
Abstract
The gastrointestinal tract, due to its role as a digestive organ and as a barrier between the exterior and interior milieus, is critically impacted by dietary, environmental, and inflammatory conditions that influence health and lifespan. Work in flies is now uncovering the multifaceted molecular mechanisms that control homeostasis in this tissue, and establishing its central role in health and lifespan of metazoans. The Drosophila intestine has thus emerged as a productive, genetically accessible model to study various aspects of the pathophysiology of aging. Studies in flies have characterized the maintenance of regenerative homeostasis, the development of immune senescence, the loss of epithelial barrier function, the decline in metabolic homeostasis, as well as the maintenance of epithelial diversity in this tissue. Due to its fundamental similarity to vertebrate intestines, it can be anticipated that findings obtained in this system will have important implications for our understanding of age-related changes in the human intestine. Here, I review recent studies exploring age-related changes in the fly intestine, and their insight into the regulation of health and lifespan of the animal.
Introduction
The intestine of aging animals suffers structural and functional decline (Kirkwood Citation2004; Biteau et al. Citation2008, Citation2010; Choi et al. Citation2008; Park et al. Citation2009; Hochmuth et al. Citation2011; Karpac et al. Citation2013; Rera et al. Citation2013, Citation2011, Citation2012). This decline limits the health of the animal, and it has been established in Drosophila that the intestine represents an important target organ with respect to genetic interventions that promote longevity (Biteau et al. Citation2010; Guo et al. Citation2014; Rera et al. Citation2013). These findings suggest that maintaining intestinal integrity is an important determinant of health and viability at the organismal level. Studies in flies have further established that the age-related decline of the intestine is caused by perturbations in a range of critical intestinal functions: (i) regeneration, (ii) barrier function, (iii) immune homeostasis, (iv) metabolism, and (v) epithelial compartmentalization. The experimental tools available in flies have allowed dissecting the molecular mechanisms causing these perturbations. An integrated model for the age-related functional decline and for the systemic consequences of this decline is thus starting to emerge. In the following, I will review this model and discuss recent findings.
Although mammals and Drosophila develop as deuterostomes and protostomes, respectively, their adult digestive tracts are structurally and functionally analogous (Pitsouli et al. Citation2009). Signaling mechanisms that control epithelial regeneration, innate immunity, and inflammation, as well as host–commensal interactions in the intestine are evolutionarily conserved (Apidianakis et al. Citation2009; Buchon, Broderick, Chakrabarti, et al. Citation2009; Colotta et al. Citation2009; Cronin et al. Citation2009; Jiang et al. Citation2009; Lee et al. Citation2009; Garrett et al. Citation2010; Grivennikov et al. Citation2010; Biteau et al. Citation2011). The Drosophila intestine thus provides a genetically accessible model that allows investigating the maintenance of homeostasis in aging gastrointestinal tracts mechanistically. It has a number of significant advantages over mammalian models: (i) ease and speed of genetic analysis, (ii) a lack of adaptive immunity, simplifying direct readouts of commensal-induced inflammation, (iii) naturally occurring, age-related intestinal dysplasia (Biteau et al. Citation2008), (iv) a relatively defined microflora, recent studies revealed only about 10 major microbial phylotypes in the fly’s gut (Ryu et al. Citation2008), and (v) short lifespan. Comprehensive studies aimed at integrating age-related changes in regeneration, immune homeostasis, size, and composition of the commensal flora, metabolic function, and intestinal barrier function, as well as at understanding the impact of these changes on overall lifespan, are thus possible for the first time.
Age-related decline of epithelial regeneration
The intestinal epithelium is a high-turnover tissue in both flies and mammals. It undergoes constant renewal and regeneration, fueled by a dedicated population of intestinal stem cells (ISCs). At the same time, the proliferative activity of ISCs is highly plastic, responding to stress and nutritional cues to preserve epithelial integrity and to adjust epithelial size to changing dietary conditions.
Aging is accompanied by a loss of proliferative homeostasis and of regenerative capacity in high-turnover tissues, including in the intestinal epithelium. Accordingly, as populations’ age, the incidence of inflammatory bowel diseases (IBDs) and gastrointestinal cancers is increasing (Lanzoni et al. Citation2008). To design rational therapies for these diseases, a better understanding of the causes and consequences of the age-related loss of proliferative homeostasis in intestinal epithelia is required.
Renewal of the Drosophila intestinal epithelium mechanistically and morphologically resembles cell turnover in the intestinal epithelium of mammals (Micchelli & Perrimon Citation2006; Ohlstein & Spradling Citation2006, Citation2007; Radtke et al. Citation2006; Biteau et al. Citation2011). ISCs are the only dividing cells in the intestinal epithelium and can give rise to at least two differentiated intestinal cell types: enteroendocrine cells (EEs) and enterocytes (ECs) (Micchelli & Perrimon Citation2006; Ohlstein & Spradling Citation2006, Citation2007). This lineage is critical for normal tissue turnover, as well as for epithelial recovery after damage or infection (Buchon, Broderick, Chakrabarti, et al. Citation2009; Buchon, Broderick, Poidevin, et al. Citation2009; Jiang et al. Citation2009). In the aging intestine, deregulation of ISC proliferation results in epithelial dysplasia. ISC proliferation strongly increases, and polyploid, mis-differentiated cells accumulate in the epithelium, disrupting its apico-basal organization, and perturbing epithelial function (Biteau et al. Citation2008, Citation2010; Buchon, Broderick, Chakrabarti, et al. Citation2009; Choi et al. Citation2008). Intestinal dysplasia is accompanied by metabolic decline, loss of barrier function, systemic infection, and ultimately death of the animal (Biteau et al. Citation2010; Rera et al. Citation2011, Citation2012; Karpac et al. Citation2013). Accordingly, limiting the rate of ISC proliferation in the aging intestine is sufficient to extend lifespan (Biteau et al. Citation2010; Rera et al. Citation2011; Guo et al. Citation2014).
ISC proliferation rates are regulated by a number of stress and growth signaling pathways, including the Jun-N-terminal Kinase (JNK), Jak/Stat, p38 MAPK, and EGFR pathways that integrate various local and systemic signals to control regenerative responses (Biteau et al. Citation2008; Amcheslavsky et al. Citation2009; Apidianakis et al. Citation2009; Buchon, Broderick, Chakrabarti, et al. Citation2009; Buchon, Broderick, Poidevin, et al. Citation2009; Chatterjee & Ip Citation2009; Jiang et al. Citation2009; Biteau et al. Citation2010; Buchon et al. Citation2010; Biteau & Jasper Citation2011; Jiang et al. Citation2011, reviewed in Biteau & Jasper Citation2011; Buchon, Broderick, et al. Citation2013). The age-related decline of proliferative homeostasis seems to be fundamentally a consequence of deregulation of the interaction between the intestinal epithelium and the commensal bacterial population, resulting in commensal dysbiosis and a condition that is reminiscent of a general inflammatory state of the epithelium that chronically and excessively stimulates proliferation of ISCs (Guo et al. Citation2014).
Immune homeostasis and host/commensal interactions
In a variety of model systems, loss of tissue homeostasis and cancer progression is associated with inflammation (Mantovani Citation2005, Citation2009; Colotta et al. Citation2009; Grivennikov et al. Citation2010). This association is particularly significant in barrier epithelia, due to the frequent and required interaction between epithelial cells (which mount innate immune responses) and the commensal microflora, but also with pathogenic microorganisms. In humans, changes in commensal populations (‘dysbiosis’) are not only associated with disorders like IBD, autoimmune and allergic diseases, obesity, and diabetes (Clemente et al. Citation2012), but also chronic inflammation and cancer (Uronis et al. Citation2009; Kaser et al. Citation2010; Claesson et al. Citation2011; Claesson et al. Citation2012). It has therefore been proposed that manipulating host/commensal interactions might be a viable avenue to promote healthy aging (Ottaviani et al. Citation2011; Biagi et al. Citation2013), yet better mechanistic insight into the interactions between the microflora, innate immune responses, and regenerative processes in the intestinal epithelium is needed (Garrett et al. Citation2010; Clemente et al. Citation2012; Holmes et al. Citation2012).
The Drosophila intestine is a productive model to explore these interactions (Buchon, Broderick, et al. Citation2013; Chambers & Schneider Citation2012). In young flies, commensal bacteria maintain basal activation of JNK and Jak/Stat activities in ISCs, ensuring low levels of epithelial renewal (Buchon, Broderick, Chakrabarti, et al. Citation2009). In older flies, however, the number of micro-organisms found in the lumen of the gut significantly increases (Ren et al. Citation2007). This dysbiosis chronically activates stress signaling pathways, inducing the epithelial dysplasia described above. Accordingly, axenically aged flies have significantly fewer mitotic ISCs than conventionally reared animals of the same age, and show a significant delay in the development of age-associated dysplasia (Buchon, Broderick, Chakrabarti, et al. Citation2009; Guo et al. Citation2014). An age-related impairment in the ability to manage the intestinal microflora thus appears to be the underlying cause of intestinal dysplasia, limiting lifespan (Guo et al. Citation2014) (Figure ).
Figure 1. Studying the pathophysiology of aging in the Drosophila intestine.
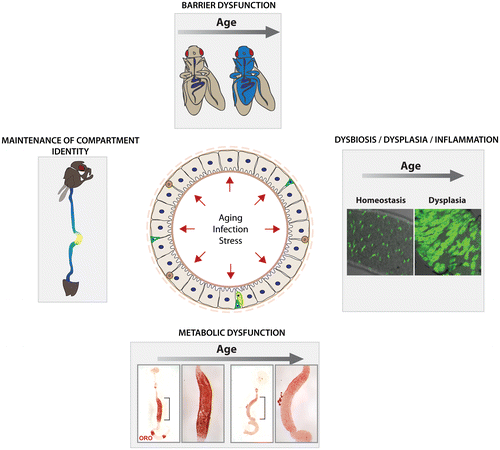
Management of the commensal flora and innate immune responses to pathogens are achieved primarily by two strategies acting in Drosophila ECs (Leulier & Royet Citation2009): (1) expression and activation of Dual Oxidase (Duox), which initiates an oxidative burst response, producing high levels of reactive oxygen species (ROS) (Ha et al. Citation2005; Ha, Lee, Park, et al. Citation2009; Lee, Seo, et al. Citation2009); and (2) activation of the immune deficiency (IMD/Relish) pathway, which activates the NFkB-like transcription factor Relish (Rel) (Buchon, Broderick, Chakrabarti, et al. Citation2009; Leulier & Royet Citation2009). Relish induces expression of antimicrobial peptides (AMPs) (Kim & Kim Citation2005). In aging intestines, the Rel/NFkB signaling pathway becomes chronically and excessively activated. Chronic activation of IMD/Relish signaling in the gut can sensitize flies to bacterial infection and shorten lifespan (Maillet et al. Citation2008; Bonnay et al. Citation2013), and in aging flies, this hyperactivation causes immune senescence and the described commensal dysbiosis (Guo et al. Citation2014). Dysbiosis, in turn, triggers the Duox-mediated ROS response, which promotes ISC over-proliferation, presumably by activating JNK signaling and by inhibiting the Nrf2 homologue CncC in ISCs (Libert et al. Citation2006; Ren et al. Citation2007; Hochmuth et al. Citation2011; Guo et al. Citation2014).
In a recent study, we have shown that this age-related deregulation of IMD/Relish signaling is caused by Foxo-mediated repression of peptidoglycan recognition proteins (PGRPs) of the SC class (Guo et al. Citation2014). PGRP-SCs are negative regulators of the IMD/Rel pathway, and over-expressing PGRP-SC2 in ECs accordingly prevents age-associated dysbiosis and dysplasia, and extends lifespan in conventionally, but not in axenically reared animals. These findings suggest that a breakdown of host/commensal interactions is central to the age-associated tissue degeneration in barrier epithelia, and identify strategies to promote tissue homeostasis by improving these interactions, thus extending lifespan (Guo et al. Citation2014). In mammals, deregulation of NFkB/AMP signaling has been associated with the pathogenesis of IBDs, suggesting that the mechanisms driving the loss of tissue homeostasis are conserved, and that modulating NFkB signaling to preserve homeostasis in the aging organism may be a promising strategy (Karin et al. Citation2006; Nenci et al. Citation2007; Xavier & Podolsky Citation2007).
The relationship between bacterial load and longevity is complex, however. In flies, commensal bacteria can have both beneficial and deleterious consequences, and completely eliminating pathogenic and commensal bacteria does not lead to lifespan extension (Brummel et al. Citation2004; Ren et al. Citation2007; Shin et al. Citation2011; Storelli et al. Citation2011). Our recent study suggests, however, that lifespan can be extended, when, rather than eliminating all bacteria from the gut, innate immune homeostasis is improved and the gut microbiota is maintained in a youthful state (Guo et al. Citation2014). The fact that PGRP-SC2 over-expression fails to extend lifespan in axenic conditions supports this interpretation.
Age-related changes in digestive function and the decline of metabolic homeostasis
Aging is associated with a loss of metabolic homeostasis, commonly caused by the deregulation of adaptive signaling interactions that coordinate energy metabolism with dietary changes (Barzilai et al. Citation2012). The mechanisms driving age-related changes in these adaptive responses remain largely unclear. Of note, the development of epithelial dysplasia is associated with a decline in the expression of trypsin genes in the gut, as well as with a general decline in energy stores (glycogen and triglycerides). Preventing dysplasia by limiting ISC proliferation rates is sufficient to restore trypsin expression, and to prevent the age-associated decline in energy stores (Biteau et al. Citation2010). Recently, we have further characterized the age-related deregulation of an adaptive metabolic response in the intestine, and the associated development of metabolic dysfunction (Karpac et al. Citation2013). Strikingly, the age-related chronic activation of Foxo in ECs that promotes innate immune senescence also perturbs the digestive function of the intestine: In young flies, activation of Foxo in ECs is required to inhibit the expression of evolutionarily conserved lipases as part of a metabolic response to dietary changes. This adaptive mechanism becomes chronically activated in the aging intestine, mediated (in part) by changes in JNK signaling. Age-related chronic JNK/Foxo activation in enterocytes is deleterious, leading to sustained repression of intestinal lipase expression and the disruption of systemic lipid homeostasis. As in the innate immune response, changes in the regulation of Foxo-mediated adaptive responses, thus contribute to the age-associated breakdown of metabolic homeostasis. The reasons for the age-associated chronic activation of Foxo remain unclear, however, and are interesting subjects for further study.
In addition to changes in the digestive function of the gut, changes in cellular metabolism of intestinal epithelial cells also play an important role in the age-related decline of this tissue. Regulating mitochondrial activity may therefore be a strategy to delay the onset of pathology and extend healthspan. In the fly intestine, this prediction has been tested using the Drosophila PGC-1 homolog (dPGC-1/spargel), a key regulator of mitochondrial energy metabolism (Rera et al. Citation2011). Strikingly, over-expression of dPGC-1 in ISC/EBs delays the onset of intestinal dysplasia and extends lifespan, suggesting that stimulating mitochondrial activity in specific cells of barrier epithelia is indeed an effective strategy to delay tissue aging and promote homeostasis.
Age-related changes in barrier function
Changes in intestinal barrier function have been reported in aging vertebrates (Katz et al. Citation1987), and a growing number of disorders have been linked to these changes, including inflammatory disorders (Farhadi et al. Citation2003; Fasano & Shea-Donohue Citation2005), multiple sclerosis (Yacyshyn et al. Citation1996), chronic heart failure (Sandek et al. Citation2008), cancer (Lin et al. Citation2012; Ullman & Itzkowitz Citation2011), and Parkinson’s disease (Forsyth et al. Citation2011). Intestinal barrier dysfunction is further common in critically ill patients and has been linked to development of the multiple organ dysfunction syndrome (Doig et al. Citation1998; Fink & Delude Citation2005; Harris et al. Citation1992). These observations suggest that barrier dysfunction plays a critical role in the etiology and/or mortality associated with many age-related diseases.
Recently, a noninvasive assay to determine intestinal integrity in individual flies has been developed that allows testing this hypothesis (Rera et al. Citation2011). Using this assay, it was shown that loss of intestinal integrity accompanies aging across a range of Drosophila genotypes and environmental conditions, and that interventions that extend lifespan (such as reduced temperature or dietary restriction) delay the onset of intestinal barrier defects, whereas interventions that shorten lifespan (such as loss of subunit b of mitochondrial complex II (sdhB)) accelerate the onset of intestinal barrier defects. Strikingly, intestinal barrier dysfunction is a better predictor of age-onset mortality than chronological age (Rera et al. Citation2012).
Flies with intestinal barrier dysfunction display increased expression of AMPs, impaired IIS and reduced metabolic stores compared with age-matched animals without intestinal barrier defects (Rera et al. Citation2012). These findings suggest that a consequence of the loss of barrier function is systemic infection, yet the exact mechanisms causing mortality in response to barrier breakdown have still to be investigated.
Using the Drosophila intestine to understand the maintenance of epithelial compartmentalization
The high rate of tissue regeneration along the gastrointestinal tract requires that mechanisms that ensure the functional diversity of newly formed intestinal cells are sustained throughout the lifespan of the organism. Such mechanisms are poorly understood, yet are likely to include short-range signaling interactions and cell autonomous cues that maintain diverse stem cell identities, as well as long-range signals that impart positional information along the gastrointestinal tract (Barker et al. Citation2010).
Recent studies have begun using the fly intestine as a model for compartment maintenance and region-specific regenerative activity (Buchon, Osman, et al. Citation2013; Li, Qi, et al. Citation2013; Marianes & Spradling Citation2013), and have suggested that compartment identities change in the aging intestinal epithelium, potentially perturbing midgut homeostasis (Buchon, Osman, et al. Citation2013). The midgut of flies can broadly be subdivided into the anterior midgut, the middle midgut, which contains an acidic gastric or copper cell region (CCR (Dubreuil Citation2004)), and the posterior midgut (PM). Finer subdivisions have been achieved by characterizing morphology and molecular makeup of intestinal epithelial cells in more detail (Buchon, Osman, et al. Citation2013; Marianes & Spradling Citation2013). ISCs can be found in each of these compartments (Hou Citation2010; Biteau et al. Citation2011; Strand & Micchelli Citation2011). ISCs in the PM express escargot (esg) and Delta (Dl), and divide asymmetrically to give rise to a precursor cell (the Dl-/esg + Enteroblast, EB), which will further differentiate into either Pdm – expressing Enterocytes (ECs) or prospero (pros) – expressing EEs (Micchelli & Perrimon Citation2006; Ohlstein & Spradling Citation2006, Citation2007). In the CCR, esg+ gastric stem cells (GSSCs) respond to stress by inducing regeneration of three different cell types: Dve+/Labial+/Cut+ Copper cells (CCs, which secrete hydrochloric acid), Dve+/weak Labial+/Cut− interstitial cells, and Pros+ endocrine cells (Strand & Micchelli Citation2011). The gastric region is maintained by a gradient of Dpp signaling activity, which is high in the gastric region and segregates GSSCs from PM ISCs (Guo et al. Citation2013; Li, Qi, et al. Citation2013).
Dpp is a Drosophila orthologue of Bmp2/4 (Affolter & Basler Citation2007), and the BMP signaling pathway has an evolutionarily conserved function in development and homeostasis of gastrointestinal epithelia (Haramis et al. Citation2004; Que et al. Citation2006; Jacobs et al. Citation2012). In the mouse, Bmp signaling establishes the esophageal epithelium (Jacobs et al. Citation2012), where Bmp-4 activity has been proposed to influence cell identities by regulating the expression of the homeotic gene Cdx (Que et al. Citation2006). In the mouse intestine, Bmp signaling controls differentiation of the secretory cell lineage (Auclair et al. Citation2007).
Similar to this role of Bmp signaling in vertebrates, Dpp is involved in multiple aspects of gastrointestinal development in Drosophila (Nakagoshi Citation2005; Mathur et al. Citation2010). Recent studies have further suggested that Dpp signaling promotes survival of ECs, recovery of ISCs into a quiescent state after injury, that it can influence ISC self-renewal, and that Dpp signaling is critical to establish regional stem cell identities during metamorphosis (Guo et al. Citation2013; Li, Zhang, et al. Citation2013; Driver & Ohlstein Citation2014; Tian & Jiang Citation2014). This multifaceted role of Dpp signaling in the regulation of intestinal regeneration is fascinating and will be of significant interest for future studies seeking to develop a comprehensive understanding of the role of Dpp in regulating intestinal homeostasis.
Concluding remarks
The Drosophila gut is rapidly emerging as the premier model system to characterize the integration of genetic and environmental conditions that influence health and lifespan. Due to its critical function as a semi-permeable barrier, its central role in the animal’s longevity, as well as its experimental accessibility, it can be anticipated that this organ will continue to provide critical insight into pathophysiological aspects of aging, as well as into new intervention strategies to extend lifespan.
Acknowledgments
Work in the author’s laboratory is supported by the National Institute on Aging (RO1 AG028127), the National Institute on General Medical Sciences (RO1 GM100196), and the National Eye Institute (R01 EY018177).
References
- Affolter M, Basler K. 2007. The Decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nature Reviews Genetics. 8:663–674.10.1038/nrg2166
- Amcheslavsky A, Jiang J, Ip YT. 2009. Tissue damage-induced intestinal stem cell division in Drosophila. Cell Stem Cell. 4:49–61.10.1016/j.stem.2008.10.016
- Apidianakis Y, Pitsouli C, Perrimon N, Rahme L. 2009. Synergy between bacterial infection and genetic predisposition in intestinal dysplasia. Proc Natl Acad Sci USA. 106:20883–20888.10.1073/pnas.0911797106
- Auclair BA, Benoit YD, Rivard N, Mishina Y, Perreault N. 2007. Bone morphogenetic protein signaling is essential for terminal differentiation of the intestinal secretory cell lineage. Gastroenterology. 133:887–896.10.1053/j.gastro.2007.06.066
- Barker N, Bartfeld S, Clevers H. 2010. Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell Stem Cell. 7:656–670.10.1016/j.stem.2010.11.016
- Barzilai N, Huffman DM, Muzumdar RH, Bartke A. 2012. The critical role of metabolic pathways in aging. Diabetes. 61:1315–1322.10.2337/db11-1300
- Biagi E, Candela M, Turroni S, Garagnani P, Franceschi C, Brigidi P. 2013. Ageing and gut microbes: Perspectives for health maintenance and longevity. Pharmacological Research. 69:11–20.10.1016/j.phrs.2012.10.005
- Biteau B, Jasper H. 2011. EGF signaling regulates the proliferation of intestinal stem cells in Drosophila. Development. 138:1045–1055.10.1242/dev.056671
- Biteau B, Hochmuth CE, Jasper H. 2008. JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell. 3:442–455.10.1016/j.stem.2008.07.024
- Biteau B, Karpac J, Supoyo S, DeGennaro M, Lehmann R, Jasper H. 2010. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genetics. 6:e1001159.10.1371/journal.pgen.1001159
- Biteau B, Hochmuth CE, Jasper H. 2011. Maintaining tissue homeostasis: dynamic control of somatic stem cell activity. Cell Stem Cell. 9:402–411.10.1016/j.stem.2011.10.004
- Bonnay F, Cohen-Berros E, Hoffmann M, Kim SY, Boulianne GL, Hoffmann JA, Matt N, Reichhart JM. 2013. big bang gene modulates gut immune tolerance in Drosophila. Proc Natl Acad Sci USA. 110:2957–2962.10.1073/pnas.1221910110
- Brummel T, Ching A, Seroude L, Simon AF, Benzer S. 2004. Drosophila lifespan enhancement by exogenous bacteria. Proc Natl Acad Sci USA. 101:12974–12979.10.1073/pnas.0405207101
- Buchon N, Broderick NA, Chakrabarti S, Lemaitre B. 2009a. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev. 23:2333–2344.
- Buchon N, Broderick NA, Poidevin M, Pradervand S, Lemaitre B. 2009b. Drosophila intestinal response to bacterial infection: activation of host defense and stem cell proliferation. Cell Host Microbe. 5:200–211.
- Buchon N, Broderick NA, Kuraishi T, Lemaitre B. 2010. Drosophila EGFR pathway coordinates stem cell proliferation and gut remodeling following infection. BMC Biol. 8:152.10.1186/1741-7007-8-152
- Buchon N, Broderick NA, Lemaitre B. 2013. Gut homeostasis in a microbial world: insights from Drosophila melanogaster. Nat Rev Microbiol. 11:615–626.10.1038/nrmicro3074
- Buchon N, Osman D, David FP, Yu Fang HY, Boquete JP, Deplancke B, Lemaitre B. 2013. Morphological and molecular characterization of adult midgut compartmentalization in Drosophila. Cell Rep. 3:1725–1738.10.1016/j.celrep.2013.04.001
- Chambers MC, Schneider DS. 2012. Pioneering immunology: insect style. Curr Opin Immunol. 24:10–14.10.1016/j.coi.2011.11.003
- Chatterjee M, Ip YT. 2009. Pathogenic stimulation of intestinal stem cell response in drosophila. J Cell Physiol. 220:664–671.10.1002/jcp.v220:3
- Choi NH, Kim JG, Yang DJ, Kim YS, Yoo MA. 2008. Age-related changes in Drosophila midgut are associated with PVF2, a PDGF/VEGF-like growth factor. Aging Cell. 7:318–334.10.1111/j.1474-9726.2008.00380.x
- Claesson MJ, Cusack S, O’Sullivan O, Greene-Diniz R, de Weerd H, Flannery E, Marchesi JR, Falush D, Dinan T, Fitzgerald G, et al. 2011. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc Natl Acad Sci USA. 108:4586–4591.10.1073/pnas.1000097107
- Claesson MJ, Jeffery IB, Conde S, Power SE, O’Connor EM, Cusack S, Harris HM, Coakley M, Lakshminarayanan B, O’Sullivan O, et al. 2012. Gut microbiota composition correlates with diet and health in the elderly. Nature. 488:178–184.10.1038/nature11319
- Clemente JC, Ursell LK, Parfrey LW, Knight R. 2012. The impact of the gut microbiota on human health: an integrative view. Cell. 148:1258–1270.10.1016/j.cell.2012.01.035
- Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A. 2009. Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis. 30:1073–1081.10.1093/carcin/bgp127
- Cronin SJ, Nehme NT, Limmer S, Liegeois S, Pospisilik JA, Schramek D, Leibbrandt A, Simoes RdM, Gruber S, Puc U, et al. 2009. Genome-wide RNAi screen identifies genes involved in intestinal pathogenic bacterial infection. Science. 325:340–343.10.1126/science.1173164
- Doig CJ, Sutherland LR, Dean Sandham JD, Fick GH, Verhoef M, Meddings JB. 1998. Increased intestinal permeability is associated with the development of multiple organ dysfunction syndrome in critically ill ICU patients. Am J Respir Crit Care Med. 158:444–451.10.1164/ajrccm.158.2.9710092
- Driver I, Ohlstein B. 2014. Specification of regional intestinal stem cell identity during Drosophila metamorphosis. Development. 141:1848–1856.10.1242/dev.104018
- Dubreuil RR. 2004. Copper cells and stomach acid secretion in the Drosophila midgut. Int J Biochem Cell Biol. 36:745–752.
- Farhadi A, Banan A, Fields J, Keshavarzian A. 2003. Intestinal barrier: an interface between health and disease. J Gastroenterol Hepatol. 18:479–497.10.1046/j.1440-1746.2003.03032.x
- Fasano A, Shea-Donohue T. 2005. Mechanisms of disease: the role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat Clin Pract Gastroenterol Hepatol. 2:416–422.10.1038/ncpgasthep0259
- Fink MP, Delude RL. 2005. Epithelial barrier dysfunction: a unifying theme to explain the pathogenesis of multiple organ dysfunction at the cellular level. Crit Care Clin. 21:177–196.10.1016/j.ccc.2005.01.005
- Forsyth CB, Shannon KM, Kordower JH, Voigt RM, Shaikh M, Jaglin JA, Estes JD, Dodiya HB, Keshavarzian A. 2011. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS One. 6:e28032.10.1371/journal.pone.0028032
- Garrett WS, Gordon JI, Glimcher LH. 2010. Homeostasis and inflammation in the intestine. Cell. 140:859–870.10.1016/j.cell.2010.01.023
- Grivennikov SI, Greten FR, Karin M. 2010. Immunity, inflammation, and cancer. Cell. 140:883–899.10.1016/j.cell.2010.01.025
- Guo Z, Driver I, Ohlstein B. 2013. Injury-induced BMP signaling negatively regulates Drosophila midgut homeostasis. J Cell Biol. 201:945–961.10.1083/jcb.201302049
- Guo L, Karpac J, Tran SL, Jasper H. 2014. PGRP-SC2 promotes gut immune homeostasis to limit commensal dysbiosis and extend lifespan. Cell. 156:109–122.10.1016/j.cell.2013.12.018
- Ha EM, Oh CT, Bae YS, Lee WJ. 2005. A direct role for dual oxidase in Drosophila gut immunity. Science. 310:847–850.10.1126/science.1117311
- Ha EM, Lee KA, Park SH, Kim SH, Nam HJ, Lee HY, Kang D, Lee WJ. 2009a. Regulation of DUOX by the Galphaq-phospholipase Cbeta-Ca2+ pathway in Drosophila gut immunity. Dev Cell. 16:386–397.10.1016/j.devcel.2008.12.015
- Ha EM, Lee KA, Seo YY, Kim SH, Lim JH, Oh BH, Kim J, Lee WJ. 2009b. Coordination of multiple dual oxidase-regulatory pathways in responses to commensal and infectious microbes in drosophila gut. Nat Immunol. 10:949–957.10.1038/ni.1765
- Haramis AP, Begthel H, van den Born M, van Es J, Jonkheer S, Offerhaus GJ, Clevers H. 2004. De novo crypt formation and juvenile polyposis on BMP inhibition in mouse intestine. Science. 303:1684–1686.10.1126/science.1093587
- Harris CE, Griffiths RD, Freestone N, Billington D, Atherton ST, Macmillan RR. 1992. Intestinal permeability in the critically ill. Intensive Care Med. 18:38–41.10.1007/BF01706424
- Hochmuth CE, Biteau B, Bohmann D, Jasper H. 2011. Redox regulation by Keap1 and Nrf2 controls intestinal stem cell proliferation in Drosophila. Cell Stem Cell. 8:1–12.
- Holmes E, Kinross J, Gibson GR, Burcelin R, Jia W, Pettersson S, Nicholson JK. 2012. Therapeutic modulation of microbiota-host metabolic interactions. Sci Transl Med. 4:137rv136.
- Hou SX. 2010. Intestinal stem cell asymmetric division in the Drosophila posterior midgut. J Cell Physiol. 224:581–584.10.1002/jcp.22194
- Jacobs IJ, Ku WY, Que J. 2012. Genetic and cellular mechanisms regulating anterior foregut and esophageal development. Dev Biol. 369:54–64.10.1016/j.ydbio.2012.06.016
- Jiang H, Patel PH, Kohlmaier A, Grenley MO, McEwen DG, Edgar BA. 2009. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell. 137:1343–1355.10.1016/j.cell.2009.05.014
- Jiang H, Grenley MO, Bravo MJ, Blumhagen RZ, Edgar BA. 2011. EGFR/Ras/MAPK signaling mediates adult midgut epithelial homeostasis and regeneration in Drosophila. Cell Stem Cell. 8:84–95.10.1016/j.stem.2010.11.026
- Karin M, Lawrence T, Nizet V. 2006. Innate immunity gone awry: linking microbial infections to chronic inflammation and cancer. Cell. 124:823–835.10.1016/j.cell.2006.02.016
- Karpac J, Biteau B, Jasper H. 2013. Misregulation of an adaptive metabolic response contributes to the agerelated disruption of lipid homeostasis in Drosophila. Cell Reports. 4:1250–1261.
- Kaser A, Zeissig S, Blumberg RS. 2010. Inflammatory bowel disease. Ann Rev Immunol. 28:573–621.10.1146/annurev-immunol-030409-101225
- Katz D, Hollander D, Said HM, Dadufalza V. 1987. Aging-associated increase in intestinal permeability to polyethylene glycol 900. Dig Dis Sci. 32:285–288.10.1007/BF01297055
- Kim T, Kim YJ. 2005. Overview of innate immunity in Drosophila. J Biochem Mol Biol. 38:121–127.10.5483/BMBRep.2005.38.2.121
- Kirkwood TB. 2004. Intrinsic ageing of gut epithelial stem cells. Mech Ageing Dev. 125:911–915.10.1016/j.mad.2004.09.004
- Lanzoni G, Roda G, Belluzzi A, Roda E, Bagnara GP. 2008. Inflammatory bowel disease: Moving toward a stem cell-based therapy. World J Gastroenterol. 14:4616–4626.10.3748/wjg.14.4616
- Lee WC, Beebe K, Sudmeier L, Micchelli CA. 2009. Adenomatous polyposis coli regulates Drosophila intestinal stem cell proliferation. Development. 136:2255–2264.10.1242/dev.035196
- Leulier F, Royet J. 2009. Maintaining immune homeostasis in fly gut. Nat Immunol. 10:936–938.10.1038/ni0909-936
- Li H, Qi Y, Jasper H. 2013. Dpp signaling determines regional stem cell identity in the regenerating adult Drosophila gastrointestinal tract. Cell Rep. 4:10–18.
- Li Z, Zhang Y, Han L, Shi L, Lin X. 2013. Trachea-derived dpp controls adult midgut homeostasis in Drosophila. Dev Cell. 24:133–143.10.1016/j.devcel.2012.12.010
- Libert S, Chao Y, Chu X, Pletcher SD. 2006. Trade-offs between longevity and pathogen resistance in Drosophila melanogaster are mediated by NFkappaB signaling. Aging Cell. 5:533–543.10.1111/ace.2006.5.issue-6
- Lin JE, Snook AE, Li P, Stoecker BA, Kim GW, Magee MS, Garcia AV, Valentino MA, Hyslop T, Schulz S, et al. 2012. GUCY2C opposes systemic genotoxic tumorigenesis by regulating AKT-dependent intestinal barrier integrity. PLoS One. 7:e31686.10.1371/journal.pone.0031686
- Maillet F, Bischoff V, Vignal C, Hoffmann J, Royet J. 2008. The Drosophila peptidoglycan recognition protein PGRP-LF blocks PGRP-LC and IMD/JNK pathway activation. Cell Host & Microbe. 3:293–303.
- Mantovani A. 2005. Cancer: inflammation by remote control. Nature. 435:752–753.10.1038/435752a
- Mantovani A. 2009. Cancer: inflaming metastasis. Nature. 457:36–37.10.1038/457036b
- Marianes A, Spradling AC. 2013. Physiological and stem cell compartmentalization within the Drosophila midgut. eLife. 2:e00886.
- Mathur D, Bost A, Driver I, Ohlstein B. 2010. A transient niche regulates the specification of Drosophila intestinal stem cells. Science. 327:210–213.10.1126/science.1181958
- Micchelli CA, Perrimon N. 2006. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature. 439:475–479.10.1038/nature04371
- Nakagoshi H. 2005. Functional specification in the Drosophila endoderm. Dev Growth Differ. 47:383–392.
- Nenci A, Becker C, Wullaert A, Gareus R, van Loo G, Danese S, Huth M, Nikolaev A, Neufert C, Madison B, et al. 2007. Epithelial NEMO links innate immunity to chronic intestinal inflammation. Nature. 446:557–561.10.1038/nature05698
- Ohlstein B, Spradling A. 2006. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature. 439:470–474.10.1038/nature04333
- Ohlstein B, Spradling A. 2007. Multipotent Drosophila intestinal stem cells specify daughter cell fates by differential notch signaling. Science. 315:988–992.10.1126/science.1136606
- Ottaviani E, Ventura N, Mandrioli M, Candela M, Franchini A, Franceschi C. 2011. Gut microbiota as a candidate for lifespan extension: an ecological/evolutionary perspective targeted on living organisms as metaorganisms. Biogerontology. 12:599–609.10.1007/s10522-011-9352-5
- Park JS, Kim YS, Yoo MA. 2009. The role of p38b MAPK in age-related modulation of intestinal stem cell proliferation and differentiation in Drosophila. Aging (Albany NY). 1:637–651.
- Pitsouli C, Apidianakis Y, Perrimon N. 2009. Homeostasis in infected epithelia: stem cells take the lead. Cell Host Microbe. 6:301–307.
- Que J, Choi M, Ziel JW, Klingensmith J, Hogan BL. 2006. Morphogenesis of the trachea and esophagus: current players and new roles for noggin and Bmps. Differentiation. 74:422–437.10.1111/dif.2006.74.issue-7
- Radtke F, Clevers H, Riccio O. 2006. From gut homeostasis to cancer. Curr Mol Med. 6:275–289.10.2174/156652406776894527
- Ren C, Webster P, Finkel SE, Tower J. 2007. Increased internal and external bacterial load during Drosophila aging without life-span trade-off. Cell Metab. 6:144–152.10.1016/j.cmet.2007.06.006
- Rera M, Bahadorani S, Cho J, Koehler CL, Ulgherait M, Hur JH, Ansari WS, Lo T Jr, Jones DL, Walker DW. 2011. Modulation of Longevity and Tissue Homeostasis by the Drosophila PGC-1 Homolog. Cell Metab. 14:623–634.10.1016/j.cmet.2011.09.013
- Rera M, Clark RI, Walker DW. 2012. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc Natl Acad Sci USA. 109:21528–21533.10.1073/pnas.1215849110
- Rera M, Azizi MJ, Walker DW. 2013. Organ-specific mediation of lifespan extension: more than a gut feeling? Ageing Res Rev. 12:436–444.10.1016/j.arr.2012.05.003
- Ryu JH, Kim SH, Lee HY, Bai JY, Nam YD, Bae JW, Lee DG, Shin SC, Ha EM, Lee WJ. 2008. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science. 319:777–782.10.1126/science.1149357
- Sandek A, Rauchhaus M, Anker SD, von Haehling S. 2008. The emerging role of the gut in chronic heart failure. Curr Opin Clin Nutr Metab Care. 11:632–639.10.1097/MCO.0b013e32830a4c6e
- Shin SC, Kim SH, You H, Kim B, Kim AC, Lee KA, Yoon JH, Ryu JH, Lee WJ. 2011. Drosophila microbiome modulates host developmental and metabolic homeostasis via insulin signaling. Science. 334:670–674.10.1126/science.1212782
- Storelli G, Defaye A, Erkosar B, Hols P, Royet J, Leulier F. 2011. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 14:403–414.10.1016/j.cmet.2011.07.012
- Strand M, Micchelli CA. 2011. Quiescent gastric stem cells maintain the adult Drosophila stomach. Proc Natl Acad Sci USA. 108:17696–17701.10.1073/pnas.1109794108
- Tian A, Jiang J. 2014. Intestinal epithelium-derived BMP controls stem cell self-renewal in Drosophila adult midgut. eLife. 3:e01857.
- Ullman TA, Itzkowitz SH. 2011. Intestinal inflammation and cancer. Gastroenterology. 140:1807–1816.10.1053/j.gastro.2011.01.057
- Uronis JM, Mühlbauer M, Herfarth HH, Rubinas TC, Jones GS, Jobin C. 2009. Modulation of the intestinal microbiota alters colitis-associated colorectal cancer susceptibility. PLoS One. 4:e6026.10.1371/journal.pone.0006026
- Xavier RJ, Podolsky DK. 2007. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 448:427–434.10.1038/nature06005
- Yacyshyn B, Meddings J, Sadowski D, Bowen-Yacyshyn MB. 1996. Multiple sclerosis patients have peripheral blood CD45RO+ B cells and increased intestinal permeability. Dig Dis Sci. 41:2493–2498.10.1007/BF02100148