
Abstract
The human uterine cervix can produce nitric oxide (NO), a free radical with an ultra‐short half‐life. The release of NO changes during pregnancy and is increased in early nonviable pregnancies compared to normal uncomplicated pregnancies. This review concentrates on the role of NO release in cervical ripening in pregnant women. Also some suggestions on future aspects are discussed.
Introduction
Nitric oxide (NO) is a small, uncharged gas molecule that is a highly reactive free radical with an extremely short half‐life of approximately 4 seconds. In the human female reproductive tract NO plays an important role in various physiological processes, including cervical ripening Citation1.
The uterine cervix has a pivotal role in the physiology of gestation and parturition; it has to be firm enough to retain the conceptus throughout the pregnancy and, on the other hand, to soften before and during labor to enable the birth of the infant. Cervical adaptation during pregnancy is a chronic process, which begins within the first trimester of pregnancy and progressively proceeds until term, and is usually described as softening, ripening, dilation, and finally, remodeling of the cervix. Obviously with this four‐step process, which must occur in sequence, complex systems must be in place to control each forward and irreversible step Citation2–4. Effacement and dilation are often associated with or contributed to uterine contractions, but it is evident that softening occurs independently of contractions. Further, effacement and dilation are often present without uterine contractions.
The cervix, which is dominated by fibrous connective tissue, is composed of an extracellular matrix consisting predominantly of collagen (70% type I and 30% type III) along with elastin, proteo‐ and glucosaminoglycans, and water, and a cellular portion consisting of smooth muscle, fibroblasts, epithelium, and blood vessels Citation5. Throughout most of gestation the cervix remains rigid and closed to secure the products of conception. A dramatic functional shift occurs during parturition to secure uncomplicated delivery. The mechanisms involved in cervical ripening are complex and are only known to a certain extent. The cascade responsible for the process of cervical ripening, and which finally enables uterine contractions to efface and dilate the cervix, is complex.
The cervical ripening is actively controlled with features similar to those in inflammation and in rearrangement of the cervical collagen fibers Citation6, Citation7. Cervical ripening is thus associated with changes in local cytokines, prostaglandins, and metalloproteases, as well as in other bioregulators which play roles in inflammation and in collagen metabolism Citation6, Citation7. These factors take part also in the regulation of NO synthesis and release Citation1, and the present review is to clarify the role of cervical NO in the ripening of the human uterine cervix.
Synthesis of nitric oxide
Nitric oxide is formed from L‐arginine Citation8 through nitric oxide synthases (NOS) Citation9, a group of enzymes that structurally resemble cytochrome P‐450 reductase Citation10. The biosynthesis of NO takes place from L‐arginine and molecular oxygen utilizing nicotine adenine dinucleotide phosphate as an electron donor, and heme, tetrahydrobiopterin, calmodulin, flavin adenine mono‐ and dinucleotide as cofactors through a reaction that consumes five electrons. The overall reaction consists of a two‐step oxidative conversion of L‐arginine to NO and L‐citrulline via Nw‐hydroxy‐L‐arginine as an intermediate, with monooxygenase I and monooxygenase II, in each step a mixed‐function oxidation taking place.
Three NOS isoenzymes have been characterized: neuronal NOS (type I, nNOS), inducible NOS (type II or iNOS), and endothelial NOS (type III or eNOS) Citation11–13. Both nNOS and eNOS are expressed constitutively, and their activity is calcium/calmodulin‐dependent, whereas the expression of iNOS is induced by bacterial lipopolysaccharides (LPS) and cytokines, independently of calcium Citation14 (Table ).
Table I. Randomized controlled trials on nitric oxide donors in cervical ripening in pregnant women.
Endothelial NOS is mainly expressed in endothelial cells Citation11 and platelets Citation15, while nNOS is expressed in the cerebellum and skeletal muscle Citation13. Inducible NOS was first cloned from activated mouse macrophages Citation12, and thereafter, it has been demonstrated in various human cells including macrophages Citation16.
Nitric oxide as a mediator
Nitric oxide serves as a highly diffusible molecule that can affect cells both directly and indirectly. The direct effects are mediated by the NO molecule itself, while the indirect ones are mediated by reactive nitrogen produced by the interaction of NO with oxygen or superoxide radicals (O2‐). At the low concentrations of NO produced through eNOS and nNOS, the direct effects prevail, while at higher concentrations of NO, produced through iNOS, the indirect effects dominate Citation17.
The formation of cyclic guanosine 3',5'‐monophosphate (cGMP) accounts for many of the direct physiological effects of NO Citation17. Nitric oxide may also interact with nonheme iron‐containing and zinc‐containing proteins, or form S‐nitrosothiols by nitrosylation Citation18.
The indirect effects of NO include oxidation, nitrosation, and nitration. Cytokine‐induced NO production mediates cytotoxicity in the target cells of macrophages Citation19. In a reaction with O2 (autooxidation) NO forms dinitrogen trioxide (N2O3), which can mediate DNA deamination and nitrosylation. By reacting with superoxide NO produces peroxynitrite (ONOO‐), which is a toxic nitrating agent and a powerful oxidant, modifying proteins, lipids, tyrosine, and nucleic acids.
Cervical nitric oxide
All three NOSs (nNOS, iNOS, and eNOS) are present in the human uterine cervix Citation20–22: neuronal NOS is localized in stroma and epithelial cells Citation22, iNOS in the epithelial cells and stromal spindle cells Citation20, and eNOS in vascular endothelium Citation20.
In animal studies, NO induced cervical ripening Citation23, and cervical NO release was elevated during labor Citation24. In a recent bovine study, relative iNOS mRNA levels were found the highest in the beginning of the third trimester and decreased significantly from towards parturition Citation25. It could be argued that the decrease in iNOS protein expression was due to changes in the composition of the extracellular matrix (ECM) in the cervix, which occur during late pregnancy and parturition Citation26. In addition, NO induces relaxation of smooth muscle in cervix Citation27.
Nitric oxide shares with tumor necrosis factor α (TNF‐α) the unique ability to initiate and to block apoptosis, depending on multiple variables that are being elucidated Citation28. Therefore, NO is both an antiapoptotic and apoptotic substance, which may arrest the cellular turnover and allow the reorganization of the collagen Citation23, Citation26. Nitric oxide may act in concert with prostaglandin E2 (PG E2) by inducing local vasodilatation and by increasing vascular permeability and leukocyte infiltration Citation29. In addition, NO may directly regulate the activity or the production of matrix metalloproteases (MMPs) Citation1, although Ledingham et al. Citation30 demonstrated that the secretion of MMP‐2 and MMP‐9 in cervical fibroblasts was not regulated by exogenous NO. If NO modulates MMPs, the action of NO both in uterus and cervix may be mediated partly by MMPs.
During normal pregnancy
In human studies, iNOS has been reported to become stimulated in the cervix during cervical ripening (Figure ) Citation20, Citation21, Citation31, although not in all studies Citation32. In addition, data are not uniform as regards the changes in expression of cervical nNOS and eNOS during term vaginal delivery: in some studies no change was seen Citation20, Citation21, but in the others, the cervical nNOS expression became stimulated Citation22.
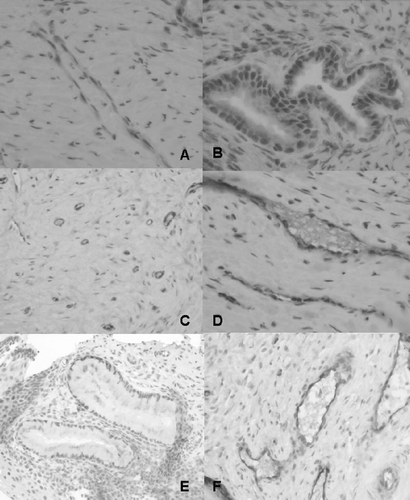
Figure 1 Immunoreactivity of iNOS and eNOS in cervixes of women with early pregnancy. Biopsy samples from mifepristone‐treated (A–D) and control women (E, F). Strong iNOS staining is seen in the endothelium in A (×200) and in the cervical glands in B (×400), but not in the control woman (E, ×200). Endothelial NOS staining is restricted to vascular endothelial cells in mifepristone‐treated (C, ×200; D, ×400) and control women (F, ×200).
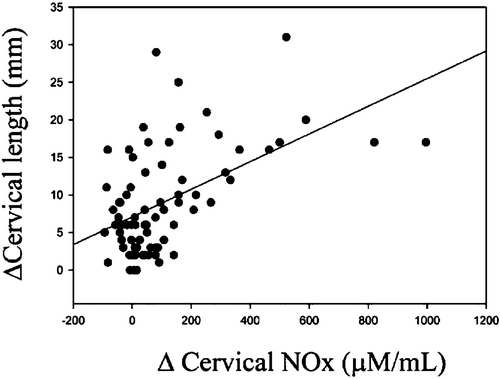
Cervical NO release increases during human pregnancy obtained by various methods Citation20–22, Citation33 (Figure ). Pregnancy‐related increase in cervical NO release has been detected also in animal studies Citation1, Citation24, Citation34–36. Cervical NO release relates to Bishop scores Citation33 and, moreover, Chiossi et al. found a significant positive correlation between changes in cervical NOx (nitric oxide metabolite) levels and Bishop score modification (P<0.01; r = 0.494), as well as between the modification of NO metabolites concentration and cervical shortening (P<0.01; r = 0.307) Citation37. Parous women have higher cervical fluid NOx concentrations than nulliparous women Citation33. A recent study showed that the iNOS gene was downregulated in the cervixes of women after term vaginal labor Citation38. This may imply that during active labor iNOS‐derived cervical NO release is no longer needed, because the primary task of cervical iNOS is in cervical ripening.
Figure 2 Cervical fluid NOx(nitric oxide metabolite) concentrations and group‐specific medians in nonpregnant and pregnant (pregn.) women (µmol/L) in logarithmic scale.
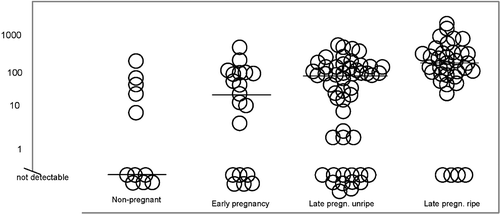
Figure 3 Correlation between changes in cervical length and changes in cervical NOx(nitric oxide metabolite), 6 hours after dinoprostone application. From reference Citation37 with permission.
In women the concentration of NOx in vaginal secretions has been reported to be elevated before preterm birth Citation39, Citation40. Although the source of this NOx is not known, both infiltrating inflammatory cells and cells in the uterine cervix may be responsible. Because NO can activate MMPs Citation41, Citation42 and induce apoptotic cell death Citation28, Citation43, the overproduction of NO may be involved in cervical ripening, fragility of membranes, and subsequent premature delivery.
In nonviable early pregnancy
Cervical NO release is increased in women with failed early intrauterine pregnancy, but not in women with tubal pregnancy Citation44. Increased cervical NO, although a very reactive molecule, is scarcely a primary cause of miscarriage. This is supported by the reduced placental expression of iNOS at the fetomaternal interface in women with spontaneous abortion when compared with that in women with early viable pregnancy Citation45. The increased cervical NO release in nonviable intrauterine pregnancy could have the following explanations. First, elevated release of NO in the dying fetus, decidua, and fetal membranes has been found in LPS‐induced abortions in animal experiments Citation46, Citation47, but no such human data exist so far. However, it appears plausible that remnants of the conceptus could have released NO excessively, which could have leaked, as either NO or NOx, into the cervical canal. This hypothesis is supported by our findings: cervical fluid NOx levels were elevated only in intrauterine miscarriages, not in tubal ones, and missed abortion, with potentially more abundant remnants of conception, was characterized by higher cervical fluid NOx levels than cases of blighted ovum Citation44. Second, miscarriage is often associated with a local inflammatory reaction in the cervix, and this may result in the stimulation of NO release through PGs or cytokines Citation1, Citation7. In addition, many other hormones, such as inhibins Citation48–50, may be involved in spontaneous abortion and may have secondarily stimulated cervical NO release. Third, increased cervical NO release may be a specific phenomenon in miscarriage, perhaps triggered by a fall in the level of progesterone either locally or in the serum (Figure ).
In postterm pregnancy
Cervical NO release is deficient in postterm pregnancy (Figure ) Citation51. It remains unclear if cervical NO deficiency is a primary phenomenon, and thus a true contributing factor to postterm pregnancy, or whether it is a reflection of relative insufficiency of PGs, cytokines, MMPs, or some other agents which may be primarily involved in cervical ripening Citation7, Citation41, Citation52–58 and which may stimulate NO release. A high progesterone level seems to downregulate the synthesis and release of cervical NO. Recent data indicate that among women with an initial postterm labor there is a significant risk for subsequent postterm deliveries Citation59, Citation60. This characteristic seems to be genetically determined Citation61, and therefore we speculate that ‘postterm genes’ are functionally linked to the genes regulating NO synthases. Such women might benefit from the administration of a vaginal NO donor when induction of labor is needed Citation62.
Figure 4 Cervical fluid nitric oxide metabolite(NOx) concentrations in women going postterm and in women delivering spontaneously at term (µmol/L). Medians are shown by lines. The detection limit of the assay was 3.8 µmol/L.
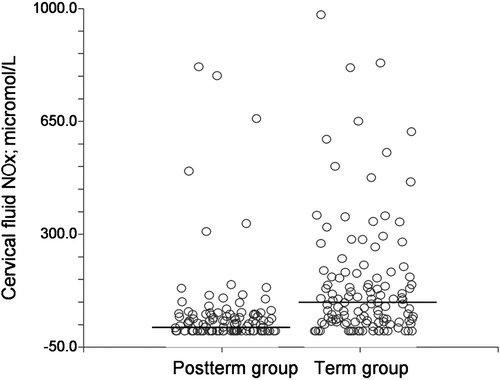
Figure 5 A simplified schematic model of the possible role of cervical nitric oxide in human cervical ripening(NOSs = nitric oxide synthases; PG = prostaglandin; MMP = matrix metalloprotease).
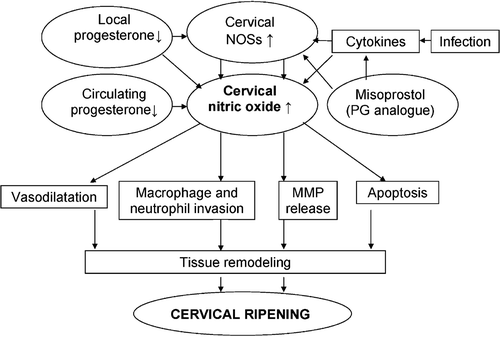
Relationships to prostaglandins
Nitric oxide and PGs operate jointly in many cells Citation63–67. Nitric oxide may either stimulate or inhibit the release of COX‐2 (cyclooxygenase), and likewise PGs may have a stimulatory or inhibitory effect on iNOS, depending on the cell type and/or the presence of cofactors Citation1, Citation67–69. Misoprostol as a PG analogue induces NO release in the uterine cervix of pregnant women, and furthermore the response of cervical NO release to PG becomes enhanced from early to late pregnancy Citation70. Thus, PG analogues such as misoprostol can perhaps initiate a chain reaction in the cervix of pregnant women; the initial NO stimulation caused by misoprostol is followed by endogenous release of PGs triggered by NO. As a result, NO, PG, and COX pathways may have a joint action in human cervical ripening, as schematically shown in Figure .
Reducing effect of progesterone
Numerous animal experiments support the view of progesterone having opposing effects on NO release in the endomyometrium and cervix; it upregulates NO release in the former, but downregulates it in the latter Citation1, Citation24, Citation34–36. The data imply a link between progesterone and cervical NO (Figure ). First, the lower the progesterone level the higher the detection rate of cervical fluid NOx in nonpregnant women Citation33, Citation70. In fact, 93% of women in the follicular phase, versus 46% of women in the luteal phase, showed detectable cervical fluid NOx. Circulating NOx levels are also higher during the follicular phase and at the time of ovulation than in the luteal phase Citation71. Second, cervical NO release was inversely related to circulating progesterone concentrations in early nonviable pregnancy Citation44. Women with threatened abortion or preterm birth have considerably lower levels of circulating progesterone than women with normal pregnancy Citation72. Progesterone insufficiency could well have stimulated cervical NO release in nonviable intrauterine pregnancy, which may be needed for ripening of the cervix during the course of miscarriage. Third, cervical NO responded to the antiprogestin mifepristone with a 17‐fold increase in cervical fluid NOx and with increased expression of iNOS in early viable pregnancy Citation31. Furthermore, mifepristone induced the appearance of iNOS in cervical glands Citation31 (Figure ).
The mechanisms behind mifepristone‐induced NO release are not known, but a progesterone receptor‐mediated pathway may be involved Citation73. Local progesterone withdrawal in the cervix brought about by mifepristone may lead specifically to the stimulation of iNOS. Alternatively, the antiglucocorticoid properties of mifepristone, blocking glucocorticoid receptors, may stimulate iNOS Citation74. This would fit well with data showing that the levels of glucocorticoid receptor decrease in the human cervix during labor Citation55. Furthermore, it is possible that mifepristone triggers an influx of inflammatory cells, such as macrophages, neutrophils, and monocytes, which are rich in iNOS. Moreover, mifepristone upregulates various MMPs and/or the secretion of cytokines in cervical cells Citation1, Citation55, Citation75. These mediators may either induce or inhibit iNOS, depending on the availability of various cofactors Citation74, Citation75. Furthermore, NO may act in concert with the COX pathway, especially with COX‐2 Citation1, Citation28, Citation65, Citation76 (Figure ). Nitric oxide in turn may soften the cervix by remodeling the ECM Citation1 (Figure ), where cytokine‐induced NOS is centrally involved Citation1, Citation20, or by inducing apoptosis of cervical cells Citation1, Citation28. Taken as a whole, there is strong evidence Citation1, Citation31, Citation34, Citation44, Citation77, Citation78 that NO in the cervix, and progesterone, are functionally related in pregnancy (Figure ). In fact, administration of progesterone is used in treatment of threatened abortion or preterm birth Citation79–83.
Possible use of nitric oxide in Obstetrics and gynecology
In animals, NO donors were found to ripen the cervix Citation77, Citation84. In women, NO donors, such as isosorbide mononitrate (IMN) Citation85–93, sodium nitroprusside (SNP) Citation94, Citation95, and glyceryl trinitrate (GTN) Citation85, Citation96–98, administered intravaginally or intracervically, ripen cervix during pregnancy (Table ). In general, NO donors appear less effective than PGs, at least in viable pregnancies, but in nonviable early pregnancy there is discrepancy concerning the effectiveness of IMN compared to misoprostol Citation91, Citation99, Citation100 (Table ). Interestingly, although the combination of NO donor and PG, simultaneously applied, did not ripen the cervix more effectively in 6 hours, it shortened the time interval between intervention and active labor Citation98. Sodium nitroprusside ripens cervix even in nonpregnant women Citation101.
Glyceryl trinitrate administrated intracervically induced cervical NO release at term Citation33, and intravaginal administration of GTN together with PG E2 (Dinoprost®) resulted in a shorter induction‐to‐vaginal‐delivery time Citation98. Isosorbide mononitrate induced COX‐2, but not COX‐1 in human cervix at term Citation102.
Nitric oxide donors are safe and have no major side effects on fetus or mother Citation62, Citation86, Citation87, Citation93, Citation100, Citation103–106. When compared with misoprostol NO donors were less effective Citation106, Citation107 but did not cause the uterine hyperstimulation Citation86, Citation108. Thus, NO donors hold promise in cervical ripening in women, although additional data are needed before they can be routinely used in clinics.
Cervical fluid NOx may perhaps be used as a marker of cervical ripeness in clinics. Clinicians could benefit from its assessment both in early and late pregnancy because this test may indicate the readiness of the cervical canal for misoprostol or mifepristone priming Citation31, Citation70. In early pregnancy, women with nonviable early pregnancy and ‘high’ cervical fluid NOx could perhaps be treated expectantly Citation44, because these women might belong to the 50% of such women who will abort spontaneously within 2 weeks Citation109. If the test result is ‘low’, priming of the cervix with PGs or mifepristone can be considered. These questions should be studied in clinical trials. Likewise, as deficient cervical NO release was related to failed progression of labor Citation51, the cervical fluid NOx level may perhaps be used to predict the likelihood of successful induction of labor. In early as well as in postterm pregnancy, NO donors may hold a promise, but this question has been studied only in postterm pregnancy so far Citation62.
No reliable biochemical marker for the identification of women at risk of preterm birth exists. Fetal fibronectin and insulin‐like growth factor‐binding protein‐1 (IGFBP‐1) have been extensively studied as marker candidates, but they have poor predictive values and there are major confounders in sample collection Citation110–112. Transvaginal ultrasonography is also insufficient in predicting preterm birth, mostly because of the wide variance in predictive value Citation113, Citation114. Cervical fluid NOx might prove to be a feasible marker for this condition. An epidemiological study in this connection would need a huge number of women, the risk of spontaneous preterm birth being 2%–3% in Finland. Therefore, we should first study women who report preterm contractions in order to see if this test can differentiate between those women who carry to term and those who deliver preterm. Some preliminary data suggest that this test could be useful in such women Citation40.
Acknowledgements
Work was financially supported by grants from the Research Funds of Helsinki University Central Hospital, the Emil Aaltonen Foundation, and the Research Foundation of Orion Corporation.
References
- Maul H., Longo M., Saade G. R., Garfield R. E. Nitric oxide and its role during pregnancy: from ovulation to delivery. Curr Pharm Des 2003; 9: 359–80
- Leppert P. C. Anatomy and physiology of cervical ripening. Clin Obstet Gynecol 1995; 38: 267–79
- Maul H., Mackay L., Garfield R. E. Cervical ripening: biochemical, molecular, and clinical considerations. Clin Obstet Gynecol 2006; 49: 551–63
- Word R. A., Li X. H., Hnat M., Carrick K. Dynamics of cervical remodeling during pregnancy and parturition: mechanisms and current concepts. Semin Reprod Med 2007; 25: 69–79
- Ludmir J., Sehdev H. M. Anatomy and physiology of the uterine cervix. Clin Obstet Gynecol 2000; 43: 433–39
- Junqueira L. C., Zugaib M., Montes G. S., Toledo O. M., Krisztan R. M., Shigihara K. M. Morphologic and histochemical evidence for the occurrence of collagenolysis and for the role of neutrophilic polymorphonuclear leukocytes during cervical dilation. Am J Obstet Gynecol 1980; 138: 273–81
- Liggins G. C. Cervical ripening as an inflammatory reaction. The cervix in pregnancy and labour. Clinical and biochemical investigation, D. A Ellwood, A. M. B Anderson. Churchill Livingstone, Edinburgh 1981; 1–9
- Palmer R. M., Ashton D. S., Moncada S. Vascular endothelial cells synthesize nitric oxide from L‐arginine. Nature 1988; 333: 664–6
- Palmer R. M., Moncada S. A novel citrulline‐forming enzyme implicated in the formation of nitric oxide by vascular endothelial cells. Biochem Biophys Res Commun 1989; 158: 348–52
- Marletta M. A. Nitric oxide synthase: aspects concerning structure and catalysis. Cell 1994; 78: 927–30
- Pollock J. S., Forstermann U., Mitchell J. A., Warner T. D., Schmidt H. H., Nakane M., et al. Purification and characterization of particulate endothelium‐derived relaxing factor synthase from cultured and native bovine aortic endothelial cells. Proc Natl Acad Sci U S A 1991; 88: 10480–4
- Xie Q. W., Cho H. J., Calaycay J., Mumford R. A., Swiderek K. M., Lee T. D., et al. Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science 1992; 256: 225–8
- Nakane M., Schmidt H. H., Pollock J. S., Forstermann U., Murad F. Cloned human brain nitric oxide synthase is highly expressed in skeletal muscle. FEBS Lett 1993; 316: 175–80
- Knowles R. G., Moncada S. Nitric oxide synthases in mammals. Biochem J 1994; 298: 249–58
- Radomski M. W., Palmer R. M., Moncada S. Characterization of the L‐arginine: nitric oxide pathway in human platelets. Br J Pharmacol 1990; 101: 325–8
- Moilanen E., Moilanen T., Knowles R., Charles I., Kadoya Y., al‐Saffar N., et al. Nitric oxide synthase is expressed in human macrophages during foreign body inflammation. Am J Pathol 1997; 150: 881–7
- Murad F. Discovery of some of the biological effects of nitric oxide and its role in cell signaling. Biosci Rep 1999; 19: 133–54
- Hogg N. The biochemistry and physiology of S‐nitrosothiols. Annu Rev Pharmacol Toxicol 2002; 42: 585–600
- Farrell A. J., Blake D. R. Nitric oxide. Ann Rheum Dis 1996; 55: 7–20
- Tschugguel W., Schneeberger C., Lass H., Stonek F., Zaghlula M. B., Czerwenka K., et al. Human cervical ripening is associated with an increase in cervical inducible nitric oxide synthase expression. Biol Reprod 1999; 60: 1367–72
- Ledingham M. A., Thomson A. J., Young A., Macara L. M., Greer I. A., Norman J. E. Changes in the expression of nitric oxide synthase in the human uterine cervix during pregnancy and parturition. Mol Hum Reprod 2000; 6: 1041–8
- Bao S., Rai J., Schreiber J. Brain nitric oxide synthase expression is enhanced in the human cervix in labor. J Soc Gynecol Investig 2001; 8: 158–64
- Chwalisz K., Shao‐Qing S., Garfield R. E., Beier H. M. Cervical ripening in guinea‐pigs after a local application of nitric oxide. Hum Reprod 1997; 12: 2093–101
- Buhimschi I., Ali M., Jain V., Chwalisz K., Garfield R. E. Differential regulation of nitric oxide in the rat uterus and cervix during pregnancy and labour. Hum Reprod 1996; 11: 1755–66
- Aalberts M., van Dissel‐Emiliani F. M., van Tol H. T., Taverne M. A., Breeveld‐Dwarkasing V. N. High iNOS mRNA and protein levels during early third trimester suggest a role for NO in prelabor cervical ripening in the bovine. Mol Reprod Dev 2007; 74: 378–85
- Leppert P. C., Kokenyesi R., Klemenich C. A., Fisher J. Further evidence of a decorin‐collagen interaction in the disruption of cervical collagen fibers during rat gestation. Am J Obstet Gynecol 2000; 182: 805–11
- Ekerhovd E., Brannstrom M., Weijdegard B., Norstrom A. Nitric oxide synthases in the human cervix at term pregnancy and effects of nitric oxide on cervical smooth muscle contractility. Am J Obstet Gynecol 2000; 183: 610–6
- Brune B., von Knethen A., Sandau K. B. Nitric oxide and its role in apoptosis. Eur J Pharmacol 1998; 351: 261–72
- Ekerhovd E., Weijdegard B., Brannstrom M., Mattsby‐Baltzer I., Norstrom A. Nitric oxide induced cervical ripening in the human: Involvement of cyclic guanosine monophosphate, prostaglandin F(2 alpha), and prostaglandin E(2). Am J Obstet Gynecol 2002; 186: 745–50
- Ledingham M. A., Denison F. C., Riley S. C., Norman J. E. Matrix metalloproteinases‐2 and ‐9 and their inhibitors are produced by the human uterine cervix but their secretion is not regulated by nitric oxide donors. Hum Reprod 1999; 14: 2089–96
- Vaisanen‐Tommiska M., Butzow R., Ylikorkala O., Mikkola T. S. Mifepristone‐induced nitric oxide release and expression of nitric oxide synthases in the human cervix during early pregnancy. Hum Reprod 2006; 21: 2180–4
- Thomson A. J., Telfer J. F., Kohnen G., Young A., Cameron I. T., Greer I. A., et al. Nitric oxide synthase activity and localization do not change in uterus and placenta during human parturition. Hum Reprod 1997; 12: 2546–52
- Vaisanen‐Tommiska M., Nuutila M., Aittomaki K., Hiilesmaa V., Ylikorkala O. Nitric oxide metabolites in cervical fluid during pregnancy: further evidence for the role of cervical nitric oxide in cervical ripening. Am J Obstet Gynecol 2003; 188: 779–85
- Ali M., Buhimschi I., Chwalisz K., Garfield R. E. Changes in expression of the nitric oxide synthase isoforms in rat uterus and cervix during pregnancy and parturition. Mol Hum Reprod 1997; 3: 995–1003
- Garfield R. E., Saade G., Buhimschi C., Buhimschi I., Shi L., Shi S. Q., et al. Control and assessment of the uterus and cervix during pregnancy and labour. Hum Reprod Update 1998; 4: 673–95
- Chwalisz K., Garfield R. E. Role of nitric oxide in the uterus and cervix: implications for the management of labor. J Perinat Med 1998; 26: 448–57
- Chiossi G., Verocchi G., Venturini P., Facchinetti F. Changes in cervical nitric oxide concentration correlate with Bishop score and cervical length modifications in prostaglandin e2‐mediated induction of labor. J Soc Gynecol Investig 2006; 13: 203–8
- Huber A., Hudelist G., Czerwenka K., Husslein P., Kubista E., Singer C. F. Gene expression profiling of cervical tissue during physiological cervical effacement. Obstet Gynecol 2005; 105: 91–8
- Nakatsuka M., Habara T., Kamada Y., Tada K., Kudo T. Elevation of total nitrite and nitrate concentration in vaginal secretions as a predictor of premature delivery. Am J Obstet Gynecol 2000; 182: 644–5
- Facchinetti F., Venturini P., Blasi I., Giannella L. Changes in the cervical competence in preterm labour. BJOG 2005; 112 Suppl 1: 23–7
- Yoshida M., Sagawa N., Itoh H., Yura S., Korita D., Kakui K., et al. Nitric oxide increases matrix metalloproteinase‐1 production in human uterine cervical fibroblast cells. Mol Hum Reprod 2001; 7: 979–85
- Biondi C., Pavan B., Lunghi L., Fiorini S., Vesce F. The role and modulation of the oxidative balance in pregnancy. Curr Pharm Des 2005; 11: 2075–89
- Leppert P. C. Proliferation and apoptosis of fibroblasts and smooth muscle cells in rat uterine cervix throughout gestation and the effect of the antiprogesterone onapristone. Am J Obstet Gynecol 1998; 178: 713–25
- Vaisanen‐Tommiska M., Mikkola T. S., Ylikorkala O. Increased release of cervical nitric oxide in spontaneous abortion before clinical symptoms: a possible mechanism for preabortal cervical ripening. J Clin Endocrinol Metab 2004; 89: 5622–6
- Marinoni E., Di Iorio R., Lucchini C., Di Netta T., Letizia C., Cosmi E. V. Adrenomedullin and nitric oxide synthase at the maternal‐decidual interface in early spontaneous abortion. J Reprod Med 2004; 49: 153–61
- Haddad E. K., Duclos A. J., Baines M. G. Early embryo loss is associated with local production of nitric oxide by decidual mononuclear cells. J Exp Med 1995; 182: 1143–51
- Ogando D. G., Paz D., Cella M., Franchi A. M. The fundamental role of increased production of nitric oxide in lipopolysaccharide‐induced embryonic resorption in mice. Reproduction 2003; 125: 95–110
- Reis F. M., Faletti A., Luisi S., Bifulco G., Cauci S., Quadrifoglio F., et al. High concentrations of inhibin A and inhibin B in ovarian serous cystadenoma: relationship with oestradiol and nitric oxide metabolites. Mol Hum Reprod 2000; 6: 1079–83
- Muttukrishna S., Jauniaux E., Greenwold N., McGarrigle H., Jivraj S., Carter S., et al. Circulating levels of inhibin A, activin A and follistatin in missed and recurrent miscarriages. Hum Reprod 2002; 17: 3072–8
- Lahiri S., Anobile C. J., Stewart P., Ledger W. L. Changes in circulating concentrations of inhibins A and pro‐alpha C during first trimester medical termination of pregnancy. Hum Reprod 2003; 18: 744–8
- Vaisanen‐Tommiska M., Nuutila M., Ylikorkala O. Cervical nitric oxide release in women postterm. Obstet Gynecol 2004; 103: 657–62
- Sennstrom M. K., Brauner A., Lu Y., Granstrom L. M., Malmstrom A. L., Ekman G. E. Interleukin‐8 is a mediator of the final cervical ripening in humans. Eur J Obstet Gynecol Reprod Biol 1997; 74: 89–92
- Sennstrom M. B., Brauner A., Bystrom B., Malmstrom A., Ekman G. Matrix metalloproteinase‐8 correlates with the cervical ripening process in humans. Acta Obstet Gynecol Scand 2003; 82: 904–11
- Stygar D., Wang H., Vladic Y. S., Ekman G., Eriksson H., Sahlin L. Increased level of matrix metalloproteinases 2 and 9 in the ripening process of the human cervix. Biol Reprod 2002; 67: 889–94
- Stjernholm‐Vladic Y., Stygar D., Mansson C., Masironi B., Akerberg S., Wang H., et al. Factors involved in the inflammatory events of cervical ripening in humans. Reprod Biol Endocrinol 2004; 2: 74
- Kelly R. W. Inflammatory mediators and cervical ripening. J Reprod Immunol 2002; 57: 217–24
- Osman I., Young A., Ledingham M. A., Thomson A. J., Jordan F., Greer I. A., et al. Leukocyte density and pro‐inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol Hum Reprod 2003; 9: 41–5
- Yoshida M., Sagawa N., Itoh H., Yura S., Takemura M., Wada Y., et al. Prostaglandin F(2alpha), cytokines and cyclic mechanical stretch augment matrix metalloproteinase‐1 secretion from cultured human uterine cervical fibroblast cells. Mol Hum Reprod 2002; 8: 681–7
- Olesen A. W., Basso O., Olsen J. Risk of recurrence of prolonged pregnancy. BMJ 2003; 326: 476
- Kistka Z. A., Palomar L., Boslaugh S. E., DeBaun M. R., DeFranco E. A., Muglia L. J. Risk for postterm delivery after previous postterm delivery. Am J Obstet Gynecol 2007; 196: 241.e1–e6
- Laursen M., Bille C., Olesen A. W., Hjelmborg J., Skytthe A., Christensen K. Genetic influence on prolonged gestation: a population‐based Danish twin study. Am J Obstet Gynecol 2004; 190: 489–94
- Bullarbo M., Orrskog M. E., Andersch B., Granstrom L., Norstrom A., Ekerhovd E. Outpatient vaginal administration of the nitric oxide donor isosorbide mononitrate for cervical ripening and labor induction postterm: a randomized controlled study. Am J Obstet Gynecol 2007; 196: 50.e1–e5
- Dong Y. L., Gangula P. R., Fang L., Yallampalli C. Nitric oxide reverses prostaglandin‐induced inhibition in ovarian progesterone secretion in rats. Hum Reprod 1999; 14: 27–32
- Ledingham M. A., Denison F. C., Kelly R. W., Young A., Norman J. E. Nitric oxide donors stimulate prostaglandin F(2alpha) and inhibit thromboxane B(2) production in the human cervix during the first trimester of pregnancy. Mol Hum Reprod 1999; 5: 973–82
- Hausman N., Beharry K. D. B. S., Nishihara K. C. B. A., Akmal Y. B. S., Asrat T. Effect of the antenatal administration of celecoxib during the second and third trimesters of pregnancy on prostaglandin, cytokine, and nitric oxide levels in rabbits. Am J Obstet Gynecol 2003; 189: 1737–43
- Gookin J. L., Duckett L. L., Armstrong M. U., Stauffer S. H., Finnegan C. P., Murtaugh M. P., et al. Nitric oxide synthase stimulates prostaglandin synthesis and barrier function in C. parvum‐infected porcine ileum. Am J Physiol Gastrointest Liver Physiol 2004; 287: G571–81
- Timoshenko A. V., Lala P. K., Chakraborty C. PGE2‐mediated upregulation of iNOS in murine breast cancer cells through the activation of EP4 receptors. Int J Cancer 2004; 108: 384–9
- Goharkhay N., Wing D. A., Pan V., McCausland V., Hanna M., Naidu Y. M., et al. The expression of EP3‐6 and inducible nitric oxide synthase messenger RNA are correlated in pregnant and misoprostol‐treated but not in nongravid or menopausal myometrium. Am J Obstet Gynecol 2002; 186: 1202–6
- Ogando D., Farina M., Ribeiro M. L., Perez Martinez S., Cella M., Rettori V., et al. Steroid hormones augment nitric oxide synthase activity and expression in rat uterus. Reprod Fertil Dev 2003; 15: 269–74
- Vaisanen‐Tommiska M., Mikkola T. S., Ylikorkala O. Misoprostol induces cervical nitric oxide release in pregnant, but not in nonpregnant, women. Am J Obstet Gynecol 2005; 193: 790–6
- Ekerhovd E., Enskog A., Caidahl K., Klintland N., Nilsson L., Brannstrom M., et al. Plasma concentrations of nitrate during the menstrual cycle, ovarian stimulation and ovarian hyperstimulation syndrome. Hum Reprod 2001; 16: 1334–9
- Gruber C. J., Huber J. C. The role of dydrogesterone in recurrent (habitual) abortion. J Steroid Biochem Mol Biol 2005; 97: 426–30
- Stjernholm‐Vladic Y., Wang H., Stygar D., Ekman G., Sahlin L. Differential regulation of the progesterone receptor A and B in the human uterine cervix at parturition. Gynecol Endocrinol 2004; 18: 41–6
- Alderton W. K., Cooper C. E., Knowles R. G. Nitric oxide synthases: structure, function and inhibition. Biochem J 2001; 357: 593–615
- Denison F. C., Riley S. C., Elliott C. L., Kelly R. W., Calder A. A., Critchley H. O. The effect of mifepristone administration on leukocyte populations, matrix metalloproteinases and inflammatory mediators in the first trimester cervix. Mol Hum Reprod 2000; 6: 541–8
- Hapangama D. K., Critchley H. O., Henderson T. A., Baird D. T. Mifepristone‐induced vaginal bleeding is associated with increased immunostaining for cyclooxygenase‐2 and decrease in prostaglandin dehydrogenase in luteal phase endometrium. J Clin Endocrinol Metab 2002; 87: 5229–34
- Chwalisz K., Garfield R. E. Regulation of the uterus and cervix during pregnancy and labor. Role of progesterone and nitric oxide. Ann NY Acad Sci 1997; 828: 238–53
- Chwalisz K., Winterhager E., Thienel T., Garfield R. E. Synergistic role of nitric oxide and progesterone during the establishment of pregnancy in the rat. Hum Reprod 1999; 14: 542–52
- Facchinetti F., Paganelli S., Comitini G., Dante G., Volpe A. Cervical length changes during preterm cervical ripening: effects of 17‐alpha‐hydroxyprogesterone caproate. Am J Obstet Gynecol 2007; 196: 453.e1–4
- da Fonseca E. B., Bittar R. E., Carvalho M. H., Zugaib M. Prophylactic administration of progesterone by vaginal suppository to reduce the incidence of spontaneous preterm birth in women at increased risk: a randomized placebo‐controlled double‐blind study. Am J Obstet Gynecol 2003; 188: 419–24
- Greene M. F. Progesterone and preterm delivery‐‐deja vu all over again. N Engl J Med 2003; 348: 2453–5
- Schindler A. E. Role of progestogens for the prevention of premature birth. J Steroid Biochem Mol Biol 2005; 97: 435–8
- Di Renzo G. C., Mattei A., Gojnic M., Gerli S. Progesterone and pregnancy. Curr Opin Obstet Gynecol 2005; 17: 598–600
- Shi L., Shi S. Q., Saade G. R., Chwalisz K., Garfield R. E. Studies of cervical ripening in pregnant rats: effects of various treatments. Mol Hum Reprod 2000; 6: 382–9
- Thomson A. J., Lunan C. B., Ledingham M., Howat R. C., Cameron I. T., Greer I. A., et al. Randomised trial of nitric oxide donor versus prostaglandin for cervical ripening before first‐trimester termination of pregnancy. Lancet 1998; 352: 1093–6
- Nicoll A. E., Mackenzie F., Greer I. A., Norman J. E. Vaginal application of the nitric oxide donor isosorbide mononitrate for preinduction cervical ripening: a randomized controlled trial to determine effects on maternal and fetal hemodynamics. Am J Obstet Gynecol 2001; 184: 958–64
- Ekerhovd E., Bullarbo M., Andersch B., Norstrom A. Vaginal administration of the nitric oxide donor isosorbide mononitrate for cervical ripening at term: a randomized controlled study. Am J Obstet Gynecol 2003; 189: 1692–7
- Li C. F., Chan C. W., Ho P. C. A comparison of isosorbide mononitrate and misoprostol cervical ripening before suction evacuation. Obstet Gynecol 2003; 102: 583–8
- Li C. F., Chan C. W., Ho P. C. A study of the efficacy of cervical ripening with nitric oxide donor versus placebo for cervical priming before second‐trimester termination of pregnancy. Contraception 2003; 68: 269–72
- Eppel W., Facchinetti F., Schleussner E., Piccinini F., Pizzi C., Gruber D. M., et al. Second trimester abortion using isosorbide mononitrate in addition to gemeprost compared with gemeprost alone: a double‐blind randomized, placebo‐controlled multicenter trial. Am J Obstet Gynecol 2005; 192: 856–61
- David M., Chen F. C. Comparison of isosorbide mononitrate (Mono Mack) and misoprostol (Cytotec) for cervical ripening in the first trimester missed abortion. Arch Gynecol Obstet 2005; 273: 144–5
- Wolfler M. M., Facchinetti F., Venturini P., Huber A., Helmer H., Husslein P., et al. Induction of labor at term using isosorbide mononitrate simultaneously with dinoprostone compared to dinoprostone treatment alone: A randomized, controlled trial. Am J Obstet Gynecol 2006; 195: 1617–22
- Osman I., MacKenzie F., Norrie J., Murray H. M., Greer I. A., Norman J. E. The ‘PRIM’ study: a randomized comparison of prostaglandin E2 gel with the nitric oxide donor isosorbide mononitrate for cervical ripening before the induction of labor at term. Am J Obstet Gynecol 2006; 194: 1012–21
- Facchinetti F., Piccinini F., Volpe A. Chemical ripening of the cervix with intracervical application of sodium nitroprusside: a randomized controlled trial. Hum Reprod 2000; 15: 2224–7
- Chan C. C., Tang O. S., Ng E. H., Li C. F., Ho P. C. Intracervical sodium nitroprusside versus vaginal misoprostol in first trimester surgical termination of pregnancy: a randomized double‐blinded controlled trial. Hum Reprod 2005; 20: 829–33
- Chanrachakul B., Herabutya Y., Punyavachira P. Randomized comparison of glyceryl trinitrate and prostaglandin E2 for cervical ripening at term. Obstet Gynecol 2000; 96: 549–53
- Sharma Y., Kumar S., Mittal S., Misra R., Dadhwal V. Evaluation of glyceryl trinitrate, misoprostol, and prostaglandin E gel for preinduction cervical ripening in term pregnancy. J Obstet Gynaecol Res 2005; 31: 210–5
- Nunes F. P., Campos A. P., Pedroso S. R., Leite C. F., Avillez T. P., Rodrigues R. D., et al. Intravaginal glyceryl trinitrate and dinoprostone for cervical ripening and induction of labor. Am J Obstet Gynecol 2006; 194: 1022–6
- Arteaga‐Troncoso G., Villegas‐Alvarado A., Belmont‐Gomez A., Martinez‐Herrera F. J., Villagrana‐Zesati R., Guerra‐Infante F. Intracervical application of the nitric oxide donor isosorbide dinitrate for induction of cervical ripening: a randomised controlled trial to determine clinical efficacy and safety prior to first trimester surgical evacuation of retained products of conception. BJOG 2005; 112: 1615–9
- de Pace V., Chiossi G., Facchinetti F. Clinical use of nitric oxide donors and L‐arginine in obstetrics. J Matern Fetal Neonatal Med 2007; 20: 569–79
- Piccinini F., Fano R. A., Volpe A., Facchinetti F. Ripening of the cervix with sodium nitroprusside in nonpregnant women. J Soc Gynecol Investig 2003; 10: 438–42
- Bullarbo M., Norstrom A., Andersch B., Ekerhovd E. Isosorbide mononitrate induces increased cervical expression of cyclooxygenase‐2, but not of cyclooxygenase‐1, at term. Eur J Obstet Gynecol Reprod Biol 2007; 130: 160–4
- Cacciatore B., Halmesmaki E., Kaaja R., Teramo K., Ylikorkala O. Effects of transdermal nitroglycerin on impedance to flow in the uterine, umbilical, and fetal middle cerebral arteries in pregnancies complicated by preeclampsia and intrauterine growth retardation. Am J Obstet Gynecol 1998; 179: 140–5
- Bates C. D., Nicoll A. E., Mullen A. B., Mackenzie F., Thomson A. J., Norman J. E. Serum profile of isosorbide mononitrate after vaginal administration in the third trimester. BJOG 2003; 110: 64–7
- Kahler C., Schleussner E., Moller A., Seewald H. J. Nitric oxide donors: effects on fetoplacental blood flow. Eur J Obstet Gynecol Reprod Biol 2004; 115: 10–4
- Radulovic N., Norström A., Ekerhovd E. Outpatient cervical ripening before first‐trimester surgical abortion: a comparison between Misoprostol and isosorbide mononitrate. Acta Obstet Gynecol Scand 2007; 86: 344–8
- Ledingham M. A., Thomson A. J., Lunan C. B., Greer I. A., Norman J. E. A comparison of isosorbide mononitrate, misoprostol and combination therapy for first trimester pre‐operative cervical ripening: a randomised controlled trial. BJOG 2001; 108: 276–80
- Maul H., Maner W. L., Saade G. R., Garfield R. E. The physiology of uterine contractions. Clin Perinatol 2003; 30: 665–76
- Condous G., Okaro E., Bourne T. The conservative management of early pregnancy complications: a review of the literature. Ultrasound Obstet Gynecol 2003; 22: 420–30
- Goepfert A. R., Goldenberg R. L., Mercer B., Iams J., Meis P., Moawad A., et al. The preterm prediction study: quantitative fetal fibronectin values and the prediction of spontaneous preterm birth. The National Institute of Child Health and Human Development Maternal‐Fetal Medicine Units Network. Am J Obstet Gynecol 2000; 183: 1480–3
- Ascarelli M. H., Morrison J. C. Use of fetal fibronectin in clinical practice. Obstet Gynecol Surv 1997; 52: S1–12
- Kekki M., Kurki T., Karkkainen T., Hiilesmaa V., Paavonen J., Rutanen E. M. Insulin‐like growth factor‐binding protein‐1 in cervical secretion as a predictor of preterm delivery. Acta Obstet Gynecol Scand 2001; 80: 546–51
- Iams J. D. Prediction and early detection of preterm labor. Obstet Gynecol 2003; 101: 402–12
- Honest H., Bachmann L. M., Coomarasamy A., Gupta J. K., Kleijnen J., Khan K. S. Accuracy of cervical transvaginal sonography in predicting preterm birth: a systematic review. Ultrasound Obstet Gynecol 2003; 22: 305–22
