Abstract
Crown rot of banana fruits is caused by a complex of fungal pathogens, the most common of which is Colletotrichum musae, and is one of the main quality defects of exported bananas. Susceptibility of banana fruits to crown rot is influenced by many pre-harvest factors. The aim of this study was to improve on the methodology for the evaluation of fruit susceptibility and to verify whether cultivation areas in Cameroon as well as seasonal variations have an influence on the susceptibility to crown rot. Fruit susceptibility was evaluated on a monthly basis throughout a year (including the dry and rainy seasons) in three banana plantations located in very different agro-ecological conditions (two in a lowland area and one in a highland area). Fruit susceptibility was determined through an internal necrotic surface (INS) assessment after artificial inoculation with C. musae. The standardization of post-inoculation environmental conditions enabled more reliable INS assessments. Fruit susceptibility was found to be significantly influenced by cultivation area (P < 0.001) since fruits grown in low altitude (Dia-dia, Koumba, 80 m) were more susceptible than fruits grown in high altitude (Ekona, 500 m). Although no seasonal effect was observed (P = 0.075), there was a highly significant date effect (P < 0.001). This was specifically the case in low-altitude plantations where fruit susceptibility was higher for some harvest dates within the rainy season. In Ekona, fruit grade and number of leaves on the banana plant were found to be significantly higher than in the two other locations, while black leaf streak disease severity was significantly lower. The potential relationship with fruit susceptibility is fully discussed.
Résumé
La pourriture du collet de la banane est causée par un complexe d'agents pathogènes fongiques, dont le plus courant est Colletotrichum musae, et est un des principaux défauts de qualité des bananes exportées. La réceptivité de la banane à l’égard de la pourriture du collet est influencée par plusieurs facteurs relatifs à la période avant-récolte. Le but de cette étude était d'améliorer la méthodologie permettant d’évaluer la susceptibilité des fruits et de vérifier si les régions de culture au Cameroun de même que les variations saisonnières avaient une influence sur la réceptivité à l’égard de la pourriture du collet. La sensibilité des fruits a été évaluée mensuellement dans le cours d'une année (y compris durant la saison sèche et la saison des pluies) dans trois plantations localisées dans des régions différentes sur le plan des conditions agroécologiques (deux dans les basses terres et une dans les montagnes). La susceptibilité des fruits a été déterminée par l’évaluation de la nécrose interne de la surface (NIS) après inoculation artificielle avec C. musae. La normalisation des conditions environnementales post-inoculation a permis d'obtenir des évaluations de la NIS plus fiables. La susceptibilité des fruits s'est avérée significativement influencée par la région de culture (P < 0.001) étant donné que les fruits cultivés dans les basses terres (Dia-Dia, Koumba, 80 m) étaient plus susceptibles que ceux cultivés en altitude (Ekona, 500 m). Bien qu'aucun effet saisonnier n'ait été observé (P = 0.075), il y avait un effet significatif relatif à la date (P < 0.001). C’était le cas particulièrement dans les plantations localisées dans les basses terres où la susceptibilité des fruits était plus grande à certaines dates de récolte durant la saison des pluies. À Ekona, le calibre des fruits et le nombre de feuilles sur les bananiers étaient significativement supérieurs aux deux autres régions, tandis que la gravité de la maladie des raies noires était significativement moindre. La relation potentielle quant à la susceptibilité des fruits y est discutée en détail.
Introduction
In Cameroon, the main quality defect of banana fruit exported to Europe is related to a post-harvest disease known as crown rot. This post-harvest disease is caused by a broad unspecific and opportunistic fungal complex. Within this complex, Colletotrichum musae (Berk. & Curt.) Arx is the most commonly described pathogenic species (Finlay & Brown, Citation1993; Krauss & Johanson, Citation2000; Lassois et al., Citation2010a). Colletotrichum musae is also responsible for banana anthracnose and wound anthracnose. At harvest, crowns of fruit appear to be healthy but after a few days of shipping, fungal infection develops superficially and internally, affecting the crowns, the pedicels and ultimately the banana pulp. The formation of clusters at the packing station involves cut sections that are penetration tracts for pathogens. Fruit contamination can occur within the field, but it mostly occurs in the washing tanks at the packing station (Shillingford, Citation1977; Finlay et al., Citation1992).
The control of post-harvest diseases of bananas intended for export mainly relies on post-harvest fungicide treatments (de Lapeyre de Bellaire & Nolin, Citation1994). Unfortunately, the chemical control method currently used is not satisfactory and has many other shortcomings, including: (i) resistance build-up in pathogen populations (Johanson & Blazquez, Citation1992; de Lapeyre de Bellaire & Dubois Citation1997); (ii) unacceptable fungicide residues in fruit; and (iii) potential human and environmental hazards with the fungicide mixtures used for post-harvest treatment of banana crowns rejected in packing stations after packing. Therefore, interest in non-chemical alternatives for the management of crown rot and other post-harvest diseases of bananas is increasing.
Geographical and seasonal variations (temperature and rainfall) have been shown to play a role in the incidence of post-harvest diseases in bananas (Lukezic et al., Citation1967; Shillingford, Citation1978; Chillet & de Lapeyre de Bellaire, Citation1996; Chillet et al., Citation2000; Krauss & Johanson, Citation2000). It has been suggested that these spatiotemporal fluctuations could reflect the variations in the quality potential of the fruit, acquired in the field. This quality potential would include two components: the fruit susceptibility to the post-harvest diseases (physiological component) and the level of fruit contamination by fungal pathogens (pathological component). Both components depend on pre-harvest factors including agronomic practices and soil–climate factors (Chillet & de Lapeyre de Bellaire, Citation1996; Chillet et al., Citation2000; Lassois et al., Citation2010b ). Alternative control methods as well as any control measures should rely on a better understanding of these pre-harvest factors as previously shown for wound anthracnose, another post-harvest disease of bananas (de Lapeyre de Bellaire et al., Citation2000; Chillet et al., Citation2006, Citation2007). However, little is known about pre-harvest factors which influence the post-harvest development of crown rot disease. Although pedo-climatic conditions and agro-technical factors are known to influence the development of this disease (Lukezic et al. Citation1967; Shillingford, Citation1978; Krauss & Johanson, Citation2000), only few studies link these fluctuations to either fruit susceptibility (Lassois et al., Citation2008, Citation2010a ) or to fruit contamination.
The aim of this work was therefore to examine the effects of geographical and seasonal variations on banana fruit susceptibility to crown rot. Preliminary studies were conducted in order to show how the standardization of the post-inoculation environmental conditions (PIEC) during the assessment of banana susceptibility to the disease by artificial inoculations could improve accuracy of assessment. In the second part, cultivation areas and seasonal variations were evaluated to understand their effects on the susceptibility of bananas to crown rot disease.
Materials and methods
Plant material
Banana fruits (Musa acuminata Colla [AAA group, Cavendish subgroup] ‘Grande Naine’) were harvested from commercial banana farms in Cameroon. Bunches were tagged at flowering. They were also covered with a plastic sleeve at this stage. Bunches were harvested at a constant physiological age (Jullien et al., Citation2008), determined when the sum of the mean daily temperature accumulated by the fruit between flowering and harvest reached 900 degree days (dd) according to the procedure described earlier (Ganry & Chillet, Citation2008). Temperatures were recorded at a weather station in the plantation so as to predict the time due for harvest. An electronic probe (Tinytag Plus, TGP-4017; Gemini Data Loggers, Chichester, UK) with regular recording intervals (every 15 min) was installed in each of the experimental plots. Average daily temperature was calculated from these data.
Evaluation of susceptibility to crown rot through artificial infection
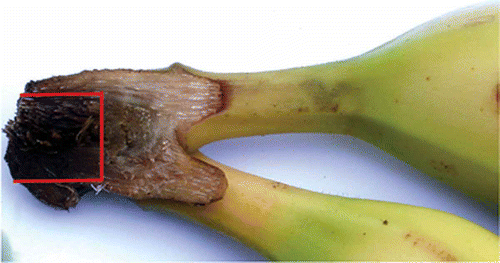
The susceptibility of banana fruits to crown rot was evaluated through artificial inoculation of banana clusters with C. musae. The second hand (occasionally the third hand was also harvested if needed) of each bunch was harvested in the field the day before experimentation (15 bunches per harvest date) and stored at 13 °C. In the laboratory, for each banana plant, the hands were divided into three to five clusters of four healthy banana fingers according to the experiments. These clusters were rid of any defects and the fingers located at the ends of the hands were systematically eliminated. The cuttings of the crowns were square, with regular and clear-cut sections in order to obtain similar crowns between the clusters. As commonly done in previous research on crown rot (Lassois et al., Citation2008; Bastiaanse et al., Citation2010), once latex exuded, crown tissues were dried with absorbent paper and sterilized by dipping in 50% alcohol. Fruits were then laid out at room temperature for 2 h so as to allow the crowns to dry. The C. musae isolate Co-CMR-65 is a standard monosporic strain, that has been previously isolated from a banana fruit harvested in Njombé (Cameroon), tested for its pathogenicity and preserved at −20 °C in a glycerol solution (30%) in order to avoid fungal variations. New cultures of the C. musae strain were initiated, 15 days before each inoculation, starting from freezed cryotubes of mycelium. Thereafter, on the fifth day, the strain was transferred to Mathur's medium (MgSO4.7H2O, 2.5 g L−1; KH2PO4, 2.7 g L−1; peptone, 1 g L−1; yeast extract, 1 g L−1; saccharose, 10 g L−1; agar, 15 g L−1) and incubated at 25 °C for 10 days. Conidia were harvested in sterile distilled water and passed through a 45 μm sterile sieve and the concentration of conidia was maintained at 104 conidia mL−1 using a Malassez cell (Marienfeld, Germany). A 50 μL droplet of the C. musae suspension was then deposited midway through the exposed surface of the crown. A sterilized filter paper was placed on the droplet in order to maintain the inoculum in place. Three hours after inoculation, the fruit clusters were packed in perforated plastic bags in commercial boxes. They were stored under different conditions according to the protocol assigned to each experiment as described below. At the end of the storage period, an evaluation of the internal progression of the crown rot was carried out. The clusters were divided into two parts and the transverse cutting of the crown allowed a visualization of the surface of rot spread into the crown (). This necrotic surface (INS) was adjusted to a rectangular shape and the length (L) and the width (W) of this surface were measured. The ‘Internal Necrosis Surface’ expressed in mm2 was calculated according to the rectangle surface formula: INS = L × W. Its average value was taken as a measure of fruit susceptibility to crown rot.
Fig. 1. Clusters of bananas divided into two parts through the transverse cutting of the crown for the visualization of internal crown rot symptoms that was adjusted to a rectangular shape for measurements of the necrotic surface.
Experimental sites
Experiments were carried out in three commercial banana farms located in representative pedo-climatic conditions, as follows: (1) Dia-dia: this plot was located in a plantation belonging to Plantation du Haut Penja (PHP), at 80 m altitude near Njombé in the Littoral region with an average temperature of 27 °C and a mean annual rainfall of 2500 mm; (2) Koumba: this plot was located in a plantation belonging to Société des Plantations du Manengouba (SPM), at 80 m altitude near Mbanga in the Littoral region with an average temperature of 26 °C and a mean annual rainfall of 2500 mm; (3) Ekona: this plot was located in a plantation belonging to Cameroun Development Corporation/Delmonte (CDC/Del Monte), at 500 m altitude on the slopes of Mount Cameroon, Moliko near Buea in the South-west region with an average temperature of 22 °C and a mean annual rainfall of 2300 mm.
The climate in the banana-producing areas of Cameroon is equatorial and it is characterized by two main and contrasting seasons: a rainy season with heavy rains from April to October and a dry season from November to March. The average temperature and rainfall during the dry season are 27 °C and 63 mm month−1 in Dia-dia and Koumba; 23 °C and 45 mm month−1 in Ekona, respectively. During the rainy season, the average temperature and rainfall are 26 °C and 309 mm month−1 in Dia-dia and Koumba; 22 °C and 301 mm month−1 in Ekona, respectively. The rainy season was considered, in this study, as a period with rainfall higher than 150 mm month−1.
Experiment 1: evaluation of fruit susceptibility through artificial infection and the standardization of post-infection environmental conditions
This experiment was conducted in September 2007, in the packing house of two commercial banana plantations: Koumba and Dia-dia. For each repetition of the experiment, 15 banana bunches were selected. Five clusters (A, B, C, D, E) were cut off from the second and third hands. The susceptibility of each cluster to crown rot was assessed as described previously. Clusters of the same treatment (A, B, C, D, E) were packed in the same commercial box and stored under stable conditions at 13 °C for different periods before necrotic assessment: A: 7 days; B: 9 days; C: 11 days; D: 13 days; E: 15 days. Fruits were allowed to ripen naturally without the use of applied ethylene. Each experiment was repeated three times for three consecutive weeks.
Experiment 2: Evaluation of geographical and seasonal effects on fruit susceptibility through artificial infection and post-infection environmental conditions similar to commercial export conditions
This experiment was conducted between April 2006 and May 2007 in Dia-dia, Koumba and Ekona. At each location, 15 banana plants were selected and labelled at the flowering stage, and three clusters were cut off from the second hand of each bunch at harvest time. The susceptibility of bananas to crown rot was evaluated as described previously. After inoculation, banana fruits were placed in commercial boxes and stored in conditions which simulate commercial exportation. Boxes were stored at 13 °C for 10 days to simulate shipping. Artificial ripening was then initiated by dipping the bananas for 5 s in an ethrel solution (2-chloroethyl phosphonic acid, 480 g L−1), and keeping the bananas at 20 °C for another three days before crown rot assessment. This experiment was repeated every month during a year.
Experiment 3: Evaluation of geographical and seasonal effects on fruit susceptibility through artificial infection and standardization of post-infection environmental conditions
This experiment was conducted between November 2007 and October 2008 in Dia-dia, Koumba and Ekona. At each location, 15 banana plants were selected and labelled at the flowering stage. Three clusters were cut off from each second hand harvested and the susceptibility of bananas to crown rot was evaluated as described previously. After inoculation, fruits were stored in stable conditions at 13 °C for 13 days in order to evaluate their susceptibility. In addition to the assessment of fruit susceptibility, some fruits and banana plant characteristics were measured at harvest stage (900 dd) in the three locations:
| - | the number of leaves (NL) on the plant, | ||||
| - | the number of hands (NH) per bunch, | ||||
| - | the grade of the median fruit (Gr) on the second hand. | ||||
Black leaf streak disease (BLSD) of bananas caused by Mycosphaerella fijiensis (Morelet) can lead to substantial reduction in the leaf area with direct effects on fruit quality and fruit growth. The severity index of BLSD was also assessed, according to the method described by Gauhl et al. (Citation1993). This method involves a visual estimation of the necrotic area per plant, scored according to a six-disease grade scale. This was done on all the leaves of each banana plant harvested at 900 dd. The assessments were conducted on a monthly basis during a year in order to evaluate seasonal effects.
Statistical analyses
The effects of geographical and seasonal variations on banana susceptibility to crown rot were analysed. This was done by subjection of the average INS values calculated for the three clusters to partially nested mixed four-way ANOVA performed with Minitab software v.15.1, each cluster being taken as an experimental unit and location, season, date and bunch as factors. The different variables were analysed by subjection of the value (INS, Gr, NH, NL and BLSD severity) to a mixed three-way ANOVA performed with Minitab software v.15.1 with location, date and bunch as factors. Multiple comparisons of the means were done by applying Tukey test at a 5% probability level.
Results
Importance of post-infection environmental conditions on necrotic assessments
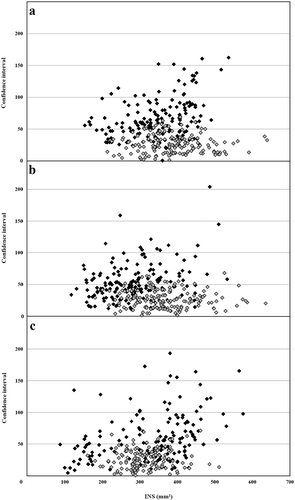
In order to evaluate the effect of the standardization of post-infection environmental conditions (PIEC) on INS assessment, we compared the data of experiment 2 (PIEC similar to commercial conditions) and data of experiment 3 (stable PIEC). This comparison showed that the confidence intervals of the means for INS were reduced at all three locations where PIEC was standardized at 13 °C and where fruit were allowed to ripen naturally ( a, b, c). For further analysis of banana susceptibility to crown rot disease, only data obtained with these new post-harvest environmental conditions was considered.
Fig. 2. Representation of different means of INS versus their confidence intervals at 95% assessed in two experiments differing by their post-inoculation environmental conditions in (a) Dia-dia, (b) Koumba and (c) Ekona. In experiment 1 (Black ![]()
Time-course of necrotic assessments under stable conditions
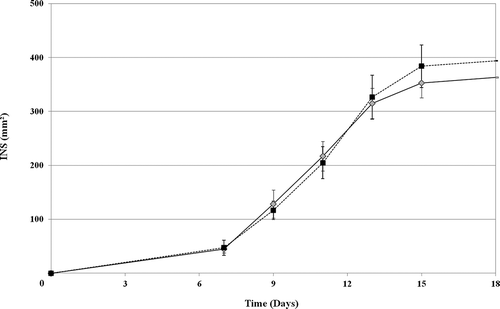
The time-course of INS at 13° C (experiment 1) is typically a sigmoid curve (). From harvest day (day 0) to day 7, necrotic surface extension was low. From the days 7 to 13, necrotic extension was linear. After day 13, the necrotic surface increased slowly and stabilization was at day 15. This time course was the same for Dia-dia and Koumba locations. As linear extension of necrotic surface ended at day 13, this date was chosen for all further estimation of the level of fruit susceptibility to crown rot.
Fig. 3. Time course of the variation of internal necrotic surface (INS) in Dia-dia (——) and Koumba (-- -- -- --). Each point represents the average mean of three replicates for each location (for each repetition INS was assessed over 15 fruits). Error bars represent standard deviation.
Geographical and seasonal variation effects on banana susceptibility to crown rot
Here, the data from experiment 3 where PIEC have been stabilized were the only ones considered. The cultivation area was found to significantly influence (P < 0.001) the susceptibility of fruit to crown rot (). Fruit susceptibility to crown rot was consistently higher in the two low-altitude plantations (average value of INS: 385.8 mm2 in Dia-dia; 366.4 mm2 in Koumba) compared with the high-altitude plantation (average value of INS: 313.4 mm2 in Ekona). As shown in , the level of fruit susceptibility to crown rot in Dia-dia () and Koumba () seems to be higher during the rainy season compared with during the dry season. However, seasonal variation over the year had no significant effect (P = 0.075) on fruit susceptibility to crown rot (). It should also be noted () that during the same season, there was a significant date effect on fruit susceptibility (P < 0.001). This date effect was especially marked in low-altitude zones ( a, b), as confirmed by the significant interaction (P = 0.011) between location and date within a season (), although no significant interaction between location and season was observed (P = 0.111). Moreover, the susceptibility level was particularly higher for both low altitude locations for harvests in the months of July, August and September ( a, b). A highly significant bunch effect was also observed (P < 0.001), as expected, from previous studies (Lassois et al., Citation2010a ). Hence, two statistically different groups were distinguished between fruits harvested in the low-altitude plantations of Dia-dia and Koumba and those harvested at Ekona ().
Table 1. Variance analysis of geographical and seasonal effects on banana susceptibility to crown rot
Table 2. Mean values and standard deviations of internal necrosis surface (INS), variation values (min – max) of INS, grade, number of hands (NH), number of leaves (NL) and black leaf streak disease (BLSD) severity measured during one year in two lowland plantations (Dia-dia and Koumba) and one highland plantation (Ekona). P is the probability
Banana fruit and banana plant characteristics at harvest
The grade of the median fruit at harvest was consistently higher (P < 0.001) in the high-altitude plantation (average value of Grade: 37.4 mm in Ekona) than in the low-altitude plantations (average value of Grade: 32.7 mm in Dia-dia; 32.5 mm in Koumba). Moreover, the number of leaves at harvest (NL) was also significantly higher (P < 0.001) in the high-altitude plantation (average value of NL: 9.6 in Ekona) as compared with the low-altitude plantations. Hence, two statistically different groups were distinguished between fruits harvested at Ekona and those harvested at Dia-dia and Koumba (). In addition, BLSD severity was significantly lower in Ekona than in the two other locations. Although the number of hands of the bunches harvested (NH) differed significantly amongst the three locations (P = 0.006), the average number of hands in Ekona did not differ from the two other locations (). As such, bananas harvested in Ekona (high-altitude plantation) had the lowest BLSDS and INS values, and the highest values of fruit grade and number of leaves as compared with bananas harvested in Dia-dia and Koumba (low-altitude plantations).
Discussion
One of the objectives of this study was to propose methods of improvements on the methodology for a more accurate assessment of banana susceptibility to crown rot by artificial inoculations. The results show that the standardization of PIEC has increased the reliability of necrotic surface assessments. Increase in necrosis under a continuous temperature of 13 °C followed a typical sigmoid curve. During the first phase of this sigmoid (0 to 7 days), the necrotic extension was discovered to be too slow to assess susceptibility variations. In the last phase (after 15 days) the extension of the crown rot was too high and the crown was almost completely rotten. It was considered that the 13 days lapse of time (end of linear progression) could permit a reliable appreciation of necrotic surface and of fruit susceptibility.
It was hypothesized that the banana cultivation area could influence fruit susceptibility to crown rot. Our data confirmed this hypothesis and provided evidence for wide variations (average INS monthly values ranged from 279 mm2 to 495 mm2 according to the location or the period) in the susceptibility of bananas to crown rot as previously seen in fruit susceptibility to wound anthracnose (Chillet et al., Citation2000). Bananas grown in both low-altitude plantations (Dia-dia and Koumba) were more susceptible to crown rot than those grown in the high altitude plantation (Ekona). Similarly, bananas grown in highland plantations in Guadeloupe were found to be less susceptible to wound anthracnose (Chillet et al., Citation2006, Citation2007). Because of low temperatures, fruits grown in high altitudes take more time to reach 900 dd than fruits grown in low altitudes. During this long growth period, they accumulate larger amounts of assimilates that could participate in the reduction of the fruit susceptibility level. Low temperatures favour assimilate availability, and affects growth (plant size) more than carbon assimilation (Pollack, Citation1990; Equiza et al., Citation1997). Such effects of low temperatures on host resistance have also been reported for other pathosystems such as Bipolaris sorokiniana/barley, Puccinia poae-nemoralis/bluegrass and Erysiphe necator/Vitis vinifera through the possible triggering of biochemical pathways associated with disease resistance (Moyer et al., Citation2010). As for banana wound anthracnose (Chillet et al., Citation2007), it was hypothesized that in such low-temperature conditions, assimilates are produced in excess as indicated by the bigger grade observed in Ekona at harvest. It has been shown, in the case of leaf rust resistance in wheat, that the setting up of the defence mechanism (phenylpropanoid pathway) represents an important energetic demand, and that several metabolic pathways that contribute to increased carbon flux through the tricarboxylic cycle are triggered concomitantly to the genes involved in phenylpropanoid metabolism (Bolton et al., Citation2008). It can then be hypothesized that the excess of assimilates contributes to this important energetic demand for secondary metabolic pathways, promoting the development of defence mechanisms.
In addition, it is interesting to note that banana plants at Ekona also had the highest number of leaves, leading to a higher source-sink (So-Si) ratio which makes the fruit less susceptible to crown rot disease as compared with fruits from plantations in Dia-dia and Koumba. These observations are in accordance with previous studies which have shown that an increased So-Si ratio enabled a significant reduction of fruit susceptibility to crown rot disease (Lassois et al., Citation2010a ).
Bananas grown in Ekona had a lower BLSD severity. This might equally account for the higher number of leaves. BLSD is a foliar disease with direct effects on yield and all indirect effects on fruit quality (Jones, Citation2000). It has been shown that BLSD and Sigatoka Disease (another related Mycosphaerella foliar disease of bananas) strongly reduce the pre-climacteric life (called the greenlife) of bananas harvested at a constant physiological age (Abadie et al., Citation2008; Chillet et al., Citation2009), whereas other abiotic stresses do not have any influence on the greenlife of bananas harvested at this same physiological age (Jullien et al., Citation2008). Because BLSD has a strong effect on greenlife, a potential effect on the susceptibility of fruit to crown rot could also be hypothesized and should be further investigated.
Fruit susceptibility to crown rot may fluctuate in the same cultivation area. Even if seasonal effect was not significant, some variations have been found in the low-altitude plantations where fruit susceptibility was particularly higher for some harvest dates within the rainy season. It has been shown that the incidence of post-harvest diseases follows a seasonal variation, disease incidence being generally higher in the rainy season (Chillet & de Lapeyre de Bellaire, Citation1996; Krauss & Johanson, Citation2000). Our results suggest, for low-altitude locations, that fruit susceptibility to crown rot might be higher for certain short periods during the rainy season. This could partially explain the higher disease incidence during this period. Nevertheless, such disease incidence variations could also be explained by higher fruit contamination by the crown rot pathogens during this period (de Lapeyre de Bellaire et al., Citation2000). During the rainy season, an increase of the microbial load in the washing tanks and the airborne spore load was noticed, especially for some species of the pathogenic complex (unpublished data).
In conclusion, we have shown some potential influence of the cultivation area on the susceptibility of bananas to the crown rot disease as well as some variations within the same cultivation area. These findings show that the implementation of alternative control methods in the framework of an integrated management strategy (Lassois et al., Citation2010b ) should perform better in highlands and particularly during the drier months of the year than in the rainy season. Our results also suggest a possible influence of black leaf streak disease that should be further studied.
Acknowledgements
The authors wish to thank the agro-industrial plantations of Cameroon: ‘Plantations du Haut-Penja’ (PHP), ‘Cameroon Development Corporation-Del Monte’ (CDC-Del Monte) and ‘Societé des plantations du Manengouba’ (SPM) for their collaboration in this project. They equally wish to thank the dynamic team of the CARBAP Phytopathology Unit at Njombé (Oscar Nguidjo, Robert Dongmo, Flauribert Tchipé, Sakio Kana, Joseph Essomé, Clément Essoh, Madeleine Ntombé, Henri Mpouli, Adolphe Moussissa, Lévi Noumtcheu and Thaddée Nkwe) for their assistance in this project.
References
- Abadie , C. , Hubert , O. , Ngando , J.E. , Ngoh , G. , Mbéguié-A-Mbéguié , D. , De Lapeyre De Bellaire , L. and Chillet , M. Evidence of the effects of Mycosphaerella leaf spot diseases on fruit quality . Memories of XVIIh ACORBAT Meeting . Guayaquil , Ecuador . Edited by: Borja , J.S. , Nogales , C. , Orrantia , C. , Paladines , R. , Quimi , V. and Tazan , L. 10–14 November 2008 . ACORBAT International .
- Bastiaanse , H. , De Lapeyre De Bellaire , L. , Lassois , L. , Misson , C. and Jijakli , M.H. 2010 . Integrated control of crown rot of banana with Candida oleophila strain O, calcium chloride and modified atmosphere packaging . J. Biol. Control , 53 : 100 – 107 .
- Bolton , M.D. , Kolmer , J.A. , Xu , W.W. and Garvin , D.F. 2008 . Lr34-mediated leaf rust resistance in wheat: Transcript profiling reveals a high energetic demand supported by transient recruitment of multiple metabolic pathways . Mol. Plant–Microbe Interact , 21 : 1515 – 1527 .
- Chillet , M. and De Lapeyre De Bellaire , L. 1996 . Elaboration de la qualité des bananes au champ. Détermination de critères de mesure . Fruits , 51 : 317 – 326 .
- Chillet , M. , De Lapeyre De Bellaire , L. , Dorel , M. , Joas , J. , Dubois , C. , Marchal , J. and Perrier , X. 2000 . Evidence for the variation in susceptibility of bananas to wound anthracnose due to Colletotrichum musae and the influence of edaphic conditions . Sci. Hortic , 86 : 33 – 47 .
- Chillet , M. , Hubert , O. , Rives , M.J. and De Lapeyre De Bellaire , L. 2006 . Effects of the physiological age of bananas on their susceptibility to wound anthracnose due to Colletotrichum musae . Plant Dis , 90 : 1181 – 1185 .
- Chillet , M. , Hubert , O. and De Lapeyre De Bellaire , L. 2007 . Relationship between physiological age, ripening and susceptibility of banana to wound anthracnose . Crop Prot , 26 : 1078 – 1082 .
- Chillet , M. , Abadie , C. , Hubert , O. , Chilin-Charles , Y. and De Lapeyre De Bellaire , L. 2009 . Sigatoka disease reduces the greenlife of bananas . Crop Prot , 28 : 41 – 45 .
- De Lapeyre De Bellaire , L. and Dubois , C. 1997 . Distribution of thiabendazole resistant Colletotrichum musae isolates from Guadeloupe banana plantations . Plant Dis , 81 : 1378 – 1383 .
- De Lapeyre De Bellaire , L. and Nolin , J. 1994 . Amélioration du contrôle du chancre sur les bananes d'exportation et traitements post-récolte . Fruits , 49 : 179 – 185 .
- De Lapeyre De Bellaire , L. , Chillet , M. , Dubois , C. and Mourichon , X. 2000 . Importance of different sources of inoculum and dispersal methods of conidia of Colletotrichum musae, the causal agent of banana anthracnose, for fruit contamination . Plant Pathol , 49 : 782 – 790 .
- Equiza , M.A. , Miravé , J.P. and Tognetti , J.A. 1997 . Differential root vs. shoot growth inhibition and its relationship with carbohydrates accumulation at low temperature in different wheat cultivars . Ann. Bot , 80 : 657 – 663 .
- Finlay , A.R. and Brown , A.E. 1993 . The relative importance of Colletotrichum musae as a crown rot pathogen on Windward Island bananas . Plant Pathol , 42 : 67 – 74 .
- Finlay , A.R. , Lubin , C. and Brown , A.E. 1992 . The banana stalk as a source of inoculum of fungal pathogens which cause crown rot . Trop. Sci , 32 : 343 – 352 .
- Ganry , J. and Chillet , M. 2008 . Methodology to forecast the harvest date of banana bunches . Fruits , 63 : 371 – 373 .
- Gauhl , F. , Pasberg-Gauhl , F. , Vuylsteke , D. and Ortiz , R. 1993 . Multilocational evaluation of black sigatoka resistance in banana and plantain , 59 Ibadan , Nigeria : IITA Research Guide no. 47. IITA . pages
- Johanson , A. and Blazquez , B. 1992 . Fungi associated with bananas crown rot on field-packed fruit from the Windward Islands and assessment of their sensitivity to the fungicides thiabendazole, prochloraz and imazilil . Crop Prot , 11 : 79 – 83 .
- Jones , D.R. 2000 . Diseases of Banana, Abaca and Enset , 544 Wallingford (UK) : CABI . pages
- Jullien , A. , Chillet , M. and Malezieux , E. 2008 . Pre-harvest growth and development, measured as accumulated degree days, determine the post-harvest green life of banana fruit . J. Hortic. Sci. Biotech , 83 : 506 – 512 .
- Krauss , U. and Johanson , A. 2000 . Recent advances in the control of crown rot of banana in the Windward Islands . Crop Prot , 19 : 151 – 160 .
- Lassois , L. , De Lapeyre De Bellaire , L. and Jijakli , M.H. 2008 . Biological control of crown rot of bananas with Pichia anomala strain K and Candida oleophila strain O . Biol. Contr , 45 : 410 – 418 .
- Lassois , L. , Bastiaanse , H. , Chillet , M. , Jullien , A. , Jijakli , M.H. and De Lapeyre De Bellaire , L. 2010a . Hand position on the bunch and source-sink ratio influence the banana fruit susceptibility to crown rot disease . Ann. Appl. Biol , 156 : 221 – 229 .
- Lassois , L. , Jijakli , M.H. , Chillet , M. and De Lapeyre De Bellaire , L. 2010b . Crown rot of bananas: pre-harvest factors involved in post-harvest disease development and integrated control methods . Plant Dis , 94 : 648 – 658 .
- Lukezic , F.L. , Kaiser , W.J. and Martinez , M.M. 1967 . The incidence of crown rot of boxed bananas in relation to microbial populations of the crown tissue . Can. J. Bot , 45 : 413 – 421 .
- Moyer , M.M. , Gadoury , D.M. , Cadle-Davidson , L. , Dry , I.B. , Magarey , P.A. , Wilcox , W.F. and Seem , R.C. 2010 . Effects of acute low temperature events on development of Erysiphe necator and susceptibility of Vitis vinifera . Phytopathology , 100 : 1240 – 1249 .
- Pollack , C.J. 1990 . The response of plants to temperature change . J. Agric. Sci. (Cambridge) , 115 : 1 – 5 .
- Shillingford , C.A. 1977 . Control of banana fruit rots and of fungi that contaminate washing water . Trop. Sci , 19 : 197 – 203 .
- Shillingford , C.A. 1978 . Climatic factors affecting post-harvest decay of Jamaican bananas . J. Agric. Univ. Puerto Rico , 63 : 45 – 49 .