Abstract
Collingsworth, P.D., R.A. Oster, C.W. Hickey, R.C. Heidinger and C.C. Kohler. 2009. Factors affecting water willow establishment in a large reservoir. Lake Reserv. Manage. 25:200–207.
We evaluated the transplanting potential of an emergent macrophyte, water willow (Justicia americana), into Rend Lake, a large, relatively unvegetated reservoir in southern Illinois. We transplanted water willow over two years into sites varying in physical conditions and using four propagule types. Transplanting in 2001 was performed to compare the durability of root crowns, plugs, and stem fragments, as well as to measure the effect of open water versus backwater sites on water willow survival. Water willow was transplanted in 2002 to explore how different planting protocols, such as planting in different months, planting different propagule types, and shoreline slope affect water willow survival. We found that water willow transplanted early in the summer had better post-winter colony survival and vigor (stem density within colonies) than transplants conducted later in the summer (Kruskal-Wallis test; P = 0.017). Rooted propagules had significantly greater first-year colony survival and ultimately exhibited greater vigor in newly established colonies. Lastly, we found that shoreline slope had a significant influence on the vigor of colonies following winter dormancy but was not significantly related to first-year colony vigor. In the summer following their initial transplanting, established water willow colonies planted on shorelines with steeper slopes (>5°) produced more plants than those planted on shallower shorelines (Kruskal-Wallis test; P = 0.003). Water willow seems to have high potential as a transplant species for reservoir management and riparian zone enhancement.
Aquatic macrophytes often play an important role in influencing species composition and angling quality of a reservoir fishery. Macrophyte density is related to predator efficiency because macrophyte stands serve as a juvenile fish refuge from predators (CitationSavino and Stein 1982, CitationGotceitas and Colgan 1987, CitationValley and Bremigan 2002). For example, CitationHayse and Wissing (1996) found high-density mats of artificial macrophytes attracted more age-0 bluegills (Lepomis macrochirus) than low-density mats, and that high-density mats reduced predation of age-0 bluegills by age-0 largemouth bass (Micropterus salmoides). Moderate densities of aquatic macrophytes seem to provide optimum foraging environments for many fish (CitationCrowder and Cooper 1982, CitationMiranda and Pugh 1997). Species such as largemouth bass utilize aquatic macrophytes to locate prey and to conceal themselves prior to ambush attacks. Larger sportfish of the family Centrarchidae are typically associated with aquatic macrophytes (CitationBain and Boltz 1992, CitationPaukert and Willis 2002).
Very large, newly created reservoirs are unique in that they are almost completely devoid of aquatic macrophytes. Factors that lead to paucity of macrophytes include fluctuations in water levels, often not coinciding with seasonal weather cycles; high inorganic turbidity, reducing the amount of light penetration; and bottom sediments of terrestrial origin (CitationSmart et al. 1996, CitationDick et al. 2004). Additionally, newly constructed reservoirs initially lack large numbers of native aquatic plant propagules to colonize the large area of immediately available habitat (CitationSmart et al. 2005). The establishment and presence of exotic plant species can be additional factors prohibiting native plant species colonization (CitationCox 1999). Consequently, an important management goal is often the establishment of appropriate vegetation.
When selecting a plant species to transplant into a reservoir, several criteria should be considered. First, transplanting native species is highly desirable because they tend not to spread to nuisance levels (CitationSmart et al. 1998). Second, species selected should produce hardy propagules appropriate for transplanting. Finally, species selected should be suited to the local environmental conditions. For example, when transplanting into a typical large Midwestern reservoir, plants selected should be resistant to high levels of turbidity, wave action, and large water level fluctuations characteristic of these systems (CitationDick et al. 2004).
American water willow (Justicia americana) possesses the traits of an appropriate transplant species and is currently being transplanted in Midwestern reservoirs (CitationStrakosh et al. 2005). Water willow is native to eastern North America and is typically found along the shorelines of lentic and lotic environments (CitationPenfound 1940). This emergent species is capable of vegetative reproduction via stem fragmentation, facilitating propagule production. Moreover, vegetative reproduction allows water willow to colonize new areas as the buoyant stems are carried away after fragmentation. Dense stands of water willow and their associated root systems are also positively correlated with substrate stability in streams (CitationFritz and Feminella 2003), which could provide an additional benefit of erosion protection for reservoirs receiving transplants (CitationRung et al. 2001).
Although water willow is currently used as a transplant species in several Midwestern states, little attention has been given to how different transplanting methods affect the survival of water willow colonies. We describe the results of two years of test plantings of water willow to assess its efficacy for use in a large reservoir nearly devoid of aquatic macrophytes. Our objectives were to (1) identify successful propagule types for transplantation and (2) identify site characteristics that maximize survival and colony expansion.
Methods
Study reservoir
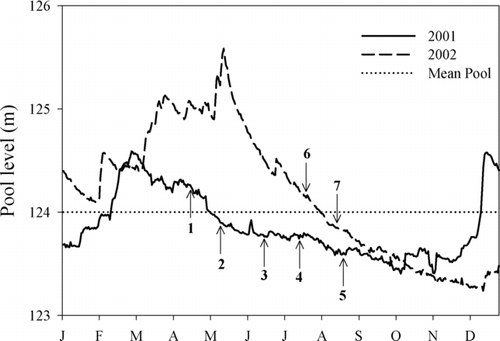
Rend Lake, located in Franklin and Jefferson Counties in southern Illinois, has a surface area of 7,649 ha with a maximum depth of 9.8 m. Normal mean pool elevation is 124 m above mean sea level. Several physical characteristics of Rend Lake make the reservoir inhospitable to establishment of native macrophytic vegetation. For example, large sporadic water-level fluctuations occur, inundating the shoreline in an unpredictable fashion (). Also, a substantial fetch generates scouring wave action along the exposed shorelines. These factors, combined with high turbidity (Secchi depths averaged 50.3 cm in 2001–2002), make establishment of aquatic macrophytes difficult. As such, submergent macrophytes are virtually nonexistent in Rend Lake, and emergent vegetation is sparse with patchy distributions of common reeds Phragmites austrailis and a few small water willow colonies along the shoreline. Although water willow colonies were of the same species transplanted during the project, the colonies were located in the back water areas that were not used during this project, and we were careful to select transplant sites that could not be naturally colonized by these established colonies.
Figure 1 Daily pool elevation in Rend Lake during 2001 and 2002. Numbers indicate when water willow transplants occurred during the study and the pool elevation at the time of transplants as follows: (1) April 2001 root crowns, (2) May 2001 plugs, (3) June 2001 stem fragments, (4) July 2001 stem fragments, (5) August 2001 stem fragments, (6) July 2002 stem fragments and rooted stem fragments, and (7) August 2002 stem fragments and rooted stem fragments. Full pool elevation of Rend Lake is 124 m above sea level, as indicated by the dotted line.
Transplant protocol
Four water willow propagules were developed during this project using various harvesting and holding techniques (). The simplest propagules were cuttings harvested from adult plants, hereafter referred to as stem fragments. Stem fragments (∼15–20 cm long) were cut to maximize the number of leaves and maintain photosynthetic activity. The second propagule was collected by uprooting root masses from dormant parent colonies. These propagules, hereafter referred to as root crowns, were harvested in the early spring prior to emergence of new plants. The third propagules, plugs, were cultivated from stem fragments collected in the fall, held over winter in an aquaculture pond, and grown outdoors for approximately one month in the spring prior to transplanting. The fourth propagules, rooted stem fragments, were cultured from stem fragments cut from parent plants in the spring and grown outdoors for one month prior to transplanting. This technique allowed the development of roots on the stem fragments but did not require the amount of time and effort necessary to produce plugs. All propagules were obtained from established water willow colonies in Kinkaid Lake, located <100 km away in Jackson County, Illinois. All stem fragment and root crown transplants were planted within 24 h of harvest.
Table 1 Experimental design of water willow transplants for transplant years 2001 and 2002. Due to annual differences in planting methods and reservoir hydrology, statistical comparisons were limited to transplants that occurred within a given year.
In April 2001, two colonies of root crowns were planted into each of five backwater sites. In May, two colonies of plugs were transplanted at three of the same backwater sites that received root crowns; however, an insufficient number of plugs were available to transplant colonies at the two remaining sites. Finally, stem fragment colonies were transplanted on a monthly basis from July through August into 10 sites in Rend Lake. Stem fragment colonies were evenly divided between the same five backwater sites and five additional open water sites to determine how site location affects water willow colony survival and vigor (). All backwater sites were located in no-wake areas naturally protected from direct wave action. All open-water sites were subject to waves created by wind or boat traffic. At each site, water willow colonies were transplanted using a 1.0-m2 grid (5 × 5) to plant 25 propagules evenly within the colony. All colonies were transplanted by placing the outer edge of the grid just within the water/shoreline interface at the present water level when plantings occurred. All transplantings during 2001 were designed to evaluate the first objective (identify successful propagule types for transplanting water willow into reservoirs).
Figure 2 Rend Lake sites that were planted with water willow propagules throughout the study. Triangles (▴) represent backwater sites planted with root crowns, plugs, and stem fragments in 2001. Diamonds represent backwater sites planted with only root crowns and stem fragments in 2001. Stars (⋆) represent open water sites planted with stem fragments only in 2001. Sites with letters represent 2002 plantings and planted with stem fragments and rooted stem fragments in July and August of 2002. Sites denoted by the letter “U” had a bottom slope of <5° whereas sites denoted with a letter “O” had a bottom slope of >5°.
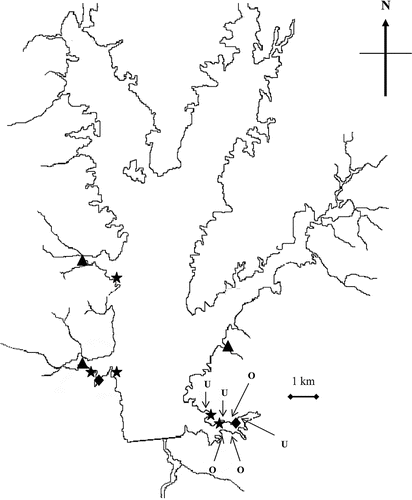
Transplanting during 2002 was conducted to evaluate site characteristics that influence colony survival and vigor. Three sites were selected with slopes <5° and three sites with slopes >5° at a distance of 1.5 m from the shoreline at the existing water level to determine how shoreline slope influences water willow transplant survival. Shoreline slope was included in 2002 transplants because native water willow colonies in Rend Lake and other southern Illinois reservoirs tend to be found along shorelines with slopes >5° (P.D. Collingsworth, Ohio University, unpublished data). In 2002, a large spring flood delayed the transplanting of water willow colonies until midsummer (). Plantings of water willow colonies were resumed in July and August at six sites located in a single backwater cove (). Three colonies of stem fragments and three colonies of rooted stem fragments were transplanted at each site in random order parallel to shoreline, for a total of six colonies per site. A 1.0-m2 quadrat (3 × 5) allowed 15 propagules per founding colony (45 stem fragments and 45 rooted stem fragments) per site. As with the previous year, all water willow colonies were transplanted by placing the outer edge of the grid just within the water/shoreline interface at the present water level on the date when plantings occurred.
Transplants conducted in 2001 and 2002 were monitored monthly through spring 2003 to determine colony survival and vigor. Colony survival was determined by counting the number of water willow colonies with at least one surviving plant on each monthly survival estimate, whereas colony vigor was determined by counting the number of individual water willow plants in a given colony on each sampling date. Colony survival and vigor was determined by dividing the number of colonies and plants at a given site on a given day by the initial number of colonies and propagules transplanted. Colony survival and vigor are reported as median percentages, which are the median values for each propagule and date combination. Therefore, colony vigor may exceed 100% as colonies grew new plants through the study. Due to annual differences in reservoir hydrology and transplanting methods, statistical tests were limited to comparison within transplanting years ().
Data analysis
Variation in colony vigor was analyzed using the nonparametric Kruskal-Wallis test. Two different estimates of colony vigor were used as dependent variables in the statistical models; first-year and post-winter colony vigor (final colony vigor estimate determined in early autumn and initial colony vigor estimate determined in the early summer the second year, respectively). For 2001 transplants, first-year and post-winter colony vigor was compared using propagule type as a main effect. Colony vigor of stem fragment transplants was also compared using site category (open water versus backwater sites) as a main effect. For 2002 transplants, first-year and post-winter colony vigor was compared using planting month, propagule type, and shoreline slope as main effects. When significant differences were found, Dunn's multiple range test was used to indicate where differences occurred (CitationDunn 1964, CitationZarr 1999). Additionally, second-year colony vigor was monitored monthly for surviving 2001 colonies through the following summer to record expansion of newly established colonies. Second-year vigor of 2001 transplants was analyzed using a repeated measures analysis of variance (ANOVA) and the profile transformation with propagule type as a main effect. The profile transformation describes differences in plant survival from adjacent months in a time series (CitationVonEnde 2001). All statistical analyses were performed using SAS software (CitationSAS 1999) at α= 0.05.
Results
2001 Transplants
In 2001, first-year vigor of water willow varied according to propagule type and planting month. As of 6 October 2001, nearly all (96%) water willow colonies planted into Rend Lake had surviving plants. Propagule type was significantly related to the vigor of water willow colonies (; Kruskal-Wallis test; P = 0.005). Rooted propagules (root crowns and plugs) maintained excellent survival of initial plants through the first year and showed some signs of modest colony growth. Root crowns had the highest colony vigor of all propagule types (median = 38.5 plants or 154%), but plug transplants also exhibited excellent colony vigor (median = 32 plants or 128%). Stem fragment colony vigor was inversely related to the amount of time the propagules spent in the reservoir. August transplants had the highest vigor of the monthly stem fragments colonies (median = 24.5 plants or 98%), while July stem fragment colonies were intermediate (median = 21.5 plants or 86%), and June stem fragments had the lowest colony vigor (median = 17 plants or 68%). Site location (open water versus backwater) was not related to water willow stem fragment first-year vigor (Kruskal-Wallis test; P = 0.546). As of October 2001, 93% of stem fragment colonies in open water and backwater sites survived. The median first-year colony vigor of stem fragments in open water and backwater sites was 80% and 92%, respectively.
Post-winter and second-year vigor was variable for the 2001 water willow colonies, following a distinct pattern relative to time of planting. In all, 71% percent of water willow colonies transplanted in 2001 survived the initial winter in the reservoir. Propagule type was again an important factor describing transplant post-winter colony vigor (; Kruskal-Wallis test; P = 0.017). Post-winter survival was excellent for root crowns and plugs, with all root crown colonies and 5 of 6 (83%) plug colonies showing signs of survival by June 2002. Post-winter survival for stem fragment colonies was moderate, with plants at 19 of 40 sites (48%) successfully surviving the first winter. Stem fragment colony vigor was related to planting month, but unlike first-year survival, post-winter colony vigor was greater for colonies transplanted early in the year than colonies planted later. Water willow colony vigor was greatest for June stem fragment colonies, intermediate for July stem fragment colonies, and lowest for August stem fragment colonies. Site location was statistically unrelated to post-winter vigor of stem fragment colonies. Again, no differences were found in vigor between open water and back water stem fragment colonies in the initial 2002 survival estimate (Kruskal-Wallis test; P = 0.460); however, post-winter colony survival was poor for open water sites, with plants in 5 of 15 sites (33%) surviving the initial winter with a median colony vigor of 13%. In backwater sites, 12 of 15 colonies (80%) survived their initial winter in the reservoir, and colony vigor was higher in backwater colonies (median colony vigor = 14 plants or 56%).
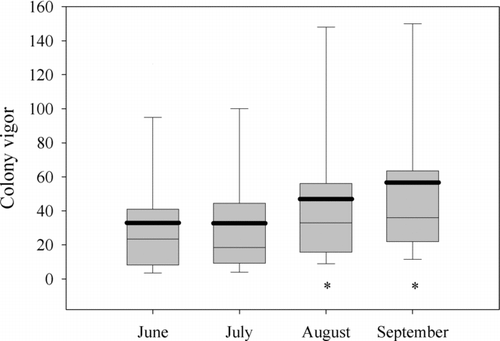
Newly established water willow colonies expanded in number of plants throughout the second summer in Rend Lake (). Propagule type had a significant effect on colony vigor throughout the second summer (Repeated Measures ANOVA, F = 4.60, df = 4, P = 0.005). From June through August, established colonies of root crowns maintained higher colony vigor than all other propagule types. New colonies transplanted with plugs and June stem fragments had the next highest colony vigor, while water willow colonies transplanted with stem fragments in July and August had the lowest colony vigor. By September 2002, no significant differences in colony vigor by propagule type were evident. Significant growth in water willow colony vigor through time was illustrated by a significant time effect (F = 19.03, df = 3, G-G adjusted P < 0.0001). However, no significant propagule × time interaction was found for colony vigor (F = 0.77, df = 12, G-G adjusted P = 0.6324), indicating that all water willow colonies from propagule types grew in a similar manner through time. Lastly, water willow colony vigor was significantly different in adjacent months, but these differences were unrelated to propagule type. Significant growth in colony vigor occurred later in the summer for all transplants, regardless of propagule type. No significant changes in colony vigor occurred between June and July (F = 0.34, df = 1, P = 0.5622), but significant increases in colony vigor occurred between July and August (F = 27.03, df = 1, P < 0.0001) and between August and September (F = 6.82, df = 1, P = 0.0141; ).
Figure 3 Colony expansion associated with different water willow propagule types planted during the summer of 2001 and monitored through the summer of 2002. Error bars represent ±1 S.D.
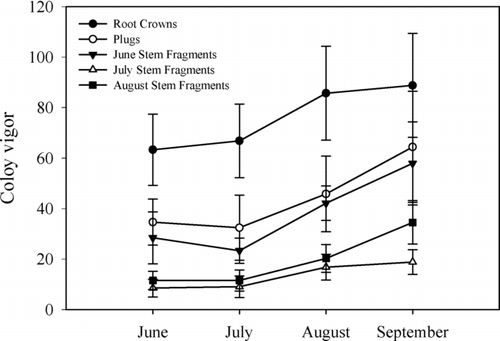
Figure 4 Water willow colony expansion through the summer of 2002. Solid lines represent quartiles and bold lines represent mean survival estimates. Monthly survival means with an asterisk were significantly greater than the means from the previous month (Repeated Measures ANOVA and Profile Transformation, P < 0.05).
2002 Transplants
First year survival of water willow colonies was excellent in 2002, with all 72 transplants surviving through 25 September. Planting month had a significant effect on the colony vigor of water willow through the first growing season (; Kruskal-Wallis test; P = 0.006). Water willow transplanted in July had 100% median colony vigor after six weeks while colonies transplants conducted in August had 87% vigor. Propagule type was also an important factor on first-year water willow vigor (Kruskal-Wallis test; P < 0.0001). Rooted stem fragments colonies survived well throughout the initial summer, with 100% median colony vigor as of September 2002. Stem fragments also experienced good first-year survival through the summer, with a median 83% colony vigor. Lastly, we found that shoreline slope was not significantly related to first-year vigor of water willow colonies (Kruskal-Wallis test; P = 0.1241). Median vigor was 87% for water willow colonies transplanted on shorelines with slopes >5°, and 93% for those on shorelines with slopes <5°.
Table 2 Median percentage of surviving plants for different water willow propagule types planted in 2001 at the end of the first summer and following winter dormancy. Survival was compared using the nonparametric Kruskal-Wallis test using the final survival estimate of the first summer (October 2001) and the initial survival estimate the next year (June 2002). Medians with different letters are significantly different (Dunn's multiple comparison test, P < 0.05).
Post-winter survival of water willow colonies transplanted summer 2002 was variable, with different factors of importance for post-winter than first-year survival (see ). Overall, 54 of the original 72 (75%) water willow colonies survived the initial winter in Rend Lake, with a median colony vigor of 93%. Planting month had a significant effect on post-winter colony vigor (Kruskal-Wallis test; P = 0.017). Median post-winter vigor was higher for water willow colonies transplanted in July (110%) than those transplanted in August (40%). Contrary to the results of first-year survival estimates, propagule type was not significantly related to post-winter colony vigor of water willow in Rend Lake (Kruskal-Wallis test; P = 0.1726). Post-winter colony survival was excellent for both propagule types, with 29 of 36 (81%) rooted stem fragment and 25 of 36 (69%) of stem fragment colonies surviving the initial winter. Median post-winter colony vigor was similar for rooted stem fragments (73%) and regular stem fragments (47%). Finally, shoreline slope was significantly related to post-winter vigor of water willow colonies (Kruskal-Wallis test; P = 0.003). Although post-winter colony survival was similar for shorelines with steep and shallow slopes (81% survival on shorelines >5° and 69% on shorelines <5°), colony vigor varied by shoreline slope. Median post-winter vigor for water willow colonies transplanted on shorelines with slopes >5° was 120%, and 40% plants for those transplanted on shorelines wit slopes <5°.
Table 3 Median percentage of surviving plants for different treatments planted in 2002 at the end of the first summer and following winter dormancy. Survival for treatment groups was compared using the nonparametric Kruskal-Wallis test using the final survival estimate (September 2002) and the initial survival estimate the next year (June 2002). Medians with different letters are significantly different (Dunn's multiple comparison test, P < 0.05).
Discussion
Propagule type was a significant factor in first-year colony vigor of water willow because rooted propagules tended to have better survival than stem fragments. Smart et al. (Citation1996, Citation1998) suggest submersed vegetation be grown in nursery pots prior to transplanting, which is consistent with our results and highlights the importance of selecting appropriate propagules to maximize survival of transplants.
From both a cost and effort prospective, the simplest propagules used were stem fragments, which required a minimal amount of effort to harvest and replant. Because these propagules lack roots, transplants have longer lag times from initial plantings to formation of established colonies. Although root crowns and plugs had better first-year survival in 2001 plantings, winter survival of June stem fragments was similar to plug transplants. Also, following winter dormancy we found no significant propagule type by time interaction in colony growth, suggesting that colonies expand through time similarly, regardless of propagule type planted. In 2002, we also found rooted stem fragments had better first-year survival than stem fragments, though winter survival was unrelated to propagule type.
Root crown transplants, which yielded more plants than any other propagule type used in our study, maintained excellent survival throughout the initial summer and expanded the following summer. However, root crowns must be harvested in the dormant state and thus are only available prior to the late spring emergence of water willow. This period usually corresponds with maximum spring flood levels in reservoirs (), making harvesting and transplanting root crowns difficult. Unlike stem fragments that can produce multiple propagules from a single host, harvesting root crowns requires the sacrifice of an entire host plant. Therefore, harvesting root crowns on a large scale could adversely affect source populations.
The results of this and a similar study (CitationDoyle et al. 1997) suggest that transplanting cultured plants provide better results than transplanting wild ones. We found planting rooted water willow, including plugs in 2001 and rooted stem fragments in 2002, resulted in better survival through the first year, and these propagules maintained excellent growth through the second year. While developing a system to culture water willow propagules requires more initial investment, large-scale and long-term reservoir enhancement projects would benefit from using cultured propagules by ensuring a dependable source of quality propagules and minimizing impacts on native water willow sources.
Perhaps the single most important factor in establishing water willow was planting time. We found that, in general, early planting resulted in increased survival and better colony expansion than later planting, probably because they have adequate time to develop roots prior to winter dormancy. Root development is crucial for energy storage during the winter and colony expansion via rhizomes the following year. From our transplants, we found that root crowns and plugs had the greatest first-year survival, winter survival, and colony expansion for the 2001 transplants. One potentially confounding factor with planting time is reservoir hydorology. During the two years of this study, water levels in Rend Lake were highest during the spring, followed by a continuous draw down throughout the summer and early fall, a common hydrological pattern for Midwestern reservoirs. Due to this precipitation pattern, water willow colonies transplanted early were planted further up shoreline than colonies transplanted later in the summer. Therefore, water willow colonies transplanted early in the spring have a higher likelihood of desiccation during the initial summer in the reservoir. In contrast, water willow colonies transplanted later in the summer were more likely to become inundated the following year as water levels rose during the spring. However, CitationStrakosh et al. (2005) found that water willow were more tolerant of prolonged periods of dessication than inundation, further strengthening the case for early planting of water willow propagules.
We also found that site characteristics, such as the location and the slope of the shoreline of the transplant sites affected water willow survival and colony vigor. Stem fragment colony vigor was not statistically different between open water and backwater sites in our first year and post-winter estimates, probably due to low sample sizes of these transplants. Colonies transplanted into open water sites had very poor post-winter survival during this study with 5 of 15 (33%) colonies surviving the initial winter compared to 12 of 15 colonies transplanted in backwater sites. These findings are consistent with CitationSmart and Dick (1999), who advocate using a “founder colony” approach by planting propagules into protected areas that, through time, will expand into different habitats. Finally, we found that shoreline slope was not related to first-year survival of transplants, but was significantly related to winter survival for 2002 transplants. The importance of shoreline slope is probably because shorelines with steeper slopes are less prone to dramatic water level fluctuations.
Water willow seems to have high potential as a transplant species for reservoir enhancement; however, in our study, we monitored new water willow colony survival in a single system using small-scale plantings. Enhancement programs at the scale necessary to provide noticeable improvements to the fisheries of large reservoirs will be conducted in multiple systems requiring a much larger number of plants than we considered; therefore, additional research is needed to identify the optimal number of propagules needed for different transplant scenarios. Future studies should investigate how the number of propagules influences the likelihood of colony establishment using a variety of propagule types and in a variety of systems.
Acknowledgments
The authors would like to thank Emily Wonnacott, Cliff O'Neal, and Brian O'Neil for their assistance in data collection and analysis. Tim Spier and Rob Colombo provided useful comments on earlier drafts of this manuscript. This research was sponsored by the Fisheries Division of the Illinois Department of Natural Resources, funded through the Federal Aid in Sportfish Restoration Act (F-140-R).
References
- Bain , M. B. and Boltz , S. E. 1992 . Effect of aquatic plant control on the microdistribution and population characteristics of largemouth bass . Trans. Am. Fish. Soc. , 121 : 94 – 103 .
- Cox , G. W. 1999 . Alien species in North America and Hawaii: impacts on natural ecosystems , Washington , D.C. : Island Press .
- Crowder , L. B. and Cooper , W. E. 1982 . Habitat structural complexity and the interaction between bluegills and their prey . Ecology , 63 : 1802 – 1813 .
- Dick , G. O. , R. M. and Smith , J. K. 2004 . Aquatic vegetation restoration in Cooper Lake, Texas: a case study , Vicksburg , MS : U.S. Army Corp of Engineers Waterways Experiment Station . Report ERDC/EL TR-05-5
- Doyle , R. D. , Smart , R. M. , Guest , C. and Bickel , K. 1997 . Establishment of native aquatic plant for fish habitat: test plantings in two north Texas reservoirs . Lake Reserv. Manage. , 13 : 259 – 269 .
- Dunn , O. J. 1964 . Multiple contrasts using rank sums . Technometrics , 6 : 241 – 252 .
- Fritz , K. M. and Feminella , J. W. 2003 . Substratum stability associated with the riverine macrophyte Justicia americana . Freshw. Biol. , 48 : 1630 – 1639 .
- Gotceitas , V. and Colgan , P. 1987 . Selection between densities of artificial vegetation by young bluegills avoiding predation . Trans. Am. Fish. Soc. , 116 : 40 – 49 .
- Hayse , J. W. and Wissing , T. E. 1996 . Effects of stem density of artificial vegetation on abundance and growth of age-0 bluegills and predation by largemouth bass . Trans. Am. Fish. Soc. , 125 : 422 – 433 .
- Miranda , L. E. and Pugh , L. L. 1997 . Relationship between vegetation coverage and abundance, size, and diet of juvenile largemouth bass during winter . N. Am. J. Fish. Manage. , 17 : 601 – 610 .
- Paukert , C. P. and Willis , D. W. 2002 . Seasonal and diel habitat selection by bluegills in a shallow natural lake . Trans. Am. Fish. Soc. , 131 : 1131 – 1139 .
- Penfound , W. T. 1940 . The biology of Dianthera americana L . Am. Midland Nat. , 24 : 242 – 247 .
- Rung , R. C. , Lutterbie , G. and Pescitelli , S. M. 2001 . An evaluation of selected emergent plant species, their potential for aquatic habitat enhancement , Illinois Department of Natural Resources Technical Report .
- SAS . 1999 . SAS/SAST User guide , Cary , NC : Version 8, SAS Institute Inc. .
- Savino , J. F. and Stein , R. A. 1982 . Predator-prey interactions between largemouth bass and bluegills as influenced by simulated submersed vegetation . Trans. Am. Fish. Soc. , 111 : 255 – 266 .
- Smart , R. M. , Doyle , R. D. and Madsen , J. D. 1996 . Establishing native submersed aquatic plant communities for fish habitat . American Fisheries Society Symposium , 16 : 347 – 356 .
- Smart , R. M. , Dick , G. O. and Doyle , R. D. 1998 . Techniques for establishing native aquatic plants . J. Aquat. Plant Manage. , 36 : 44 – 49 .
- Smart , R. M. and Dick , G. O. 1999 . Propagation and estabilishment of aquatic plants: a handbook for ecosystem restoration projects , Vicksburg , MI : U.S. Army Corps of Engineers, Waterways Experiment Station . Report A-99-4
- Smart , R. M. , Dick , G. O. and Snow , J. R. 2005 . Update to the propogation and establishment of aquatic plants handbook , Vicksburg , MS : U.S. Army Corp of Engineers Waterways Experiment Station . Report ERDC/EL TR-05-4
- Strakosh , T. R. , Gido , K. B. , Guy , C. S. and Eitzman , J. L. 2005 . The response of water willow Justicia americana to different water inundation and dessication regimes . N. Am. J. Fish. Manage. , 25 : 1476 – 1485 .
- Valley , R. D. and Bremigan , M. T. 2002 . Effects of macrophyte bed architecture on largemouth bass foraging: implications of exotic macrophyte invasions . Trans. Am. Fish. Soc. , 131 : 234 – 244 .
- VonEnde , C. N. 2001 . “ Repeated-measures analysis ” . In Design and analysis of ecological experiments , 2nd , Edited by: Scheiner , S. M. and Gurevitch , J. 134 – 157 . NY : Oxford University Press .
- Zarr , J. H. 1999 . Biostatistical analysis , 4th , NJ : Prentice Hall, Upper Saddle River .