/lines planted in the URN were: (a) a ‘Sun II’-backcross line with Pc94, (b) ‘AC Assiniboia’, (c) ‘Triple Crown’, (d) ‘Leggett’, (e) ‘HiFi’ (2008 and 2009 only), and (f) ‘Makuru’, as a susceptible check.
Urediniospores from each sample were transferred to four to five seedlings of ‘Makuru’ with a sterile spatula and a small amount of water. The seedlings had been treated with maleic hydrazide (0.277 g L−1 of water) to suppress emergence of secondary leaves. Inoculated seedlings were incubated in a dew chamber (Model I-60D, Percival Scientific, Perry, Iowa) overnight at 18 °C, and then were grown in a greenhouse at 18–25 °C with a 16-h photoperiod supplemented with high-pressure sodium lighting. To minimize cross-contamination, the seedlings in each pot were grown inside a transparent plastic cone connected to a filtered air supply under gentle positive pressure. Approximately 7 d after inoculation, the seedlings in each pot were thinned to a single seedling with the primary leaf trimmed to contain a single uredinium. Approximately two weeks after inoculation, urediniospores from individual uredinia were collected with micro-cyclone collectors into size #00 gelatin capsules. A suspension of urediniospores from each uredinium in 400 μL of light industrial oil (Bayol, Esso Canada) was sprayed onto susceptible ‘Makuru’ seedlings for one further urediniospore increase before inoculation onto a set of differential oat lines described below.
Races of P. coronata f. sp. avenae were determined as described by Chong et al. (Citation2000), using a standard set of 16 primary differentials and three supplemental differentials shown in . Seedlings were evaluated for infection type (IT) 12–14 d after inoculation using a 0 to 4 scale (Murphy, Citation1935), with ITs of 0, 1 and 2 being considered as low (avirulent) and ITs of 3 and 4 as high (virulent). The susceptible cultivar, ‘Makuru’, was included in the differential set as a check for infection levels. In addition, two putative new Pc genes from A. sterilis, temporarily designated temp_pc97 (recessive) and temp_Pc98, were included in the differential sets for evaluation of their effectiveness against isolates from the eastern prairie region.
Table 1. Frequencies (%) of virulence on 19 oat crown rust differentials and two putative new genes of Puccinia coronata f. sp. avenae isolates collected from wild oat (Avena fatua) and cultivated oat (A. sativa) in commercial farm fields in Manitoba and eastern Saskatchewan of Canada during 2007–2009, and statistical significance of difference in frequency of virulence to each differential between isolates from wild oat and cultivated oat
Frequencies of virulence of isolates from wild oat and cultivated oat on the 19 differentials each year were compared by Fisher's exact test using SasINT (SAS OnlineDoc® Version 9.1.3; SAS; Institute Inc. Citation2000–2004, Cary, NC) to determine if there were significant differences in frequencies of virulence between isolates from wild oat and cultivated oat in each of the three years.
Samples obtained from surveys each year were separated into 30 to 40 groups (approximately three to five samples per group). Urediniospores from individual groups were used to inoculate seedlings of ‘Makuru’ for urediniospore increase. The urediniospores subsequently collected from each of the groups (termed composite isolates hereafter) were then used to inoculate a set of resistant lines to detect rare races. The resistant set was composed of ‘HiFi’, ‘Leggett’, ‘Summit’ (2008 and 2009 only), and two single-gene lines (with temp_pc97 and temp_Pc98). Isolates established from susceptible pustules were inoculated onto differential sets for race identification described above.
Results
Incidence and severity of oat crown rust in the eastern prairie region of Canada
In 2007, crown rust was first observed on wild oat in trace amounts in Manitoba on 6 July. Prolonged dry conditions and heat in July and early August slowed crown rust development. Crown rust developed again as temperatures returned to near normal around mid August. Late-planted fields in western Manitoba and eastern Saskatchewan were observed to have 40–60% crown rust severities during the third week of August. Most of these fields were at heading or early milk stage, and most likely would be damaged by the rust if grown to maturity.
Crown rust was much lighter across the eastern prairie region in 2008 than in 2007. June was unusually cool with overnight temperatures often below 10 °C. This was followed by a very warm July, further hindering the development of crown rust. It was not until 11 August that trace amounts of crown rust were first observed in Manitoba, about a month later than usual. Wild oat plants in areas adjacent to the alternate host, buckthorn, near Carman, MB, were still free of crown rust in late July and early August, even though aecia were found on this alternate host during the last week of June. In a normal year, wild oat plants in these locations would mostly have been killed by crown rust by this time. Crown rust remained at trace levels until 29 August. Even late-planted oat fields escaped damage by crown rust in 2008.
The crown rust situation in 2009 was very similar to that in 2008, except that the incidence of crown rust was even lighter and outbreak of the rust was even later in 2009. The unusually low temperatures in June was followed by temperature remaining well below normal in July and early August, with overnight temperature often dipping below 10 °C. Aecia were found on buckthorn in the Carman region on 2 July. However, wild oat plants and commercial oat fields in close proximity to this buckthorn area were free of crown rust in July and early August. Subsequently, trace amounts of crown rust were observed on wild oat on 15 August in Manitoba. Because of the late and light outbreak of crown rust in 2009, the majority of the rust samples were obtained from wild oat during the first two weeks in September. Only a small number of samples were obtained from commercial oat fields in 2009, as much of the oat crop either had matured or had been harvested by September.
Virulence of oat crown rust
A total of 748 single-pustule isolates were established from samples obtained from wild oat and cultivated oat during 2007–2009. The frequencies of virulence of these isolates to the Pc genes of the 19 oat crown rust differentials are shown in . In isolates from wild oat and cultivated oat, virulence frequencies to Pc38 and Pc39 were over 92.0% and 88.0%, respectively, during 2007–2009. The frequency of virulence to Pc48 in isolates from wild oat was between 8.2–14.1%, and in isolates from cultivated oat was between 11.5–18.2%. The frequency of virulence to Pc68 in isolates from wild oat was 45.9% in 2007 and was very similar to this level in 2008 and 2009. Frequencies of virulence to Pc50, Pc54 and Pc58 were mostly below 10.0%. Frequencies of virulence to Pc40, Pc46, Pc51, Pc56 and Pc59 were between 30.6% and 72.7%. Virulence frequency to Pc64 was ≤ 30.0%. Frequencies of virulence to Pc52, Pc62 and Pc96 were below 20.0%. Frequency of virulence to Pc45 in isolates from wild oat increased from 0.6% in 2007 to 19.2% in 2008, and then to 30.8% in 2009. In isolates from cultivated oat, the increase in frequency of virulence to Pc45 occurred only in 2009. The frequency of virulence to Pc68 was significantly higher (P < 0.001) in isolates from cultivated oat than in isolates from wild oat in all three years (). In contrast, the frequencies of virulence to Pc64 and Pc96 were significantly higher (P < 0.05) in isolates from wild oat than in isolates from cultivated oat in 2007, and the frequency of virulence to Pc45 was also significantly higher (P < 0.001) in isolates from wild oat in 2008 ().
During 2007–2009, virulence to Pc91 was found in a single isolate from wild oat in 2009. Virulence to Pc94 was found in three isolates from wild oat (1.8%) and in one isolate from cultivated oat in 2007. Virulence to Pc94 was also found in one isolate from wild oat in 2009. Virulence to temp_pc97 or temp_Pc98 was found in less than 2.0% of the isolates from wild oat in 2007 and 2009. In isolates from cultivated oat, virulence to temp_pc97 was found in four isolates (3.1%) in 2007. Virulence to temp_Pc98 was found in one isolate in 2008 and in one isolate in 2009.
Races of P. coronata f. sp. avenae
Using 19 oat crown rust differentials, a total of 341 races were identified from the combined total of 748 single-pustule isolates established from samples from wild oat and cultivated oat during 2007–2009. Of the 341 races, 287 were identified in isolates from wild oat and 108 in isolates from cultivated oat. For the individual years, 101 races were identified in isolates from wild oat in 2007, 116 races in 2008 and 150 races in 2009. In isolates from cultivated oat, 81 were identified in 2007, 33 in 2008 and 18 in 2009. Among the races identified from wild oat, only 24 of the 287 races (8.4%) were found in all three years. In isolates from cultivated oat, only four of the 108 races (3.7%) were found in all three years. These four races were also among the 24 races found in all three years in isolates from wild oat.
A large proportion of the races (73–80%) identified from wild oat and cultivated oat each year was represented by a single isolate (). Because of the large number of races identified in all three years, those found at less than 1.5% of the isolates each year were excluded from . LQLK-96 (6.5% of the isolates) and LQBK-96 (5.9%) were the two most common races in isolates from wild oat in 2007. BRBG (3.8%) and DRBB (3.2%) were the two most common races in 2008. In 2009, LRBG (3.2%) was the most common race, followed by BRBB, BRLG, and NRBB (all 2.7%). In isolates from cultivated oat, BRBB (7.7%) and LRBG (4.6%) were the most common races in 2007. LRBG (10.2%) and NRBG (10.2%) were the most common races in 2008. In 2009, JRLG, LRBB, LRCG and LRLB (9.1%) were the most common races. However, because of small sample size, the most common races identified in 2009 were each represented by only two isolates out of a total of 22. This figure might not be as reliable as those identified from a larger sample, such as in 2007, when the most common race BRBB (7.7%) was represented by 10 isolates out of a total of 130 isolates.
Fig. 1. Frequency and distribution of races of Puccinia coronata f. sp. avenae from the eastern prairie region of Canada ranked according to number of isolates per race during 2007–2009. For example, 71.2% of the races from wild oat in 2007 and 88.5% of the races from cultivated oat (Avena sativa) in 2007 were represented by a single isolate. Combining the years, 61.2% of the races from wild oat (A. fatua) and 73.9% of the races from cultivated oat were represented by a single isolate. Races were identified by the North American nomenclature system for P. coronata f. sp. avenae (Chong et al., Citation2000) using 19 oat crown rust differentials.
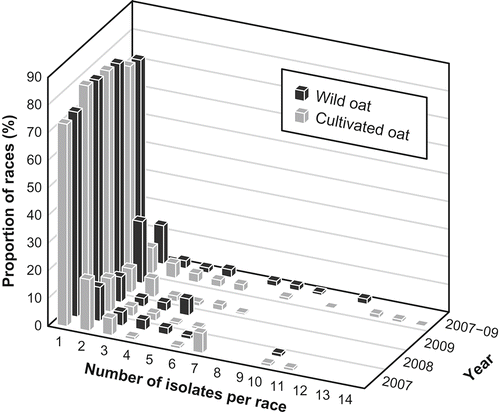
Table 2. Frequency (%) of races of Puccinia coronata f. sp. avenae identified in collections of isolates from wild oat (Avena fatua) and cultivated oat (A. sativa) in commercial farm fields in Manitoba and eastern Saskatchewan during 2007–2009
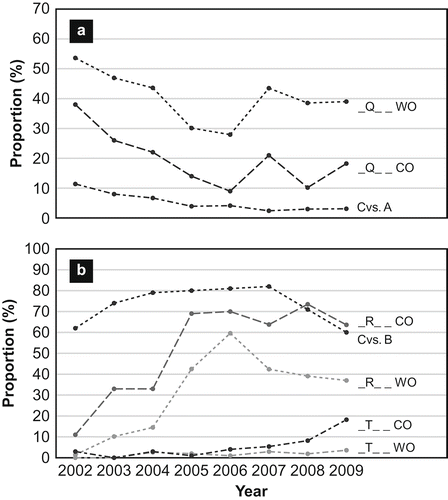
Changes in the prairie crown rust population as a result of selection by the resistance genes being deployed in the eastern prairie region can be more easily followed by considering the reactions on just the second set of differentials (Pc38, Pc39, Pc48 and Pc68), as indicated by the second letter of the race code (Chong et al., Citation2008). For example, the _Q_ _ designation represents races that are virulent to Pc38 and Pc39 and avirulent to Pc48 and Pc68, and the _R_ _ designation represents races that are virulent to Pc38, Pc39 and Pc68 and avirulent to Pc48. During 2007–2009, isolates in the wild oat population were dominated by two race groups ( ). The _Q_ _ group of races accounted for 43.5% of the isolates in 2007, whereas the _R_ _ group accounted for 42.4% of the isolates. There was a slight decline in isolates in these two groups after 2007. In isolates from cultivated oat, the _R_ _ group was the most dominant, representing over 63.0% of the isolates in all three years, and the _Q_ _ group was much smaller, representing 18.2% to 21.0% of the isolates during this period (and ). Virulence to Pc48 was found in race groups, _D_ _, _F_ _, _S_ _, and _T_ _ . In 2007, five _T_ _ races were found in 2.9% of isolates from wild oat, three in 1.9% of the isolates in 2008, and seven in 3.6% of the isolates in 2009 (). In isolates from cultivated oat, there were seven _T_ _ races (2.9%) in 2007, four (8.2%) in 2008, and four (18.2%) in 2009 (). Cultivars such as ‘Dumont’ and ‘Riel’ (Pc38, Pc39), ‘AC Assiniboia’ and ‘Ronald’ (Pc38, Pc39, Pc68), ‘Triple Crown’ (Pc48), and recently released ‘Summit’ (Pc48, Pc68), are susceptible to these _T_ _ races.
Fig. 2. a, Proportions (%) of Puccinia coronata f. sp. avenae isolates identified to be races with virulence to the Pc38, 39-gene combination from wild oat (_Q_ _ WO) and cultivated oat (_Q_ _ CO) in the eastern prairie region of Canada from 2002–2009, and proportions (%) of the total oat areas in Manitoba planted to cultivars with the Pc38, 39-gene combination (Cvs. A). b, Proportions (%) of isolates identified to be races with virulence to the Pc38, 39, 68-gene combination from wild oat (_R_ _ WO) and cultivated oat (_R_ _ CO) and races with virulence to the Pc38, 39, 48, 68-gene combination from wild oat (_T_ _ WO) and cultivated oat (_T_ _ CO) in the eastern prairie region from 2002–2009, and proportions (%) of the total oat areas in Manitoba planted to cultivars with the Pc38, 39, 68-gene combination (Cvs. B). The 2002–2006 race data from Chong et al. (Citation2008) were included to facilitate discussion. The cultivar information was obtained from the Manitoba Crop Insurance Reports – 2002 to 2009. Races were coded by the avirulence and virulence reactions on just the second set of differentials (Pc38 Pc39, Pc48, Pc68), as indicated by the second letter of the race code (Chong et al., Citation2000).
In the isolates from wild oat, the three isolates (1.8%) with virulence to Pc94 in 2007 were identified to be BRBG-94, and the isolate with virulence to Pc94 in 2009 was BSGB-94, 96. Ten races (17.6%) were virulent to Pc96 in 2007, five (5.8%) in 2008, and five (2.7%) in 2009. Three races (1.8%) were virulent to temp_pc97 in 2007, and four races (1.8%) in 2009. Two races (1.2%) were virulent to temp_Pc98 in 2007, and three (1.4%) in 2009. Only a single isolate (DRBG-91) from wild oat was found to have virulence to Pc91 during 2007–2009 (). In isolates from cultivated oat, virulence to Pc96 was found in eight races (7.7%) in 2007. Virulence to temp_Pc98 in 2008 was found in one isolate (BTGG-98) in 2008 and in one isolate (HQPK-98) in 2009. The one isolate (LRBC-94, 96) found in 2007 with virulence to Pc94 also had virulence to Pc96.
In 2007, only one out of 170 isolates (0.6%) from wild oat was virulent to Pc45. In 2008, 25 races in 19.2% of the isolates from wild oat had virulence to this gene. Fifteen of these races belonged to the _Q_ _ group, three in the _R_ _ group, and five in the _S_ _ group. In 2009, the number of Pc45-virulent races increased to 54 (30.8% of isolates). Twenty-nine of these races were found in the _Q_ _ group, five in the _R_ _ group, nine in the _S_ _ group, and two in the _T_ _ group. None of the isolates from cultivated oat had virulence to Pc45 in 2007 and 2008 (). In 2009, five of the 18 races (22.7% of isolates) identified were virulent to Pc45. Two of the races belonged to the _R_ _ group and three to the _Q_ _ group.
Uniform rust nurseries
A total of 92 races were identified in 113 isolates established from samples from selected oat lines and cultivars in the URN during 2007–2009. Twenty-nine were _Q_ _ races, 24 were _R_ _ races, four were _S_ _ races, and five were _T_ _ races. Virulence to Pc96 was found in two _Q_ _ races and in one _S_ _ race in 2007, and in two _Q_ _ races in 2008. Virulence to temp_Pc98 was found in one isolate (BBLB-98) in 2007. Virulence to Pc45 was found in three races of the _R_ _ group and in two races of the _S_ _ group in 2007. Three _S_ _ races with virulence to Pc45 were identified in isolates from URN in 2009. Two of these were the same _S_ _ races found in URN in 2007. Forty of the 92 races from the URN were also found in isolates from wild oat and cultivated oat.
Resistant sets
No susceptible pustules were found on ‘HiFi’ and ‘Stainless’, or on single-gene lines with temp_pc97 and temp_Pc98, after inoculation with composite isolates from each of the three years. Several susceptible pustules were found on the Pc94-line and ‘Leggett’ in 2007 and 2009. The isolates established from these pustules were identified to be BRBB-94 and LRBG-94 in 2007. The nine isolates from the Pc94-line and ‘Leggett’ in 2009 were all identified to be NRLB-94.
Discussion
The incidence of crown rust in 2008 and 2009 was one of the lightest observed in the eastern prairie region. Based on records dating back to 1973, only 1988 and 1989 (Chong & Seaman, Citation1989, Citation1990) appeared to have similar light incidence of crown rust on wild oat and susceptible oat lines as seen in 2008 and 2009. The first appearance of crown rust on 11 August in 2008 and on 15 August in 2009 was the latest ever seen in the eastern prairie region. Prior to 2008, the latest incidence of crown rust reported in the eastern prairie region since 1973 was 25 July in both 1980 and 1989 (Harder & Clark, Citation1981; Chong & Seaman Citation1990). Despite light crown rust in all three years, the crown rust population in the eastern prairie region was just as diverse as in previous years (Chong et al., Citation2008). A large number of races were identified each year. Also, a large proportion of the races (> 73.0%) identified each year were represented by a single isolate. As reported previously (Chong et al., Citation2008), in some years there were significant differences between isolates from wild oat and cultivated oat in frequencies of virulence to several Pc genes. Pc64 and Pc96 have never been deployed in the prairie region, and it is not known why there was a significant difference (P < 0.05) between isolates from wild oat and cultivated oat in frequencies of virulence to these two genes. In the case with Pc68, the difference between isolates from wild oat and cultivated oat in frequency of virulence to this gene was also significant (P < 0.01–0.001) during 2002–2005 (Chong et al., Citation2008), when the major shift in virulence to Pc68 finally occurred in the prairie rust population (Chong et al., Citation2008) after cultivars with the Pc38, 39, 68-gene combination had become widely grown (). During 2007–2009, the difference between the two collections of isolates in frequency of virulence to Pc68 was even more significant (P < 0.001) (). When cultivars with a specific resistance gene dominate so much of the hectarage, they clearly would have an effect if the frequency of virulence to this gene was based on isolates just from commercial oat fields. Using 2007 as an example, 81.5% of the oat hectarage was planted to cultivars with Pc38, Pc39 and Pc68 () Thus, 8 of every 10 samples obtained from commercial oat fields would contain isolates with virulence to the Pc38, 39, 68-gene combination, assuming fields planted to all cultivars were randomly distributed in the prairie region. This extensive deployment of Pc68 in oat cultivars in the Canadian prairie region would explain why the frequency of virulence to Pc68 was consistently higher in isolates from cultivated oat than in isolates from wild oat (). This extensive deployment of Pc68 would also explain why isolates of the _R_ _ races were up to seven times more abundant than isolates of the _Q_ _ races in isolates from cultivated oat ( and ). This lower frequency of _Q_ _ races was a result of isolates with virulence only to Pc38 and Pc39 (i.e. _Q_ _ races) being filtered out in fields planted to cultivars with the Pc38, 39, 68-gene combination. On the other hand, wild oat plants in the prairie region have no resistance to select for or against any isolates in the rust population. This would explain why isolates of the _Q_ _ races were far more abundant in the wild oat collection than in the cultivated oat collection in all three years, but would not explain why these isolates were relatively more abundant during 2007–2009 than in previous years. From 2002 to 2006, there has been a gradual decline of isolates of the _Q_ _ races in the rust population (). Regression analysis showed this decline was significant (P < 0.05–0.01) in isolates from both wild oat and cultivated oat (J. Chong, unpublished). Thus, the proportions of these isolates seen during 2007–2009 were not consistent with the trend of decline of these isolates from 2002 to 2006 (). Since hectarage of cultivars with the Pc38, 39-gene combination has already declined to ≤ 3.1% (), factors other than host selection pressure were involved in maintaining an overabundance of these isolates during 2007–2009.
The huge increase in frequency of virulence to Pc45 in the eastern prairie region in 2008 and 2009 () was unprecedented. This gene has never been deployed in Canada, and virulence to Pc45 has been very low or not detected in most years since 1974 (Chong & Kolmer, Citation1993; Chong & Zegeye, Citation2004; Chong et al., Citation2008). In the USA, frequency of virulence to Pc45 has always been higher, particularly in the winter oat region (Leonard, Citation2003; Carson, Citation2008). A further increase in virulence to this gene was observed in 2008 and 2009 in the USA, after cultivars ‘Morton’ and ‘Kame’ became more widely grown (M. Carson, personal communication). Seedling tests revealed that these cultivars either have Pc45 or another gene, the corresponding virulence to which in the pathogen is associated with virulence to Pc45 (M. Carson, personal communication). The increase in virulence to Pc45 in the USA coincided with the increase in virulence to this gene during the same period in the eastern prairie region. As an unusual increase in virulence to Pc45 was also observed in Ontario in 2008 (D. Falk, personal communication), it is most likely that the increase in virulence to Pc45 in Ontario and in the prairie region was a result of isolates migrating into these regions from the USA.
Movement of cereal rust spores from the USA into the prairie region of Canada occurs annually along the ‘Puccinia Pathway’ (Roelfs & Groth, Citation1989). Such movement of crown rust urediniospores was shown to be significant in 2002, when crown rust isolates typically found in the USA were found in abundance in the prairie rust population (Chong et al., Citation2008). In other years, migration of isolates via urediniospores into the prairie region of Canada might have occurred but gone unnoticed. One reason is that USA and the eastern prairie region have races in common (Chong et al., Citation2000), due to similar extensive use of Pc38 and Pc39, and to a lesser extent of Pc48 and Pc68, in the United States (Carson, Citation2008). Another reason is that migrants might be completely overwhelmed by the local crown rust population as epidemics initiated by aeciospores from the alternate host generally occur much earlier than urediniospores arriving from the USA. The very low incidence and very late outbreak of local crown rust epidemics in the eastern prairie region in 2008 and 2009 might have provided the conditions favouring establishment of migrants from the USA. Presently, virulence to Pc38 and Pc39 is very high and virulence to Pc68 is very low in the USA (Carson, Citation2008). Migration of isolates with this virulence pattern into the prairie region from the USA would explain why _Q_ _ races continued to persist in abundance in the wild oat collection during 2007–2009 (). As the crown rust population overwinters in the eastern prairie region via the alternate host, it remains to be seen how long these isolates, as well as isolates with the unnecessary virulence to Pc45, would persist in the rust population in the absence of host selection pressure.
Since the release of ‘Leggett’ in 2004, hectarage of this cultivar has increased to 17.9% of the total oat hectarage in Manitoba in 2009, but isolates with virulence to Pc94 were still rarely found in the prairie rust population (). Isolates with virulence on ‘HiFi’ were even rarer, even though hectarage of this cultivar has increased to 3.0% in 2009. Only a single isolate with virulence on ‘HiFi’ was found during 2007–2009. Since ‘HiFi’ has other resistance gene(s) in addition to Pc91 (McMullen et al., Citation2005), it is not known to what degree this additional gene(s) might have contributed to the overall resistance of ‘HiFi’. Single-gene lines with Pc91 are now available (provided by Dr Curt McCartney) for characterizing the resistance conferred by this gene in the absence of other resistance genes. Such lines could be important for identifying races virulent to cultivars like ‘Stainless’ and ‘Souris’, which might have Pc91 but may not have additional resistance genes as in ‘HiFi’. ‘Summit’, which has Pc48 and Pc68, is still under limited production for certified seed in 2010, but isolates with this virulence combination (_T_ _ races) were already present in 3.6% of the isolates from wild oat in 2009 and in 8.2% of the isolates from cultivated oat in 2008.
Virulence to the putative new gene temp_pc97 (J. Chong, unpublished) was ≤ 3.1% or not detected in some years during 2007–2009, and virulence to gene temp_Pc98 (J. Chong, unpublished) was present at a frequency between 1.2% and 4.5%. Linkage studies showed that these two putative new genes are not linked to Pc38, Pc39, Pc48, Pc68, Pc94 and Pc96, and also are not linked to each other (J. Chong, unpublished). There was F2 segregation in crosses of temp_pc97 or temp_Pc98 with Pc91, but the number of susceptible plants was very small. As ‘HiFi’ was used in this linkage study, and as this cultivar is known to carry additional resistance genes (McMullen et al., Citation2005), a small number of susceptible plants, e.g. one out of 64 or 256 plants, would be expected in the F2 populations that were segregating for three or four genes. It is most likely that temp_pc97 and temp_Pc98 are not linked to Pc91. Repeating the linkage study using a single-gene line of Pc91 would be more conclusive.
Since 2002, when ‘HiFi’ was first used as an oat crown rust differential, virulence to this cultivar (carrying Pc91 and additional resistance genes) was found in only three isolates. Virulence to Pc91 has not been detected in any combination with Pc94, Pc96, temp_pc97 or temp_Pc98. Combined virulence also has not been detected to temp_pc97 and temp_Pc98, since temp_pc97 and temp_Pc98 were first used as supplemental differentials in 2005 and 2007, respectively. Combined virulence also has not been detected to temp_pc97 and Pc48. As virulence to temp_pc97 or temp_Pc98 is already present in the prairie rust population, it is imperative that temp_pc97 and temp_Pc98 be used in combination with other effective Pc genes, namely Pc91 and (or) Pc94, to prolong the effective lifespan of each of the resistance genes.
Acknowledgements
We are grateful to the co-operators for the care of the uniform rust nurseries in Brandon and Morden, MB, and in Indian Head and Regina, SK. The excellent technical assistance of Michael Shillinglaw in preparing and is gratefully acknowledged.
Related Research Data
References
- Aung , T. , Chong , J. and Leggett , M. The transfer of crown rust resistance gene Pc94 from a wild diploid to cultivated hexaploid oat . Proceedings of the 9th European and Mediterranean Cereal Rusts & Powdery Mildews Conference . 2–6 Sept 1996 , Lunteren, Netherlands. pp. 167 – 171 . Wageningen : European and Mediterranean Cereal Rust Foundation .
- Brouwer , J.B. 1983 . Inheritance and breeding potential of resistance to Puccinia coronata avenae in oats. Ph.D. thesis, Department of Agricultural Genetics and Biometry , University of Sydney .
- Carson , M.L. 2008 . Virulence frequencies in oat crown rust in the United States from 2001 through 2005 . Plant Dis. , 92 : 379 – 384 .
- Chong , J. and Brown , P.D. 1996 . Genetics of resistance to Puccinia coronata f. sp. avenae in two Avena sativa accessions . Can. J. Plant Pathol. , 18 : 286 – 292 .
- Chong , J. and Kolmer , J.A. 1993 . Virulence dynamics and phenotypic diversity of Puccinia coronata f. sp. avenae in Canada during 1974–1990 . Can. J. Bot. , 71 : 248 – 255 .
- Chong , J. and Seaman , W.L. 1989 . Virulence and distribution of Puccinia coronata in Canada in 1988 . Can. J. Plant Pathol. , 11 : 439 – 442 .
- Chong , J. and Seaman , W.L. 1990 . Distribution and virulence of Puccinia coronata in Canada in 1989 . Can. J. Plant Pathol. , 19 : 176 – 180 .
- Chong , J. and Seaman , W.L. 1997 . Incidence and virulence of Puccinia coronata f. sp. avenae in Canada in 1995 . Can. J. Plant Pathol. , 19 : 176 – 180 .
- Chong , J. and Zegeye , T. 2004 . Physiologic specialization of Puccinia coronata f. sp. avenae, the cause of oat crown rust, in Canada from 1999 to 2001 . Can. J. Plant Pathol. , 26 : 97 – 108 .
- Chong , J. , Gruenke , J. , Dueck , R. , Mayert , W. and Woods , S. 2008 . Virulence of oat crown rust [Puccinia coronata f. sp. avenae] in Canada during 2002–2006 . Can. J. Plant Pathol , 30 : 115 – 123 .
- Chong , J. , Leonard , K.J. and Salmeron , J.J. 2000 . A North American system of nomenclature for Puccinia coronata f. sp. avenae . Plant Dis. , 84 : 580 – 585 .
- Fleischmann , G. and Baker , R.J. 1971 . Oat crown rust race differential: replacement of the standard differential varieties with a new set of single resistance gene lines derived from Avena sterilis . Can. J. Bot. , 49 : 1433 – 1437 .
- Harder , D.E. 1975 . Crown rust of oats in Canada in 1974 . Can. Plant Dis. Surv. , 55 : 63 – 65 .
- Harder , D.E. and Clark , R.V. 1981 . Distribution and virulence of Puccinia coronata avenae in Canada in 1980 . Can. J. Plant Pathol. , 3 : 235 – 238 .
- Hoffman , D.L. , Chong , J. , Jackson , E.W. and Obert , D.E. 2006 . Characterization and mapping of a crown rust resistance gene complex (Pc58) in TAM O-301 . Crop Sci. , 46 : 2630 – 2635 .
- Leonard , K.J. 2003 . Regional frequencies of virulence in oat crown rust in the United States from 1990 through 2000 . Plant Dis. , 87 : 1301 – 1310 .
- Leonard , K.J. , Huerta-Espino , J. and Salmeron , J.J. 2005 . Virulence of oat crown rust in Mexico . Plant Dis. , 89 : 941 – 948 .
- Martens , J.W. and Dyck , P.L. 1989 . Genetics of resistance to rust in cereals from a Canadian perspective . Can. J. Plant Pathol. , 11 : 78 – 85 .
- McCallum , B.D. , Fetch , T. and Chong , J. 2007 . Cereal rust control in Canada . Aust. J. Agric. Res. , 58 : 639 – 647 .
- McMullen , M.S. , Doehlert , D.C. and Miller , J.D. 2005 . Registration of ‘HiFi’ oat . Crop Sci. , 45 : 1664
- Murphy , H.C. 1935 . Physiologic specialization in Puccinia coronata avenae . U.S. Dept. Agric. Tech. Bull. , : 433
- Peterson , R.F. , Campbell , A.B. and Hannah , A.E. 1948 . A diagrammatic scale for estimating rust intensity on leaves and stems of cereals . Can. J. Res. C. , 26 : 496 – 500 .
- Roelfs , A.P. and Groth , J.V. 1989 . Epidemiology of the cereal rusts in North America . Can. J. Plant Pathol. , 11 : 86 – 90 .
- Rooney , W.L. , Rines , H.W. and Phillips , R.L. 1994 . Identification of RFLP markers linked to crown rust resistance gene Pc91 and Pc92 in oat . Crop Sci. , 34 : 940 – 944 .
- SAS Institute Inc . 2000–2004 . SasINT (SAS OnlineDoc® Version 9.1.3; SAS , Cary, NC, , USA : Institute Inc. 2000–2004 .
- Simons , M.D. and Murphy , H.C. 1955 . A comparison of certain combinations of oat varieties as crown rust differentials . U.S. Dep. Agric. Tech. Bull. , 1112 : 1 – 22 .