
Abstract
Apart from within the Arumbera Sandstone that spans the Ediacaran–Cambrian boundary, the entire Neoproterozoic succession of the Amadeus Basin has been generally believed to be devoid of metazoan fossils, despite features of possible biogenic origin having been described in the earliest geological mapping reports. Herein are described a suite of curious, rounded impressions preserved upon the surface of a sandstone bed from the basal Neoproterozoic unit, the quartzitic Heavitree Formation that dates to ca 850 Ma. Compared with rounded structures of both organic and inorganic origin, these features resemble fossils of stranded medusae, both modern and ancient, and thereby add a potentially pelagic lifeform to previously described burrowing and sessile forms of likely metazoans, which are preserved as fossils/trace fossils within the Tonian period of the Neoproterozoic elsewhere in the basin. An organic origin for these features would support the contention that the Amadeus Basin harbours the earliest evidence that macroscopic life flourished, albeit briefly, some 215 million years prior to the start of the Ediacaran period and 250 million years before metazoans successfully colonised the late Ediacaran seas.
KEY POINTS
Curious rounded impressions upon a scree sample of Heavitree Formation (ca 850 Ma) are best compared with fossils of stranded jellyfish, both modern and ancient.
Such impressions add to the phyla seemingly extant within the Amadeus Basin during the Tonian Period (early Neoproterozoic).
Such phyla suggest that metazoan life evolved some 250 Ma earlier than generally believed, although became extinct locally some 10 Ma later.
Extinction occurred at ca 840 Ma when changing climate caused the enclosed Amadeus Basin to evolve from conducive shallow aquatic conditions to a harsh evaporitic salt pan.
Two later forays into metazoan evolution within the basin are evidenced by burrow-like tubular structures at ca 720 Ma and the Arumberia fossils at ca 560 Ma.
Introduction
Ediacaran fossils were first encountered in central Australia during commercial geological exploration near Mount Skinner in the southwestern Georgina Basin (). Discoidal in shape, the fossils were located within the Central Mount Stuart Formation, of late Ediacaran age, and although originally described by Wade (Citation1969) as representing two forms of medusae (Hallidaya brueri and Skinnera brooksi), Hall et al. (Citation2020) concluded them to be unique tri-radial metazoan genera, while Retallack and Broz (Citation2020) reinterpreted them as the upper and lower casts of the same feature, possibly colonial bacteria or fungi. Structures resembling Hallidaya brueri were also discovered within the lower Arumbera Sandstone, of latest Ediacaran age (), in the adjacent Amadeus Basin, as was a small Rangea fossil (see Glaessner, Citation1969, figure 9 A) and an association of “very common non-branching infilled endichnial worm burrows, which are more or less parallel to the bedding” (Glaessner, Citation1969, p. 389), later classified as Planolites (Glaessner, Citation1984) and concluded to be the oldest metazoan trace fossil in the Alice Springs region.
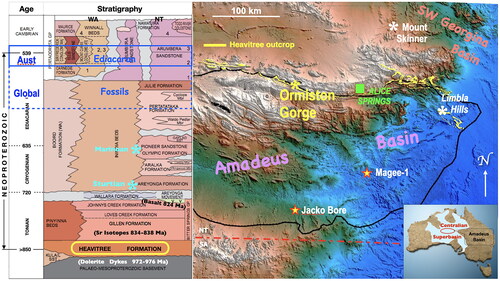
Figure 1. Eastern Amadeus Basin, showing Heavitree Formation outcrop (yellow) and locations discussed herein, along with stratigraphic column of the Neoproterozoic succession highlighting the Heavitree Formation. Tonian/Cryogenian boundary (from Halverson et al., Citation2020); blue stars mark the stratigraphic position of the two near global Cryogenian glaciations. The solid blue box encloses stratigraphic occurrences of Ediacaran fossils within the basin, extended by the dashed blue box to their generally accepted oldest age globally.
The basal member of the Arumbera Sandstone (dated to ca 560 Ma; Retallack & Broz, Citation2020) also revealed “peculiar grooves radiating from several centres” that Glaessner and Walter (Citation1975, p. 1) formally described as the remains of the cup-shaped body fossil Arumberia banksi. The similar morphology of Arumberia to microbially induced sedimentary structures, along with the outcome of various flume experiments, induced considerable discussion about their proposed biogenic origin. However, Allen and Haines (Citation2018) and Retallack and Broz (Citation2020) concluded Arumberia to indeed be biogenic, either cnidarian, metazoan or lichen, as they appear to be time-restricted and display several features distinct from the flume-derived features. Nevertheless, the debate continues, as Arumberia has most recently been described as “the remains of extinct, filamentous organisms (microbial or algal?) that occupied very shallow water and emergent environments across the globe” (McMahon et al., Citation2021, p. 1).
Tubular trace fossils with funnel-shaped upper ends were described by Walter (Citation1980) from the Elkera Formation, which underlies the Hallidaya/Skinnera-bearing Central Mount Stuart Formation, within the southwestern Georgina Basin. Although virtually identical to the walled burrow Laevicyclus, because of their ca 600 Ma age these burrows were concluded to be abiogenic fluid escape structures. Plummer (Citation2021), however, revisited this conclusion in the light of similar features discovered within two units of the Amadeus Basin that pre-date the Elkera Formation, proposing that the basin had witnessed at least two failed trials of metazoan evolution during the early and mid Neoproterozoic.
Geological setting of the Amadeus Basin
The Amadeus Basin is a vast intra-cratonic depocentre covering some 170 000 km2 of central Australia, largely within the southern reaches of the Northern Territory () but extending westwards into Western Australia. Initial deposition within the basin during the early Neoproterozoic post-dates dolerite dyke intrusions dated to between 972 and 976 Ma (baddeleyite and zircon dates from Wyborn et al., Citation1998) and continued with intermittent breaks into the middle Paleozoic. The Johnnys Creek Formation, comprising redbeds and dolostones that are separated from the basal Heavitree Formation by the evaporites and carbonates of the Gillen and Loves Creek formations, contains spilitic lavas that are dated to 824 Ma (Glikson et al., Citation1996), while 87Sr/86Sr isotopes from the Gillen Formation indicate ages of 838 and 834 Ma (Walter et al., Citation2000). Meanwhile, the contact between the Heavitree Formation and overlying Gillen Formation immediately south of Ormiston Gorge () is mappable as a conformable and transitional boundary, suggesting the depositional age for the basal quartzitic Heavitree Formation to be ca 850 Ma.
Heavitree Formation
The Heavitree Formation was described by Clark (Citation1974) as comprising four members, namely (from the base): locally developed interlaminated shales and siltstones (Undoolya Siltstone Member); fine- to medium-grained quartzose sandstones, locally conglomeratic at the base (Temple Bar Sandstone Member); then, above a scoured base, a series of thickly bedded pebble conglomerates to pebbly sandstones (Fenn Gap Conglomerate Member); and capped with fine- to medium-grained quartzose sandstones (Blatherskite Quartzite Member).
Although averaging between 200 and 300 m in thickness (Lindsay, Citation1999), the Heavitree Formation is thickest in the northeast corner of the basin where paleocurrent data indicate rivers, flowing from both the southeast and the north, merged before debouching their load to create a large delta exceeding 500 m thick (Plummer, Citation2015). Aside from the common presence of crossbeds and ripple marks, mudcracks of both desiccation and synaeresis origin also occur. Additionally, the presence of ‘possible invertebrate tracks’ was reported from three locations along 240 km of the north to northeastern margin of the basin (Wells et al., Citation1970), although no detailed descriptions or illustrations were provided.
The basin-wide arrangement of sedimentary facies that constitute the Heavitree Formation is not known because outcrops occur only in the northeast and southwest corners of the basin and along its northern margin. Where the Heavitree/basement contact is exposed in Heavitree Gap, just south of Alice Springs township, the basal unit comprises grey shales and siltstones (Undoolya Siltstone Member). Within the basin, the formation neither outcrops nor is marked by a discernible seismic event, and only two wells have penetrated basement (). In Magee-1, situated on the rising eastern flank of the basin but sitting off the deltaic pile, the Heavitree Formation comprises only 6.3 m of dark grey shale overlain by a predominantly medium-grained sandstone (Wakelin-King & Austin, Citation1992). In Jacko Bore (originally drilled as Mt Kitty-1), located in the south-central part of the basin, the Heavitree sandstone facies are absent, and dark grey argillaceous siltstones and dolomicrites of the basal Gillen Formation lie directly upon the granodiorite basement (Adderley, Citation2014). Similarly, the upper boundary of the Heavitree Formation is rarely exposed but outcrops immediately south of Ormiston Gorge where, within the uppermost several metres, limestone interbeds occur as the formation transitions conformably into the overlying carbonate/evaporite Gillen Formation.
The sandy facies of the Heavitree Formation may thus be limited to deltaic and shoreline deposits that are restricted to the margins of the basin and wedge-out or interfinger with grey shales and siltstones typical of more distal deposition, as at Heavitree Gap and in Magee-1. These in turn appear to be replaced in the centre of the basin by black shales and dolomicrites that constitute the basal Gillen Formation, as encountered in Jacko Bore. The Gillen Formation is typified by carbonates that are stromatolitic, as are the carbonates in the overlying Loves Creek, Johnnys Creek and Wallara formations, suggesting that, as stromatolites release free oxygen as a by-product (e.g. Des Marais, Citation1991), at times during the Tonian period the Amadeus Basin was likely a local ‘oxygen oasis’ conducive to the early development of metazoan life.
New evidence of pre-Ediacaran life?
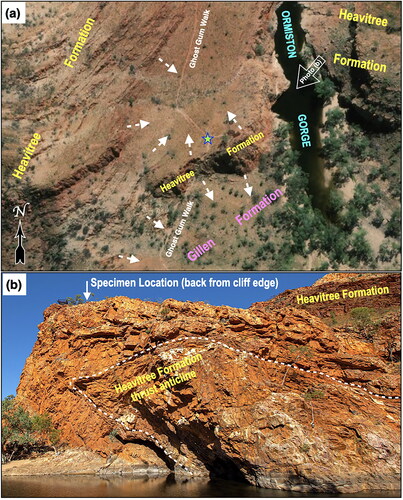
When the possibility of pre-Ediacaran life in the Amadeus Basin was recently mooted (Plummer, Citation2021, Citation2022), one of us (JDG) revisited a photograph taken in 2017 of a slab of Heavitree quartzite from adjacent to the pathway of the Ghost Gum Walk that crosses the Heavitree Formation on the western flank of Ormiston Gorge in the MacDonnell Ranges, 135 km west of Alice Springs ( and ). After scaling a thrust-faulted anticlinal wall of quartzitic Heavitree Formation from its southern start, the western arm of the Ghost Gum Walk crosses outcrop and/or scree of the formation that alone forms the imposing western rampart of Ormiston Gorge (). Although a scree sample, the slab in question is located upon the scree-slope composed entirely of quartzitic Heavitree Formation atop the outcropping thrust anticline () behind which looms a high quartzitic ridge composed solely of Heavitree Formation. As the surface of alternating outcrop and scree upon which the specimen lies is accessed only by detritus shed from Heavitree Formation outcrops (), the specimen cannot have been derived from any other formation.
Figure 2. (a) Satellite image (from open access www.mapsassist.com) showing the trackway of the Ghost Gum Walk adjacent to Ormiston Gorge cutting through (b) the thrust anticline of Heavitree Formation. Dashed arrows in (a) indicate surface dips showing that the specimen (blue star) can only have been derived from the Heavitree Formation, while the open white arrow indicates the viewpoint for the photograph of the Heavitree thrust anticline in (b).
However, being a scree sample resting upon a complexly faulted and folded Heavitree substrate, the stratigraphic position within the Heavitree sequence from which it was derived (whether Temple Bar Sandstone Member or Blatherskite Quartzite Member) cannot be determined. Nevertheless, the presence of associated ripple marks and desiccation cracks indicates that the sands forming the Heavitree Formation originated as part of a shoreline environment that covered the northern margin of the basin at ca 850 Ma. Although the specimen remains in the field, as it lies at a remote location within the Tjoritja/West Macdonnell National Park, is too large for easy hand carriage and borders a footpath (so requires replacing), negotiations are ongoing for its collection and removal. As a consequence of the sample remaining in the field, standard paleontological examination techniques, such as sectioning and geochemical or biomarker analysis, cannot be undertaken, so the following interpretations are, by necessity, comparative only. Nevertheless, when the sample is ultimately collected, while it remains alone and unique it will only be subjected to non-destructive analysis.
Fossilised onto the surface of the quartzitic slab are several rounded impressions, 3–5 cm in diameter and generally comprising a slightly raised outer, coronal rim surrounding an annular ‘skirt’ within which lies a slightly raised core (, and ). Rather than being a depression within a continuous sand layer, such as typifies subsidence after gas or minor fluid escape, the annular ‘skirt’ is bordered by a rim comprising a minor buildup of sand, while itself being devoid of the sand that forms the surrounding layer, suggesting that a foreign body was present to prevent deposition. Emanating from the raised central core of one of the impressions, which is asymmetric in shape, are weakly defined lineations that extend beyond the rim ().
Figure 3. Slab of Heavitree quartzite bearing several rounded structures with raised centres and rims impressed upon a stippled sandstone surface typical of algal mats. Impressions are indicated by green arrows (enlarged in ) and by the blue arrow (in ). Coin diameter: 3 cm. Diagonal dark bars are plant shadows.

Figure 4. (a) Close-up of two of the Heavitree Formation structures (green arrows on ) showing a central ‘body’ and annular ‘skirt’ upon a generally stippled surface (‘elephant-skin’ weathering) suggestive of a fossilised algal mat. (b) Comparison image of impressions left by modern jellyfish stranded on a Somerset, UK, beach in July Citation2022 (reproduced from BBC West under licence CC BY-NC-ND 4.0).

Figure 5. (a) Close-up of a flat sandstone surface of Heavitree Formation adjacent to Ormiston Gorge showing ‘elephant skin’ texture. (b) Close-up of a Heavitree Formation structure (blue arrow in ) impressed into a sandstone layer displaying ‘elephant skin’ texture: note the asymmetric central feature from which faint lineations emanate, suggestive of possible drag marks (the dark NE–SW bar across the centre is a shadow). (c) Modern stranded jellyfish: note drag lineations (Cordner, Citation2020, reproduced from Shields Gazette under licence CC BY-NC-ND 4.0). (d) Fossil of late Cambrian medusoid stranded upon a rippled, shoreface sandstone from central Wisconsin (Hagadorn et al., Citation2002, reproduced with written permission from the Geological Society of America).

The form of the impressions does not display characteristics typical of structures produced either inorganically by fluid escape, such as sand volcanoes, or organically by metazoan burrowing or resting activity, as, in the broken edge through one of the impressions (top left beneath coin), no vertical evacuation pipe-like structure is evident. Fluid escape structures are, however, common within the uppermost unit of the Heavitree Formation typically creating, in a nearby outcrop, sand volcanoes on various scales that result in an irregularly mounded bedding surface (detailed in Plummer & Gorter, Citation2024) rather than the generally flattish surface upon which the impressions are preserved.
As with other generally flat sandstone surfaces of Heavitree Formation, the overall bedding plane upon which the impressions are preserved displays a generally stippled appearance (‘elephant skin’ texture; and ) suggestive of agglutination of individual sand grains typical of algal or microbial mats that Gehling (Citation1986, Citation1999) and Slagter et al. (Citation2022), amongst others, conclude to be essential in preventing the sand from post-depositional erosion and thus enabling fossilisation. The general lack of predation and bioturbation during the Neoproterozoic also increased the probability of preservation and fossilisation of any soft-bodied metazoans.
The imprints are not concordant with the ‘elephant skin-textured’ algal/biomat surface, but rather disrupt it ( and ), and therefore are not a component of the biomat. Rather, the imprints are defined by an absence of the enclosing sediment layer, which effectively nullifies them from being fossilised algal or fungal colonies, such as those depicted by El Albani et al. (Citation2010), Glaessner (Citation1969) or Grazhdankin and Gerdes (Citation2007), or the Beltanelliformis of Bobrovskiy et al. (Citation2018) that are all typified by concentric buildups. Nor do the Heavitree impressions resemble either the mould or cast of the rhizome holdfast of macroalgae, as described by Wang et al. (Citation2020), as they lack all evidence of the characteristic filamentous rhizoids growing out of the central rhizome holdfast, specifically the Gemmaphyton-type holdfast Longfengshania that is reportedly of Tonian age. Furthermore, these various macroalgal holdfasts are preserved in muddy dolostones and black shales indicative of a quiescent depositional environment, whereas the Heavitree Formation impressions are preserved upon a high-energy shoreline sandstone likely to have been non-conducive for the growth, let alone preservation, of such delicate macroalgal structures.
The absence of sediment within the annular skirt of the Heavitree Formation imprints is not typical of Aspidella, an Ediacaran (and older) fossil form that has been the subject of numerous studies and reviews (Burzynski et al., Citation2017; Gehling et al., Citation2000; MacGabhann, Citation2007; amongst others), nor indeed of Aspidella fossils that occur in a nearby outcrop of the Blatherskite Quartzite Member, the upper unit of the Heavitree Formation described by Plummer and Gorter (Citation2024). Other rounded to circular Aspidella-like fossils include Triforillonia, a member of the Ediacaran Avalon assemblage in Newfoundland, and the temporally more widely distributed Nimbia, a discoidal fossil arguably of cnidarian or microbial origin. Triforillonia comprises an outline (less than 3.0 cm across) of three intersecting semicircles encasing a raised central ‘rosette’ that occupies the entire internal cavity (Gehling et al., Citation2000). Nimbia, on the other hand, is small (generally less than 1.5 cm diameter) and comprises a raised outer rim encasing a flat internal disc with or without a central tubercle (MacGabhann, Citation2007). Unlike Triforillonia and Nimbia, the Heavitree Formation impressions are essentially circular and considerably larger, and feature a flat annular ‘skirt’ between their raised coronal rim and raised core. Finally, the form of the Heavitree Formation impressions does not conform to the evacuation cavities of eroded diagenetic concretions or nodules (e.g. Biek, Citation2002; Hudson et al., Citation2023), nor do they display evidence of diagenetic ionic migration (absence of liesegang rings, etc), or to crystal clusters, whether syn- or post-sedimentary.
The imprints do, however, strongly resemble impressions of present-day stranded jellyfish ( and ) as well as fossil impressions of jellyfish stranded on Ediacaran (Srivastava, Citation2012) and Cambrian shorelines (; Hagadorn & Belt, Citation2008; Hagadorn et al., Citation2002; Sappenfield et al., Citation2017). The annular ‘skirt’ surrounding the raised asymmetric core, from which faint lineations emanate (), is interpreted to be where the umbrella-like bell and central body of a jellyfish came to rest, the presence of which prevented deposition of the enclosing sand layer and creating the overall outline. The faint lineations, which extend below the level of the overarching bell, are interpreted as linear drag marks scored onto the substrate during the final phase of stranding, comparable with that of recent jellyfish strandings (). According to Sappenfield et al. (Citation2017), stranded medusae typically have raised central cores, as seen in the Heavitree Formation examples, because, in an attempt to free themselves, they pump their bodies, which forces sand into their internal cavities, thereby sealing their fate.
Aside from the uncertainty of whether or not these impressions are of biogenic origin, it has been suggested that, because the exact stratigraphic position of the sample cannot be determined, the features impressed upon it have no value in a chrono-paleontological sense. We would argue, however, that with potential paleontological samples being so rare in the early to mid Neoproterozoic, the mere presence of any sample, be it in situ or scree, is of immense value. If many of the samples from the Ediacara Hills in South Australia described by Sprigg (Citation1947, Citation1949) had been dismissed because of their scree nature, the global geological community would not now be referring to a biota or a geological period bearing that name.
So, are there now four?
Although cognisant of the fact that many of the earliest fossil discoveries within the Neoproterozoic that were initially interpreted as medusae have subsequently been shown to be the holdfast attachments of frond-like creatures (i.e. Aspidella) with others reinterpreted as algal colonies, we are confident not only that these impressions within the Heavitree Formation are of organic origin but also that their characteristics most strongly resemble those of stranded medusae, both modern and ancient. If so, these features would take to four the number of different genera of likely fossils/trace fossils encountered within the quartzitic facies of the Heavitree Formation. From Limbla Hills in the northeast of the basin (), Lindsay (Citation1991) described vertical tubular structures and an enigmatic structure, both forms, he believed, being the result of the burrowing activity of metazoans. While concurring with a burrowing origin for the vertical tubular structures, Plummer (Citation2021) interpreted the enigmatic structure as the body fossil of possibly two superimposed cup-shaped carapaces of sessile metazoans that lived partially buried within the sediment. Additionally, several Aspidella, the holdfasts of frondose metazoans, and a possible horizontal burrow have also been reported from outcrops of the uppermost Heavitree Formation immediately south of Ormiston Gorge (Plummer & Gorter, Citation2024).
The above scenario would suggest, therefore, that burrowing metazoans, sessile (both cup-shaped and frondose) metazoans and pelagic metazoans—a veritable metazoan menagerie in the making—were seemingly all extant within the Amadeus Basin of central Australia at ca 850 Ma. This occurrence falls some 100 million years after the start of the predicted 950–800 Ma age range for metazoan evolution as determined at the molecular level (Petersen et al., Citation2004, 2007; Sperling & Stockey, Citation2018, ). Although the validity of the molecular clock is much debated (e.g. Budd & Mann, Citation2020), its timing is supported by Bezerra et al. (Citation2021), who concluded that the initial emergence within eukaryotes of aerobic enzymes critical for metazoan evolution was a Mesoproterozoic, perhaps even a late Paleoproterozoic, event. The ca 850 Ma age of these Amadeus Basin impressions also links well with other late Tonian fossils dated to ca 770 Ma in Kazakhstan (Meert et al., Citation2011) and central India (Choudhuri, Citation2023). However, this early flourish of metazoan life within the Amadeus Basin ultimately became a ‘failed evolutionary trial’ (Plummer, Citation2022) when the basin, an isolated intra-cratonic basin within the Rodinia Supercontinent, evolved from the conducive shallow aquatic setting of Heavitree Formation time into the extensive hot and arid evaporitic salt pan of Gillen Formation time, within which such early life, with no means to migrate to safer havens, could not survive. The next foray into metazoan within the Amadeus Basin occurred some 120 million years later, as evidenced by a suite of burrow-like tubular structures described in Plummer (Citation2021).
Conclusion
The Heavitree Formation, the basal unit within the Amadeus Basin of central Australia, conservatively dates to ca 850 Ma. Within its predominantly deltaic and shoreline sandstone facies, four distinct potential lifeforms interpreted as fossils or trace fossils have been identified some 280 km apart, namely vertical burrows, cup-shaped body fossils, frondose holdfasts and, as described herein, several possible medusae. Together, these fossils suggest that at least four metazoan genera were extant and active during the Tonian period of the Neoproterozoic era, some 215 million years before the start of the Ediacaran period and 250 million years earlier than the generally accepted oldest animal macrofossils. Changing environmental conditions throughout the intra-cratonic Amadeus Basin from shallow aquatic to arid terrestrial, however, terminated this early metazoan evolutionary flourish. Without doubt, this proposal that a macroscopic ‘biota’ of Tonian age thrived within the Amadeus Basin will be met with scepticism, but we are hopeful that it will stimulate further search globally for pre-Ediacaran life forms. Whatever the origin of these various features ultimately proves to be, however, will add to the overall debate on the timing of metazoan origins.
Acknowledgements
The authors wish to thank several anonymous reviewers whose various comments improved the comprehensiveness of the paper.
Disclosure statement
No potential conflict of interest was reported by the authors.
Data availability statement
The authors confirm that the data supporting the findings of this study are within the article.
References
- Adderley, D. (2014). Mt Kitty 1 basic well completion report. Northern Territory Geological Survey Report PR2015-0010. Santos Ltd.
- Allen, H., & Haines, P. (2018). Arumberia banksii: How well can MISS tell time? [Paper presentation]. Australian Geoscience Council Convention: Big Issues and Ideas in Geoscience, Abstracts, 290.
- BBC West. (2022). Warning as hundreds of jellyfish wash up on Somerset beaches. BBC News, 7 July 2022.
- Bezerra, B. S., Belato, F. A., Mello, B., Brown, F., Coates, C. J., de Moraes Leme, J., Trindade, R. I. F., & Costa-Paiva, E. M. (2021). Evolution of a key enzyme of aerobic metabolism reveals Proterozoic functional subunit duplication events and an ancient origin of animals. Scientific Reports, 11(1), 15744. https://doi.org/10.1038/s41598-021-95094-4
- Biek, B. (2002). Concretions and nodules in North Dakota. North Dakota Geological Survey. Retrieved from https://www.dmr.nd.gov/ndgs/ndnotes/concretions/concretions.asp
- Bobrovskiy, I., Hope, J. M., Krasnova, A., Ivantsov, A., & Brocks, J. J. (2018). Molecular fossils from organically preserved Ediacara biota reveal cyanobacterial origin for Beltanelliformis. Nature Ecology & Evolution, 2(3), 437–440. https://doi.org/10.1038/s41559-017-0438-6
- Budd, G. E., & Mann, R. P. (2020). Survival and selection biases in early animal evolution and a source of systematic overestimation in molecular clocks. Interface Focus, 10(4), 20190110. https://doi.org/10.1098/rsfs.2019.0110
- Burzynski, G., Narbonne, G. M., Dececchi, T. A., & Dalrymple, R. W. (2017). The ins and outs of Ediacaran discs. Precambrian Research, 300, 246–260. https://doi.org/10.1016/j.precamres.2017.08.012
- Choudhuri, A. (2023). The advent of motile life in the Proterozoic: Scepticism and reality [Paper presentation]. Presentation & Abstract T3/O-03, Vindhyan Supergroup: Recent Advances, Challenges & Opportunities (VISACOP) Conference, Geological Survey of India, Special Publication, 128, 39.
- Clark, D. (1974). Heavitree Quartzite stratigraphy and structure near Alice Springs, N.T. Northern Territory Geological Survey Record, 74/9, 37 pp.
- Cordner, C. (2020). Beware of the jellyfish—and here’s what to do if you are stung by one. Shields Gazette, 7th August 2020.
- Des Marais, D. J. (1991). Microbial mats, stromatolites and the rise of oxygen in the Precambrian atmosphere. Global and Planetary Change, 5(1-2), 93–96. https://doi.org/10.1016/0921-8181(91)90130-O
- El Albani, A., Bengtson, S., Canfield, D. E., Bekker, A., Macchiarelli, R., Mazurier, A., Hammarlund, E. U., Boulvais, P., Dupuy, J-J., Fontaine, C., Fürsich, F. T., Gauthier-Lafaye, F., Janvier, P., Javaux, E., Ossa, F. O., Pierson-Wickmann, A-C., Riboulleau, A., Sardini, P., Vachard, D., Whitehouse, M., & Meunier, A. (2010). Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago. Nature Letters, 466(7302), 100–104. https://doi.org/10.1038/nature09166
- Gehling, J. G. (1986). Algal binding of siliciclastic sediments: A mechanism for the preservation of Ediacaran fossils [Paper presentation]. 12th International Sedimentological Congress, Abstracts, 117.
- Gehling, J. G. (1999). Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks. PALAIOS, 14(1), 40–57. https://doi.org/10.2307/3515360
- Gehling, J. G., Narbonne, G. M., & Anderson, M. M. (2000). The first named Ediacaran body fossil, Aspidella terranovica. Palaeontology, 43(3), 427–456. https://doi.org/10.1111/j.0031-0239.2000.00134.x
- Glaessner, M. F. (1969). Trace fossils from the Precambrian and basal Cambrian. Lethaia, 2(4), 369–393. https://doi.org/10.1111/j.1502-3931.1969.tb01258.x
- Glaessner, M. F. (1984). The Dawn of Animal Life: A Biohistorical Study. Cambridge Earth Science Series, Cambridge University Press, 256 pp.
- Glaessner, M. F., & Walter, M. R. (1975). New Precambrian fossils from the Amadeus Basin, Northern Territory, Australia. Bureau of Mineral Resources, Geology & Geophysics Record, 1975/17, 23.
- Glikson, A. Y., Stewart, A. J., Ballhaus, C. G., Clarke, G. L., Feeken, E. H. J., Leven, J. H., Sheraton, J. W., & Sun, S-S. (1996). Geology of the western Musgrave Block, central Australia, with particular reference to the mafic–ultramafic Giles Complex. Australian Geological Survey Organisation Bulletin, 239, 206.
- Grazhdankin, D., & Gerdes, G. (2007). Ediacaran microbial colonies. Lethaia, 40(3), 201–210. https://doi.org/10.1111/j.1502-3931.2007.00025.x
- Hagadorn, J. W., & Belt, E. S. (2008). Stranded in upstate New York: Cambrian Scyphomedusae from the Potsdam Sandstone. PALAIOS, 23(7), 424–441. https://doi.org/10.2110/palo.2006.p06-104r
- Hagadorn, J. W., Dott, R. H., Jr, & Damrow, D. (2002). Stranded on a Late Cambrian shoreline: Medusae from central Wisconsin. Geology, 30(2), 147–150. https://doi.org/10.1130/0091-7613(2002)030
- Hall, C. M. S., Droser, M. L., Clites, E. C., & Gehling, J. G. (2020). The short-lived but successful tri-radial body plan: A view from the Ediacaran of Australia. Australian Journal of Earth Sciences, 67(6), 885–895. https://doi.org/10.1080/08120099.2018.1472666
- Halverson, G., Porter, S., & Shields, G. (2020). Chapter 17: The Tonian and Cryogenian periods. In F. M. Gradstein, J. G. Ogg, M. D. Schmitz, & G. M. Ogg (Eds.), Geologic Time Scale 2020. Elsevier BV. https://doi.org/10.1016/C2020-1-02369-3
- Hudson, J. D., Paxton, R. B., Andrews, J. E., Dennis, P. F., & Marca, A. D. (2023). Classic Scottish sandstone concretions formed from hot water. Geology Today, 39(6), 231–235. https://doi.org/10.1111/gto.12455
- Lindsay, J. F. (1991). New evidence for ancient metazoan life in the Late Proterozoic Heavitree Quartzite, Amadeus Basin, central Australia. In R. J. Korsch & J. M. Kennard (Eds.), Geological and geophysical studies in the Amadeus Basin, Central Australia (pp. 91–95). Bureau of Mineral Resources, Geology and Geophysics. Bulletin 236.
- Lindsay, J. F. (1999). Heavitree Quartzite, a Neoproterozoic (ca 800–760 Ma), high-energy, tidally influenced, ramp association, Amadeus Basin, central Australia. Australian Journal of Earth Sciences, 46(1), 127–139. https://doi.org/10.1046/j.1440-0952.1999.00693.x
- MacGabhann, B. A. (2007). Discoidal fossils of the Ediacaran biota: A review of current understanding. In P. Vickers-Rich & P. Komarower (Eds.), The rise and fall of the Ediacaran biota (pp. 297–313). Geological Society of London. Special Publication 286. https://doi.org/10.1144/SP286.21
- McMahon, W. J., Davies, N. S., Liu, A. G., & Went, D. J. (2021). Enigma variations: Characteristics and likely origin of the problematic surface texture Arumberia, as recognised from an exceptional bedding plane exposure and the global record. Geological Magazine, 159(1), 1–20. https://doi.org/10.1017/S0016756821000777
- Meert, J. G., Gibsher, A. S., Levashova, N. M., Grice, W. C., Kamenov, G. D., & Ryabinin, A. B. (2011). Glaciation and ∼770 Ma Ediacara (?) fossils from the Lesser Karatau Microcontinent, Kazakhstan. Gondwana Research, 19(4), 867–880. https://doi.org/10.1016/j.gr.2010.11.008
- Peterson, K. J., Lyons, J. B., Nowak, K. S., Takacs, C. M., Wargo, M. J., & McPeek, M. A. (2004). Estimating metazoan divergence times with a molecular clock. Proceedings of the National Academy of Sciences of the United States of America, 101(17), 6536–6541. https://doi.org/10.1073/pnas.0401670101
- Peterson, K. J., Summons, R. E., & Donoghue, P. C. J. (2007). Molecular palaeobiology. Palaeontology, 50(4), 775–809. https://doi.org/10.1111/j.1475-4983.2007.00692.x
- Plummer, P. S. (2015). Heavitree Quartzite: Its place within the Centralian Superbasin [Paper presentation]. Annual Geoscience Exploration Seminar (AGES) 2015. Northern Territory Geological Survey Record, 2015-002 (pp. 83–91).
- Plummer, P. S. (2021). Was the Amadeus Basin of Central Australia a crucible for pre-Ediacaran macro-biotic evolutionary trials? Transactions of the Royal Society of South Australia, 145(2), 125–142. https://doi.org/10.1080/03721426.2021.1935585
- Plummer, P. S. (2022). Failed evolutionary trials: Should we expect to find fossil evidence? Academia Letters, Article 5212, 6. https://doi.org/10.20935/AL5212
- Plummer, P. S., & Gorter, J. D. (2024). Aspidella and water escape structures from the ca 850 Ma (Tonian) Heavitree Formation, Amadeus Basin, Central Australia. Australian Journal of Earth Sciences, 71(3), 361–369. https://doi.org/10.1080/08120099.2023.2290248
- Retallack, G. J., & Broz, A. P. (2020). Arumberia and other Ediacaran–Cambrian fossils of central Australia. Historical Biology, 33(10), 1964–1988. https://doi.org/10.1080/08912963.2020.1755281
- Sappenfield, A. D., Tarhan, L. G., & Droser, M. L. (2017). Earth’s oldest jellyfish strandings: A unique taphonomic window or just another day at the beach? Geological Magazine, 154(4), 859–874. https://doi.org/10.1017/S0016756816000443
- Slagter, S., Hao, W., Planavsky, N. J., Konhauser, K. O., & Tarhan, L. G. (2022). Biofilms as agents of Ediacara-style fossilisation. Scientific Reports, 12(1), 10. https://doi.org/10.1038/s41598-022-12473-1
- Sperling, E. A., & Stockey, R. G. (2018). The temporal and environmental context of early animal evolution: Considering all the ingredients of an “Explosion”. Integrative and Comparative Biology, 58(4), 605–622. https://doi.org/10.1093/icb/icy088
- Sprigg, R. G. (1947). Early Cambrian(?) jellyfishes from the Flinders Ranges, South Australia. Transactions of the Royal Society of South Australia, 71(2), 202–224.
- Sprigg, R. G. (1949). Early Cambrian jellyfishes of Ediacara, South Australia, and Mount John, Kimberley District, Western Australia. Transactions of the Royal Society of South Australia, 73(1), 72–96.
- Srivastava, P. (2012). Ediacaran discs from the Jodhpur sandstone, Marwar Supergroup, India: A biological diversification or taphonomic interplay? International Journal of Geosciences, 03(05), 1120–1126. https://doi.org/10.4236/ijg.2012.35113
- Wade, M. (1969). Medusae from uppermost Precambrian or Cambrian sandstones, central Australia. Palaeontology, 12(3), 351–365.
- Wakelin-King, G., & Austin, L. (1992). Magee 1: EP38, Northern Territory, well completion report. Pacific Oil & Gas Pty. Ltd. (unpublished)
- Walter, M. R. (1980). Adelaidean and Early Cambrian stratigraphy of the southwestern Georgina Basin: Correlation chart and explanatory notes. Bureau of Mineral Resources Australia Report, 214, 21.
- Walter, M. R., Veevers, J. J., Calver, C. R., Gorjan, P., & Hill, A. C. (2000). Dating the 840–544 Ma Neoproterozoic interval by isotopes of strontium, carbon, and sulfur in seawater, and some interpretative models. Precambrian Research, 100(1-3), 371–433. https://doi.org/10.1016/S0301-9268(99)00082-0
- Wang, Y., Wang, Y., Tang, F., & Zhao, M. (2020). Ediacaran macroalgal holdfasts and their evolution: A case study from China. Palaeontology, 63(5), 821–840. https://doi.org/10.1111/pala.12485
- Wells, A. T., Forman, D. J., Ranford, L. C., & Cook, P. J. (1970). Geology of the Amadeus Basin, central Australia. Bureau of Mineral Resources, Geology and Geophysics Bulletin, 100, 222.
- Wyborn, L., Hazell, M., Page, R., Idnurm, M., & Sun, S. (1998). A newly discovered major Proterozoic granite alteration system in the Mount Webb region, central Australia, and implications for Cu–Au mineralisation. Australian Geological Survey Organisation, Research Newsletter, 28, 1–6.