
Abstract
Puccinia triticina Eriks., the causal agent of wheat leaf rust, was collected across Canada in 2010 to determine the virulence spectrum present in the pathogen population. From 314 infected leaf collections, 399 single pustule isolates were recovered, including 341 from Manitoba and Saskatchewan, 38 from Ontario, nine from Quebec and 11 from Prince Edward Island (PEI). When analysed for virulence on 16 standard differential lines, 41 different virulence phenotypes were identified. The most common were MLDS (26.1%), TDBJ (22.8%) and TDBG (11.8%). In Manitoba and Saskatchewan, the most common phenotypes were MLDS (29.6%), TDBJ (25.5%) and TDBG (13.8%), which were also abundant in 2009. In Ontario, the most common were MBTN (23.7%), TDBJ (10.5%), MCGJ, MCTN and MLDS (each at 7.9%), whereas in Quebec, each of the nine isolates analysed had a different phenotype. In PEI, the more commonly isolated phenotypes were MBNQ (six isolates), MBTN and MHNQ (two isolates each). Each population had unique virulence phenotypes, and the eastern populations were more diverse than the Manitoba and Saskatchewan population. Virulence trends from previous years continued in 2010, with increasing levels of virulence found for Lr9 and Lr17 while decreasing virulence was found for Lr2a, Lr14a and Lr24. There was no virulence found in 2010 for Lr19, Lr21, Lr22a, Lr29, Lr32 or Lr52.
Résumé
Puccinia triticina Eriks, l’agent causal de la rouille brune, a été collecté partout au Canada en 2010 afin de déterminer le spectre de virulence dans la population de l’agent pathogène. À partir de 314 collections de feuilles infectées, 399 isolats à pustules individuelles ont été récupérés, y compris 341 en provenance du Manitoba et de la Saskatchewan, 38 de l’Ontario, 9 du Québec et 11 de l’Île-du-Prince-Édouard (Î.-P.-É.). Lorsque testés sur 16 lignées différentielles standards pour la virulence, 41 phénotypes de virulence différents ont été identifiés. Les plus communs étaient MLDS (26.1%), TDBJ (22.8%) et TDBG (11.8%). Au Manitoba et en Saskatchewan, les phénotypes les plus communs étaient MLDS (29.6%), TDBJ (25.5%) et TDBG (13.8%), qui abondaient en 2009 également. En Ontario, les plus courants étaient MBTN (23.7%), TDBJ (10.5%) MCGJ, MCTN et MLDS (7.9% chacun), tandis qu’au Québec, chacun des neuf isolats analysés avait un phénotype différent. À l’Î.-P.-É., les phénotypes les plus couramment isolés étaient MBNQ (six isolats), MBTN et MHNQ (deux isolats chacun). Chaque population possédait des phénotypes de virulence uniques et les populations de l’est étaient plus diversifiées que celle du Manitoba et de la Saskatchewan. Les tendances des années précédentes quant à la virulence se sont maintenues en 2010, sauf en ce qui a trait à la fréquence de virulence à l’égard des gènes de virulence Lr9 et Lr17, qui a augmenté, et la fréquence de virulence à l’égard des gènes Lr2a, Lr14a et Lr24, qui a décru. Aucune virulence n’a été détectée en 2010 à l’égard des gènes Lr19, Lr21, Lr22a, Lr29, Lr32 ou Lr52.
Introduction
Wheat leaf rust, caused by Puccinia triticina Eriks. (Anikster et al. Citation1997) (= P. recondita Rob. ex Desmaz. f. sp. tritici), is one of the most common and destructive diseases on wheat in Canada and other parts of the world. The disease reduces yield by 5–25%, depending on the susceptibility of the crop, severity of the epidemic, fungicide application and the timing of infection (McCallum et al. Citation2007). Canadian virulence surveys for P. triticina have been conducted annually since 1931 to monitor changes in the pathogen population, determine the frequency of virulence to key resistance genes, provide a historic record of shifts in virulence and detect groups of genetically similar isolates (Wang et al. Citation2010; McCallum et al. Citation2016). This knowledge can be used along with information on the resistance genes in current and future wheat cultivars to make informed decisions when breeding wheat for leaf rust resistance.
The main objective of this study was to determine the frequency and distribution of virulence phenotypes from P. triticina isolates collected across Canada in 2010, using 16 near-isogenic wheat lines inoculated at the seedling stage, to monitor changes in the pathogen population. A secondary objective was to determine the virulence spectrum for representative isolates of all the virulence phenotypes found, on five additional near-isogenic wheat lines with adult plant resistance genes and 13 additional near-isogenic lines with seedling resistance genes.
Materials and methods
Infected wheat leaves were collected from individual fields and nurseries from June to September in 2010 in various locations throughout Prince Edward Island (PEI), Ontario, Quebec, Manitoba, Saskatchewan and Alberta. The leaves were air dried at 20–27oC for ~12–24 h then stored separately for 2–4 months at 5oC. Puccinia triticina urediniospores from individual collections were then scraped off from infected leaves using a metal spatula and a small amount of water, and inoculated onto the susceptible wheat cultivar ‘Little Club’ by rubbing the leaves with a mixture of urediniospores, water and Tween 20 as described previously (McCallum & Seto-Goh Citation2005). Each pot of ‘Little Club’ seedlings was pretreated with 50 mL maleic hydrazide solution (0.36 g L−1 concentration) ~5 d after seeding to prevent the emergence of secondary leaves and to produce larger uredinia with abundant sporulation. A plastic cone ~25 cm in height with an open top was placed over each pot to minimize cross contamination. Inoculated plants were placed into a dew chamber (Model I-60D, Percival Scientific, Perry, IA) with nearly 100% relative humidity for ~17 h to allow the urediniospores to germinate and initiate the infection process, then placed in a greenhouse at 20 ± 4oC with supplemental high-pressure sodium lighting. Approximately 7 d after inoculation chlorotic spots appeared, indicating areas of infection. The leaves were then trimmed so that a single isolated uredinium remained on the upper edge of each trimmed leaf. Cross contamination was minimized by removing all extra leaves. At ~14 d after inoculation, urediniospores were collected from a single isolated uredinium into a 00 gelatin capsule using a vacuum suction micro-collector, mixed with a light mineral oil (Bayol, Esso Canada, Toronto, ON), and sprayed onto a 7-d-old set of wheat seedlings which included ‘Thatcher’ and 16 single resistance gene ‘Thatcher’ near-isogenic lines. Two single uredinial isolates were typically evaluated from each rust collection, although sometimes one or three isolates per collection were analysed. Approximately 12 seeds of each ‘Thatcher’ near-isogenic line were planted in a clump, and the clumps were evenly spaced in a fibre flat (25 × 15 cm). Plants were pretreated with maleic hydrazide as described above, which helped to maintain green leaves without changing the resistance of the wheat plants or virulence of the P. triticina isolates. After inoculation, the plants were dried for at least 1 h to allow the oil to evaporate and then incubated and maintained as described above. Infection types produced on the 12 standard leaf rust (Lr) differential lines [Set 1: Lr1 (RL6003a), Lr2a (RL6016), Lr2c (RL6047), Lr3 (RL6002); Set 2: Lr9 (RL6010), Lr16 (RL6005), Lr24 (RL6064), Lr26 (RL6078); Set 3: Lr3ka (RL6007), Lr11 (RL6053), Lr17 (RL6008), Lr30 (RL6049)] were used to determine the three letter code according to the virulence phenotype nomenclature (Long & Kolmer Citation1989). RL6003a is a reselection of RL6003 that has only the resistance gene Lr1. Four supplemental differential lines [Set 4: LrB (RL6051), Lr10 (RL6004), Lr14a (RL6013), Lr18 (RL6009)] were added to provide additional virulence information about the isolates, resulting in a four letter code. All isolates were also inoculated onto ‘Thatcher’ and Lr1 +LrCen (RL6003b, a reselection from RL6003 containing both Lr1 and LrCen). The resistance gene temporarily named LrCen was previously identified in the Thatcher-Lr1 near-isogenic line RL6003 (McCallum & Seto-Goh Citation2006b). Infection types to all the differential near-isogenic lines were rated 12 d after inoculation. Isolates that produced infection types ‘;’ (hypersensitive flecks), ‘1’ (small uredinia with necrosis) and ‘2’ (small to medium sized uredinia with chlorosis) were considered avirulent to the differential line, and those that produced infection types ‘3’ (medium sized uredinia without chlorosis or necrosis) and ‘4’ (large uredinia without chlorosis or necrosis) were considered virulent to the line (Long & Kolmer Citation1989). Inoculations were repeated if the infection response was not clear.
At least one isolate from most of the unique virulence phenotypes identified was inoculated onto adult plants of ‘Thatcher’ and five ‘Thatcher’ near-isogenic lines (Lr12 [RL6011], Lr13 [RL4031], Lr22a [RL6044], Lr35 [RL6082], or Lr37 [RL6081]). Single plants of each ‘Thatcher’ near-isogenic line and ‘Thatcher’ were grown together in a 15-cm-diameter pot in a greenhouse at day/night temperatures of 25/18°C with supplemental high-pressure sodium lighting. Plants were trimmed so that only two or three culms per plant remained. Flag leaves from each of the plants within a pot were inoculated with a single pustule isolate, as described previously for the seedling inoculation. Inoculated plants were dried for over 1 hour to prevent cross contamination then incubated overnight in a dew chamber and grown in the greenhouse as described previously. Infection types were evaluated 14 d after inoculation. This subset of isolates was also tested on 13 additional ‘Thatcher’ near-isogenic lines at the seedling stage (Lr2b [RL6019], Lr3bg [RL6042], Lr14b [RL6006], Lr15 [RL6052], Lr19 [RL6040], Lr20 [RL6092], Lr21 [RL6043], Lr23 [RL6012], Lr25 [RL6084], Lr28 [RL6079], Lr29 [RL6080], Lr32 [RL6086] and Lr52). These isolates were also retested on seedling plants of the set of 16 ‘Thatcher’ near-isogenic lines mentioned previously to confirm their reactions, since many of these resistance genes (particularly Lr18 and LrB) are sensitive to temperature and other conditions. Inoculation, incubation and rating were as described previously for seedling evaluation.
Results and discussion
During 2010, wheat leaf rust was first observed on 9 June on spring wheat at Winkler, Manitoba. This is relatively early in the growing season, which led to a widespread and relatively severe epidemic in Manitoba on susceptible wheat cultivars that were not protected with foliar fungicide application (McCallum & Seto-Goh Citation2011); however, most fields were treated with a foliar fungicide application. In Ontario, leaf rust was found in 26 of 28 wheat fields surveyed during the 2010 growing season, and the mean severity was 2.4 on a 0 (no disease) to 9 (severely diseased) rating scale (Xue & Chen Citation2011).
Wheat leaf samples infected with Puccinia triticina were collected by the authors and collaborators across Canada. There were 314 infected leaf collections analysed – 212 from Manitoba, 37 from Saskatchewan, 34 from Ontario, nine each from Quebec and Nova Scotia, seven from Prince Edward Island (PEI), and three each from Alberta and British Columbia. Single pustule P. tritici isolates were recovered from the collections in each province except for those from Alberta, British Columbia and Nova Scotia, in which we could not recover a viable isolate. In total, 399 single pustule isolates were analysed for virulence, including 341 from Manitoba and Saskatchewan, 38 from Ontario, nine from Quebec and 11 from PEI.
There were 41 different virulence phenotypes found among the 399 isolates analysed (). The most common virulence phenotypes found in Canada during 2010 were MLDS (26.1%), TDBJ (22.8%) and TDBG (11.8%) ( and ). These virulence phenotypes were also the most frequently found in Canada during the period 2007–2009 () (McCallum et al. Citation2016). Since the majority of isolates (341/399) were from Manitoba and Saskatchewan, these three pathotypes were also the most frequently found in this region; MLDS (29.6%), TDBJ (25.5%), TDBG (13.8%). MLDS and TDBJ were also the most frequently found virulence phenotypes in the North Central region of the USA during 2010 (Kolmer et al. Citation2012). Leaf rust inoculum blows into Manitoba and Saskatchewan from this North Central region in the USA so there is often a close similarity in the P. triticina populations in these regions. Within Manitoba and Saskatchewan, there were 19 unique virulence phenotypes, some of which have been increasing in recent years, including MFNS (5.3%), TBBG (5.0%), MDPS (4.4%) and MFPS (4.1%). Three virulence phenotypes found in 2010 (PBJQ, MDTS and TDGJ) had not been detected previously in this region (McCallum et al. Citation2016); these are different from each other but similar to virulence phenotypes previously found in this region.
Table 1. Frequency and distribution of virulence phenotypes of Puccinia triticina identified by infection types to selected resistance genes.
Table 2. Frequency (%) of predominant P. triticina virulence phenotypes in Canada from 2000 to 2010.
Of the 38 isolates tested for virulence from Ontario, there were 18 virulence phenotypes. The most commonly found were MBTN (23.7%), TDBJ (10.5%) and MCGJ, MCTN and MLDS (each at 7.9%) (). These were also found frequently in Ontario from 2008–2009 (McCallum et al. Citation2008, Citation2009). There were four virulence phenotypes found in Ontario during 2010 (TCRF, TBBG, PCQR and MCTT) that have not been found in the past 15 years in Ontario (McCallum et al. Citation2016).
There were nine virulence phenotypes (MCGJ, MCTP, MFDS, MFGJ, NBBT, PCQR, PCQT, TBQH and TBRK) found from nine isolates tested from Quebec (). Of these, TBQH, PCQT, PCQR and MCTP were not found since 1998 (McCallum et al. Citation2016). Relatively few isolates have been analysed from Quebec annually so it is difficult to reach firm conclusions, but the population appears to be very diverse since there is a high frequency of unique phenotypes, many of which were not found in previous years.
There were four virulence phenotypes (MBNQ, MBTN, MHNQ and MCSQ) found among 11 isolates from PEI (). Only MBNQ and MHNQ were found in PEI in previous years, although isolates from PEI have only been analysed since 2006, and from that time to 2009, only 36 isolates were analysed in total. MBTN and MCSQ were not previously found in PEI, although MBTN was a common virulence phenotype in Ontario in recent years. As with Quebec and Ontario, the P. triticina population in PEI appears to be more diverse for virulence than that found in western Canada. The PEI population is distinct from the populations in western Canada, but also very different from the Quebec and Ontario populations, as three of the four virulence phenotypes found in PEI were not found in the rest of Canada, similar to previous years. There were no isolates found in PEI that were virulent to Lr2a, Lr2c, Lr9 or Lr18; however, virulence to Lr16 was detected in PEI but not in the rest of the country ().
Table 3. Frequencies of virulence of Puccinia triticina in Canada in 2010 to lines of wheat with single Lr genes for leaf rust resistance and comparison to frequencies from 2009.
For the Manitoba and Saskatchewan samples, the frequencies of virulence to most resistance genes stayed relatively stable compared with 2009 (). However, the frequencies of virulence to Lr2a, Lr14a and Lr24 declined slightly from 2009, whereas virulence to Lr9 and Lr17 increased slightly, following trends in recent years () (McCallum et al. Citation2016). All of these genes are found in wheat cultivars from the USA which can drive the selection for virulence in the P. triticina population (Kolmer et al. Citation2012) that blows into Canada; however, of these, only Lr2a and Lr14a are found commonly in Canadian wheat cultivars (McCallum et al. Citation2016).
Fig. 1 Frequency of virulence (%) from 2000–2010 in the Manitoba and Saskatchewan population of P. triticina to near-isogenic lines containing Lr2a, Lr9, Lr14a, Lr17 or Lr24.
aData from McCallum & Seto-Goh (Citation2003, Citation2004, Citation2005, Citation2006a, Citation2006b, Citation2008, Citation2009) and McCallum et al. (Citation2010, Citation2011, Citation2013).
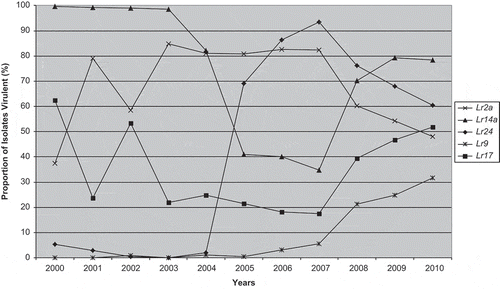
In Ontario, the frequencies of virulence to most resistance genes tested were similar to previous years (), except to Lr9 (7.9%) which is higher than it was previously, following a recent trend of increasing virulence to Lr9 across Canada (). Virulence to Lr18 in 2010 was moderately high in Ontario (23.7%), whereas it was not detected in western Canada, similar to previous years.
The frequency of virulence to Lr18 was the highest in Quebec (66.7%) among all regions surveyed, and in 2010 was the highest this has been in Quebec since 2001 () (McCallum et al. Citation2016). There was no virulence on Lr9 in Quebec, in contrast to the relatively high level of virulence in western Canada (31.7%), emphasizing the differences between the western and eastern populations of P. triticina in Canada.
At least one isolate from all of the unique virulence phenotypes identified was additionally tested for virulence on 12 ‘Thatcher’ near-isogenic lines (Lr2b, Lr3bg, Lr14b, Lr15, Lr19, Lr20, Lr21, Lr23, Lr25, Lr28, Lr29, Lr32 and Lr52) at the seedling stage (). We tested 96 isolates representing all 41 unique virulence phenotypes (as defined by the standard set of 16 differentials) found in Canada during 2010. Results for 59 isolates with differential responses on selected single-gene lines are shown in . All 96 isolates tested had similar responses to Lr2b as they did to Lr2a, were avirulent to Lr19, Lr21, Lr29, Lr32 and Lr52 and were virulent to Lr15 (data not shown). All isolates were also avirulent to Lr25 except 243–1 PBDQ and 244–2 PBJQ (both from Saskatchewan) (data not shown). There were 37, 80, 73, 30 and 44 virulent isolates for Lr3bg, Lr14b, Lr20, Lr23 and Lr28, respectively, among the 96 isolates tested. These results were similar to those from past years.
Table 4. Infection types of 59 representative isolates from Canada in 2010 tested on near isogenic lines of ‘Thatcher’ at the seedling (Lr3bg, Lr14b, Lr20, Lr23, Lr28) or adult-plant stage (Lr12 and Lr37).
These same 96 isolates were also tested for virulence on ‘Thatcher’ near-isogenic wheat lines with Lr12, Lr13, Lr22a, Lr35 or Lr37 at the adult plant stage. All isolates were avirulent to Lr22a, and all but three (243–2 PBJQ, 244–2 PBJQ and 292–2 NBBT) were virulent to Lr13, only six isolates (66–1 MDDS, 184–3 MDPS, 271–1 MBTN, 273–2 PCLR, 290–1 MCTP, 310–1 MHNQ) were virulent to Lr35 (data not shown). There were 78 and 67 isolates virulent to Lr12 and Lr37, respectively ( and data not shown). In past years, nearly all isolates were virulent to Lr12 and Lr13 and all isolates tested to date have been avirulent to Lr22a. The frequency of virulence to Lr37 has been high, whereas virulence to Lr35 was at a relatively low frequency.
Overall the P. triticina population in Manitoba and Saskatchewan followed trends from previous years, and was distinct from the eastern Canadian populations. The Ontario and Quebec populations had some virulence phenotypes in common, but most were unique to one province or the other. The Quebec and Ontario populations were more diverse than the Manitoba and Saskatchewan population, similar to what has been observed in previous years. There were no virulence phenotypes found in all four regions in Canada but there were some that were found in both Manitoba and Saskatchewan and either Ontario or Quebec. There are a number of potential reasons for the differences between the eastern and western populations. The inoculum originates each year from the USA and the Great Plains region contributes to the inoculum in western Canada, whereas the eastern USA growing regions are the source of the inoculum for the eastern Canadian population and the populations in these regions in the USA are significantly different (Kolmer et al. Citation2012). Additionally, the wheat cultivars grown in western and eastern Canada are different, with winter wheat predominating over spring wheat in eastern Canada and spring wheat predominating in western Canada, which could also contribute to differences in these populations. There may also be greater potential for over-wintering in eastern Canada than western Canada. These differences have been seen commonly in previous years (Kolmer Citation1991; Wang et al. Citation2010). The PEI population was different from the other eastern populations, which could be due to the geographic separation of the wheat growing regions in Ontario and Quebec from PEI. The inoculum from the USA coming into PEI would be from the eastern seaboard of the USA, whereas for Ontario and Quebec would be closer to the Ohio Valley region.
Some of the commonly used genes in Canadian wheat cultivars include Lr1, Lr10, Lr13, Lr16, Lr21, Lr22a and Lr34 (McCallum et al. Citation2016). Evolution of virulence in the pathogen population has rendered Lr1, Lr10 and Lr13 relatively ineffective in isolation, whereas no virulence for Lr21, Lr22a and Lr34 was detected in Canada prior to 2010. Virulence to Lr21 was detected for the first time in North America during 2010 when two Lr21 virulent phenotypes (TFBGQ and TFBJQ) were found in the North-Central USA (Kolmer et al. Citation2012). The frequencies of these phenotypes in the sample of the population from this region were relatively low (6% for TFBGQ and 2% for TFBJQ). Virulence to Lr21 was not detected in Canada in 2010, but virulent isolates could have gone undetected if the proportion of isolates with Lr21 virulence was very low in the population. The information generated in this study can be integrated with virulence results from previous years and used along with information on the resistance genes in current and future wheat cultivars to make informed decisions when breeding wheat in Canada for leaf rust resistance. Continuous surveillance of the pathogen population in Canada is essential for early detection of new virulence phenotypes that can attack the Lr genes in Canadian wheat cultivars and cause serious losses due to leaf rust epidemics.
Acknowledgements
We thank André Comeau, Harpinder Randhawa and Richard Martin who sent samples for analysis, and other cooperators who grew trap rust nurseries.
References
- Anikster Y, Bushnell WR, Eilam T, Manisterski J, Roelfs AP. 1997. Puccinia recondita causing leaf rust on cultivated wheats, wild wheats, and rye. Can J Bot. 75:2082–2096.
- Kolmer JA. 1991. Phenotypic diversity in two populations of Puccinia recondita f,sp. tritici in Canada during 1931–1987. Phytopathology. 81:311–315.
- Kolmer JA, Long DL, Hughes ME. 2012. Physiologic specialization of Puccinia triticina on wheat in the United States in 2010. Plant Dis. 96:1216–1221.
- Long DL, Kolmer JA. 1989. A North American system of nomenclature for Puccinia recondita f.sp tritici. Phytopathology. 79:525–529.
- McCallum BD, Fetch T, Chong J. 2007. Cereal rust control in Canada. Aust J Agric Res. 58:639–647.
- McCallum BD, Hiebert CW, Cloutier S, Bakkeren G, Rosa SB, Humphreys DG, Marais GF, McCartney CA, Panwar V, Rampitsch C, et al. 2016. A review of wheat leaf rust research and the development of resistant cultivars in Canada. Can J Plant Pathol. 38:1–18.
- McCallum BD, Seto-Goh P. 2003. Physiologic specialization of wheat leaf rust (Puccinia triticina) in Canada in 2000. Can J Plant Pathol. 25:91–97.
- McCallum BD, Seto-Goh P. 2004. Physiologic specialization of Puccinia triticina, the cause of wheat leaf rust, in Canada in 2001. Can J Plant Pathol. 26:109–120.
- McCallum BD, Seto-Goh P. 2005. Physiologic specialization of wheat leaf rust (Puccinia triticina) in Canada in 2002. Can J Plant Pathol. 27:90–99.
- McCallum BD, Seto-Goh P. 2006a. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2003. Can J Plant Pathol. 28:208–213.
- McCallum BD, Seto-Goh P. 2006b. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2004. Can J Plant Pathol. 28:566–576.
- McCallum BD, Seto-Goh P. 2008. Physiologic specialization of Puccinia triticina in Canada in 2005. Can J Plant Pathol. 30:124–132.
- McCallum BD, Seto-Goh P. 2009. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2006. Can J Plant Pathol. 31:80–87.
- McCallum BD, Seto-Goh P. 2011. Leaf rust and stripe rust of wheat in Manitoba and eastern Saskatchewan in 2010. Can Plant Dis Surv. 91:98.
- McCallum BD, Seto-Goh P, Xue A. 2010. Physiological specialization of Puccinia triticina in Canada in 2007. Can J Plant Pathol. 32:229–236.
- McCallum BD, Seto-Goh P, Xue A. 2011. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2008. Can J Plant Pathol. 33:541–549.
- McCallum BD, Seto-Goh P, Xue A. 2013. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2009. Can J Plant Pathol. 35:338–345.
- Wang X, Bakkeren G, McCallum B. 2010. Virulence and molecular polymorphisms of the wheat leaf rust fungus Puccinia triticina in Canada from 1997 to 2007. Botany. 88:575–589.
- Xue AG, Chen Y. 2011. Disease of spring wheat in eastern Ontario in 2010. Can Plant Dis Surv. 91:105–106. Available from: http://www.cps-scp.ca/cpds.shtml