Abstract
Nishihiro, J. and I. Washitani. 2009. Quantitative evaluation of water-level effects on “regeneration safe-sites” for lakeshore plants in Lake Kasumigaura, Japan. Lake Reserv. Manage. 25:217–223.
We evaluated the effects of water-level regime on the area of “regeneration safe-sites” for seed germination and initial establishment of seedlings of helophytes occurring in the emergent vegetation zone of Lake Kasumigaura, Japan. Based on the results of previous studies and an original experiment, potential regeneration safe-sites (PRS) for helophytes were defined as sites in which the ground surface was inundated for less than 3 consecutive days from 1 April to 15 May. We estimated and compared the areas of PRS both before and after the implementation of an artificial water-level regime in the mid-1970s. Comparisons were made using data on daily water levels during 1960–1969 and 1996–2005, topographic profiles of the lakeshore in 1967 and 2005, and areas of the emergent vegetation in 1972 and 1997. We estimated that the current area of PRS in the lake has been reduced to 24% of past levels. This reduction was accompanied by both a complete loss of a spring drawdown and a decrease in the area of emergent vegetation. The areas of PRS under various water management regimes indicated that a relatively slight lowering of the spring water level could cause a marked increase in the area of PRS (e.g., a 10-cm decrease in the managed target water level could result in a doubling of the area of PRS). Our results suggest a water-level management policy that includes a spring drawdown can be effective in the restoration of helophyte species diversity through the recovery of PRS.
Lakeshore vegetation plays a vital role in the maintenance of biodiversity and ecosystem functioning in lake ecosystems; however, the vegetation of many lakes has been reduced by various human activities, including the construction of artificial levees on lakeshores (CitationElias and Meyer 2003) and the deterioration of water quality (CitationBrinson and Malvarez 2002, CitationSchmieder 2004, CitationMaemets and Freiberg 2005). In addition to these well-recognized impacts, manipulations of the water level of a lake and/or its patterns of fluctuation are other important factors affecting the abundance and species composition of lakeshore vegetation (CitationCrivelli and Grillas 1995, CitationShay et al. 1999, CitationCoops and Hosper 2002).
Lake water levels are often strongly affected by human demands through management related to water usage and/or flood control, and any decisions regarding water-level management require a consensus among stakeholders (CitationSteinman et al. 2002). During such consensus-building processes, it is critical to predict the effects of various water-level management options on both lakeshore vegetation, which is crucial to lake ecosystem integrity, and other important ecological and social issues under consideration.
Although the effects of water-level regime on lakeshore plants can vary among species and among growth stages within a species (CitationNilsson and Keddy 1988, CitationHill et al. 1998), research should focus on the groups of species most important to biodiversity and ecosystem functioning and on the life-stages that strongly affect the viability of the plant population. Marsh plants growing in the lakeshore emergent vegetation are an important component of the plant diversity of a lake; furthermore, these species play important roles in lake ecosystems, such as forming the basis of the littoral food web (CitationWetzel 2001) and providing food and habitat for many animal species (CitationSchmieder 2004). Marsh plants are highly sensitive to changes in water regime: overly high water levels have caused declines in or the extermination of their populations (Citationvan der Valk 1981, CitationGalinato and van der Valk 1986, CitationSchneider 1994, CitationLenssen et al. 1998, CitationBaldwin et al. 2001, CitationBudelsky and Galatowitsch 2004). Furthermore, germination and seedling establishment stages of marsh plants can be severely inhibited by inundation (CitationGalinato and van der Valk 1986, CitationBaldwin et al. 1996, CitationLenssen et al. 1998, CitationSeabloom et al. 1998, CitationNishihiro et al. 2004a). Inhibition of regeneration by seeds can lead not only to the immediate disappearance of above-ground populations of annual plants, but also to decreases in the size and genetic diversity of perennial plant populations (CitationWatkinson and Powell 1993). A quantitative evaluation of the effects of water-level policy changes on the temporal and spatial availability of “safe-sites” for seed germination and seedling establishment of lakeshore marsh plants is therefore essential for the planning of appropriate management strategies to preserve biodiversity and the integrity of ecosystem functioning.
In Lake Kasumigaura, the second largest lake in Japan, the species richness of lakeshore vegetation has been rapidly declining since the implementation of a new water-level policy in the mid-1970s, which stabilized the water level throughout the year (CitationMiyawaki et al. 2004, CitationNishihiro and Washitani 2007). Our previous studies in the remnant lakeshore reed-marsh suggest that although species rich soil seed banks are still retaining in the soil, the opportunity for seed germination and/or seedling establishment is largely reduced by the inundation (CitationNishihiro et al. 2004a, Citation2004b). The reduction in regeneration safe-sites is suspected as a major cause of the recent decline of the species diversity.
In this study, we evaluated and compared the areas of potential regeneration safe-sites (PRS) of lakeshore marsh plants in Lake Kasumigaura before and after the implementation of the new policy. We also predicted the area of PRS under various potential water regimes. Based on previous studies of the environmental requirements for seed germination and seedling establishment of plants growing in lakeshore reed-marsh (CitationNishihiro et al. 2004a, Citation2004b), PRS were defined as areas of emergent vegetation that escaped severe inundation and were estimated using lake water levels and lakeshore topography. In addition, the validity of these criteria was verified in a small-scale lakeshore experiment.
Methods
Study site
Lake Kasumigaura (also known as Nishiura; 35° N, 140° E) is a shallow freshwater lake with a surface area of 17,200 ha and average and maximum water depths of 4 and 7 m, respectively. The annual mean air temperature and annual precipitation recorded at the nearest meteorological station (Tsuchiura Meteorological Station) are 14.7° C and 1153.6 mm, respectively (1996–2005 average). The construction of concrete levees over the vegetation along all the lakeshore and deterioration of water quality have heavily damaged the littoral vegetation. Although the natural littoral vegetation once consisted of at least 423 ha of emergent and 748 ha of submerged vegetation as late as 1972, only a fragmented belt of emergent vegetation (183 ha in total), dominated by Phragmites australis W. Clayton, remained between open water and the concrete levees by the 1990s, and submerged plants had completely disappeared (CitationSakurai 1981, CitationNishihiro and Washitani 2007). The loss of submerged vegetation might accelerate the area decrease of the emergent vegetation. The loss of plant species diversity in the emergent vegetation is also recognized; at least 15 species recorded in 1970s were not observed in 1990s or later (Nishihiro [forthcoming 2009]).
After the 1975 implementation of intense artificial water-level control through the operation of a water gate in a downstream river of the lake, the natural, seasonal water-level fluctuations (i.e., a drawdown in winter–spring and an increase in summer–autumn) have been almost completely lost. Details of the patterns of water-level fluctuations before and after the implementation of the water management regime are described in the Results section.
Evaluation of the area of potential regeneration safe-sites
Previous studies suggest that the phenology of seedling emergence and the effects of inundation on the mortality of emerged seedlings are similar for a large number of plants occurring in the reed-marsh shore of Lake Kasumigaura. For example, seedling emergence in at least 42 species, including 23 annual and 19 perennial, was concentrated from April to early May (CitationNishihiro et al. 2004a), and a significant reduction in seedling emergence occurred in 19 of 24 species under experimental inundated conditions (CitationNishihiro et al. 2004b). Furthermore, our observational study revealed that the naturally emerged seedlings of marsh plants suffered high mortality when the ground surface was inundated continuously for more than 3 days (CitationNishihiro et al. 2004a). The PRS of the lakeshore marsh plants were quantified base on the assumptions that PRS occur in the existing emergent vegetations, and that the PRS area is the range of ground surface inundated for less than 3 consecutive days during the period of seedling establishment (i.e., 1 April to 15 May). We calculated the area of the PRS using data on areas of emergent vegetation, topographic profiles of the emergent vegetation, and daily water levels of the lake.
The area of emergent vegetation was quantified from vegetation maps in 1972 and 1997, which represent past and current periods, respectively (). The proportion of ground surface in PRS condition was calculated as the number of ground points that satisfied the topographic criteria of PRS (described above) divided by the total number of points for which ground height was measured. We used topographic profiles of 32 transects, which were set perpendicular to the shoreline and across the beds of lakeshore vegetation for the past and current periods. Measuring the ground height at 0.5-m intervals along the transects yielded 2539 and 2313 points of ground-height data for the past and current period, respectively (). The inundation of a ground point was determined by comparing the ground height of the point and the daily water level recorded once a day at the center of the lake by the Ministry of Land, Infrastructure, and Transport of Japan (MLIT; ).
Table 1 Sources of data for the estimation of the areas of potential regeneration safe-sites under past, current, and potential future lake water management. MLIT: data obtained by the Ministry of Land, Infrastructure, and Transport of Japan.
Areas of PRS under given water regimes were calculated using area and topographic data of the current emergent vegetation and the potential lake water level, which was simulated using the daily water level during the current regime (). Under this regime, the lake water level was approximately maintained at a target water level throughout the year. Although rainfall caused daily fluctuations in the water level, monthly averages nearly coincided with the target water level. In our calculations, the daily water level for each year during 1996–2005 (with the exception of 1999, an exceptionally high water-level year) was adjusted so that the monthly average corresponded to a certain target water level.
Effect of inundation of seedling mortality
To validate the above-mentioned criteria of PRS, i.e., sites inundated for less than 3 consecutive days during the period from 1 April to 15 May, we tested the effects of natural inundation on the seedling mortality of 12 indigenous plants common to the remnant reed beds of Lake Kasumigaura (), including Carex capricornis, which is endangered in Japan (Environmental Agency of Japan 2000). We collected seeds of each species in 2001 from a remnant reed bed, and in early May 2002 we sowed 10 replicates of 50 seeds, moist-chilled (i.e., stratification) for 8 weeks, of each species in plastic pots (25 cm diameter × 10 cm height) filled with sand. The pots were placed in plastic containers, and the water level was maintained at 5 cm below the sand surface to maintain field capacity. Seedlings were grown until the cotyledons had fully expanded and were thinned to 25 per pot in early June. On 15 June 2002, five pots of each species were placed along the shoreline of Lake Kasumigaura in a restored vegetation patch (36°3′ N, 140°13′ E) sheltered by stone wave protection where the sand surface was under 10 cm of water (inundated treatment). The other five pots per species were placed along the shore where the sand surface was 10 cm above the water (non-inundated treatment). The number of live seedlings was recorded daily for both the inundated and non-inundated treatments until ⩾ 95% of seedlings had died. Seedlings that had wilted or floated away were considered dead.
Table 2 Survival rate of 12 helophytic species after exposure to the inundation treatment. All seedlings in the non-inundated treatment survived during the same period.
Results
Lakeshore topography and lake water level
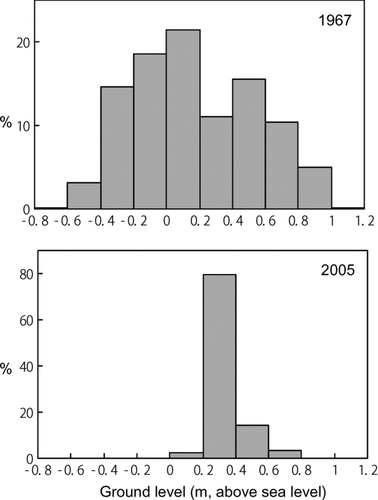
Although the locations of transects were identical in 1967 and 2005, the topographic profiles demarcating zones of emergent vegetation were markedly different: variation in ground height drastically declined in 2005 (). In particular, emergent vegetation with ground heights lower than 0.2 m a.s.l and those higher than 0.4 m a.s.l. suffered severe losses ().
Figure 1 Relative frequency of the ground heights of emergent vegetation in 1967 (mean ± SD: 0.176 ± 0.366, n = 2539) and 2005 (0.324 ± 0.104, n = 2313).
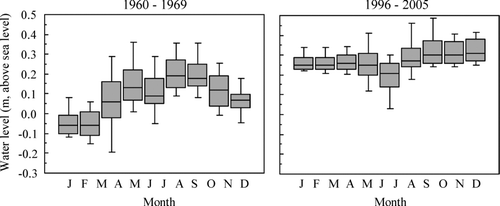
Prior to the implementation of the new water-level policy (1960–1969), there were distinct seasonal fluctuations in water level, with a drawdown in spring and an increase in autumn (). During this period, the daily average lake water level in spring (1 April to 15 May) ranged from −0.18 to 0.23 m, with an average of−0.05 m a.s.l. During more recent years (1996–2005), however, water levels ranged from 0.13 to 0.63 m, with a mean of 0.26 m a.s.l., and were significantly higher than in the earlier period.
Figure 2 Seasonal water-level patterns in Lake Kasumigaura, Japan. The monthly average, 25th–75th percentiles, and 10th–90th percentiles are shown prior to water-level management using a water gate (1960–1969, left graph) and for more recent years (1996–2005, right graph) subsequent to the implementation of the management regime.
Area of potential regeneration safe-sites
The area of emergent vegetation in 1997 (183 ha) was only 43.3% of that in 1972 (423 ha). Although there was some among-year variation from 1960 to 1969, 56.8 ± 7.2% (mean ± SD; n = 10) of the area of emergent vegetation met the PRS conditions; however, from 1996 to 2005 only 32.0 ± 16.9% (n = 10) of the area of emergent vegetation was considered PRS. The current total area of PRS was significantly lower than that in the past, although the area of estimated PRS depended on the various criteria used to define these sites ().
Table 3 Estimated area of potential regeneration safe-sites (PRS) in the past and current lake conditions under different criteria: the ground surface is inundated for less than d consecutive days during the period from 1 April to 15 May (d= 1, 2, …7). Means and SDs were shown with the results of the 1 way ANOVAs.
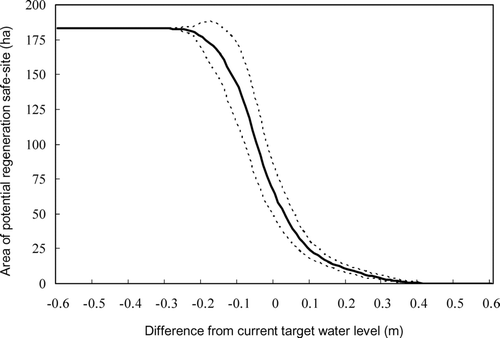
Variation in the area of predicted PRS under various water-level management regimes indicated that a slight change in the target water level from its current value could cause a marked change in the area of PRS. For example, a 10-cm increase in the target water level could cause a 37% reduction in the area of PRS, whereas a 10-cm decrease could double the area of PRS ().
Figure 3 Shifts in the estimated area of potential regeneration safe-sites for lakeshore helophytes due to changes in the target water level used for lake water management. Solid and dotted lines represent the mean (n = 9) and 95% confidence intervals, respectively.
Effect of inundation on seedling mortality
When inundated, all seedlings except those of Eleocharis wichurae died within 6 days (). In contrast, all seedlings in the non-inundated treatment survived during the same period. Regardless of species, more than 30% of seedlings died within 3 days of being inundated ().
Discussion
Tolerance of seedlings to inundation
Many plant species have evolved physiological mechanisms to enable their seeds to detect the appropriate timing and/or micro-environmental conditions for germination (CitationHarper 1977). However, various unpredictable biotic and abiotic factors affect successful seedling establishment under natural conditions (CitationSilvertown and Dickie 1981). Although many empirical studies have suggested that marsh plants are adapted to germinate under non-inundated conditions during the germination season (CitationGalinato and van der Valk 1986, CitationBaskin et al. 1993, CitationBaldwin et al. 1996, Citation2001, CitationLenssen et al. 1998, CitationSeabloom et al. 1998, CitationNishihiro et al. 2004a, Citation2004b), little is known about the tolerance of emerged seedlings to inundation (but see CitationStreng et al. 1989). During only 3 days of inundation, we observed high mortality of the emerged seedlings of 11 of 12 species, suggesting that our definition of PRS was valid. The effects of inundation on seedling mortality under natural lakeshore conditions, however, may be even more severe than those under experimental conditions that offered protection from wave action, which along with moving debris in the water can exacerbate seedling mortality (CitationStreng et al. 1989). Our estimated area of PRS may therefore be somewhat larger than that occuring in nature.
Some E. wichurae seedlings survived during the inundation treatment; however, both seed germination and seedling emergence of this species are significantly reduced by inundation (CitationNishihiro et al. 2004b). Thus, inundation can also negatively affect the regeneration of E. wichurae.
Effects of water level on regeneration safe-sites
Our results suggest that during the last ∼30 years the area of PRS has diminished to 24% of its historic level. The primary causes of this decline include both the implementation of a new water-level management policy resulting in high water during spring and decreases in the area of lakeshore emergent vegetation due to the alteration of shore topography through the construction of concrete levees and/or shore erosion. The seasonal water-level fluctuations that occurred up until the implementation of the new policy in 1975 were largely influenced by the seasonality of precipitation in the watershed; monsoons cause high amounts of precipitation during summer and autumn, whereas precipitation is relatively light in the winter and early spring. The spring drawdown of lake water is crucial for generating seasonal germination safe-sites for most lakeshore plants that germinate in spring on exposed ground during periods of low water (CitationNishihiro et al. 2004a, Citation2004b).
Low variation in ground height along the current lakeshore may be a result of stabilization of the lake-water level. In particular, the loss of ground with height lower than 0.2 m () can be explained as a result of erosion during a period when the water level had been maintained at ∼ 0.25 m a.s.l. (). Conversely, the loss of the ground higher than 0.4 m is thought to be a result of the construction of the concrete levees on the emergent vegetation during 1970–1996 (CitationMiyawaki et al. 2004).
Our evaluation showed that the area of PRS can be largely changed even if the change in the target water level is small () because the topographic features of lakeshore emergent vegetation exhibit low variation in ground height (). A rising spring water level may disproportionately reduce the number of opportunities for regeneration of lakeshore plants. In contrast, the area of PRS could be doubled by lowering the target water level by only 10 cm; therefore, decreasing the target water level by 10–20 cm would be an effective method of conserving the diversity of lakeshore plants. The period needed to allow seedlings to grow to a flood-tolerant height is currently unclear. The optimal period of spring drawdown for diversity conservation should be carefully monitored through adaptive management.
Although the improvement of water-level regime can contribute to the recovery of plant diversity, it is not sufficient to achieve the long-term recovery of the degraded vegetation of the lake. Other efforts such as the area recovery of the vegetation zone are indispensable to the restoration of lakeshore vegetation. In 2002, the reconstruction of an artificial littoral zone to restore shoreline vegetation was implemented at Lake Kasumigaura (CitationNishihiro et al. 2006), and monitoring of the restored vegetation has continued (CitationNishihiro and Washitani 2007). Such restoration efforts can enhance the effects of the water-level improvement.
Acknowledgments
We thank The Kasumigaura River Office of Ministry of Land, Infrastructure, and Transport, Japan for providing the data of the lake-water level and lakeshore topography. We are grateful to Naomi Hamada for her assistance with collection of topographic data. This research was partly financed by Grant-in-Aid for Young Scientists (B) 19710198 to J. Nishihiro.
Notes
*The locations of transects and surveyed topographic profiles were identical in 1967 and 2005.
*P < 0.0001.
References
- Baldwin , A. H. , McKee , K. L. and Mendelssohn , I. A. 1996 . The influence of vegetation, salinity, and inundation on seed banks of oligohaline coastal marshes . Am. J. Bot. , 83 : 470 – 479 .
- Baldwin , A. H. , Egnotovich , M. S. and Clarke , E. 2001 . Hydrologic change and vegetation of tidal freshwater marshes: field, greenhouse, and seed-bank experiments . Wetlands , 21 : 519 – 531 .
- Baskin , C. C. , Baskin , J. M. and Chester , E. W. 1993 . Germination ecology of Lepiochloa panicoides, a summer annual grass of seasonally dewatered mudflats . Acta Oecol. , 14 : 693 – 704 .
- Brinson , M. M. and Malvarez , A. I. 2002 . Temperate freshwater wetlands: types, status, and threats . Environ. Conserv. , 29 : 115 – 133 .
- Budelsky , R. A. and Galatowitsch , S. M. 2004 . Establishment of Carex stricta Lam. seedlings in experimental wetlands with implications for restoration . Plant Ecol. , 175 : 91 – 105 .
- Coops , H. and Hosper , S. H. 2002 . Water-level management as a tool for the restoration of shallow lakes in the Netherlands . Lake Reserv. Manage. , 18 : 293 – 298 .
- Crivelli , A. J. and Grillas , P. 1995 . Responses of vegetation to a rise in water level at Kerkini Reservoir (1982–1991), a Ramsar site in Northern Greece . Environ. Manage. , 19 : 417 – 430 .
- Elias , J. E. and Meyer , M. W. 2003 . Comparisons of undeveloped and developed shorelands, northern Wisconsin, and recommendations for restoration . Wetlands , 23 : 800 – 816 .
- Environmental Agency of Japan . 2000 . Threatened Wildlife of Japan: Red Data Book , 2nd , Tokyo : Japan Wildlife Research Center . in Japanese
- Galinato , M. and van der Valk , A. 1986 . Seed germination traits of annuals and emergents recruited during drawdown in the Delta Marsh, Manitoba, Canada . Aquat. Bot. , 26 : 89 – 102 .
- Harper , J. L. 1977 . “ Population biology of plants ” . London : Academic Press .
- Hill , N. M. , Keddy , P. A. and Wisheu , I. C. 1998 . A hydrological model for predicting the effects of dams on the shoreline vegetation of lakes and reservoirs . Environ. Manage. , 22 : 723 – 736 .
- Lenssen , J. P.M. , ten Dolle , G. E. and Blom , C. W.P.M. 1998 . The effect of flooding on the recruitment of reed marsh and tall forb plant species . Plant Ecol. , 139 : 13 – 23 .
- Maemets , H. and Freiberg , L. 2005 . Long- and short-term changes of the macrophyte vegetation in strongly stratified hypertrophic Lake Verevi . Hydrobiologia , 547 : 175 – 184 .
- Miyawaki , S. , Nishihiro , J. , Nakamura , K. and Washitani , I. 2004 . Spatio-temporal pattern and factors of vegetation decline in Lake Kasumigaura . Jpn. J. Conserv. Ecol. , 9 : 45 – 55 . in Japanese with English summary
- Nilsson , C. and Keddy , P. A. 1988 . Predictability of change in shoreline vegetation in a hydroelectric reservoir, Northern Sweden . Can. J. Fish. Aquat. Sci. , 45 : 1896 – 1904 .
- Takamura , N. 2009 . “ Indicators of ecosystem integrity in lake littoral area ” . In New perspective in the ecosystem restoration studies , Tokyo , , Japan : Kyouritsu-Shuppan . [forthcoming 2009] (in Japanese)
- Nishihiro , J. , Miyawaki , S. , Fujiwara , N. and Washitani , I. 2004a . Regeneration failure of lakeshore plants under an artificially altered water regime . Ecol. Res. , 19 : 613 – 623 .
- Nishihiro , J. , Araki , S. , Fujiwara , N. and Washitani , I. 2004b . Germination characteristics of lakeshore plants under an artificially stabilized water regime . Aquat. Bot. , 79 : 333 – 343 .
- Nishihiro , J. , Nishihiro , M. A. and Washitani , I. 2006 . Assessing the potential for recovery of lakeshore vegetation: species richness of sediment propagule banks . Ecol. Res. , 21 : 436 – 445 .
- Nishihiro , J. and Washitani , I. 2007 . Restoration of lakeshore vegetation using sediment seed banks; studies and practices in Lake Kasumigaura, Japan . Global Environ. Res. , 11 : 171 – 177 .
- Sakurai , Y. 1981 . Changes of flora, vegetation area and biomass of aquatic plants in the recent progress of eutrophication in Lake Kasumigaura . Research Report of the National Institute of Environmental Studies , 22 : 229 – 279 . in Japanese with English summary
- Schmieder , K. 2004 . European lake shores in danger – concepts for sustainable development . Limnologica , 34 : 3 – 14 .
- Schneider , R. L. 1994 . The role of hydrologic regime in maintaining rare plant communities of New York's coastal plain pondshores . Biol. Conserv. , 68 : 253 – 260 .
- Seabloom , E. W. , van der Valk , A. G. and Moloney , K. A. 1998 . The role of water depth and soil temperature in determining initial composition of prairie wetland coenoclines . Plant Ecol. , 138 : 203 – 216 .
- Shay , J. M. , de Geus , P. M.J. and Kapinga , M. R.M. 1999 . Changes in shoreline vegetation over a 50-year period in the Delta Marsh, Manitoba, in response to water levels . Wetlands , 19 : 413 – 425 .
- Silvertown , J. W. and Dickie , J. B. 1981 . Seedling survivorship in natural populations of nine perennial chalk grassland plants . New Phytol. , 88 : 555 – 558 .
- Steinman , A. , Havens , K. and Hornung , L. 2002 . The managed recession of Lake Okeechobee, Florida: integrating science and natural resource management . Cons. Ecol. , 6 ( 2 ) : 17 http://www.ecologyandsociety.org/vol6/iss2/art17
- Streng , D. R. , Glitzenstein , J. S. and Harcombe , P. A. 1989 . Woody seedling dynamics in an east Texas floodplain forest . Ecol. Monogr. , 59 : 177 – 204 .
- van der Valk , A. 1981 . Succession in wetlands: a Gleasonian approach . Ecology , 62 : 688 – 696 .
- Watkinson , A. R. and Powell , J. C. 1993 . Seedling recruitment and the maintenance of clonal diversity in plant populations—a computer simulation of Ranunculus repens . J. Ecol. , 81 : 707 – 717 .
- Wetzel , R. G. 2001 . Limnology , 3rd , 1006 London : Academic Press .