Abstract
Soil population densities and spatial patterns of sclerotia produced by Sclerotinia sclerotiorum were determined in six commercial lettuce production fields with varying histories of lettuce drop in Yuma County, AZ. Each field was divided into eight approximate equal sections and in each section, a plot (10 × 20 m) was randomly selected for soil sampling. Soil cores were collected along both diagonals at 2 m intervals in each plot and sclerotia populations were determined by wet sieving. The average soil sclerotia population per plot ranged from 0.0 to 5.8/100 g soil. The highest and the lowest percent of individual soil samples with ≥ 1 sclerotia/100 g soil per plot were 63% and 7%, respectively. Three indices of dispersions – variance/mean, Morisita’s index and Lloyd’s index – calculated for each of the six fields indicated that sclerotia distributions were aggregated. However, in none of the fields could the sclerotia distributions be adequately described by the Poisson, negative binomial or Neyman type A distributions.
Résumé
Les densités des populations vivant dans le sol et les profils spatiaux des sclérotes produits par Sclerotinia sclerotiorum ont été déterminés dans six champs d’Yuma County, en Arizona, où l’on produit commercialement de la laitue et dans lesquels l’historique de la pourriture sclérotique de la laitue diffère. Chaque champ a été divisé en huit sections à peu près égales et, dans chaque section, une parcelle (10 m × 20 m) a été choisie aléatoirement en vue d’en échantillonner le sol. Les carottes de sol ont été prélevées dans chaque parcelle, à tous les 2 m, sur les deux diagonales, et les populations de sclérotes ont été identifiées par criblage par voie humide. La population moyenne de sclérotes par parcelle variait de 0,0 à 5,8/100 g de sol. Le pourcentage le plus élevé d’échantillons de sol affichant un sclérote ou plus par 100 g de sol par parcelle était de 63% et le plus bas, de 7%. Trois indices de dispersion – moyenne et variance, indice de Morisita et indice de Lloyd – , calculés pour chacun des six champs, ont indiqué que les distributions de sclérotes étaient agrégatives. Toutefois, les distributions de sclérotes n’ont pu être adéquatement décrites dans aucun des champs par la courbe de Poisson, par distribution binomiale négative ou par distribution de Neyman type A.
Introduction
Lettuce drop is one of the most destructive diseases of lettuce (Lactuca sativa L.) and has been reported in all lettuce-growing regions of the world (Dillard & Grogan, Citation1985; Subbarao, Citation1998). In the USA, the disease regularly occurs in the two primary lettuce-producing states of Arizona and California. Yield losses vary from 1% to nearly 75% depending on conditions, but under ideal disease conditions an entire field may be lost (Purdy, Citation1979; Subbarao, Citation1998). The disease is caused by two closely related soilborne fungi, Sclerotinia sclerotiorum (Lib.) de Bary and S. minor Jagger. Although both fungi are present in the lettuce-growing areas of Arizona and California, their distribution is somewhat distinct. In the cooler region of coastal California, S. minor is the predominant species. However, in the desert production areas of Yuma Co, Arizona, and Imperial Co, California, which are quite distinctive from other western production areas in regard to soil and climate, S. sclerotiorum is the predominant species (Subbarao, Citation1998).
Both fungi produce sclerotia, masses of hyphae surrounded by a tough melanin layer, that serve as both survival structures and primary inoculum for subsequent lettuce crops. Sclerotia of S. minor infect the host directly by producing mycelium. However, sclerotia of S. sclerotiorum can infect the host either directly by producing mycelium or indirectly by producing apothecia and subsequently ascospores (Subbarao et al., Citation1994; Chitrampalam et al., Citation2008). In desert production areas of Arizona and California, the specific conditions required to prime the sclerotia to produce apothecia are not commonly encountered and thus, the disease is primarily initiated by myceliogenic germination of sclerotia (Subbarao, Citation1998; Matheron & Porchas, Citation2004).
Sclerotia of S. sclerotiorum can survive up to seven years and their longevity is affected by location of sclerotia within the soil profile, duration of burial and soil temperature and moisture (Subbarao et al., Citation1994; Matheron, & Porchas, Citation2005). Sclerotia at depths greater than 10 cm have reduced viability and are generally not capable of causing disease due to unfavourable O2 and CO2 levels, low soil temperature, high soil moisture, and colonization of sclerotia by antagonistic microorganisms (Grogan et al., Citation1980; Imolehin & Grogan, Citation1980; Subbarao, Citation1998). In contrast, sclerotia within 8 cm of the soil surface and 2 cm of the lettuce taproot have high rates of successful infection (Subbarao et al., Citation1994).
Commercially acceptable cultivars with adequate levels of resistance are not currently available. Thus, current management strategies for lettuce drop rely primarily on fungicides such as iprodione and boscalid. However, the level of disease management achieved with fungicides is not dramatic and is also inconsistent (Matheron & Porchas, Citation2005; Chitrampalam et al., Citation2008). A greater reduction in disease may be achieved with an integrated disease management approach, which includes an increased focus on reducing sclerotia populations in the soil.
To effectively assess the impact of soilborne sclerotia on lettuce drop and assess the impact of management strategies on sclerotial populations, estimates of soilborne sclerotia populations need to be available. For effective sclerotia sampling and reliable density estimations to be possible, preliminary information regarding densities and distribution in commercial fields are needed. In previous studies on soil sclerotial distribution for various soilborne fungi, including Macrophomina phaseolina (Tassi) Goid., Verticillium dahlia Kleb. and Sclerotinia minor (Smith & Rowe, Citation1984; Dillard & Grogan, Citation1985; Mihail & Alcorn, Citation1987), results revealed that soilborne sclerotia exhibited aggregated spatial distributions to varying degrees depending on differences in ecology among fungal species. However, no such studies have been conducted for S. sclerotiorum. Although work by Hao & Subbarao Citation(2003) investigated mean soil populations of sclerotia in California fields and the associated aggregated pattern of disease incidence, that study did not describe the underlying soil distribution patterns of sclerotia. This deficiency in descriptive data is even more acute in regard to the unique desert production areas of the southwestern USA and its effect on the ecology of S. sclerotiorum.
Sampling for soilborne pathogens usually involves collecting soil samples at intervals along one of several pre-determined paths through a field such as an X, W or diagonal pattern (Lin et al., Citation1979; Nicot et al., Citation1984). Alternatively, sampling sites can be selected by using simple random sampling, stratified random sampling, cluster sampling or other survey sampling techniques (Mihail & Alcorn, Citation1987). Sampling methods (random vs. cluster sampling) become more important than sampling size when disease is aggregated. In previous studies, sampling along X-shaped paths covering a smaller demarcated area within the field provided information about the area for both random and aggregated pathogen populations in computer-simulated fields (Lin et al., Citation1979). The same study also suggested that where the population is aggregated, samples should be taken from multiple areas across the entire field.
The objective of this study was to determine soil population densities of sclerotia of S. sclerotiorum in the prime winter lettuce production areas around Yuma, AZ, in fields with varied histories of disease incidences. Spatial patterns of sclerotia in soil were established along with mean aggregate size for those populations determined to have a clumped distribution. These data are required for the subsequent development of general soil sampling strategies necessary for improved lettuce drop disease management in desert agroecosystems.
Materials and methods
Field sampling
Six lettuce fields of approximately 8 hectares, each with a history of lettuce drop, were selected for sampling. All fields were located in the prime winter lettuce production area around Yuma, AZ, and were regularly cropped to lettuce using conventional lettuce production practices for the area. The selection of fields was assisted by the University of Arizona Vegetable Extension Specialist, Dr. Mike Matheron, who was familiar with all fields in question. Fields A, B and C had severe outbreaks of lettuce drop in the previous season, estimated at > 50% disease incidence. Fields D, E and F had moderate levels of lettuce drop, estimated at 20–30%. The soil and climatic conditions were similar across this sampling area and all fields had been recently harvested and disc harrowed to incorporate crop residue.
The sampling strategy suggested by Lin et al. Citation(1979) for aggregated distributions was used with minor modification for this study. To determine the sclerotia distribution pattern in different parts of the field, each field was divided into eight approximate equal parts and one plot (10 × 20 m) was randomly selected in each part for sampling. In each plot, 24 soil cores were collected across both diagonals at 2 m intervals by inserting a 2.65 cm-diameter soil auger to a depth of 7 cm. Each soil sample was placed in individual paper bags with plot and diagonal position recorded and returned to the laboratory. Thus, a total of 192 samples were collected from each field.
Sclerotium population density determination
Soil samples were air dried in paper bags at room temperature and stored until analysis. Immediately prior to analysis, each sample was homogenized and mixed thoroughly. From each sample, 100 g of soil was wet sieved through a US Std. series # 18 (The W. S. Tyler Company, Ohio, USA) and the recovered sclerotia were enumerated by examination under a stereo microscope at 60× (Subbarao et al., Citation1994). For each plot, descriptive statistics such as mean, variance and range for sclerotia/100 g soil were calculated. The sampling data from all plots of each field were then combined and analysed field-wise both by individual soil cores values (192 soil cores) and by plot means (8 plot means). Analysis of variance was performed on mean sclerotia of different fields to check for significance differences in the mean population among lettuce fields (Sigmastat Software Inc., San Jose, California, USA).
Analysis of spatial pattern of sclerotia in soil
Three indices of dispersion; the variance to mean ratio, Morisita’s index of dispersion, and Lloyd’s index of patchiness (Mihail & Alcorn, Citation1987), were calculated for each plot by sample (n = 24) and for each field both by plot (n = 8) and by sample (n = 192) to test for deviation from unity (Lin et al., Citation1979). Morisita’s index of dispersion was calculated by using the following formula: Id=n [(Σ(x2) – Σx)/((Σx) 2– Σx)], where n is number of sampling units and × is the number of sclerotia per sampling unit (Mihail & Alcorn, Citation1987). Lloyd’s index of patchiness was calculated using the following formula: LIP = X*/where X* is the mean crowding and
is the mean population (Shukla et al., Citation1985). Mean crowding (X*) was calculated by X* =
((S2/
)–1) where
is the mean and S2 is the variance (Lloyd, Citation1967). Values that are significantly greater than 1 indicate an aggregated spatial pattern (Southwood, Citation1978). Significant departure of index of dispersion from 1.0 was tested using the chi-square test statistic (Ludwig & Reynolds, Citation1988).
The spatial pattern within plots and the effect of varying the size/interval of the sampling units (the distance between the two sampling points) on the detection of underlying spatial patterns were analysed using Hill’s two-term local quadrat variance method (TTLQV) (Ludwig & Reynolds, Citation1988). Data gathered from a series of contiguous sites in each diagonal across plots were grouped into different blocks by adding together adjacent sampling site data, and variance for these blocks was calculated as follows:
The Poisson, Neyman type A and negative binomial discrete frequency distributions were fit to the dataset from each field by using the PADIS statistical program (Lopez & Velazquez, Citation1997). This program assesses the degree to which the dataset was described by the particular theoretical distribution by using a chi-square goodness of fit technique. An agreement with Poisson would indicate a random spatial pattern and an agreement with the negative binomial or Neyman type A distribution would indicate an aggregate distribution (Mihail & Alcorn, Citation1987).
Determination of sclerotia viability
To determine the potential of each sclerotium collected from the field to cause disease, viability of sclerotia was ascertained for all the collected sclerotia from three of the six lettuce fields. Sclerotia were surface sterilized in 10% household bleach for 1 minute and washed twice with sterilized water. Each sclerotium was individually placed on a 2% water agar plug (1 cm diam) in a Petri plate and incubated at 20 °C for a week. The per cent sclerotia germination (viability) was calculated for each field.
Table 1. Descriptive statistics for sclerotia populations of Sclerotinia sclerotiorum in six lettuce fields
Results
Sclerotia population density determination
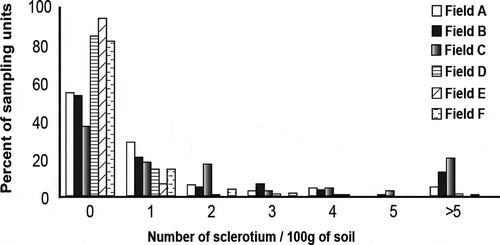
The number of sclerotia of S. sclerotiorum per 100 g of soil from lettuce production fields ranged from 0 to 42 (). More than 80% of the samples from fields D, E and F did not contain any sclerotia (). Field C and field E contained the minimum (36%) and maximum number (93%) of samples with no sclerotia, respectively. Field C had the highest percentage of samples (20%) with more than 5 sclerotia per 100 g of soil. The mean number of sclerotia per 100 g of soil for individual fields ranged from 0.08 to 2.9. Fields B and C had the highest mean number of sclerotia (2.9 sclerotia/100 g of soil) and field E had the lowest mean number of sclerotia (0.08 sclerotia/100 g soil).
Fig. 1. Frequency distribution of sclerotia densities of Sclerotinia sclerotiorum in six lettuce fields.
Analysis of spatial pattern of sclerotia in soil
When indices of dispersion were calculated per plot, all three indices indicated that sclerotia were aggregated in all plots in fields B and C (Table S1). In field A, an aggregated distribution was indicated by all three indices in five plots, by two indices in one plot, and by one index in one plot. However, in one of the eight plots in field A, an aggregate distribution was not supported by any of the three indices. In fields D, E and F, at least one of the three indices of dispersion indicated an aggregate distribution in three, one and five plots, respectively. When the indices of dispersion were calculated per field using data from individual soil cores (n = 192), all indices indicated aggregated distributions in fields A, B and C. However, when the indices were calculated per field using mean data from each plot (n = 8), no indices indicated aggregated distributions, revealing a uniformity in mean plot sclerotia population across each field (Table S1). For fields D, E and F, most indices indicated aggregated distributions of sclerotia when the indices were calculated per field using individual soil core data (n = 192). Once again, when the indices were calculated per field using mean data from each plot, no indices indicated aggregated distributions in fields D and F. However, in field E, the Morisita’s index indicated an aggregated distribution (Table S1).
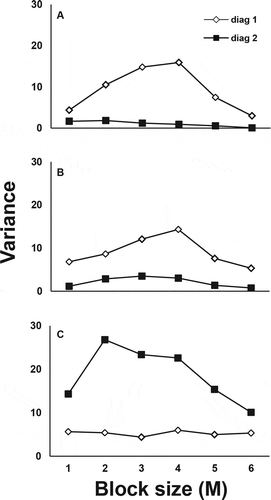
The analysis of spatial pattern within each plot by two term local quadrat (TTLQ) method revealed that sclerotial distribution was aggregated along 46% of the diagonals from all fields combined ( and ). Aggregate sizes calculated by TTLQ analysis ranged from 2 to 12 metres in diameter (). The number of sampling units (soil cores) required to estimate the field sclerotia populations with an allowable sampling error of 20% of the mean was 178, 152 and 55 for field A, B and C, respectively, which is below the number of sampling units (n = 192) used in this study (). However, the number of sampling units required to estimate the field sclerotia populations within allowable sampling error were 334, 499 and 214 for fields D, E and F, respectively, which were higher than the number of sampling units used in this study (). When estimations of field populations were based upon plot means, the number of plots required to estimate field populations was higher than the number of plots used in this study in five of six lettuce fields ().
Table 2. Distribution patterns of sclerotia of S. sclerotiorum within two diagonals of the same plots of six lettuce fields
Fig. 2. Graph of variance peaks for random, uniform and clumped patterns of sclerotia of Sclerotinia sclerotiorum from two diagonals of the same plot for representative plots from field A (A), field B (B) and field C (C) lettuce fields.
The test for goodness of fit of observed sclerotial distributions to theoretical distribution models indicated that the sclerotial distributions were not adequately described by any of three distribution models tested; Poisson, negative binomial and Neyman Type A distributions (). However, analysis by plot within each field indicated that the sclerotial distributions for all plots in fields A, B and C, except plot 7 in field B, fit to at least one of three theoretical distribution models (Table S2). Two plots from field A best fit to the Poisson distribution model. The other plots in fields A, B and C best fit to either negative binomial, Neyman type A or both distribution models. In fields D and F, only three and five plots, respectively, had sufficient sclerotia frequency to run goodness of fit tests to fit theoretical distribution models. However, only two plots from field D and four plots from field F fit to at least one of the theoretical distribution model (Table S2). None of the plots in field E had enough sclerotia frequency to run a goodness of fit to any of three theoretical distribution models (Table S2).
Determination of sclerotia viability
All sclerotia recovered from soil samples of fields D, E and F, which totalled 57, 17 and 74, respectively, were tested for viability. All sclerotia (100%) regenerated Sclerotinia colonies in seven days of incubation and thus were considered viable.
Table 3. A precision of sampling size used in estimating sclerotia populations of Sclerotinia sclerotiorum in six lettuce fields
Discussion
The mean sclerotia populations of S. sclerotiorum in the six lettuce field selected for this study located near Yuma, AZ, and with a prior history of lettuce drop, ranged from 0.08 to 2.9 sclerotia/100 g of soil. The only other comparative data from lettuce fields comes from previous work by Hao & Subbarao Citation(2005), which provided estimates of the sclerotia population of S. sclerotiorum in nine lettuce fields in the San Joaquin Valley, CA. In that work, the sclerotial population averaged 0.06 sclerotia/100 cm3 of soil. Several factors could have contributed to the higher sclerotial populations in this study compared with the study by Hao & Subbarao Citation(2005) such as fungicide use or method of irrigation, both of which are known to significantly impact the incidence of lettuce drop (Subbarao, Citation1998). However, one factor that likely contributed more substantially is the intensive lettuce cropping system used in the desert production areas around Yuma, AZ, as compared with the San Joaquin Valley. In desert lettuce production systems, the higher value of the winter lettuce crop and a more limited availability of land suitable for lettuce production encourages continuous lettuce production and discourages winter crop rotation, which would likely contribute to a build-up of inoculum over time. When rotation is practiced, selection of alternate crops would likely play a role in differential soil sclerotia populations as well. For example, crop rotation with a non-host crop such as broccoli significantly reduced soil populations of sclerotia of S. sclerotiorum in the Salinas Valley, CA, where broccoli rotation is commonly used (Hao & Subbarao, Citation2003). Crop rotation with broccoli and other non-host crops is common in San Joaquin Valley lettuce production as well. However, in the Yuma area, crops such as cauliflower, cabbage and celery, which are also susceptible to S. sclerotiorum, are occasionally rotated with lettuce and this likely can contribute to the higher populations of sclerotia found in these soils.
Table 4. Goodness of fit of the distribution of sclerotia of Sclerotinia sclerotiorum in six lettuce fields to statistical distribution
Sclerotial populations in fields A, B and C were more than 10 times higher than that in fields D, E and F. Complete histories of the six lettuce fields regarding prior disease problems or cultural practices were not available, so the reason for the higher level of sclerotia population in the first three fields could not be determined. However, disease incidences for each sampled field immediately prior to harvest and the period of soil sampling were approximated by the extension specialist who helped in selecting the fields for this study (Dr. Mike Matheron). Considering what is known about lettuce drop epidemiology, whereby each infected head results in the proliferation of sclerotia that function as inoculum in subsequent crops (Subbarao, Citation1998), the higher end-of-season incidence of disease estimated in fields A, B and C compared with fields D, E and F would explain some of the differences in post-harvest sclerotia population observed between fields. This study did not determine the impact of the estimated soil sclerotia densities on disease incidence in the subsequent lettuce crops in each of the sampled fields. However, a previous study did establish the relationship between sclerotia densities and lettuce drop incidence for all three major lettuce types commonly grown in the desert production areas: crisphead, romaine and leaf lettuces (Chitrampalam et al., Citation2010). Therefore, using the estimated numbers of sclerotia in soil from the sampling strategy established in this study, and the relationship between soil sclerotia populations and subsequent incidence of lettuce drop established in the previous study, one could predict the potential disease incidence in a prospective production field.
The distribution of sclerotia of S. sclerotiorum in soil was non-random in all six lettuce fields and the high variance-to-mean ratios indicated an aggregated distribution of sclerotia, similar to that of several other soilborne sclerotia-forming fungi (Smith & Rowe, Citation1984; Mihail & Alcorn, Citation1987). Morisita’s index of dispersion, Lloyd index of patchiness and the two-term local quadrat variance method also suggested an aggregated distribution. In a few plots, a random or uniform distribution pattern was observed along the plot diagonals, but this was due to sclerotial counts of zero in most of the sampling units of those particular diagonals. The consistent aggregated pattern of sclerotia of S. sclerotiorum, where ascospores are rarely produced in desert environments, was similar to that of the closely related S. minor, which rarely produces ascospores (Dillard & Grogan, Citation1985). Indeed, the indices of dispersion obtained for S. sclerotiorum in this study (1.15 to 29.6) were higher than that obtained for other sclerotia-forming soilborne fungi such as S. minor (1.6 to 2.9) and M. phaseolina (1.52 to 6.19) (Dillard & Grogan, Citation1985; Mihail & Alcorn, Citation1987). One reason for the higher indices of dispersion for S. sclerotiorum could be that S. minor and M. phaseolina usually produce large numbers of small sclerotia, in contrast to lower numbers of large sclerotia by S. sclerotiorum, and these may be more easily dispersed from the site of origin, thereby reducing the size of aggregate. Moreover, M. phaseolina also produces asexual conidia regularly in addition to sclerotia which could also potentially influence the size of aggregate by spreading the pathogen more widely.
Lettuce fields are usually disked soon after crop harvest, and lettuce residues containing large numbers of recently formed sclerotia are incorporated and concentrated in the area immediately surrounding the previously infected lettuce head. This contributes to an aggregated pattern of recoverable sclerotia in the following season. The incidence of lettuce drop from mycelial infection shows a similar aggregated pattern (Dillard & Grogan, Citation1985; Hao & Subbarao, Citation2005). Interestingly, studies have shown that the incidence of lettuce drop due to ascospore infection has an aggregated pattern as well, suggesting that ascospore dispersal in lettuce fields is also localized (Hao & Subbarao, Citation2005).
Analysis of soil sclerotia population across fields (n = 6) for goodness of fit to theoretical distribution models showed that the observed sclerotia distribution in each field was not adequately described by either Poisson, negative binomial or Neyman type A models, even though indices of dispersal suggested an aggregated distribution. A similar failure of fitting of observed distribution to the theoretical distribution models was also observed with microsclerotia populations of Macrophomina phaseolina collected from cultivated soils in Arizona where the calculated indices distributions also indicated an aggregated distribution (Mihail & Alcorn, Citation1987). However, analysis of soil population data across plots (n = 24) revealed that sclerotia distributions in most plots were adequately described by either negative binomial or Neyman type A models, representing aggregated distribution patterns, and in agreement with the indices of dispersal. The available theoretical distribution models representing aggregated distributions are perhaps not optimal for describing sclerotia of Sclerotinia and Macrophomina in soil and new distribution models may be required to more accurately represent their distribution in cultivated field soils.
The precision of sample size used in this study to estimate sclerotia population was tested using a 20% standard error as suggested by Southwood Citation(1978). Based upon data from individual soil cores, the predicted sample size required to attain this level of precision was lower for fields A, B and C than the sample size used in this study (n = 192). However, the sample size required in determining the sclerotia population for fields D, E and F was higher than the sample size used in this study. The reason for the low number of soil samples required to determine sclerotia population in fields from A to C was due to the presence of sclerotia in a higher percentage of sampling units and all of the eight plots in all three fields had mean sclerotia populations between 0.5 to 5.8/100 g of soil. The standard error calculated with 192 soil samples for fields A, B and C (10, 18 and 19%, respectively) was lower than the allowable standard error (20%). However, the standard error calculated for fields D, E and F (25, 31 and 20%, respectively) were equal to or higher than the allowable standard error. The reason for higher requirement of sampling units and high standard error in fields D and E was due to the presence of low sclerotia population in these two fields. Since the number of soil samples required to determine sclerotia population for most fields was equal to or below the sample size used in this study, a sample size of 192 cores is a reasonable sized sampling to determine sclerotia population in fields which have a history of high incidence of lettuce drop. For fields with a lower incidence of lettuce drop, this sampling size may not be sufficient for precision. However, if the disease potential is not as high, a less precise estimation of soil population may be acceptable.
The calculated potential sampling error based upon plot means was higher than the allowable sampling error (20%) in five out of six fields sampled (22 to 40%), although the sampling error in fields A, B and F (27, 22 and 22%, respectively) was very close to the allowable sampling error. To achieve a sampling error of 20% would require 15, 10 and 9 plots for fields A, B and F, respectively. Note that almost doubling the number of plots for field A (8 plots to 15 plots) only reduced the standard error by 7. Thus, the sample size of 8 plots used in this study to estimate sclerotia population appears to be reasonable for a field with high incidence of lettuce drop. Again, if the disease potential is not high, a less precise estimation of soil population may be acceptable.
In summary, the sclerotia populations of S. sclerotiorum in winter lettuce production fields in the deserts of Arizona were higher than that reported in other areas and followed an aggregated distribution. The number of sampling units used in this study was sufficient to determine the observed sclerotia population within a reasonable sampling error and thus these data can be utilized as foundations for subsequent studies on the ecology and management S. sclerotiorum in desert agroecosystems. Although this study estimated the sclerotia population of S. sclerotiorum and determined their distribution in lettuce fields, it did not assess the potential impact of this inoculum on subsequent lettuce production. Thus, a future study on the relationship between sclerotium density and disease in desert winter production will be critical for the future development of more effective management strategies.
Supplemental data
Supplemental data for this article can be accessed here: http://dx.doi.org/10.1080/07060661.2013.841758
Supplemental Tables
Download MS Word (31.3 KB)Acknowledgements
This work was supported in part by the University of Arizona College of Agriculture and Life Sciences, the Arizona Iceberg Lettuce Research Council and the Arizona Department of Agriculture. We would like to thank Monica Alvarado and Chad Cox for assistance in collecting and processing the field soil samples.
References
- Chitrampalam, P., Figuli, P.J., Matheron, M.E., Subbarao, K.V., & Pryor, B.M. (2008). Biocontrol of lettuce drop caused by Sclerotinia sclerotiorum and S. minor in desert agroecosystems. Plant Dis., 92, 1625–1634.
- Chitrampalam, P., Turini, T.A., Matheron, M.E., & Pryor, B.M. (2010). Effect of sclerotia density and irrigation on disease incidence and on efficacy of Coniothyrium minitans in suppressing lettuce drop caused by Sclerotinia sclerotiorum. Plant Dis., 94, 1118–1124.
- Dillard, H.R., & Grogan, R.G. (1985). Relationship between sclerotial spatial pattern and density of Sclerotinia minor and the incidence of lettuce drop. Phytopathology, 75, 90–94.
- Elliot, J.M. (1971). Some methods for the statistical analysis of samples of benthic invertebrates. Freshwater Biological Association Science Publication No. 25. Ambleside, U.K., Freshwater Biological Association. 144 p.
- Grogan, R.G., Sall, M.A., & Punja, Z.K. (1980). Concepts for modeling root infection by soilborne fungi. Phytopathology, 70, 361–363.
- Hao, J.J., & Subbarao, K.V. (2003). Effects of broccoli rotation on lettuce drop caused by Sclerotinia minor and on the population density of sclerotia in soil. Plant Dis., 87, 159–166.
- Hao, J.J., & Subbarao, K.V. (2005). Comparative analyses of lettuce drop epidemics caused by Sclerotinia minor and S. sclerotiorum. Plant Dis., 89, 717–725.
- Imolehin, E.D., & Grogan, R.G. (1980). Effects of oxygen, carbon dioxide and ethylene on growth, sclerotial production, germination, and infection by Sclerotinia minor. Phytopathology, 70, 1158–1161.
- Lin, C.S., Pousjinsky, G., & Mauer, M. (1979). An examination of five sampling methods under random and clustered disease distributions using simulation. Can. J. Plant Sci., 59, 121–130.
- Lloyd, M. (1967). Mean crowding. J. Anim. Ecol., 36, 1–30.
- Lopez-Collado, J., & Osada-Velazquez, H.K. (1997 ). PADIS v. 1.01. Programma de Ajuste a Distributiones dIScreatas.
- Ludwig, A.J., & Reynolds, F.J.(1988). Statistical Ecology. Hoboken, NJ: Wiley-Interscience. 337 pages.
- Matheron, M.E., & Porchas, M. (2004). Activity of boscalid, fenhexamid, fluazinam, fludioxonil, and vinclozolin on growth of Sclerotinia minor and S. sclerotiorum and development of lettuce drop. Plant Dis., 88, 665–668.
- Matheron, M.E., & Porchas, M. (2005). Influence of soil temperature and moisture on eruptive germination and viability of sclerotia of Sclerotinia minor and S. sclerotiorum. Plant Dis., 89, 51–54.
- Mihail, J.D., & Alcorn, A.M. (1987). Macrophomina phaseolina: Spatial patterns in a cultivated soil and sampling strategies. Phytopathology, 77, 26–1131.
- Nicot, P.C., Rouse, D.I., & Yandell, B.S. (1984). Comparison of statistical methods for studying spatial pattern of soilborne plant pathogen in the field. Phytopathology, 74, 1399–1402.
- Purdy, L.H. (1979). Sclerotinia sclerotiorum: History, diseases and symptomology, host range, geographic distribution and impact. Phytopathology, 69, 875–880.
- Shukla, R.P., Tandon, P.L., & Suman, C.L. (1985). Spatial distribution of different stages of mango stone weevil, Sternochetus mangiferae (Fabricius) (Coleoptera:Curculionidae). Agric. Ecosyst. Environ., 12, 135–140.
- Smith, V.L., & Rowe, R.C. (1984). Characteristics and distribution of propagules of Verticillium dahliae in Ohio potato field soils and assessment of two assay methods. Phytopathology, 74, 553–556.
- Southwood, T.R.E. (1978). Ecological Methods, 2nd ed. New York: Chapman and Hall Press. 524 pages.
- Subbarao, K.V. (1998). Progress toward integrated management of lettuce drop. Plant Dis., 82, 1068–1078.
- Subbarao, K.V., Dacuyan, S., Koike, S.T., & Jackson, L.E. (1994). Evaluation of three quantitative assays for Sclerotinia minor. Phytopathology, 84, 1471–1475.