
Abstract
Root-lesion nematodes (Pratylenchus spp.) are important pests of potato (Solanum tuberosum L.), particularly in sandy soils and in the presence of Verticillium dahliae. We assessed the effects of crop rotation and soil management practices (reduced tillage, cover crops and composted manure applications) on population densities of P. neglectus on potato in a sandy loam soil in southern Alberta. Crops in rotation included potato (P), dry bean (DB, Phaseolus vulgaris L.), wheat (W, Triticum aestivum L.), sugar beet (SB, Beta vulgaris L.), oats (O, Avena sativa L.) and timothy (T, Phleum pratense L.). The rotations included ‘conventional’ and ‘conservation’ versions of 3-year (DB-W-P) and 4-year (W-SB-DB-P) rotations, as well as 5-year (W-SB-W-DB-P) and 6-year (O/T-T-T-SB-DB-P) rotations. Conservation practices included autumn cover crops and incorporation of compost as a substitute for inorganic fertilizer. Population densities of P. neglectus in the potato phase of each rotation were assessed in autumn of 2006 and 2007, and at the beginning, middle and end of 2008, 2009 and 2010 growing seasons. Pratylenchus neglectus populations were affected by rotation length but not soil management practices; population densities in the 3-year rotations were larger than in most other rotations at most sample dates. Potato yields in the 3-year conventional rotation were consistently lower than in longer rotations. In the 3-year rotation, potato was preceded by wheat, which is known to be a good host for P. neglectus. We speculate that greater population build-up on wheat in the year before potato, rather than rotation length or crop diversity per se, was the cause of the larger population densities in the 3-year rotations than in longer rotations.
Résumé
Les nématodes des racines (Pratylenchus spp.) sont d’importants ravageurs des pommes de terre (Solanum tuberosum L.), particulièrement de celles qui poussent dans des sols sablonneux, en présence de Verticillium dahliae. Nous avons évalué les effets des rotations des cultures et des pratiques de gestion des sols (travail réduit du sol, plantes de couverture et applications de fumiers compostés) sur les densités de population de P. neglectus sur la pomme de terre poussant dans les loams sableux du sud de l’Alberta. Les rotations des cultures incluaient les pommes de terre (PT), le haricot (H, Phaseolus vulgaris L.), le blé (B, Triticum aestivum L.), la betterave à sucre (BS, Beta vulgaris L.), l’avoine (A, Avena sativa L.) et la phléole des prés (PP, Phleum pratense L.). Les rotations incluaient les versions « traditionnelle » et « axée sur la conservation » de trois ans (H-B-PT) et de quatre ans (B-BS-H-PT), de même que les rotations de cinq ans (B-BS-B-H-PT) et de six ans (A/PP-PP-PP-BS-H-PT). Les pratiques axées sur la conservation incluaient les plantes de couverture automnales et l’incorporation de compost comme engrais biologique. Les densités des populations de P. neglectus durant la phase « pomme de terre » de chaque rotation ont été évaluées à l’automne 2006 et à l’automne 2007, ainsi qu’au début, au milieu et à la fin des saisons de croissance de 2008, 2009 et 2010. Les populations de P. neglectus ont été perturbées par la durée des rotations, mais pas par les pratiques de gestion des sols. Les densités des populations au cours des rotations de trois ans étaient plus élevées que durant les autres types de rotations, et ce, à la plupart des dates d’échantillonnage. Les rendements de pommes de terre obtenus de la rotation traditionnelle de trois ans étaient invariablement plus bas que ceux des autres types de rotations. Dans la rotation de trois ans, les pommes de terre étaient précédées par le blé qui est reconnu comme étant un hôte de prédilection de P. neglectus. Nous avançons l’hypothèse voulant que la plus grande augmentation de populations chez le blé au cours de l’année précédant la pomme de terre, plus que la durée de la rotation ou que la variété de culture en soi, a engendré les densités plus élevées de populations au cours des rotations de trois ans plutôt qu’au cours des rotations plus longues.
Introduction
Root-lesion nematodes (Pratylenchus spp.) can be serious pests of potato. Pratylenchus penetrans (Cobb) Schuurmans-Stekhoven has been the focus of most research on the role of root-lesion nematodes in potato production, as it is known to interact with Verticillium dahliae Kleb. to cause the Potato Early Dying (PED) disease complex (e.g. Rowe & Powelson Citation2002). Pratylenchus neglectus is often associated with potato production in western North America (e.g. Davis et al. Citation1992; Hafez et al. Citation1992; Umesh & Ferris Citation1994; Mahran et al. Citation2010). In greenhouse and microplot studies, this species has been shown to reproduce on potato and reduce its growth (Olthof Citation1990; Umesh & Ferris Citation1994; Hafez et al. Citation1999), although there appears to be some geographical variation in the association, with populations in Manitoba and Idaho showing apparently limited capacity to reproduce on potato (Hafez et al. Citation1999; Mahran et al. Citation2010). The role of P. neglectus in PED has not been studied extensively, although one study suggests that there may be variation among geographically isolated populations of P. neglectus with respect to their ability to contribute to PED (Hafez et al. Citation1999).
Potato production on irrigated, sandy soils in southern Alberta has grown rapidly, with acreage planted to potatoes expanding from 5182 ha in 1998 to 16 582 ha in 2003 (Alberta Agriculture Rural Development Citation2013). This increase in potato production has occurred in tandem with similar increases in production of dry beans and sugar beets, and producers are interested in identifying rotations and production practices for these crops that will conserve soil resources. Cereal crops are commonly used in rotations with potato, dry beans or sugar beets in western Canada. Pratylenchus neglectus has a wide host range (Castillo & Vovlas Citation2007), but it is most often associated with cereal and grain legume crops (Riley & Kelly Citation2002; Castillo & Vovlas Citation2007; Smiley & Machado Citation2009). Population densities of P. neglectus have been positively associated with frequency of cereal cropping (Riley & Kelly Citation2002) and damage to those crops (Smiley & Machado Citation2009).
In order to assess the economic and environmental aspects of a range of crop rotations and production practices, a multifaceted long-term rotation experiment was established in southern Alberta in 2000 (Li et al. Citation2015). As a component of this broader study, the objective of the research presented here was to compare the effects of preceding crop rotations and conservation verses conventional production practices on population densities of P. neglectus on potato.
Materials and methods
Field experiment
The crop rotation experiment was established in spring of 2000 at the Agriculture and Agri-Food Canada Vauxhall Substation (50°03′N, 112°09′W, elev. 781 m) on an Orthic Brown Cherozemic soil (Chin series). Texture in the 0–15 cm depth was sandy loam (0.52 kg kg−1 sand, 0.34 kg kg−1 silt, 0.14 kg kg−1 clay). The 30 year normal (1981–2010) mean annual precipitation was 352 mm with a mean annual air temperature of 5.8°C. The principal crops were ‘Russet Burbank’ potato (P), dry bean (DB), wheat (W), sugarbeet (SB), oats (O) and timothy (T) grown in four basic rotation sequences with lengths of 3–6 years, with ‘conventional’ and ‘conservation’ versions of the 3- and 4-year rotations, for a total of six rotations that included potato (). Each crop of each rotation sequence was represented in any given year in the experiment, which enabled us to sample the potato phase of each rotation in every year. Each rotation was replicated four times in a randomized complete block design. Plot dimensions were 10 × 18.3 m with a 2.1 m buffer zone between each plot.
Table 1. Characteristics of crop rotations and soil-nutrient management practices preceding potato, and average marketable and total potatoes for the period 2001–2009.
Conservation rotations were characterized by the following practices: (1) use of autumn-seeded cover crops of rye (Secale cereale L.); (2) incorporation of composted cattle manure as a substitute for inorganic P fertilizer prior to potato, (3) direct seeding or reduced tillage where possible, and (4) narrow-row direct harvested dry beans (Li et al. Citation2015). The reduced tillage included the use of a single pass chisel plough (3-year conservation) or dammer-diker (4-, 5- and 6-year conservation rotations) the autumn before potato in conservation rotations whereas the conventional rotations were moldboard ploughed to a depth of 25 cm (). The compost was derived from straw-bedded feedlot manure and had been turned multiple times with a tractor-pulled windrow turner over a 3 month active thermophilic phase and then allowed to cure for a further 3 months before use (Larney & Olson Citation2006). The compost was applied at 42 Mg ha−1 wet weight in the autumn prior to potatoes in the 4- and 5-year conservation rotations, and to the 6-year rotation. Compost was applied to the 3-year conservation rotation at a lower rate of 28 Mg ha−1.
Nematode sampling
Nematode sampling commenced in the autumn of 2006 after a complete cycle of the 6-year rotation and two full cycles of the 3-year rotation. Only the potato phases of the six rotations were sampled each year. Plots were sampled immediately after potato harvest in 2006 and 2007. In each of 2008, 2009 and 2010, samples were taken in the early season (just before or after potato planting, May–early June), mid-season (late July–early August), and immediately after potato harvest (late September). At each sample date, twenty 2-cm diameter cores were taken to a 25 cm depth from each plot and composited. The samples were kept refrigerated and shipped to the Pacific Agri-Food Research Centre in Agassiz, BC, where nematode populations were analysed. Nematodes were extracted from 50 mL soil subsamples using Baermann pans (Forge et al. Citation2007) and Pratylenchus nematodes were enumerated using an inverted microscope. In 2008 and 2009, potato root fragments were picked from the mid-season samples and endoparasitic nematodes were extracted over 7 days in a mist chamber (Ingham Citation1994).
The population of Pratylenchus was identified as P. neglectus on the basis of morphological characteristics, particularly the presence of two lip annules, lack of spermatheca and absence of males in the population (Castillo & Vovlas Citation2007; Yu Citation2008). The morphology-based identification was confirmed by molecular analyses. A portion of the 28S region of rDNA was amplified using nematode-specific forward and reverse primers D2A (CAAGTACCGTGAGGGAAAGTTG) and D3B (TCGGAAGGAACCAGCTACTA), respectively (De Ley et al. Citation1999). Amplicons were sequenced and compared with sequences for P. neglectus in GenBank. The sequence data were also filed with GenBank (accession numbers KM200578, KM200579).
Data analyses
Nematode counts were expressed as nematodes per 100 mL soil and transformed to log(x + 100) before final analyses to correct heteroscedasticity. For data from the 3-year and 4-year rotations only, the main-factor and interaction effects of production practices (conventional vs. conservation), rotation length and sample date were analysed using Proc Mixed in SAS (SAS, Inc., Cary, NC). Sample dates were treated as repeated measures and blocks were designated as random variables. Subsequently, data from all rotations were separated by season of sampling (spring, summer, autumn) and for each season, the main-factor and interaction effects of rotation length and year were analysed. The analysis for autumn included 2006 and 2007 data whereas analyses for spring and summer were based on 2008–2010 data only. The SLICE procedure was used to test the effect of rotation length and compare means at each sample date.
Results and discussion
Rotation system effects on P. neglectus population densities
There were no significant effects of production system (conventional vs. conservation) on P. neglectus population densities, either alone or as an interaction with length of rotation (3-year vs. 4-year) (data not shown). The conservation rotations differed from the conventional rotations in that a rye cover crop was planted each autumn, there was less tillage, and compost was applied each autumn preceding potato. The conservation and conventional rotations thus differed in several ways that could have had competing influences on P. neglectus populations and therefore resulted in no net effect. For example, the compost amendments and general enhancement of soil organic matter in the conservation rotations (Li et al. Citation2015) could have fostered a more suppressive soil food web leading to suppression of P. neglectus as has been observed in other systems (e.g. Abawi & Widmer Citation2000; Oka & Yermiyahu Citation2002; Forge & Kempler Citation2009; Oka Citation2010). In contrast, the autumn rye cover crop would likely have promoted P. neglectus population build-up by extending the season with a host species, thereby negating any suppression due to organic matter and soil food web enhancement.
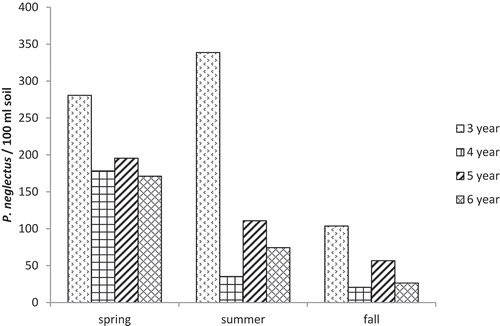
Because there was no significant effect of production system or system × rotation length interaction, subsequent analyses were conducted on 3-year and 4-year data pooled over the two production systems. The 3-year rotation (B-W-P) resulted in the greatest population densities on potato. There was a significant date × rotation effect, with significant (P < 0.05) effects of rotation length on six of the 10 sample dates (). At all of these dates, population densities in the 3-year rotation were significantly greater than in at least one other rotation, and there were few significant differences among the 4-, 5- and 6-year rotations. There was also a significant rotation effect for each season of sampling (). Population densities in the 3-year rotation were significantly greater than in 4-year and 6-year rotations in all three seasons, and greater than in the 5-year rotation in spring and autumn samples.
Table 2. Effect of rotation length on population densities of Pratylenchus neglectus in soil (nematodes per 100 mL soil) planted to potato, by sample date. Analyses were conducted on log(X + 100) transformed data; data presented are means of raw data.
Fig. 1 Effect of rotation length on population densities of P. neglectus in soil planted to potato (nematodes per 100 mL soil), by season of sampling. Analysis of variance and mean separations were conducted on log(X + 100) transformed data.
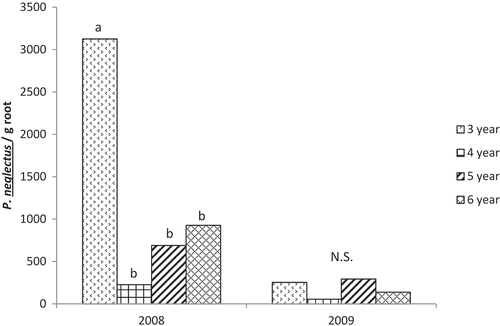
The effect of the 3-year rotation on P. neglectus population densities was manifest as significantly greater numbers of nematodes per g potato root in the 3-year rotation than in other rotations in 2008 but not in 2009 (). There have been conflicting reports on the host status of potato for P. neglectus (Mahran et al. Citation2010). Our root sample data included population densities in excess of 3000 nematodes per g root, which strongly suggests that potato was a good host for the population of P. neglectus at the test site. This is consistent with most previous studies of the potato–P. neglectus interaction (Olthof Citation1990; Umesh & Ferris Citation1994; Hafez et al. Citation1999), although a more recent study in Manitoba indicated there was no population increase on potato and very low population densities in roots from field plots (<50 nematodes g−1) (Mahran et al. Citation2010), leading the authors to speculate that ‘Russet Burbank’ potato may be a poor host for P. neglectus.
Fig. 2 Effect of rotation length on population densities of P. neglectus in roots of potato (nematodes g−1 root). Analysis of variance and mean separations were conducted on log(X + 100) transformed data. N.S. = non-significant effect of rotation in analysis of variance.
In addition to being longer rotations with a greater diversity of crops, the 4-, 5- and 6-year rotations differed from the 3-year rotation in that dry beans rather than wheat immediately preceded potato. As P. neglectus is well known to parasitize and multiply rapidly on wheat, we speculate that the sequence of wheat-potato was the primary cause of the increased population densities measured in potatoes in the 3-year rotation. All of the crops used in this experiment can host P. neglectus (Castillo & Vovlas Citation2007), but there are no data directly comparing reproductive potential of the nematode on wheat to any of the other crops. The 5-year rotation fostered significantly greater population densities than the 6-year rotation in autumn 2006 and the 4-year rotation in spring 2008 and spring 2009 (). It is worth noting that the 5-year rotation had wheat-bean in the 2 years preceding potato whereas the 4- and 6-year rotations both had sugar beet-bean in the 2 years preceding potato. This specific effect of wheat is significant, as it indicates that it may not be necessary to rotate away from potato for a significant length of time as long as wheat does not immediately precede the potato crop.
Alternatively, the lower P. neglectus population densities could be the result of the greater diversity of crop species, perhaps fostering beneficial changes in soil microbial ecology resulting in nematode suppression. Further experimentation with long rotations with wheat at differing positions in the rotations, relative to potato, would help elucidate the cause of high population densities in the 3-year rotation observed in this study.
Relationships with potato yields
The damage potential of P. neglectus on potato is not well understood as the majority of research on interactions between root-lesion nematodes and potato has focused on P. penetrans. Population densities of P. neglectus were generally inversely correlated with yields across the rotations, with the 3-year rotations having lower cumulative yields than other rotations, and with the highest yields on the 5-year conservation rotation which was significantly greater than all rotations except the 4-year and 6-year conservation rotations (). Attempts to correlate P. neglectus population densities with yields across individual plots did not yield significant correlation coefficients as a result of high plot-to-plot variability. Symptoms of Potato Early Dying (PED) were evident, particularly in the 3-year conventional plots in 2007. PCR with primers specific for the ITS1 and ITS2 regions of Verticillium dahliae rDNA confirmed the presence of V. dahliae in the plots (L. Kawchuk, personal communication). We speculate that because root-lesion nematodes can act synergistically with V. dahliae to cause PED, the lower P. neglectus pressure in the longer rotations contributed to reduced intensity of PED relative to the 3-year rotation. Additional research using controlled inoculation of microplots under field conditions to further elucidate relationships between P. neglectus population densities, V. dahliae and potato yields under Alberta growing conditions is warranted.
In conclusion, our data demonstrate that short rotations with wheat can significantly increase population densities of P. neglectus in fields subsequently planted to potato. It is not possible to draw firm conclusions about whether this effect is the direct result of having wheat as the crop immediately preceding potato or some other aspect of the shorter and less diverse rotation, such as shifts in nematode-antagonistic soil microflora. The well-recognized affinity of P. neglectus with cereal crops leads us to speculate that greater population build-up on wheat the year before potato was the primary cause of the greater population densities on potato in the 3-year rotations than in the longer rotations. We also speculate that the greater population densities in the 3-year rotations contributed to the greater incidence of PED and lower potato yields than in the longer rotations.
Acknowledgements
We thank Jim Sukeroff, Dale Reynolds and Vauxhall Substation staff and summer students for field assistance.
Additional information
Funding
References
- Abawi GS, Widmer TL. 2000. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl Soil Ecol. 15:37–47.
- Alberta Agriculture Rural Development [Internet]. 2013. Alberta Irrigation Information 2012; [ cited 2014 Jun 27]. Available from: http://www1.agric.gov.ab.ca/$Department/deptdocs.nsf/all/irr7401
- Castillo P, Vovlas N. 2007. Pratylenchus (Nematoda: Pratylenchidae): Diagnosis, Biology, Pathogenicity and Management: Nematology Monographs and Perspectives. Leiden, The Netherlands : Brill; p. 6.
- Davis JR, Hafez SL, Sorensen LH. 1992. Lesion nematode suppression with the Butte potato and relationships to Verticillium wilt. Am Potato J. 69:371–383.
- De Ley P, Felix MA, Frisse LM, Nadler SA, Sternberg PW, Thomas WK. 1999. Molecular and morphological characterization of two reproductively isolated species with mirror image anatomy (Nematoda: Cephalobidae). Nematology. 2:591–612.
- Forge TA, Kempler C. 2009. Organic mulches influence population densities of root-lesion nematodes, soil health indicators, and root growth of red raspberry. Can J Plant Pathol. 31:241–249.
- Forge TA, Kimpinski J. 2007. Nematodes. In: Gregorich EG, Carter MR, editors. Sampling and methods of analysis. 2nd ed. Boca Raton (FL): CRC Press; p. 415–425.
- Hafez SL, Al-Rehiayani S, Thornton M, Sundararaj P. 1999. Differentiation of two geographically isolated populations of Pratylenchus neglectus based on their parasitism of potato and interaction with Verticillium dahliae. Nematropica. 29:25–35.
- Hafez SL, Golden AM, Rashid F, Handoo Z. 1992. Plant-parasitic nematodes associated with crops in Idaho and eastern Oregon. Nematropica. 22:193–202.
- Ingham RE. 1994. Nematodes. In: Weaver RW, editor. Methods of soil analysis. Part 2 – Microbiological and biochemical properties. Madison (WI): Soil Science Society of America; p. 459–490.
- Larney FJ, Olson AF. 2006. Windrow temperatures and chemical properties during active and passive aeration composting of beef cattle feedlot manure. Can J Soil Sci. 86:783–797.
- Li L, Larney FJ, Angers DA, Pearson DC, Blackshaw RE. 2015. Surface soil quality attributes following 12 years of conventional and conservation management on irrigated rotations in southern Alberta. Soil Sci Soc Amer J. 79:930–942.
- Mahran A, Tenuta M, Shinners-Carenelly T, Mundo-Ocampo M, Daayf F. 2010. Prevalence and species identification of Pratylenchus spp. in Manitoba potato fields and host suitability of ‘Russet Burbank’. Can J Plant Pathol. 32:272–282.
- Oka Y. 2010. Mechanisms of nematode suppression by organic soil amendments – A review. Appl Soil Ecol. 44:101–115.
- Oka Y, Yermiyahu U. 2002. Suppressive effects of composts against the root-knot nematode Meloidogyne javanica on tomato. Nematology. 4:891–898.
- Olthof Th HA. 1990. Reproduction and parasitism of Pratylenchus neglectus on potato. J Nematol. 22:303–308.
- Riley IT, Kelly SJ. 2002. Endoparasitic nematodes in cropping soils of western Australia. Aust J Exp Agr. 42:49–56.
- Rowe RC, Powelson ML. 2002. Potato early dying: Management challenges in a changing production environment. Plant Dis. 86:1184–1193.
- Smiley RW, Machado S. 2009. Pratylenchus neglectus reduces yield of winter wheat in dryland cropping systems. Plant Dis. 93:263–271.
- Umesh KC, Ferris H. 1994. Influence of temperature and host plant on the interaction between Pratylenchus neglectus and Meloidogyne chitwoodi. J Nematol. 26:65–71.
- Yu Q. 2008. Species of Pratylenchus (Nematoda: Pratylenchidae) in Canada: Description, distribution, and identification. Can J Plant Pathol. 30:477–485.