Abstract
Popovičová, J. 2009. Water quality assessment and ecoregional comparison of a reservoir in east-central Indiana. Lake Reserv. Manage. 25:155–166.
This study assessed water quality of a reservoir in an agricultural watershed of east-central Indiana, examined the effects of a thermal and oxygen regime on cycling of nutrients, and compared the results to ecoregional data and reference guidelines. Two locations were monitored biweekly from May through September 2007 for pH, temperature, dissolved oxygen, alkalinity, Secchi disk transparency (SD), chlorophyll a, and nutrient concentrations. The reservoir did not stratify during the monitoring season, although both anoxia and reoxygenation of the hypolimnion were observed. These conditions affected nitrogen (N) and phosphorus (P) cycling because nitrification was found to occur in the hypolimnion, and both internal load and water column mixing affected the concentrations and distribution of P. The reservoir was characterized as a eutrophic water body based on SD, total N and chlorophyll a concentrations, while total P concentrations classified the reservoir as slightly hypereutrophic (TSI = 73). This P overload has shifted the system toward N-limiting conditions (molar TN:TP = 18). Comparison of trophic parameters to Ecoregion 55 data placed this reservoir within the 75th percentile, and all parameters exceeded the U.S. Environmental Protection Agency ecoregional reference guidelines. I discuss a potential restoration of this water body to comply with the ecoregional nutrient criteria and to avoid future deterioration associated with N limitation.
The consequences of cultural eutrophication negatively affect designated uses of lakes and reservoirs, and restoration of eutrophic or hypereutrophic conditions will depend on the natural levels of nutrients influenced by regional environmental characteristics (CitationOmernik 1987, Citation1995, CitationFulmer and Cooke 1990, CitationHeiskary et al. 1987, CitationSmith et al. 2003). Ecoregions—land areas with similar land cover, land use, soils and natural vegetation (CitationOmernik 1987)—have been employed to assess ecoregional differences in lake and stream water quality, establish reference conditions for nutrients for development of management strategies, and to determine restoration potential for lakes and reservoirs (CitationHeiskary et al. 1987, CitationHughes and Larsen 1988, CitationFulmer and Cooke 1990, CitationSmith et al. 2003, CitationDodds et al. 2006). However, the validity and success of this approach varies. Ecoregional characterization was successfully applied to lakes in Minnesota and reservoirs in Ohio and Kansas (CitationHeiskary et al. 1987, CitationFulmer and Cooke 1990, CitationDodds et al. 2006). This approach, however, was not appropriate for classification of water quality distribution among Northeastern and Michigan lakes (CitationJenerette et al. 2002, CitationCheruvelil et al. 2008). A local monitoring program was found to be more accurate and useful for setting lake-specific conditions and management than the utilization of regional criteria due to large variations in nutrients and chlorophyll a relationship among lakes and reservoirs, even within the same region (CitationHatch 2003, CitationHavens 2003).
Prairie Creek Reservoir, Indiana, serves for fisheries, recreation, and as a secondary drinking water supply. The reservoir is located in east-central Indiana in Ecoregion 55, Eastern Corn Belt Plains, where eutrophication problems are among the worst in the state of Indiana (CitationIDEM 2006, CitationJones and Medrano 2006). About 75% of the land in the ecoregion is used for crops such as corn and soybeans. The ecoregion is characterized by gently rolling till plains, has few natural lakes, and the soil is poorly drained. Until 2005 water quality investigations at the reservoir were limited to one-time sampling of trophic parameters in 1996 and 2002 (CitationJones and Barnes 2005, CitationJones and Medrano 2006). In 2006 CitationPopovičová (2008) found high concentrations of phosphorus and anoxic conditions; however, analyses of total nitrogen (TN) and total phosphorus (TP), the key parameters to determine trophic status, were not included. To date, analysis of phytoplankton composition has been almost nonexistent even though some cases of algal blooms had been reported to local laboratories. CitationJones and Medrano (2006) found that blue-green algae comprised 41% of the total plankton in 2002, and Cylindrospermopsis raciborskii was measured at 84,247 cells/mL although no toxin was detected (CitationJones and Sauter 2005). The goal of the present study was to assess eutrophication of Prairie Creek Reservoir, compare the results to the ecoregional data, and determine its restoration potential. Specific objectives were to assess the extent of anoxia and its effects on distribution and cycling of nutrients, determine the limiting nutrient, and analyze available data for Ecoregion 55 to compare the reservoir to ecoregional conditions and reference guidelines.
Materials and methods
Prairie Creek Reservoir (5.1 km2) is located in a watershed (44 km2) of the White River in east-central Indiana (). Land use in the watershed is dominated by agriculture (64.7%), and 13.3% is reserved for green space (CitationDMMPC 2007). Soil is drained by subsurface tiles and surface drains. Mean reservoir depth is 4.7 m, maximum depth is 9.1 m and maximum volume of water is 23.9 × 109 m3.
Figure 1 Location of Prairie Creek Reservoir and sampling sites.
Biweekly sampling and monitoring was performed in the north (8.5 m depth) and center (7 m depth) of the reservoir between 10:00 and 15:00 from May through September 2007 (). Samples for nutrients and alkalinity were collected from the epilimnion at 0.5 m depth and from hypolimnion at 0.5 m from the bottom using a horizontal beta sampler. Samples were transported in high-density polyethylene NalgeneTM bottles on ice, stored at 4°C, and analyzed within 24 hours of collection (CitationAPHA 1998).
Temperature, dissolved oxygen (DO), and chlorophyll a were measured in-situ at 0.5-m intervals from surface to bottom using a Hydrolab Sonde DS5 (Hach, Inc., Loveland, CO). Secchi disk (SD) transparency was measured according to CitationCarlson and Simpson (1996), and a pocket weather meter (Kestrel 2000, Nielsen-Kellerman, Boothwyn, PA) was used to measure wind speed at the time of sample collection. Chlorophyll a concentrations were calculated for the euphotic zone (euphotic depth = 1.7 × SD; CitationScheffer 2004). Alkalinity was analyzed by titration with a 0.02 N sulfuric acid, nitrates (NO3-N) by the cadmium reduction method 4500-NO3-E, and ammonia (NH3-N) by the salicylate method (CitationAPHA 1998). Samples for analysis of soluble reactive phosphorus (SRP) were filtered through a membrane filter (0.45 μm) and then analyzed by the ascorbic acid method 4500-P-E (CitationU.S. EPA 1997, CitationAPHA 1998). Total nitrogen was analyzed by persulfate digestion, and TP was determined using the ascorbic acid method after digestion with acid persulfate. Quality assurance and quality control procedures were employed according to standard methods (CitationAPHA 1998).
Descriptive statistics and analysis of variance for determination of differences among sampling sites, months, and depths were determined using MiniTab® 15.1.0.0 (Minitab Inc., State College, PA). To satisfy the requirement of normal distribution and equal variances, data were tested for normality using the Kolmogorov-Smirnov test and the Leven test for homogeneity of variances. Due to lack of normality, all data were log-transformed. Spearman's rho correlation analysis was used to determine relationships between variables. Trophic state index (TSI) for SD, chlorophyll a, and TP was calculated for July–August data using Carlson's equations (CitationCarlson 1977), and the CitationKratzer and Brezonik (1981) equation was used to calculate TSI based on TN.
Data for Indiana lakes and reservoirs in Ecoregion 55 were obtained from Jones and Medrano (Citation2006; N = 36), and a median and range for each relevant parameter were calculated. The reference nutrient criteria for lakes and reservoirs in Level III Ecoregion 55 and Ecoregion VI were compared to the collected data (CitationU.S. EPA 2000).
Results
Physicochemical water quality parameters
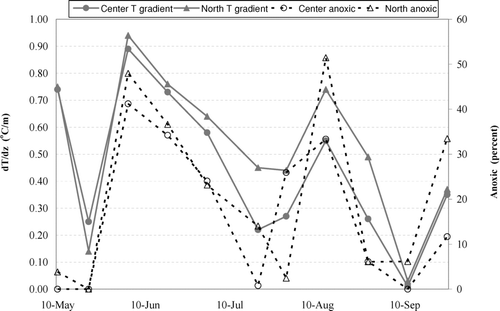
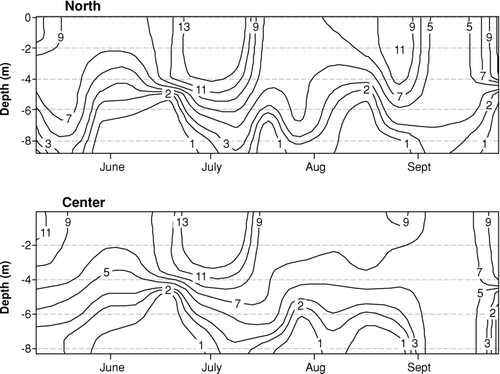
During the entire monitoring period, results for pH and water temperature did not violate Indiana water quality standards (CitationIAC 2008). In situ pH was significantly higher in the epilimnion but no temporal differences were identified (). Hypolimnetic temperatures in the center location were higher, and the temperature gradient at both locations indicated lack of thermal stratification at the reservoir (dT/dz < 1°C; CitationStefan et al. 1996; and ). Although not statistically different, the center location had lower temperature gradients (0.01°C/m ⩽ dT/dz ⩽ 0.89°C/m) than did the north location (0.03°C/m ⩽ dT/dz ⩽ 0.94°C/m; and ).
Table 1 Water quality parameters measured at the reservoir from May to September 2007.
Figure 2 Thermal regime at CitationPrairie Creek Reservoir in 2007.
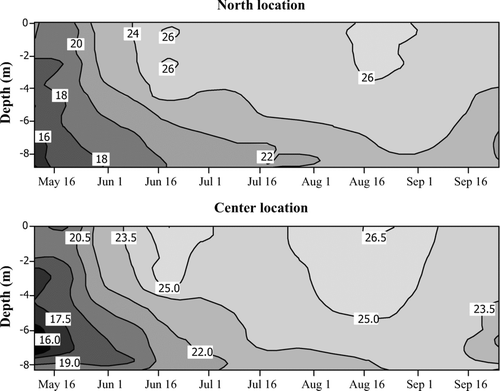
Figure 3 Temperature gradient and anoxic conditions at the reservoir in 2007.
Epilimnetic alkalinity showed a decreasing trend at both locations from May until the end of September indicating photosynthetic activity. Hypolimnetic alkalinity increased significantly (p< 0.01) in July and August from that measured in the epilimnion and was slightly higher at the north location (). At the center location in mid-August the hypolimnetic alkalinity decreased to the original level measured in spring while at the north location high alkalinity persisted until the end of the month (). The results imply that decomposition was most intense in July and August and persisted longer at the north location.
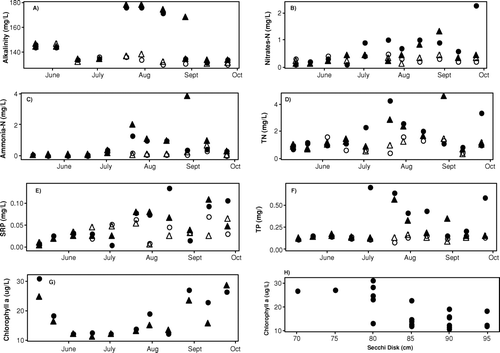
Figure 4 Time series graph for a) alkalinity, b) nitrates c) ammonia-N, d) total nitrogen, e) soluble reactive phosphorus, f) total phosphorus, g) chlorophyll a, and h) chlorophyll a vs. Secchi disk. Symbols in graphs a) through f) represent: ◯ Center epilimnion, Δ North epilimnion. • Center hypolimnion, ▴ North hypolimnion; in graph g) • represents Center and ▴ North location.
The results for DO violated the Indiana water quality standard of 5 mg/L in 95% of the hypolimnion samples and in 5% of epilimnion samples (CitationIAC 2008; ). Epilimnetic DO concentrations did not exhibit significant temporal differences, although they were significantly higher (p< 0.001) than in the hypolimnion (; ). Hypolimnetic DO concentrations measured on 4 June 2007 indicated the establishment of anoxic conditions with a DO concentration of 0.31 mg/L at the north and 0.32 mg/L at the center location. In general, anoxia persisted until the end of September, although several days of hypolimnetic reoxygenation and mixing were recorded (). At the north location DO concentration in the hypolimnion increased to 1.94 mg/L on 30 July, after which anoxia persisted to varying depths until the end of the monitoring season. At the center, more shallow location, hypolimnetic DO concentrations increased on 20 July (1.37 mg/L), 28 August (1.24 mg/L), and 11 September (5.89 mg/L), whereas 25 September was marked by anoxia (0.4 mg/L).
Figure 5 Vertical and temporal variability of dissolved oxygen concentrations at CitationPrairie Creek Reservoir in 2007.
Trophic parameters
The months characterized by anoxic conditions in the reservoir were also characterized by increasing concentrations of nutrients ( and ). With one exception, NO3-N concentrations met the Indiana water quality standard for aquatic life (1.6 mg/L, n = 44; ; CitationIAC 2008). Epilimnetic NO3-N concentrations showed little temporal difference even as chlorophyll a concentration increased (); however, hypolimnetic concentrations were significantly higher (p< 0.05; ) and increased throughout the monitoring period, especially, at the center location where reoxygenation of the hypolimnion occurred more frequently. The maximum NO3-N concentration was measured on 25 September (2.3 mg/L) at the center following the increase of hypolimnetic DO concentration to 5.89 mg/L ().
Concentrations of NH3-N in the hypolimnion and epilimnion were significantly different (p < 0.001; ; ), although temporal differences were found only in the hypolimnion (p < 0.01). Epilimnetic NH3-N concentrations increased at the end of the season when mixing of the water column was observed. Increased hypolimnetic NH3-N concentrations were measured in July and August, and the maximum was reached in August at the north location (3.8 mg/L). On 25 September concentrations approached those found in May, although NH3-N production in the hypolimnion was still occurring. Concentrations of TN followed the temporal trend of NO3-N and NH3-N with a greater temporal variability in the epilimnion. Hypolimnetic TN concentrations were significantly greater (p < 0.05) than in the epilimnion (; ) and, in general, had a trend similar to NH3-N in that concentrations increased until July, then declined, and the maximum was reached at the end of August at the north location (). Concentrations of TN characterized the reservoir as eutrophic (TSI = 56; CitationKratzer and Brezonik 1981).
Contrary to the distribution of N compounds, the soluble reactive phosphorus (SRP) concentrations increased in both epilimnion and hypolimnion as summer progressed, and hypolimnetic concentrations exceeded those found in the epilimnion starting mid-July (; ). The increased SRP concentrations in the hypolimnion are typical of anoxic conditions that began in late May and, with the exception of some mixing periods, persisted until the end of August in the center and until September in the north. Concentrations of TP in the hypolimnion were significantly higher than in the epilimnion (p < 0.001; ; ), even though only hypolimnetic TP concentrations showed significant temporal differences (p < 0.05) and reached their maximum in July (; ). High concentrations of TP found at the reservoir characterized this water body as slightly hypereutrophic (TSI = 73; CitationCarlson 1977).
Algal biomass, as measured by chlorophyll a concentration, reached its minimum in June when “stratification” (dT/dz) was greatest and increased until the end of the monitoring period, reaching its maximum in September (, and ). The late spring decline measured in June could be attributed to the clear-water phase that often occurs at the end of spring (CitationLampert et al. 1986, CitationScheffer 2004) due to depletion of available nutrients caused by the spring bloom of diatoms. Although not significantly different, chlorophyll a concentrations were slightly higher at the center location, where more mixing occurred and higher SRP concentrations were measured (). Concentrations of chlorophyll a characterized the reservoir as a eutrophic water body (TSI = 58). Secchi disk transparency correlated inversely with chlorophyll a concentrations (r2= 0.67, p< 0.001; ; ). Maximum water transparency was measured in June (0. 93 m) and the minimum in August (0.80 m; ), although no significant temporal differences were found. Similar to TSI calculated from TN and chlorophyll a, SD results were characteristic of a eutrophic reservoir (TSI = 62).
Discussion
The results of this study demonstrate distribution and cycling of nutrients in a polymictic reservoir where both anoxic periods and intermittent mixing were observed. A 2006 monitoring study found weak stratification of the water column in July (dT/dz = 1.13 and 1.3°C/m in the center and north location, respectively; CitationPopovičová 2008); however, this was not the case in 2007. This type of thermal regime would be expected for the geometry ratio (A0.25: zmax) of 5.2 (CitationStefan et al. 1996). Contrary to the findings of Stefan that anoxic conditions never occur in lakes with the ratio >5, however, anoxia was observed in this reservoir. Furthermore, reoxygenation of the hypolimnion was observed, especially at the shallower center location, which can be attributed to lack of thermal stratification, depth, and contribution of additional factors such as wave action and mixing caused by wind and motorboats that control vertical mixing (CitationScheffer 2004, CitationKelton and Chow-Fraser 2005). The first two weeks of July at this reservoir experienced especially heavy motorboat traffic (). Local weather data revealed that storm events occurred prior to monitoring days when hypolimnetic reoxygenation was measured. Precipitation was recorded 18 July (5.6 mm), 19 July (16 mm), 16–25 August (8.9–60.7 mm), and 7–9 September (3.8–41.6 mm). Major storms can have significant effects on water column mixing (CitationHerb and Stefan 2004) and, consequently, can induce oxygenation of the hypolimnion (). Depth may play a role as well because the center, more shallow location (∼7 m) was characterized by a lower temperature gradient and stronger winds (mean speed of 8.85 km/h vs. 8.05 km/h) making it more susceptible to mixing and, thus, hypolimnetic reoxygenation (), even though significant differences in DO concentrations were not found between the monitored locations.
The alternating periods of anoxia and mixing affected the cycling and distribution of nutrients throughout the water column. In shallow reservoirs water mixing can enhance exchange of nutrients between the sediment and the water column (CitationScheffer 2004). Increased epilimnetic concentrations during the monitoring season were observed especially for SRP, although NO3-N and TN showed some variability as well (). Increased concentrations of NO3-N in the hypolimnion during the season were the consequence of organic matter decomposition and subsequent nitrification of ammonia due to reoxygenation of the water column. Ammonification of detritus creates suitable conditions for nitrification when sufficient oxygen becomes available at the top of the sediment (CitationLijklema 1994, CitationScheffer 2004). In stratified lakes the zones of nitrification move throughout the water column depending on the presence of oxygen and ammonia, and move to the sediment-water interface when stratification ceases to occur (CitationStewart et al. 1982). In shallower, mixed lakes (e.g., Prairie Creek Reservoir), however, nitrification mainly occurs at the sediment-water interface in the presence of any available oxygen (CitationStewart et al. 1982, CitationLijklema 1994, CitationScheffer 2004). Increased NO3-N concentrations in the hypolimnion are especially evident in the center, shallower location where several events of reoxygenation were measured ( and ). Additionally, nitrification can be enhanced by diurnal temperature fluctuations that can induce vertical mixing of the water column, and, consequently, oxygen replenishment (CitationHerb and Stefan 2004, CitationScheffer 2004). However, the biweekly water column measurements of temperature (T) and DO are not adequate for investigation of diurnal variations at this reservoir. The results of this study demonstrate that nitrification occurred in the hypolimnion as a result of water column mixing and reoxygenation.
High concentrations of NH3-N in the hypolimnion are expected when anoxic conditions and higher amounts of decaying algae are present as summer progresses (CitationScheffer 2004). Increased production of NH3-N due to ammonification in the bottom sediment resulted in subsequent increase in concentrations of NO3-N throughout the season. Concentrations of NH3-N increased significantly in July and then decreased as the season progressed with one exception recorded at the north location in late August. Hypolimnetic NO3-N concentrations, however, increased throughout July and August due to nitrification of ammonia generated when dissolved oxygen was present as a result of mixing. Concentrations of about 1 mg/L of DO were found to support nitrification, while conditions favorable to denitrification are around 0.2 mg/L (CitationKnowles et al. 1981). In this study the concentrations of DO in the hypolimnion increased above 1 mg/L on several occasions, leading to nitrification ( and ).
Vertical mixing of water and variations in DO concentrations at the sediment-water interface also influence P cycling between the sediment and water column (CitationHerb and Stefan 2004). In shallow lakes this mixing can have two effects on P fate (CitationScheffer 2004). Periods of reduced mixing and turbulence can lead to anoxic conditions at the sediment surface that cause release of P (CitationNürnberg and Peters 1984, CitationNürnberg 1994, CitationScheffer 2004, CitationKelton and Chow-Fraser 2005), while during periods of increased mixing P is distributed throughout the water column. Resuspension and oxygenation of sediment is related to lake size and depth, and this reservoir is classified as a water body where both forms of P release occur (CitationScheffer 2004). This is supported by the present study where internal P release, as well as increased epilimnetic concentrations of SRP, were observed, indicating the contribution of P from the sediment and hypolimnion as a result of anoxic conditions and intermittent mixing (, and ). The estimated internal P load calculated from the in situ summer increases in water column TP concentration was 1018 mg/m2 at the north and 1803 mg/m2 at the center location based on the maximum water column concentrations measured in July at both locations (CitationNürnberg 1987).
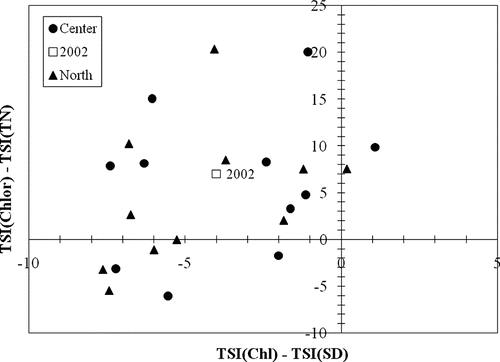
Although high summer TP concentrations characterized this reservoir as slightly hypereutrophic, the symptom of excessive algae scum was not observed in the reservoir. The absence of extensive algal growth, which would be expected at the measured TP concentrations, could be attributed to several factors such as lack of stratification, mixing of the water column, reoxygenation of the hypolimnion, nutrient limitations, and nonalgal turbidity. First, in a lake with a mixed water column, epilimnetic concentrations of chlorophyll a are lower than in a stratified lake because algae is not confined to the surface as it is in a stratified lake (CitationHavens 2003). Furthermore, historical data suggest an increasing trend in TP loading at the reservoir (CitationJones and Barnes 2005) that has caused a shift in nutrient limitation (). The mean summer TN:TP ratio of 18 indicates a co-limitation of N and P and a shift toward N-limiting conditions (CitationElser et al. 1990, CitationGuildford and Hecky 2000, CitationDzialowski et al. 2005). A decrease in N/P ratio develops with increasing eutrophication and can be related to higher P load from the watershed (CitationLijklema 1994). Other studies found that lakes initially identified as P-limited had shifted to N-limitation due to excessive P loading (CitationNürnberg and Peters 1984, CitationElser et al. 1990, CitationHavens 1995, CitationJames et al. 1995, CitationHavens et al. 1996). Finally, nonalgal turbidity can play a significant role in phytoplankton production due to light limitation (CitationScheffer 2004, CitationCarlson and Havens 2005). Indiana lakes and reservoirs were found to be affected by nonalgal turbidity (CitationJones and Medrano 2006); however, the results for this reservoir showed that nonalgal turbidity does not significantly limit algal growth because the correlation between TSI(SD) and TSI(Chlor) is stronger (r2= 0.65, p < 0.001) than correlation between TSI(SD) and TSI(TP) (r2= 0.26, p = 0.245; Carlson and Havens 2005). Additionally, differences between the TSI results for TN, SD and chlorophyll a fit in the upper left quadrant of a deviation plot, suggesting the effect of nitrogen (.) Therefore, algal growth at the reservoir is most likely limited by nitrogen rather than nonalgal turbidity, although bioassay analyses would be useful to confirm these findings in the future (CitationCarlson and Havens 2005).
Table 2 Historical changes in trophic levels at Prairie Creek Reservoir.
Figure 6 TSI deviation plot for Prairie Creek Reservoir in 2007. Data for 2002 are from CitationJones and Medrano (2002).
Comparison to lakes and reference conditions of Ecoregion 55
Compared to other Indiana lakes and reservoirs in Ecoregion 55, the percent of water column under oxic conditions was higher in Prairie Creek Reservoir than in 50% of water bodies of this ecoregion (). However, these ecoregional conditions only reflect an instantaneous state measured on the day of monitoring that can change depending on weather patterns, wind, boat traffic, and stratification (CitationScheffer 2004, CitationKelton and Chow-Fraser 2005, CitationKnowlton and Jones 2006). The lowest DO concentrations at Prairie Creek Reservoir were measured in June when about 40% of the bottom water column was anoxic; similar conditions were also observed in 2006 (CitationPopovičová 2008). Considering the nutrient concentrations in this reservoir, the lack of thermal stratification alleviated anoxia, reduced the extent of the anoxic layer, and prevented an even a greater release of TP from the sediment (CitationScheffer 2004).
Table 3 Comparison of CitationPrairie Creek Reservoir 2007 data to other Indiana lakes and reservoirs in Ecoregion 55 and the reference values.
The Secchi disk transparency in the reservoir was only slightly lower than the median for Ecoregion 55 (). Concentrations of TN were within the 50th percentile of the ecoregional levels, while NO3-N, NH3-N, TP, and chlorophyll a concentrations were within the 75th percentile. A review of the ecoregional data acquired between 1999 and 2003 (CitationJones and Medrano 2005) showed that in 2002 the reservoir had better water quality in comparison to other lakes and reservoirs of the same ecoregion; nitrates were within the 25th percentile, TN and TP values were within the 50th percentile, and chlorophyll a was in the 75th percentile of the ecoregional data. In 2005 Jones and Barnes reported that the Indiana TSI (ITSI) for this water body, a scoring system developed specifically for lakes and reservoirs in the state, changed from mesotrophic (ITSI = 24) to hypereutrophic status (ITSI = 55; Jones and Barnes 2005, Jones and Medrano 2006).
The SD transparency decreased significantly from 2.7 m in 1996 to 0.7 m in 2002 (CitationJones and Barnes 2005). These results demonstrate degradation of the reservoir water quality due to eutrophication between 1996 and 2007 that placed the reservoir among 10% of hypereutrophic water bodies in the state (). This decrease in water quality is a result of nutrient loading, increased load of suspended solids, and absence of adequate land management practices. Historical trends in TSI (CitationCarlson 1977) show that while levels of N remained stable between 1996 and 2007, P load has increased considerably, consequently affecting SD transparency and chlorophyll a concentrations, although SD transparency and chlorophyll a concentrations did not change significantly between 2002 and 2007 (). These trends also explain the shift toward the N-limited system (CitationGuildford and Hecky 2000, CitationDzialowski et al. 2005) because N limitation has been found to occur in lakes with excessive P input (CitationNürnberg and Peters 1984, CitationJames et al. 1995, CitationHavens et al. 1996). This finding is of great consequence for the management of this reservoir because N-limiting conditions produce blooms of green and blue-green algae that can generate toxins and obnoxious taste and odor compounds (CitationHavens 1995, CitationHavens et al. 1996, CitationJohnston and Jacoby 2003). Decreasing N inputs, however, might not decrease cyanobacteria growth in N-limited systems (CitationScheffer 2004), and management of both N and P is necessary to control production of cyanobacteria (CitationDzialowski et al. 2005). Current management practices in the reservoir watershed are limited to only minimum tillage agriculture; use of other strategies such as buffer strips, livestock fencing, use of constructed wetlands at the reservoir inlets and in-lake management practices have not been implemented.
To determine the reservoir restoration potential, water quality parameters were compared to the reference guidelines and the corresponding TSI proposed by the CitationU.S. EPA (2000) because Indiana does not have sufficient data for development of its own criteria. All parameters measured in 2007 exceeded the reference guidelines for the ecoregion ( and ), and the attainment of the proposed levels would restore the reservoir water quality to the level found in 1996 (CitationU.S. EPA 2000, CitationJones and Sauter 2005; and ). Such comparisons, however, could be negatively affected by inadequate long-term data for this water body needed to account for temporal variability. CitationKnowlton and Jones (2006) showed that reservoirs with long-term means for TP and chlorophyll a concentrations in compliance with the reference nutrient criteria often exceeded those criteria when individual values or seasonal averages were considered. Numeric criteria should therefore be applied only to long-term averages; otherwise, compliance with the criteria can be misclassified. Consequently, monitoring at this reservoir should continue to obtain at least four seasons of data to assess typical conditions and variability (CitationKnowlton and Jones 2006).
The suitability of an ecoregional approach to guide restoration and management of Indiana lakes and reservoirs has not been established. Ecoregional comparisons performed in other states have used extensive databases of lakes data spanning several years (CitationJenerette et al. 2002, CitationHavens 2003, CitationDodds et al. 2006, CitationKnowlton and Jones 2006, CitationCheruvelil et al. 2008), while Indiana lakes and reservoirs were monitored once between 1994 and 1998 and once between 1999 and 2003. In both monitoring periods only one measurement was performed per water body (CitationJones and Sauter 2005, CitationJones and Medrano 2006). Although CitationJones and Medrano (2006) attempted to classify lakes and reservoirs according to ecoregions, significant differences among them were not established. Additionally, the results for Ecoregion 55 demonstrate very high variability, especially in SD and chlorophyll a concentrations (). If reference conditions were based on these data, reference levels for chlorophyll a would be set at 0.87 μg/L (lower 25th percentile) or 0.66 μg/L (trisection method), and reference conditions for SD would be set at 1.8 m (25th percentile) or at 2.7 m (trisection method); such conditions, especially for chlorophyll a, would be unattainable for this reservoir.
Prairie Creek Reservoir showed high temporal variability of nutrients caused by lack of stratification and intermittent reoxygenation of the hypolimnion. Trophic parameters showed worsening of eutrophic conditions and development of N limitation. Comparison of trophic variables to the ecoregional levels placed the reservoir in the 75th percentile. Restoration would be possible if management practices were implemented to prevent further deterioration and to avoid potential cyanobacterial blooms in the future that could threaten its uses for recreation and drinking water (CitationDodds et al. 2006). However, the comparison analysis was limited by availability of historical data for this reservoir as well as Ecoregion 55. Therefore, further monitoring is necessary to establish ecoregional reference conditions for Indiana lakes and reservoirs and to assess significant differences in water quality among ecoregions. Such assessment should also consider classification of lakes and reservoirs within smaller groups, such as eight-digit hydrologic units (CitationCheruvelil et al. 2008) or subregions (CitationHughes and Larsen 1988, CitationSmith et al. 2003) due to large variability of reference data and natural background concentrations (CitationSmith et al. 2003). Additionally, further long-term investigation of this reservoir would be beneficial to assess temporal variability of trophic parameters and determine more accurate lake-specific management strategies. Finally, an investigation of diurnal changes in temperature, dissolved oxygen and nutrients would be valuable to gain a better understanding of nitrification-denitrification processes and nutrient cycling in the reservoir.
Acknowledgments
The author is grateful to Diana Fiallos Celi for her work on field monitoring and laboratory analyses. This project was funded by Ball State University (BSU), Indiana Academy of Science, and a Lilly V grant.
Notes
†Epilimnion.
††Hypolimnion.
+Soluble reactive phosphorus.
++Euphotic zone.
*Depth difference significant at p< 0.05.
**Depth differences significant at p< 0.01.
***Depth differences significant at p< 0.001.
†data from CitationJones and Sauter (2005).
*data from CitationJones and Medrano (2006).
+calculated from CitationKratzer and Brezonik (1981).
††calculated from CitationCarlson (1977).
†epilimnetic values from July and August for the ecoregion (data from CitationJones and Medrano 2006); data in parentheses denote minimum and maximum.
*reference guidelines for Ecoregion 55; median value from 4 seasons—25th percentile % for each season (CitationU.S. EPA 2000).
+reference guidelines for Ecoregion VI (CitationU.S. EPA 2000).
††July and August epilimnetic values.
**nitrate + nitrite.
References
- APHA . 1998 . Standard methods for the examination of water and wastewater , 20th , Edited by: Clesceri , L. S. , Greenberg , A. E. and Eaton , A. D. Washington , D.C. : American Public Health Association .
- Carlson , R. E. 1977 . A trophic state index for lakes . Limnol. Oceanogr. , 22 ( 2 ) : 361 – 369 .
- Carlson , R. E. and Simpson , J. 1996 . A coordinator's guide to volunteer lake monitoring methods , North American Lake Management Society .
- Carlson , R. E. and Havens , K. E. 2005 . Graphical methods for the interpretation of relationships between trophic state variables . Lake Reserv. Manage. , 21 : 107 – 118 .
- Cheruvelil , K. S. , Soranno , P. A. , Bremigan , M. T. , Wagner , T. and Martin , S. L. 2008 . Grouping lakes for water quality assessment and monitoring: The roles of regionalization and spatial scale . Environ. Manage. , 41 : 425 – 440 .
- DMMPC . 2007 . “ Prairie Creek Reservoir Master Plan ” . In Delaware-Muncie Metropolitan Plan Commission http://www.whiteriverwatershedproject.org/prairiecreekmasterplan.html Accessed 19 June 2008
- Dodds , W. K. , Carney , E. and Angelo , R. T. 2006 . Determining ecoregional reference conditions for nutrients, Secchi depth and chlorophyll a in Kansas lakes and reservoirs . Lake Reserv. Manage. , 22 ( 2 ) : 151 – 159 .
- Dzialowski , A. R. , Wang , S. , Lim , N. , Spotts , W. W. and Huggins , D. G. 2005 . Nutrient limitation of phytoplankton growth in central plains reservoirs, USA . J. Plankton Res. , 27 : 587 – 595 .
- Elser , J. J. , Marzolf , E. R. and Goldman , C. R. 1990 . Phosphorus and nitrogen limitation of phytoplankton growth in the freshwaters of North America: A review and critique of experimental enrichments . Can. J. Fish. Aquat. Sci. , 47 ( 7 ) : 1468 – 1477 .
- Fulmer , F. G. and Cooke , G. D. 1990 . Evaluating restoration potential of 19 Ohio reservoirs . Lake Reserv. Manage. , 6 ( 2 ) : 197 – 206 .
- Guildford , S. J. and Hecky , R. E. 2000 . Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol . Oceanogr. , 45 : 1213 – 1223 .
- Hatch , L. K. 2003 . Factors affecting Iowa lake and reservoir water quality . Lake Reserv. Manage. , 19 ( 2 ) : 150 – 159 .
- Havens , K. E. 1995 . Secondary nitrogen limitation in a subtropical lake impacted by non-point source agricultural pollution . Environ. Pollut. , 89 ( 3 ) : 241 – 246 .
- Havens , K. 2003 . Phosphorus – algal bloom relationships in large lakes on South Florida: Implications for establishing nutrient criteria . Lake Reserv. Manage. , 19 ( 3 ) : 222 – 228 .
- Havens , K. E. , Aumen , N. G. , James , R. T. and Smith , V. H. 1996 . Rapid ecological changes in a large subtropical lake undergoing cultural eutrophication . Ambio , 25 ( 3 ) : 150 – 155 .
- Heiskary , S. C. , Wilson , B. and Larsen , D. P. 1987 . Analysis of regional patterns in lake water quality: Using ecoregions for lake management in Minnesota . Lake Reserv. Manage. , 3 : 337 – 344 .
- Herb , W. R. and Stefan , H. G. 2004 . Temperature stratification and mixing dynamics in a shallow lake with submersed macrophytes . Lake Reserv. Manage. , 20 ( 4 ) : 296 – 308 .
- Hughes , R. M. and Larsen , D. P. 1988 . Ecoregions: an approach to surface water protection . Journal of Water Pollution Control Federation , 60 ( 4 ) : 496 – 493 .
- IAC . 2008 . Indiana Administrative Code, Title 327 Water pollution Control Board, Article 2: Water Quality Standards
- IDEM . 2006 . Indiana water quality Indiana's integrated water monitoring and assessment report , Indiana Department of Environmental Management .
- James , R. T. , Smith , V. H. and Jones , B. L. 1995 . Historical trends in the Lake Okeechobee ecosystem . Arch. Hydrobiol. , 107 ( 1 ) : 49 – 69 .
- Jenerette , G. D. , Lee , J. , Waller , D. W. and Carlson , R. E. 2002 . Multivariate analysis of the ecoregion delineation for aquatic systems . Lake Reserv. Manage. , 29 ( 1 ) : 67 – 75 .
- Jones , W. W. and Barnes , B. 2005 . Indiana lake water quality assessment report for 1994-1998 , Bloomington , Indiana : Indiana University, School of Public and Environmental Affairs .
- Jones , W. W. and Sauter , S. 2005 . Distribution and abundance of Cylindrospermopsis raciborkii in Indiana lakes and reservoirs , Bloomington , Indiana : Indiana University, School of Public and Environmental Affairs .
- Jones , W. W. and Medrano , S. 2006 . Indiana lake water quality assessment report for 1999–2003 , Bloomington , Indiana : Indiana University, School of Public and Environmental Affairs .
- Johnston , B. R. and Jacoby , J. M. 2003 . Cyanobacterial toxicity and migration in a mesotrophic lake in western Washington, USA . Hydrobiologia , 495 : 79 – 91 .
- Kelton , N. and Chow-Fraser , P. 2005 . A simplified assessment of factors controlling phosphorus loading from oxygenated sediments in a very shallow eutrophic lake . Lake Reserv. Manage. , 21 ( 3 ) : 223 – 230 .
- Knowles , R. , Lean , D. R.S. and Chan , Y. K. 1981 . Nitrous oxide concentrations in lakes: variations with depth and time . Limnol. Oceanogr. , 26 ( 5 ) : 855 – 866 .
- Knowlton , M. F. and Jones , J. R. 2006 . Natural variability in lakes and reservoirs should be recognized in setting nutrient criteria . Lake Reserv. Manage. , 22 ( 2 ) : 161 – 166 .
- Kratzer , C. R. and Brezonik , P. L. 1981 . A Carlson type trophic state index for nitrogen in Florida lakes . Water Res. Bull. , 17 ( 4 ) : 713 – 715 .
- Lampert , W. , Fleckner , W. , Rai , H. and Taylor , B. E. 1986 . Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase . Limnol. Oceanogr. , 31 ( 3 ) : 478 – 490 .
- Lijklema , L. 1994 . Nutrient dynamics in shallow lakes: Effects on changes in loading and role of sediment-water interactions . Netherlands: Hydrobiologia , 275 ( 1 ) : 335 – 348 .
- Nürnberg , G. K. 1987 . A comparison of internal phosphorus load in lakes with anoxic hypolimnia: Laboratory incubations versus in situ hypolimnetic phosphorus accumulation . Limnol. Oceanogr. , 32 ( 5 ) : 1160 – 1164 .
- Nürnberg , G. K. 1994 . Phosphorus release from anoxic sediments: What we know and how we can deal with it . Limnetica , 10 ( 1 ) : 1 – 4 .
- Nürnberg , G. K. and Peters , R. H. 1984 . The importance of internal phosphorus load to the eutrophication of lakes with anoxic hypolimnia . Verh. Internat. Verein. Limnol , 22 ( 1 ) : 190 – 194 .
- Omernik , J. M. 1987 . Ecoregions of the conterminous United States . Ann. Assoc. Amer. Geogr. , 77 ( 1 ) : 118 – 125 .
- Omernik , J. M. 1995 . Ecoregions: A framework for managing ecosystems . The George Wright Forum , 12 ( 1 ) : 35 – 50 .
- Popovičová , J. 2008 . Water quality monitoring and assessment of the Prairie Creek Reservoir in Delaware County, Indiana . Proceeding of the Indiana Academy of Science. , 117 ( 2 ) : 124 – 135 .
- Scheffer , M. 2004 . Ecology of shallow lakes , Netherlands : Kluwer Academic Publishers .
- Smith , R. A. , Alexander , R. B. and Schwartz , G. E. 2003 . Natural background concentrations of nutrients I streams and rivers of the conterminous United States . Environ. Sci. Technol. , 37 ( 14 ) : 3039 – 3047 .
- Stefan , H. G. , Hondzo , M. , Fang , X. , Eaton , J. G. and McCormick , J. H. 1996 . Simulated long-term temperature and dissolved oxygen characteristics of lakes in the north-central United States and associated with fish habitat limits . Limnol. Oceanogr. , 41 ( 5 ) : 1124 – 1135 .
- Stewart , W. D. , Preston , F. R.S. , Peterson , H. G. and Christofi , N. 1982 . Nitrogen cycling in eutrophic freshwater . Phil. Trans. R. Soc. Lond. , 296 : 491 – 509 .
- U.S. EPA . 1997 . “ Monitoring and assessing water quality ” . In Volunteer stream monitoring: a methods manual , U.S. Environmental Protection Agency .
- U.S. EPA . 2000 . Ambient water quality criteria recommendations: Information supporting the development of state and tribal nutrient criteria: Lakes and reservoirs in nutrient ecoregion VI. , U.S. Environmental Protection Agency . 822-B-00-008