Abstract
Compost teas are fermented watery extracts of composted materials that are used for their beneficial effects on plants including antimicrobial activities. Non-aerated compost teas (NCT) prepared from seaweed compost, shrimp powder compost, and chicken, bovine and sheep manure composts were tested for their effect on the in vitro growth of tomato root pathogens and for their effect on the development of damping-off diseases caused by Pythium ultimum and Rhizoctonia solani. Results showed that NCT prepared from the five types of compost had the ability to greatly reduce mycelial growth of P. ultimum, R. solani, Fusarium oxysporum f. sp. radicis-lycopersici and Verticillium dahliae in vitro. Sterilization of the NCT caused complete or partial loss of their inhibitory effect on mycelial growth of the pathogens. Application of NCT decreased the percentage of necrotic seedlings in P. ultimum-inoculated tomato seeds from 42% to 100%, but did not reduce necrosis in R. solani-inoculated seeds. When applied on tomato seedlings inoculated with damping-off pathogens, various NCT also increased seedling fresh and/or dry weights.
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the authors.
Résumé
Les thés de compost sont des extraits aqueux fermentés de matériaux compostés. On reconnaît aux thés de compost des effets bénéfiques sur les plantes attribuables à leur activité antimicrobienne. Des thés de composts non-aérés (TCNA) à base de compost d'algues, de compost de poudre de crevette et de composts de fumier de poulet, de bovin et de mouton ont été testés afin d'évaluer leurs effets sur la croissance in vitro de différents agents pathogènes de la tomate. L'effet des TCNA sur le développement de la fonte des semis, causée par Pythium ultimum et Rhizoctonia solani, a également été évalué. Les résultats ont montré que les TCNA à base des cinq types de composts réduisent la croissance mycélienne de P. ultimum, R. solani, Fusarium oxysporum f. sp. radicis-lycopersici et Verticillium dahliae in vitro. La stérilisation des TCNA a par ailleurs conduit à une perte complète ou partielle de leur effet inhibiteur sur la croissance mycélienne des agents pathogènes. Appliqués sur les semences de tomate inoculées avec P. ultimum, les TCNA ont réduit de 42 à 100% le pourcentage de semences présentant des lésions nécrotiques. Les TCNA n'ont pas eu un tel effet chez les semences inoculées avec R. solani. L'application de certains TCNA sur les semences de tomate inoculées avec les agents pathogènes de la fonte des semis a résulté en une augmentation du poids sec et/ou humide des jeunes plants.
Introduction
Synthetic fungicides remain the most widely used control measure against fungal plant diseases. Although relatively effective, synthetic fungicides have two major drawbacks: their lack of long-term efficacy caused by the development of resistance in plant pathogens and their adverse effects on human health and the environment (Avis, Citation2007). Therefore, there remains an urgent need for efficient and reliable disease control measures in field and greenhouse crops.
A possible alternative to the use of synthetic fungicides for the control of plant disease is to exploit the antimicrobial activities of compost teas as they provide means for controlling plant pathogens that are considered safer for health and the environment (Siddiqui et al., Citation2009). Compost teas are fermented watery extracts of composted materials that are used for their beneficial effects on plants including antimicrobial activities (Litterick et al., Citation2004).
The potential factors affecting the efficacy of compost teas include the target pathogen and host plant and the preparation methodologies of the teas (compost type, aeration, fermentation additives, duration of fermentation, etc.) (Scheuerell & Mahaffee, Citation2006a ). In previous reports, the type of compost was shown to be a critical factor in the efficacy of the produced tea. Indeed, many authors reported that teas prepared from manure-based composts are generally more effective against plant pathogens than teas from plant-based composts (Weltzien, Citation1991; Al-Dahmani et al., Citation2003; Haggag & Saber, Citation2007). Conversely, others have concluded that the type of the compost does not directly affect the efficacy of the tea and that both manure-based composts and other compost types can provide an efficient tea to control plant pathogens (Elad & Shtienberg, Citation1994; Scheuerell & Mahaffee, Citation2006a ).
Compost teas are reported to control plant pathogens through different mechanisms. The vast consortium of beneficial microbial antagonists present in compost teas is the most reported factor influencing their efficacy (Weltzien, Citation1991; Elad & Shtienberg, Citation1994; McQuilken et al., Citation1994; Cronin et al., Citation1996; Scheuerell & Mahaffee, Citation2004, Citation2006b ; Diánez et al., Citation2006; Gea et al., Citation2009; Koné et al., Citation2010). The microorganisms present in the tea may act as pathogen antagonists through their ability to compete for nutrients and/or space (Al-Mughrabi et al., Citation2008), to destroy pathogens by parasitism (El-Masry et al., Citation2002), to produce antimicrobial compounds, and/or to induce systemic resistance in plants (Zhang et al., Citation1998). Other work suggested that physicochemical properties of the compost teas, namely plant nutrients and organic molecules such as humic or phenolic compounds (Hoitink et al., Citation1997; Siddiqui et al., Citation2008), may protect the plant against disease through improved nutritional status, direct toxicity toward the pathogen and/or induced systemic resistance.
Among the described benefits of compost teas is their ability to decrease foliar plant disease in tomatoes (Elad & Shtienberg, Citation1994; Al-Dahmani et al., Citation2003; Haggag & Saber, Citation2007; Koné et al., Citation2010). However, work on the effect of compost teas on soilborne tomato pathogens is scarce. In this study, non-aerated compost teas (NCT) prepared from various compost types were tested for their effect on the growth of different tomato (Solanum lycopersicum L.) root pathogens and for their effect on the development of damping-off diseases caused by Pythium ultimum Trow and Rhizoctonia solani Kühn.
Materials and methods
Microbial material
Four tomato root pathogens were employed to determine the suppressiveness of NCT. Pythium ultimum (strain 447), R. solani (strain 171), Fusarium oxysporum f. sp. radicis-lycopersici (FORL) Jarvis & Shoemaker (strain 304), and Verticillium dahliae Kleb. (strain 175) were maintained on potato dextrose agar (PDA) (Becton Dickinson, Sparks, MD) at 24 °C. Isolates were obtained from the Laboratoire de diagnostic en phytoprotection (MAPAQ, Québec, Canada).
Preparation of compost teas
Five commercially available composts produced from bovine manure (Voilà, S. Boudrias, Laval, Canada), chicken manure (Fertilo, Fafard, St-Bonaventure, Canada), sheep manure (Voilà, S. Boudrias), shrimp powder (Voilà, S. Boudrias) or seaweed (BioMax, Premier Tech, Rivière-du-Loup, Canada) were used for the preparation of non-aerated compost teas (NCT). NCT were prepared as described in Koné et al. (Citation2010). Briefly, one litre of each compost was homogenized and incorporated in five litres of agricultural well water (St-Frédéric-de-Beauce, Canada) in a 10-litre plastic container. The container was loosely covered and incubated in the dark for 14 days at 21 °C. The NCT were stirred on day 7 and day 14 of incubation. Following the incubation period, the mixture was filtered through eight layers of cheesecloth. The resulting NCT were conserved at 4 °C until use.
Effect of NCT on mycelial growth
The effect of NCT on mycelial growth of tomato root pathogens was measured by individually incorporating the prepared NCT and agricultural well water (control) into warm (45 °C) PDA at 15% (v/v) as described by Koné et al. (Citation2010). In addition, the effect of sterilized NCT on mycelial growth was evaluated by incorporating autoclaved (121 °C, 20 min) or microfiltered (0.2 μm) NCT and agricultural well water (control) into PDA (15%, v/v) as described above. A 5-mm agar plug covered with actively growing mycelium of each of the four tomato root pathogens was inoculated at the centre of each Petri dish. The dishes were incubated for 36 h to 14 days, depending on the pathogen, at 21 °C in the dark. After the incubation period, mycelial growth was measured as the average of two perpendicular diameters of the colony. All experiments were conducted according to a completely randomized design with four replicates. The experiments were repeated twice.
Effect of NCT on damping-off of tomato
Inoculum for the damping-off trials was prepared by inoculating 10 agar plugs (5 mm diameter) covered with actively growing mycelia of P. ultimum or R. solani in 100 mL of potato dextrose broth (PDB) (Becton Dickinson) and incubated at 24 °C on a rotary shaker (150 rpm) for 2 weeks. After the incubation, the culture medium containing the fungal biomass was homogenized in a blender and centrifuged at 5500 × g (24 °C). The supernatant was discarded and the fungal pellets were washed in sterile water and centrifuged (5500 × g, 24 °C). The supernatant was discarded and the fungal cells were resuspended in sterile agricultural well water. The final concentration was adjusted using a haemacytometer to 107 propagules mL−1 for R. solani and 108 propagules mL−1 for P. ultimum.
The efficacy of NCT against damping-off diseases was assessed by placing five tomato seeds ‘Beefsteak’ (Veseys Seeds, Charlottetown, PEI) on sterile Whatman paper in 60 mm sterile Petri dishes. Seeds were treated with one mL of each of the NCT or one mL of agricultural water (control). The treated seeds were then inoculated with one mL of either agricultural water, R. solani or P. ultimum inoculum prepared as previously described. The seedlings were incubated in the dark for 3 weeks at 23 °C. Following incubation, the percentage of germinated seeds and the percentage of seeds showing necrotic lesions were noted. Fresh and dry weights of the seedlings were measured. Experiments were conducted according to a completely randomized design with five replicates. Each experimental unit consisted of five tomato seeds. The experiments were repeated twice.
Statistical analysis
Analysis of variance (ANOVA) was carried out using the GLM procedure of SAS (SAS Institute, Citation1999) and, when significant (P < 0.05), means were separated using Fisher's protected least significant difference (LSD) test. ANOVA of the experimental data showed no significant difference between the two experiments in both the mycelial growth and damping-off trials. The data were therefore combined and analysed as a single experiment for each trial.
Results
Effect of NCT on mycelial growth
NCT significantly reduced mycelial growth of all root pathogens when compared with the water control (). NCT prepared from sheep, bovine and chicken manure composts had the highest inhibitory effect against P. ultimum, reducing mycelial growth by 97–100%. NCT from seaweed and shrimp composts also significantly reduced mycelial growth of P. ultimum, albeit to a lesser extent (40–44% inhibition). Teas prepared from sheep, bovine and chicken manure composts and from shrimp compost also significantly inhibited R. solani growth (89–100% inhibition relative to the control). NCT prepared from seaweed compost showed a significant but lower inhibitory effect against R. solani. All prepared NCT significantly inhibited the mycelial growth of FORL when compared with the control. There was no significant difference between the efficacies of each tested NCT in controlling growth of FORL. NCT prepared from sheep, bovine and chicken manure composts and from shrimp compost completely inhibited V. dahliae growth. Seaweed compost NCT also significantly inhibited V. dahliae albeit to a lesser extent (67%) than the other NCT.
Table 1. Effect of non-aerated compost teas (NCT) on mycelial growth of Pythium ultimum, Rhizoctonia solani, Fusarium oxysporum f. sp. radicis-lycopersici (FORL) and Verticillium dahliae in vitro.
Sterilization by autoclaving generally caused the NCT to lose all inhibitory effect against P. ultimum and R. solani in vitro (). However, autoclaved shrimp NCT still significantly inhibited P. ultimum growth by 86%. Autoclaved bovine manure NCT also significantly inhibited R. solani, albeit to a much lower extent (11% relative to the control) than the non-sterilized NCT. Conversely, sterilization by autoclaving generally caused only a reduction of the inhibition of FORL (25–43% inhibition relative to the control) and V. dahliae (14–21% relative to the control) (). Only autoclaved shrimp NCT lost all inhibition against FORL growth and only seaweed and chicken manure NCT lost all inhibition against V. dahliae.
Table 2. Effect of autoclaved non-aerated compost teas (NCT) on mycelial growth of Pythium ultimum, Rhizoctonia solani, Fusarium oxysporum f. sp. radicis-lycopersici (FORL) and Verticillium dahliae in vitro.
Sterilization by microfiltration caused a loss of NCT inhibition for all NCT-pathogen combinations with the exception of a reduced inhibition of V. dahliae with seaweed, bovine manure and sheep manure NCT (9–21% inhibition relative to the control) and of P. ultimum with seaweed NCT (73%) (). In addition, microfiltered NCT from shrimp compost, chicken manure compost and bovine manure compost significantly stimulated the growth of P. ultimum and FORL.
Table 3. Effect of microfiltered non-aerated compost teas (NCT) on mycelial growth of Pythium ultimum, Rhizoctonia solani, Fusarium oxysporum f. sp. radicis-lycopersici (FORL) and Verticillium dahliae in vitro.
Effect of NCT on tomato damping-off
NCT application on healthy (not inoculated with either P. ultimum or R. solani) seeds did not significantly influence the percentage of germination, the percentage of necrotic seedlings, the fresh and dry weights of the seedlings as compared with the water control (data not shown).
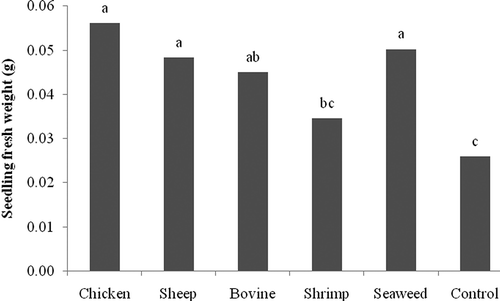
In P. ultimum inoculated seeds, application of NCT did not significantly affect the percentage of germination. However, application of any of the NCT significantly reduced the percentage of necrotic seedlings by 58% (shrimp compost) to 100% (bovine manure compost) (). Fresh weight was enhanced significantly by all NCT, except NCT prepared from shrimp compost () although dry weights of the seedlings were not significantly different in any of the treatments (data not shown).
Fig. 1. In vivo effect of compost teas made from chicken, sheep, and cow manure, and from shrimp and seaweed composts on necrotic tomato seedlings inoculated with Pythium ultimum. Means with the same letter are not significantly different (P = 0.05) according to Fisher's protected LSD test.
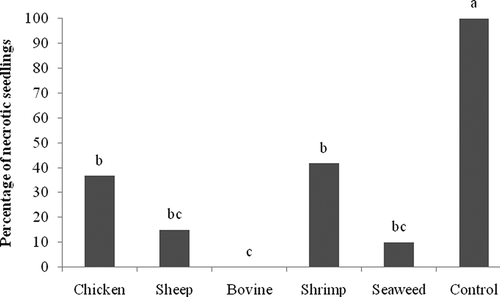
Fig. 2. In vivo effect of compost teas made from chicken, sheep, and cow manure, and from shrimp and seaweed composts on fresh weight of tomato seedlings inoculated with Pythium ultimum. Means with the same letter are not significantly different (P = 0.05) according to Fisher's protected LSD test.
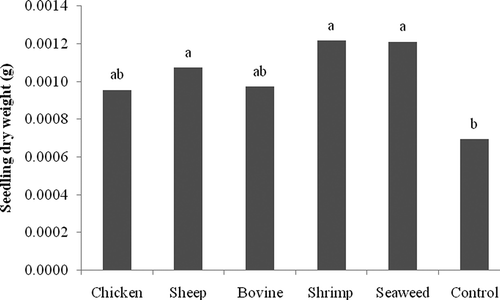
In R. solani inoculated seeds, percentage of germination and percentage of necrotic seedlings were 88 and 100%, respectively. Treatment with NCT did not provide significant differences in percentages of germination or necrotic seedlings when compared with the water control (data not shown). However, application of NCT produced from sheep manure, shrimp and seaweed composts significantly increased fresh weight () and dry weight () of the seedlings.
Fig. 3. In vivo effect of compost teas made from chicken, sheep, and cow manure, and from shrimp and seaweed composts on fresh weight of tomato seedlings inoculated with Rhizoctonia solani. Means with the same letter are not significantly different (P = 0.05) according to Fisher's protected LSD test.
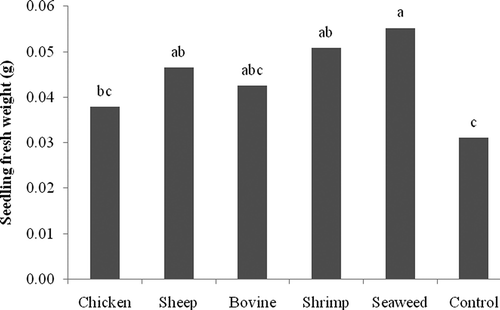
Fig. 4. In vivo effect of compost teas made from chicken, sheep, and cow manure, and from shrimp and seaweed composts on dry weight of tomato seedlings inoculated with Rhizoctonia solani. Means with the same letter are not significantly different (P = 0.05) according to Fisher's protected LSD test.
Discussion
This study showed that NCT prepared from the five types of compost (seaweed compost, shrimp powder compost, and chicken, bovine and sheep manure composts) had the ability to greatly reduce the mycelial growth of P. ultimum, R. solani, FORL and V. dahliae. Among the tested NCT, those produced from chicken, bovine and sheep manure composts gave the highest inhibition of all tested pathogens, reducing the mycelial growth by 78–100% (). Shrimp compost NCT was as effective as the manure-based NCT in inhibiting the mycelial growth of all pathogens except the oomycete P. ultimum. NCT prepared from seaweed compost was less effective than NCT from manure-based composts in inhibiting the pathogens, with the exception of a high (70%) inhibition of FORL. Previous reports have shown that compost teas affect the mycelial growth of numerous plant pathogens (e.g. Scheuerell & Mahaffee, Citation2004, 2006a, 2006b; Haggag & Saber, Citation2007; Al-Mughrabi et al., Citation2008; Siddiqui et al., Citation2009; Koné et al., Citation2010). In particular, aerated grape marc compost tea greatly inhibited the mycelial growth of numerous soilborne fungi, including FORL, F. oxysporum f. sp. lycopersici, F. oxysporum f. sp. radicis-cucumerinum, R. solani, V. dahliae and V. fungicola (Diánez et al., Citation2006).
Sterilization of the NCT by autoclaving or microfiltration caused complete (100%) or partial loss of the inhibitory effect on mycelial growth for all the tested teas. This indicated that the microbial populations in the NCT were at least in part responsible for the direct antimicrobial activity as reported previously (Weltzien, Citation1991; Elad & Shtienberg Citation1994; McQuilken et al., Citation1994; Cronin et al., Citation1996; Zhang et al., Citation1998; El-Masry et al., Citation2002; Scheuerell & Mahaffee, Citation2004; Diánez et al., Citation2006; Gea et al., Citation2009; Koné et al., Citation2010). Mechanisms of action underlying the efficacy of compost teas to control plant pathogens have been reported as single or multiple mechanisms involving microbial antagonism (through antibiosis, parasitism, competition for nutrients/space or induced plant resistance) (Zhang et al., Citation1998; El-Masry et al., Citation2002; Al-Mughrabi et al., Citation2008) or suppressive physicochemical properties (improved nutrient status of the plant, toxic compounds or induced resistance) (Hoitink et al., Citation1997; Siddiqui et al., Citation2008).
Application of the NCT on healthy seedlings did not result in any adverse effects, such as increased percentage of necrotic seedlings, reduced germination, reduced fresh or dry weight, on the tomato seedlings, showing that NCT are not phytotoxic. Previous work had shown that compost tea prepared from bovine manure or municipal solid waste composts had toxic effects on lettuce seedlings (Carballo et al., Citation2009).
When NCT were applied to P. ultimum-inoculated tomato seeds, the percentage of germinated seeds was not affected. However, results demonstrated a drastic decrease in the percentage of necrotic seedlings by 42% (shrimp powder compost tea) to 100% (bovine manure compost tea) relative to the control. This is in agreement with the results of Cummings et al. (Citation2009) where the percentage of spinach (Spinacia oleracea L.) seed emergence was not improved by compost teas, yet successful control of post-emergence seedling damping-off by P. ultimum was achieved using a compost tea prepared from vermicompost and shrimp powder. Scheuerell and Mahaffee (Citation2004) also showed that compost tea prepared from yard trimmings compost and, in particular, vermicomposts had a suppressive effect on P. ultimum damping-off by increasing the number of healthy plants.
In R. solani trials, none of the tested NCT increased germination nor decreased necrotic seedlings. Other work had shown that compost tea prepared from vermicompost and shrimp powder was able to increase spinach seedling emergence and reduce pre-emergence damping-off by R. solani. However, post-emergence damping-off was not affected by this compost tea (Cummings et al., Citation2009).
In some cases, such as with seaweed compost tea, it was shown that NCT were more efficient in reducing the severity of P. ultimum damping-off on tomato seedlings than it was in inhibiting the mycelial growth of the pathogen in vitro. The additional indirect effect of these NCT against P. ultimum in vivo might have been provided by particular microorganisms or compounds within the NCT that, while not directly inhibitory to P. ultimum mycelial growth in vitro, had a possible effect on the induction of intrinsic plant defences that helped stave off the disease as has been reported in other systems (Zhang et al., Citation1998; Haggag & Saber, Citation2007; Siddiqui et al., Citation2009).
In both damping-off trials, various NCT increased the fresh and dry weights of the tomato seedlings in the presence of the pathogens while such effect of NCT was not observed on healthy seedlings. More precisely, NCT prepared from chicken, sheep and bovine manure composts as well as shrimp powder compost increased seedling fresh weight in the P. ultimum trial and NCT prepared from sheep manure, shrimp powder and seaweed composts increased the fresh and dry weights of R. solani-inoculated seedlings. Cummings et al. (Citation2009) reported that aerated compost teas made with vermicompost and shrimp powder also increased total dry weight of spinach seedlings in the absence of significant control of post-emergence damping-off caused by P. ultimum and R. solani. This would indicate that specific compost teas may improve the nutritional status of the plant in the presence of pathogens. Other work has shown that compost teas would also have the secondary effect of improving plant growth and nutrition through a fertilizing effect (Hargreaves et al., Citation2009). Previous physicochemical analysis of the five composts used in this study (Koné et al., Citation2010) revealed that these compost teas would be relatively weak fertilizers based on the low concentration of macro- and microelements in these teas. However, it is possible that the microbial component of these teas may help promote the growth of the seedling through the production of plant growth regulators or other activities as has been shown previously (Kang et al., Citation2006; Mehnaz & Lazarovits, Citation2006; Tsavkelova et al., Citation2006; Gravel et al., Citation2007; Avis et al., Citation2008).
With regard to potential microbial safety issues, previous work with these NCT showed that E. coli was not present in any of the NCT (Koné et al., Citation2010) although some NCT showed the presence of coliforms. While coliforms in NCT have been reported to fall below detectable levels when applied to plants (Welke, Citation2005), care should be taken to limit or eliminate potential human pathogens from compost teas, in particular if preparation includes additives destined to enhance microbial growth (Ingram & Millner, Citation2007).
Overall, results from this study showed that NCT from various compost types provided a significant inhibition of the mycelial growth of soilborne pathogens of tomato and significantly reduced tomato seedling damping-off by P. ultimum. NCT prepared from sheep and bovine manure composts consistently provided the highest inhibition both in vitro and in vivo. Although the complete or partial loss of in vitro efficacy upon sterilization of the NCT indicates that microbial communities are a crucial factor in direct suppressive effects (competition, parasitism, antibiosis) against the pathogens, the induction of plant defence reactions by particular microorganisms or by organic and inorganic compounds in the NCT cannot be ruled out. Finally, specific NCT caused an increase in seedling weight, which may indicate that the microbial communities in the NCT are producing plant growth-promoting substances in the presence of the pathogens. Future work will attempt to identify microbial antagonists in highly suppressive NCT as well as identify compost tea preparation parameters that enhance populations of these antagonists.
Acknowledgements
The authors thank Kim Roger-Bergeron for technical assistance. Antoine Dionne was supported by a scholarship from the Fonds québécois de recherche sur la nature et les technologies (FQRNT). This work was supported by the Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec (MAPAQ).
Notes
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the authors.
References
- Al-Dahmani , J.H. , Abassi , P.A. , Miller , S.A. and Hoitink , H.A.J. 2003 . Suppression of bacterial spot of tomato with foliar sprays of compost extracts under greenhouse and field conditions . Plant Dis. , 87 : 913 – 919 .
- Al-Mughrabi , K.I. , Berthélémé , C. , Livingston , T. , Burgoyne , A. , Poirier , R. and Vikram , A. 2008 . Aerobic compost tea, compost and a combination of both reduce the severity of common scab (Streptomyces scabiei) on potato tubers . J. Plant Sci. , 3 : 168 – 175 .
- Avis , T.J. 2007 . Antifungal compounds that target fungal membranes: applications in plant disease control . Can. J. Plant Pathol. , 29 : 323 – 329 .
- Avis , T.J. , Gravel , V. , Antoun , H. and Tweddell , R.J. 2008 . Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity . Soil Biol. Biochem. , 40 : 1733 – 1740 .
- Carballo , T. , Gil , M.V. , Calvo , L.F. and Moran , A. 2009 . The influence of aeration system, temperature and compost origin on the phytotoxicity of compost tea . Compost Sci. Util. , 17 : 127 – 239 .
- Cronin , M.J. , Yohalem , D.S. , Harris , R.F. and Andrews , J.H. 1996 . Putative mechanism and dynamics of inhibition of the apple scab pathogen Venturia inaequalis by compost extracts . Soil Biol. Biochem. , 28 : 1241 – 1249 .
- Cummings , J.A. , Miles , C.A. and Du Toit , L.J. 2009 . Greenhouse evaluation of seed and drench treatments for organic management of soilborne pathogens of spinach . Plant Dis. , 93 : 1281 – 1292 .
- Diánez , F. , Santos , M. , Boix , A. , De Cara , M. , Trillas , I. , Aviles , M. and Tello , J.C. 2006 . Grape marc compost tea suppressiveness to plant pathogenic fungi: role of siderophores . Compost Sci. Util. , 14 : 48 – 53 .
- Elad , Y. and Shtienberg , D. 1994 . Effect of compost water extracts on grey mold (Botrytis cinerea) . Crop Protect. , 13 : 109 – 114 .
- El-Masry , M.H. , Khalil , A.I. , Hassouna , M.S. and Ibrahim , H.A.H. 2002 . In situ and in vitro suppressive effect of agricultural composts and their extracts on some phytopathogenic fungi . World J. Microbiol. Biotechnol. , 18 : 551 – 558 .
- Gea , F.J. , Navarro , M.J. and Tello , J.C. 2009 . Potential application of compost teas of agricultural wastes in the control of the mushroom pathogen Verticillium fungicola . J. Plant Dis. Protect. , 116 : 271 – 273 .
- Gravel , V. , Antoun , H. and Tweddell , R.J. 2007 . Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: possible role of indole acetic acid (IAA) . Soil Biol. Biochem. , 39 : 1968 – 1977 .
- Haggag , W.M. and Saber , M.S.M. 2007 . Suppression of early blight on tomato and purple blight on onion by foliar sprays of aerated and non-aerated compost teas . J. Food Agric. Environ. , 5 : 302 – 309 .
- Hargreaves , J.C. , Adl , M.S. and Warman , P.R. 2009 . Are compost teas an effective nutrient amendment in the cultivation of strawberries? Soil and plant tissue effects . J. Sci. Food Agric. , 89 : 390 – 397 .
- Hoitink , H.A.J. , Stone , A.G. and Han , D.Y. 1997 . Suppression of plant diseases by composts . HortScience , 32 : 184 – 187 .
- Ingram , D.T. and Millner , P.D. 2007 . Factors affecting compost tea as a potential source of Escherichia coli and Salmonella on fresh produce . J. Food Protect. , 70 : 828 – 834 .
- Kang , B.R. , Yang , K.Y. , Cho , B.H. , Han , T.H. , Kim , I.S. , Lee , M.C. , Anderson , A.J. and Kim , Y.C. 2006 . Production of indole-3-acetic acid in the plant-beneficial strain Pseudomonas chlororaphis O6 is negatively regulated by the global sensor kinase GacS . Curr. Microbiol. , 52 : 473 – 476 .
- Koné , S.B. , Dionne , A. , Tweddell , R.J. , Antoun , H. and Avis , T.J. 2010 . Suppressive effect of non-aerated compost teas on foliar fungal pathogens of tomato . Biol. Control , 52 : 167 – 173 .
- Litterick , A.M. , Harrier , L. , Wallace , P. , Watson , C.A. and Wood , M. 2004 . The role of uncomposted materials, compost, manures and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural crop production – a review . Crit. Rev. Plant Sci. , 23 : 453 – 479 .
- McQuilken , M.P. , Whipps , J.M. and Lynch , J.M. 1994 . Effects of water extracts of a composted manure-straw mixture on the plant pathogen . Botrytis cinerea. World J. Microbiol. Biotechnol. , 10 : 20 – 26 .
- Mehnaz , S. and Lazarovits , G. 2006 . Inoculation effects of Pseudomonas putida, Gluconacetobacter azotocaptans, and Azospirillum lipoferum on corn plant growth under greenhouse conditions . Microb. Ecol. , 51 : 326 – 335 .
- SAS Institute . 1999 . SAS/STAT User's Guide , Cary , NC : SAS Institute . Version 8
- Scheuerell , S.J. and Mahaffee , W.F. 2004 . Compost tea as a container medium drench for suppressing seedling damping-off caused by Pythium ultimum . Phytopathology , 94 : 1156 – 1163 .
- Scheuerell , S.J. and Mahaffee , W.F. 2006a . Variability associated with suppression of gray mold (Botrytis cinerea) on geranium by foliar applications of nonaerated and aerated compost teas . Plant Dis. , 90 : 1201 – 1208 .
- Scheuerell , S.J. and Mahaffee , W.F. 2006b . “ Compost teas: alternative approaches to the biological control of plant diseases ” . In Microbial ecology of aerial plant surfaces , Edited by: Bailey , M.J. , Lilley , A.K. , Timms-Wilson , T.M. and Spencer-Phillips , P.T.N. 165 – 179 . Wallingford , , UK : CABI .
- Siddiqui , Y. , Meon , S. , Ismail , M.R. and Ali , A. 2008 . Trichoderma-fortified compost extracts for the control of choanephora wet rot in okra production . Crop Prot. , 27 : 385 – 390 .
- Siddiqui , Y. , Meon , S. , Ismail , R. and Rahmani , M. 2009 . Bio-potential of compost tea from agro-waste to suppress Choanephora cucurbitarum L. the causal pathogen of wet rot of okra . Biol. Control , 49 : 38 – 44 .
- Tsavkelova , E.A. , Yu , S. , Cherdyntseva , T.A. and Netrusov , A.I. 2006 . Microbial producers of plant growth stimulators and their practical use: a review . Appl. Biochem. Microbiol. , 2 : 117 – 126 .
- Welke , S.E. 2005 . The effect of compost extract on the yield of strawberries and the severity of . Botrytis cinerea. J. Sustain. Agr. , 25 : 57 – 68 .
- Weltzien , H.C. 1991 . “ Biocontrol of foliar fungal disease with compost extracts ” . In Microbial ecology of leaves , Edited by: Andrews , J.H. and Hirano , S.S. 430 – 450 . New York , NY : Springer-Verlag .
- Zhang , W. , Han , D.Y. , Dyck , W.A. , Davis , K.R. and Hoitink , H.A.J. 1998 . Compost and compost water extract-induced systemic acquired resistance in cucumber and Arabidopsis . Phytopathology , 88 : 450 – 455 .