Abstract
One hundred and sixty-nine wheat leaf samples infected with leaf rust (Puccinia triticina Eriks.) were collected across Canada in 2009. Three hundred and ten single uredinial isolates were analysed for their virulence spectra. The most common virulence phenotypes identified in Canada were TDBJ (28.1%), MLDS (20.6%) and TDBG (15.8%). There were eight virulence phenotypes identified among 12 isolates from Alberta and British Columbia – the most common were TDBJ (four isolates) and MBBJ (two isolates). There were 20 unique virulence phenotypes identified among 269 isolates from Manitoba and Saskatchewan. The most common virulence phenotypes were TDBJ (30%), MLDS (23%) and TDBG (18%). In Ontario, there were 13 virulence phenotypes identified from 20 isolates – the most common were TBRK (four isolates), MCGJ (three isolates), NBBT and TDBJ (two isolates each). Each of the four isolates from Quebec was a unique virulence phenotype – TBJS, MCPS, TBHK and TBPK. In Prince Edward Island, there were four virulence phenotypes (MCQJ, MCNQ, MCQK and MHNQ) identified among five isolates. The regions were different from each other for most common virulence phenotypes, but some phenotypes were in common between western Canada and Ontario. Many phenotypes were unique to a particular region and some were not found in previous Canadian surveys. Virulence phenotypes in eastern and western Canada differed for virulence to genes Lr2a, Lr2c and Lr18.
Résumé
En 2009, 169 échantillons de feuilles de blé infectées par la rouille brune (Puccinia triticina Eriks.) ont été collectés partout au Canada. Trois cent dix isolats mono-urédiniaux ont été analysés en fonction de leurs spectres de virulence. Les phénotypes de virulence identifiés le plus couramment au Canada ont été TDBJ (28,1 %), MLDS (20,6 %) et TDBG (15,8 %). On a identifié 8 phénotypes de virulence parmi 12 isolats provenant d'Alberta et de Colombie-Britannique, les plus communs étant TDBJ (4 isolats) et MBBJ (2 isolats). On a identifié 20 phénotypes de virulence uniques parmi 269 isolats provenant du Manitoba et de la Saskatchewan. Les phénotypes les plus communs étaient TDBJ (30 %), MLDS (23 %) et TDBG (18 %). En Ontario, 13 phénotypes de virulence ont été identifiés à partir de 20 isolats, les plus courants étant TBRK (4 isolats), MCGJ (3 isolats) ainsi que NBBT et TDBJ (2 isolats chacun). Chacun des quatre isolats provenant du Québec correspondait à un phénotype de virulence unique : TBJS, MCPS, TBHK et TBPK. À l’Île-du-Prince-Édouard, on a identifié quatre phénotypes de virulence parmi cinq isolats : MCQJ, MCNQ, MCQK et MHNQ. En ce qui a trait à la plupart des phénotypes les plus courants, les provinces différaient les unes des autres, mais certains phénotypes étaient communs à l'Ouest canadien et à l'Ontario. Plusieurs phénotypes étaient propres à une région en particulier et certains n'avaient jamais été détectés au cours d’études canadiennes précédentes. Les phénotypes de virulence de l'est et de l'ouest du Canada différaient sur le plan de la virulence quant aux gènes Lr2a, Lr2c et Lr18.
Introduction
There were approximately 10 million ha seeded to wheat in Canada during 2009, including 6.8 million ha of hexaploid spring wheat (Triticum aestivum L.), 2.2 million ha of durum wheat (Triticum turgidum L. ssp. durum (Desf.) Husn.), and 0.3 million ha of winter wheat (T. aestivum) (Anonymous, Citation2012). Each of these wheat types (spring, winter and durum) are further subdivided into various classes based on end-use quality parameters, such as bread and pasta making attributes (McCallum & DePauw, Citation2008). Canada western red spring (CWRS) wheat accounted for approximately 6.1 million ha of the hexaploid spring wheat area planted, with other wheat classes accounting for the remaining seeded area. The most commonly grown CWRS cultivars in 2009 were ‘Lillian’, ‘Harvest’ and ‘Superb’, grown on 17.5%, 15.9% and 8.5%, respectively, of the total area seeded to CWRS cultivars (Anonymous, Citation2009). The genetics of leaf rust resistance in ‘Lillian’ and ‘Harvest’ have not been determined, although it appears that ‘Lillian’ has Lr30 and Lr34 along with other genes (McCallum et al., Citation2011a ; B. McCallum, unpublished data), while ‘Harvest’ has the non-functional allele for Lr34. ‘Superb’ was demonstrated to have Lr2a and Lr10 (McCallum & Seto-Goh, Citation2010 a). In inoculated field trials in 2009, ‘Lillian’ was resistant, ‘Harvest’ was moderately resistant and ‘Superb’ was susceptible. There was also 0.52 million ha seeded to wheat in eastern Canada, with winter wheat in Ontario accounting for 0.40 million ha and the rest primarily spring wheat in Ontario and the other eastern provinces (Anonymous, Citation2012).
Wheat leaf rust, caused by Puccinia triticina Eriks. (Anikster et al., Citation1997) (= P. recondita Rob. ex Desmaz. f. sp. tritici), is a historical and current production problem on wheat in Canada and other parts of the world. The disease reduces yield by 5–25%, depending on the susceptibility of the crop, fungicide application and the timing of infection (McCallum et al., Citation2007). Virulence surveys have been conducted annually in Canada since 1931 to monitor changes in the P. triticina population. These surveys can detect new virulence phenotypes, provide a historic record of shifts in virulence to specific resistance genes, and detect groups of genetically similar isolates (Wang et al., Citation2010). Knowledge on the virulence of the P. triticina population and the resistance genes in Canadian wheat cultivars can be used to make informed decisions on breeding and developing new wheat cultivars with effective resistance genes (McCallum & DePauw, Citation2008).
The main objective of this study was to determine the frequency and distribution of virulence phenotypes of P. triticina collected across Canada, using 16 near-isogenic wheat lines inoculated at the seedling stage. A second objective was to determine the virulence spectrum for a set of representative isolates containing all the virulence phenotypes found in 2009, on six additional near-isogenic wheat lines with adult plant resistance genes, and 12 additional near-isogenic lines with seedling resistance.
Materials and methods
Infected wheat leaves were collected from individual fields and nurseries from June–September, 2009 in various locations throughout Prince Edward Island (PEI), Ontario (ON), Quebec (QC), Manitoba (MB), Saskatchewan (SK), Alberta (AB) and British Columbia (BC). The leaves were air dried at 20–27 °C for approximately 12 to 24 h and stored separately for 2–4 months at 5 °C. Puccinia triticina urediniospores from individual collections were subsequently scraped off from infected leaves using a metal spatula and a small amount of water, and inoculated onto the susceptible wheat cultivar ‘Little Club’ by rubbing the leaves with a mixture of urediniospores, water and Tween 20 as described previously (McCallum & Seto-Goh, Citation2005). Each pot of seedlings was pretreated with 50 mL maleic hydrazide solution (0.36 g L−1) approximately five days after seeding to prevent the emergence of secondary leaves and to produce larger uredinia with abundant sporulation. A plastic cone, approximately 25 cm in height, with an open top was placed over each pot to minimize cross-contamination. Inoculated plants were placed into a dew chamber (Model I-60D, Percival Scientific, Perry, IA) with nearly 100% relative humidity at 17 °C for approximately 17 h to allow the urediniospores to germinate and initiate the infection process. Plants were then placed in a greenhouse at 20 ± 4 °C with supplemental high-pressure sodium lighting. Approximately seven days after inoculation, chlorotic spots appeared, indicating areas of infection. The leaves were then trimmed so that single isolated uredinia remained on the upper edge of each trimmed leaf. Cross-contamination was minimized by removing all extra leaves. At approximately 14 days after inoculation, urediniospores were collected from a single isolated uredinium into a 00 gelatin capsule, using a vacuum suction micro-collector, mixed with a light mineral oil (Bayol, Esso Canada, Toronto, ON) and sprayed onto a 7-day-old set of wheat seedlings which included ‘Thatcher’ and 16 single resistance gene ‘Thatcher’ near-isogenic lines. A pot of ‘Thatcher’ seedlings was used to increase the isolate for subsequent inoculations and long-term storage. Two single uredinial isolates were typically evaluated from each rust collection, although sometimes one or three isolates per collection were analysed. Approximately 12 seeds of each ‘Thatcher’ near-isogenic line were planted in a clump and the clumps were evenly spaced in a fibre flat (25 × 15 cm). Plants were pretreated with maleic hydrazide as described above. After inoculation, the plants were dried for at least 1 h to allow the oil to evaporate and then incubated and maintained as described above. Infection types produced on the 12 standardized differentials [Set 1: Lr1 (RL6003a), Lr2a (RL6016), Lr2c (RL6047), Lr3 (RL6002); Set 2: Lr9 (RL6010), Lr16 (RL6005), Lr24 (RL6064), Lr26 (RL6078); Set 3: Lr3ka (RL6007), Lr11 (RL6053), Lr17 (RL6008), Lr30 (RL6049)] were used to determine the three-letter code according to the virulence phenotype nomenclature (Long & Kolmer, Citation1989). Four supplemental differentials [Set 4: LrB (RL6051), Lr10 (RL6004), Lr14a (RL6013), Lr18 (RL6009)] were added to provide additional virulence information about the isolates, resulting in a four-letter code. All isolates were also inoculated onto ‘Thatcher’ and Lr1 +LrCen (RL6003b). The resistance gene temporarily named LrCen was previously identified in the ‘Thatcher-Lr1’ near-isogenic line RL6003 (McCallum & Seto-Goh, Citation2006b). Infection types to all the differential near-isogenic lines were rated 12 days after inoculation. Isolates that produced infection types ‘;’ (hypersensitive flecks), ‘1’ (small uredinia with necrosis), and ‘2’ (small- to medium-sized uredinia with chlorosis) were considered avirulent and those that produced infection types ‘3’ (medium-sized uredinia without chlorosis or necrosis) and ‘4’ (large uredinia without chlorosis or necrosis) were considered virulent (Long & Kolmer, Citation1989). Inoculations were repeated if the infection response was not clear.
At least one isolate from most of the unique virulence phenotypes identified was inoculated onto adult plants of ‘Thatcher’ and six ‘Thatcher’ near-isogenic lines (Lr12 [RL6011], Lr13 [RL4031], Lr22a [RL6044], Lr35 [RL6082], and Lr37 [RL6081]). In previous studies, selected isolates were also tested on the ‘Thatcher-Lr34’ near-isogenic line but all isolates had nearly the same avirulent reaction on this line so it was omitted. Single plants of each line were grown together in a 15 cm diameter pot in the greenhouse as described previously. Plants were trimmed so that only two culms per plant remained, and flag leaves of all plants within a pot were inoculated with a single uredinial isolate. The inoculated plants were incubated overnight at 17 °C in a dew chamber and grown in the greenhouse at 20 ± 4 °C as described previously. Infection types were evaluated 14 days after inoculation. This subset of isolates was also tested on 12 additional ‘Thatcher’ near-isogenic lines at the seedling stage (Lr2b [RL6019], Lr3bg [RL6042], Lr14b [RL6006], Lr15 [RL6052], Lr19 [RL6040], Lr20 [RL6092], Lr21 [RL6043], Lr23 [RL6012], Lr25 [RL6084], Lr28 [RL6079], Lr29 [RL6080], Lr32 [RL6086], and Lr52). These isolates were also retested on seedling plants of the set of 16 ‘Thatcher’ near-isogenic lines mentioned previously to confirm the reactions using the methods for seedlings described previously.
Results and discussion
During 2009, the growing season in western Canada was relatively cool, which slowed the development of wheat leaf rust. The first observance of leaf rust in Manitoba was during early July, which is much later than normal (McCallum & Seto-Goh, Citation2010b ). There was also less inoculum coming into western Canada from the north-central United States than would normally be the case due to non-conducive weather and wind patterns for spread (Kolmer et al., Citation2011). The net result was a very low level of wheat leaf rust infection in western Canada compared with other years.
In Ontario, leaf rust was commonly observed only after the first week of July when wheat crops were at the milk to dough development stages. The disease was at low to medium levels of infection and did not result in significant yield reductions (Xue & Chen, Citation2010). The relatively low temperatures and frequent periods of rain in June and early July, and the high temperatures in late July and August, were likely responsible for the widespread infection but relatively low severity of leaf rust observed in Ontario.
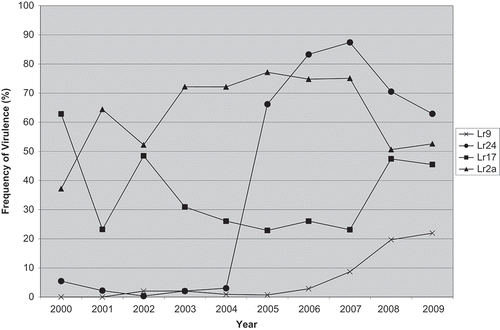
There were 169 leaf samples infected with P. triticina collected throughout Canada for analysis. From these samples, 310 single uredinial isolates were analysed for their virulence spectra (). The population from MB and SK made up 269 of the total 310 isolates from across Canada. As a result, the most predominant isolates in Canada were the same as those from MB and SK, with TDBJ at 28.1%, MLDS at 20.6% and TDBG at 15.8%. This was similar to the situation in 2008 (). The virulence phenotypes TDBJ and TDBG were the most common phenotypes between 2006 and 2008, but in 2009, MLDS became the second most frequently found virulence phenotype, and has been steadily increasing in proportion since 2007 (). There were eight virulence phenotypes identified among the 12 isolates from AB and BC – the most common were TDBJ (four isolates) and MBBJ (two isolates). All the virulence phenotypes found in AB and BC were also found in other regions of Canada, except MCBS which was only found in this region. There were two virulence phenotypes (NBBT and MDPS) found in 2009 that have not been found in this region in the past 14 years.
Table 1. Frequency and distribution of virulence phenotypes of Puccinia triticina identified by infection types to selected resistance genes in Canada in 2009
Table 2. Frequency (%) of predominant Puccinia triticina virulence phenotypes in Canada from 2000 to 2009
There were 20 unique virulence phenotypes identified among the 269 isolates from MB and SK. The most common virulence phenotypes were TDBJ (30%), MLDS (23%) and TDBG (18%). These were also the most common in this region in 2008 at 29.1%, 20.5% and 28.5%, respectively. There were seven virulence phenotypes found in MB and SK in 2009 that were not found in this region for the past 14 years (PBDQ, TBJS, MJDS, MNDS, TMGJ, TNBQ and TNPS). The most common virulence phenotypes found in the North Central United States in 2009 were MLDS (44.9%) and TDBG (37.2%), similar to those found in MB and SK, as the North Central United States is the main source of inoculum for this area (Kolmer et al., 2011).
In ON there were 13 virulence phenotypes identified from 20 isolates – the most common were TBRK (four isolates), MCGJ (three isolates), NBBT and TDBJ (two isolates each). From a larger sample (71 isolates) in 2008, the most common phenotypes identified were MFDS (28.2%), MLDS (8.5%), MBTN and MDDS (5.6% each). This could indicate a change in the ON population; however, the 2009 sample is relatively small to permit firm conclusions. There were four virulence phenotypes found in 2009 in ON that were not found in the previous 14 years – MCGJ, NBBT, TCRK and TFTS.
Each of the four isolates from QC was a unique virulence phenotype – TBJS, MCPS, TBHK and TBPK. None of these virulence phenotypes were found in QC during the past 11 years. None of these virulence phenotypes were found outside of QC in 2009 except TBJS. The population from Quebec appeared to be fairly distinct from the populations in both ON and PEI, as in previous years.
In PEI there were four virulence phenotypes (MCQJ, MCNQ, MCQK and MHNQ) identified among five isolates. Of these, MCNQ and MCQJ were not found in the previous three years from PEI although, similar phenotypes were found previously in Canada. All of these virulence phenotypes were unique to PEI, which demonstrates the differentiation of the PEI population from the other Canadian populations. This has been observed each year since 2006 in which samples from PEI were first analysed (McCallum & Seto-Goh, Citation2009; McCallum et al., Citation2010, Citation2011b ).
Virulence data on each of the differential lines in each region of Canada is presented in . The number of isolates sampled in some regions was too low to determine if meaningful shifts in the virulence of the population had occurred from previous years. However, the MB and SK sample with 269 isolates can be analysed for virulence changes in the population. Virulence to Lr9 increased slightly from 2008 which follows a trend of increasing virulence since 2005 (). Virulence to both Lr24 and Lr17 was similar to that seen in 2008, although virulence to both of these resistance genes had been fluctuating over recent years. Virulence to Lr2a declined between 2007 and 2008 due to the increase in the MLDS virulence phenotype which is avirulent to Lr2a, but was relatively stable between 2008 and 2009 (). Virulence to Lr18 was relatively common in ON and QC compared with western Canada where it was rare (). Alternatively, virulence to Lr24 was common in MB and SK (68%) whereas it was much lower in ON (30%) and not found in QC or PEI.
Table 3. Frequencies of virulence of Puccinia triticina in Canada in 2009 to lines of wheat with single Lr genes for leaf rust resistance
Fig. 1. Frequency of virulence (%) from 2000–2009 in the Manitoba and Saskatchewan population of P. triticina to near-isogenic lines containing Lr2a, Lr14a, Lr16, Lr17 or Lr24.
Eighty-one isolates were selected to represent each of the virulence phenotypes found in each region. These isolates were tested on 13 supplementary differentials at the seedling stage (Lr2b, Lr3bg, Lr14b, Lr15, Lr19, Lr20, Lr21, Lr23, Lr25, Lr28, Lr29, Lr32 and Lr52) and on five differential lines at the adult plant stage (Lr12, Lr13, Lr22a, Lr35 or Lr37). To present the results, isolates with the same virulence phenotype from the same region are represented by a single isolate in and therefore only 52 isolates of the 81 tested are shown. For the 13 supplementary seedling differentials, all isolates were virulent to Lr15 (except for 09-163-3 NBBT), avirulent to Lr19, Lr21, Lr29, Lr32, Lr52 and varied for virulence to Lr2b, Lr3bg, Lr14b, Lr20, Lr23, Lr25, Lr28. All isolates virulent to Lr2a were also virulent to Lr2b and all isolates avirulent to Lr2a were also avirulent to Lr2b. All isolates were virulent to Lr14b except five: 09-121-1 PBDQ, 09-121-2 PBDQ, 09-124-1 PBDQ, 09-137-2 MCTN, 09-140-3 MCGS. All isolates were avirulent to Lr25 except: 09-121-1 PBDQ, 09-121-2 PBDQ and 09-124-1 PBDG. Seventy-two of the isolates were virulent to Lr20, 62 were virulent to Lr3bg, 43 were virulent to Lr23 and 42 were virulent to Lr28 of the 81 isolates tested ().
Table 4. Infection types of 52 representative isolates from Canada in 2009 tested on near isogenic lines of ‘Thatcher’ with adult-plant resistance genes and additional seedling resistance genes
For the adult plant differential tests all isolates were avirulent to Lr22a and Lr35 (except for 09-7-1 MLDS, 09-164-1 MLDS, 09-164-4 TBJS, 09-168-1 TDBJ which were virulent to Lr35 but avirulent to Lr22a). There were 74 virulent isolates to Lr12, 75 virulent isolates for Lr13 and 58 virulent isolate for Lr37 of the 81 isolates tested ().
Overall, the leaf rust populations found in Canada during 2009 were similar to those found in previous years. There were a number of new virulence phenotypes found in each region, although the frequency of virulence to the resistance genes tested did not change significantly from 2008. The population in AB and BC shared most virulence phenotypes with MB and SK; however, there were some phenotypes unique to this region. The ON and QC populations were different from each other and from the rest of Canada. There were some virulence phenotypes in common between Ontario and western Canada but most were unique to Ontario. As in previous years, the PEI population was different from all other regions in Canada and contained unique virulence phenotypes.
Acknowledgements
We thank André Comeau, Harpinder Randhawa and Richard Martin for providing samples for analysis, and other cooperators who grew trap rust nurseries.
Notes
Related Research Data
References
- Anikster , Y. , Bushnell , W.R. , Eilam , T. , Manisterski , J. and Roelfs , A.P. 1997 . Puccinia recondita causing leaf rust on cultivated wheats, wild wheats, and rye . Can. J. Bot. , 75 : 2082 – 2096 .
- Anonymous (2009). Canadian Wheat Board, wheat variety survey 2009.the Canadian Wheat Board http://www.cwb.ca (http://www.cwb.ca)
- Anonymous . 2012 . Statistics Canada . Field crop reporting series – 3401. Table 001–0010. ,
- Kolmer , J.A. , Long , D.L. and Hughes , M.E. 2011 . Physiologic specialization of Puccinia triticina on wheat in the United States in 2009 . Plant Dis. , 95 : 935 – 940 .
- Long , D.L. and Kolmer , J.A. 1989 . A North American system of nomenclature for Puccinia recondita f. sp. tritici . Phytopathology , 79 : 525 – 529 .
- McCallum , B.D. and Depauw , R.M. 2008 . A review of wheat cultivars grown in the Canadian prairies . Can. J. Plant Sci. , 88 : 649 – 677 .
- McCallum , B.D. and Seto-Goh , P. 2003 . Physiologic specialization of wheat leaf rust (Puccinia triticina) in Canada in 2000 . Can. J. Plant Pathol. , 25 : 91 – 97 .
- McCallum , B.D. and Seto-Goh , P. 2004 . Physiologic specialization of Puccinia triticina, the cause of wheat leaf rust, in Canada in 2001 . Can. J. Plant Pathol. , 26 : 109 – 120 .
- McCallum , B.D. and Seto-Goh , P. 2005 . Physiologic specialization of wheat leaf rust (Puccinia triticina) in Canada in 2002 . Can. J. Plant Pathol. , 27 : 90 – 99 .
- McCallum , B.D. and Seto-Goh , P. 2006a . Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2003 . Can. J. Plant Pathol. , 28 : 208 – 213 .
- McCallum , B.D. and Seto-Goh , P. 2006b . Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf, in Canada in 2004 . Can. J. Plant Pathol. , 28 : 566 – 576 .
- McCallum , B.D. and Seto-Goh , P. 2008 . Physiologic specialization of Puccinia triticina in Canada in 2005 . Can. J. Plant Pathol. , 30 : 124 – 132 .
- McCallum , B.D. and Seto-Goh , P. 2009 . Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2006 . Can. J. Plant Pathol. , 31 : 80 – 87 .
- McCallum , B.D. and Seto-Goh , P. 2010a . The inheritance of leaf rust resistance in the wheat cultivars ‘Superb’, ‘McKenzie’ and ‘HY644’ . Can. J. Plant Pathol. , 32 : 387 – 395 .
- McCallum , B.D. and Seto-Goh , P. 2010b . Leaf rust and stripe rust of wheat in Manitoba and eastern Saskatchewan in 2009 . Can. Plant Dis. Surv. , 90 : 108
- McCallum , B.D. , Fetch , T. and Chong , J. 2007 . Cereal rust control in Canada . Aust. J. Agric. Res. , 58 : 639 – 647 .
- McCallum , B.D. , Seto-Goh , P. and Xue , A. 2010 . Physiological specialization of Puccinia triticina in Canada in 2007 . Can. J. Plant Pathol. , 32 : 229 – 236 .
- McCallum , B.D. , Humphreys , D.G. , Somers , D.J. , Dakouri , A. and Cloutier , S. 2011a . Allelic variation for the rust resistance gene Lr34/Yr18 in Canadian wheat cultivars . Euphytica , 183 : 261 – 274 .
- McCallum , B.D. , Seto-Goh , P. and Xue , A. 2011b . Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2008 . Can. J. Plant Pathol. , 33 : 541 – 549 .
- Wang , X. , Bakkeren , G. and McCallum , B. 2010 . Virulence and molecular polymorphisms of the wheat leaf rust fungus Puccinia triticina in Canada from 1997 to 2007 . Botany , 88 : 575 – 589 .
- Xue, A.G., & Chen, Y. (2010). Disease of spring wheat in eastern Ontario in 2009. Can. Plant Dis. Surv., 90, 114–115. http://www.cps-scp.ca/cpds.shtml (http://www.cps-scp.ca/cpds.shtml)