
Abstract
The cheek teeth of Debruijnia kostakii n. sp. from the Early Miocene of Karydia are only slightly smaller than those of the largest extant species of the Spalacinae (Spalax giganteus), but show characteristics that are reminiscent of its Paleogene cricetid ancestor. The peculiar combination of large size and primitive dental morphology of this species bridges the gap between Debruijnia and Pliospalax and thereby supports the hypothesis that these genera form the core of the complex evolutionary history leading to extant Spalax.
Introduction
The fossiliferous lacustrine deposit of Karydia was discovered by Mr. Dimitris Foussekis and the author during a prospection trip in the early nineties of the last century. Karydia 1 refers to the small test sample taken from the, at the time abandoned, quarry on the west side of Karydia Hill. Karydia 2 and 3 refer to lignitic clay beds at the top of two superposed sediment cycles exposed in the quarry on the east side of the hill. Lithostratigraphical correlation of the deposits exposed in these two quarries was impossible due to lack of exposure and the presence of faults. Samples of several tons of matrix from Karydia 2 were collected and wet-screened during three consecutive years. This means that the large spalacine that will be described below occurs at a rate of about four cheek teeth per ton of matrix.
The fossil record of the fossorial muroid rodents assembled in the subfamily Spalacinae can be traced to the Late Oligocene of south eastern Europe (De Bruijn et al. Citation2013; De Bruijn et al., Citation2015), but the group is probably much older. The evolutionary history of the mole-rats is largely based on isolated cheek teeth from south eastern European and Middle Eastern localities, that is to say, from the core of the area occupied by the extant representatives (Figure ). Although the number of sites that have yielded spalacine teeth is quite large the number of specimens per site is usually limited. This makes the evaluation of samples difficult because, in addition to the regular individual variation of details of the occlusal pattern, the morphology of the occlusal surface of these semi-hypsodont teeth changes as a result of attrition (De Bruijn et al., Citation2015).
Figure 1. Map of Europe, the Middle East and North Africa showing the geographical range of the extant Spalacinae and the position of the locality Karydia.
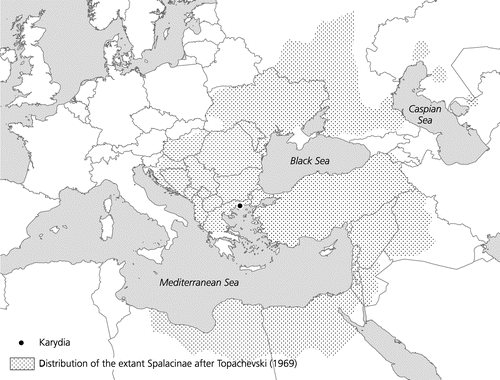
The similarity in overall dental architecture of all Spalacinae (rather high-crowned robust teeth with thick enamel, cusps incorporated into the lophs, metaconid of the m1 large and situated at the level of the anteroconid) and the limited geographical range occupied by its representatives through time suggests that it is a very closely knit clade. Representatives from different time-slices show a, seemingly random, mosaic-like combination of primitive (low-crowned and complex) and derived (high-crowned and simple) dental characteristics. This ‘homogeneity’ and the lack of polarity in the development of the dental features through time resulted in the recognition of only four, poorly delimited, fossil genera (Vetuspalax, De Bruijn, Marković & Wessels Citation2013, Debruijnia, Ünay, Citation1996, Heramys Klein Hofmeijer & De Bruijn, Citation1985 and Pliospalax Kormos, Citation1932 = Sinapospalax Sarıca & Sen, Citation2003 see De Bruijn et al., Citation2013, Citation2015). Since Spalacine species do not cluster into readily recognizable groups generic allocation remains subjective. This peculiar property is exemplified by the material that will be described below.
Methods
The material described below is housed in the collections of the department of Earth Sciences of Utrecht University, the Netherlands. Measurements have been taken with an Ortholux microscope with mechanical stage and measuring clocks and are given in 0.1 mm. units. Upper molars are indicated by M1–3, lower molars by m1–3. Specimens are all figured as from the left side. If the original is from the right side its number on the plate has been underlined. All pictures were taken with a Jeol table SEM.
Systematic paleontology
Order Rodentia Bowdich, 1821
Family Muridae Illiger, 1811
Subfamily Spalacinae Gray, Citation1821
Extinct genera included: Vetusspalax, Debruijnia, Heramys and Pliospalax
Genus Debruijnia Ünay, Citation1996
Type species Debruijnia arpati Ünay, Citation1996
Type locality and level: Keseköy, Anatolia, MN3
Other species included: Debruijnia sp. (in Ünay and Göktas, Citation1999), Debruijnia kostakii n. sp.
Debruijnia kostakii n.sp.
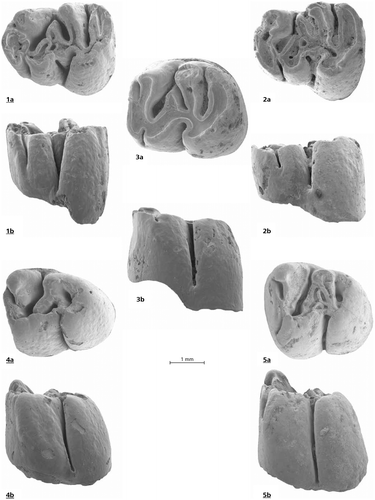
Figures and
Figure 2. Debruijnia kostakii from Karydia 2: 1 a, b, M1 inverse No. 1027, 3 a, b, M1 No. 1021, 4, M1 No. 1023, 5 a, b, M2 inverse No. 1035, 6 a, b, M2 No. 1031, 7 a, b, M3 No.1038, 8 a, b, M3 No. 1039. Debruijnia kostakii from Karydia 3. Figure M1 No. 1041.
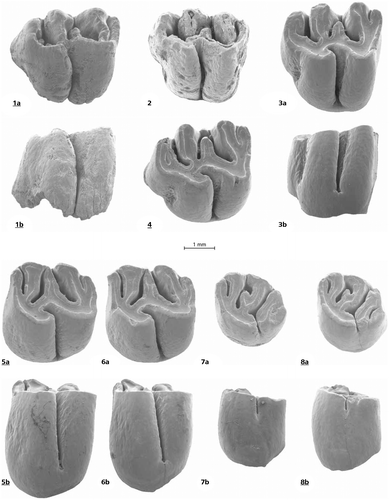
Figure 3. Debruijnia kostakii from Karydia 2: 1 a, b, m1 No. 1014, 2 a, b, m1 No. 1013, 3 a, b m2 No. 1016. 4 a, b, m3 No. 1019, 5 m3 No. 1020.
Selected reference: Debruijnia kostakii n. sp. Debruijnia n. sp (large) in Theocharopoulos (Citation2000, p. 86).
Derivatio nominis: I dedicate this species to the late Constantinos D. Theocharopoulos (Kostaki for friends) who showed admirable character and stamina when we collected the Karydia material in the rain.
Holotype: m1 dext. KR 2, nr. 1014, Figure (1a & 1b).
Type locality: Karydia 2, (KR 2), Greece, MN4.
Diagnosis: Debruijnia kostakii is a large spalacine. The anteroloph of the M1 is separated from the protocone by a shallow notch and the anterior arm of the protocone is either complete or interrupted. The metalophule is transverse in some M1, but connected to the posteroloph in others. The sinus of the five-lophed M3 remains open lingually until an advanced stage of wear. The m1 is elongate and its metaconid does not fuse with the anteroconid until a late wear stage. The protosinusid is progressively less developed from m1 through m3. A long free-ending posterior arm of the hypoconid is present in fresh m1.
Differential diagnosis: Most teeth of Debruijnia kostakii are somewhat larger than those of P. sinapensis Sarıca & Sen, Citation2003 (MN10), P. canakkalensis Ünay, Citation1981(MN7/8) and P. primitivus Ünay,Citation1978 (MN7/8). Their dental pattern is in many respects intermediate between the type species of the genus D. arpatti and the oldest known Pliospalax: P. marmarensis Ünay, Citation1990. The degree of lophodonty and hypsodonty in D. kostakii is similar as in Pliospalax, but its cheek teeth differ in retaining plesiomorph details such as the presence of the anterior arm of the protocone in the M1 and the free-ending posterior arm of the hypoconid in the m1. In these respects D. kostakii it is closer to Debruijnia than to Pliospalax.
Description
M1. The depth of the notch that separates the anterocone from the protocone and the development of the anterior arm of the protocone of the M1 show considerable individual differences. The mesoloph is strong, but never reaches the labial edge of the occlusal surface.
M2. All five M2 are similar. Their sinuses reach almost to the base of the crowns, their protolophules are directed forwards and connect to the anteroloph just labially of the protocone while the metalophules are directed posteriorly and connect to the posterolophs just labial of the hypocones.
M3. The two available M3 are very similar, both have five lophs and a sinus that does not reach as close to the base of the crown as in the M1 and M2.
m1.The anterior outline of the m1 is like in most cricetids, because the anteroconid is not fused with the far forwards situated metaconid as in Pliospalax. The labially open protosinusid reaches rather far down. The short robust posterior arm of the protoconid ends free. The mesolophid is strong. The long posterior arm of the hypoconid fuses with the posterolophid in a rather early wear stage.
m2. The protosinusid of the m2 does not reach far down. The robust posterior arm of the protoconid ends free and the mesolophid and the posterior arm of the hypoconid are absent.
m3. The protosinus of the m3 is very weak. The posterior arm of the protoconid is directed postero-lingualy and may connect to the entoconid. The short, rudimentary hypolophulid is directed towards the hypoconid.
Conclusions
The age estimate for the Karydia assemblage is late Early Miocene (MN4) (Theocharopoulos, Citation2000), an age that is intermediate between that for Keseköy (MN3) (Ünay et al. Citation2003), type locality of the type species of Debruijnia, and for Paşalar (MN5) (Ünay et al. Citation2003), the locality that yielded the oldest Pliospalax. The dental morphology of D. kostakii bridges the gap between Debruijnia and Pliospalax which suggests that these are closely related. Its cheek teeth are, however, larger than those of any of the younger species of Pliospalax. It seems therefore unlikely that there is a direct ancestor – descendant relationship between P. kostakii and a known species of Pliospalax. The occurrence of seemingly random combinations of dental characteristics in species of different genera shows that the complex evolution of the Spalacinae is poorly understood. The available record suggests that Debruijnia and Pliospalax are part of the main lineage leading to the extant Spalacinae, while Heramys with its high-crowned cheek teeth and reduced M3, and Vetusspalax with its relatively brachyodont simple teeth represent extinct branches.
Funding
This work was supported by the Utrecht University and Kempen en co merchant bank, Amsterdam.
Disclosure statement
No potential conflict of interest was reported by the author.
Acknowledgements
I am grateful to the directory of the B.E.A.K brick factory in Komotini for the hospitality in their quarries as well as in the factory for sample drying. A number of colleagues have helped me collecting, drying and wet-screening. Among those my special gratitude goes to Constantinos S. Doukas for his diplomacy and for paving my way and to the late Constantinos D. Theocharopoulos for his dedication, enthusiasm and sense of humor. I thank my colleague Wilma Wessels for her support and criticizm. Last, but not least I thank Utrecht University and Kempen en co merchant bank for financial support. Tilly Bouten helped in making the SEM pictures and Margot Stoete (both Utrecht University) made the plates and figure one.
References
- de Bruijn H, Marković Z, Wessels W. 2013. Late Oligocene rodents from Banovići. Palaeodiversity. 6:63–105.
- de Bruijn H, Bosma AA, Wessels W. 2015. Are the Rhizomyinae and the Spalacinae closely related? Contradistinctive conclusions between genetics and palaeontology. Palaeobiodivers. Palaeoenviron. 95:257–269.10.1007/s12549-015-0195-y
- Gray JE. 1821. On the natural arrangement of vertebrose animals. London Med Repos. 15:296–310.
- Klein Hofmeijer G, de Bruijn H. 1985. The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 4, The Spalacidae and Anomalomyidae. Proc Koninklijke Akademie van Wetenschappen, Series B. 88:185–198.
- Kormos T. 1932. Neue Pliozäne Nagetiere aus der Moldau [New Pliocene roodents from Moldavia]. Paläont Z. 14:193–199.10.1007/BF03041633
- Sarıca N, Sen S. 2003. Spalacidae. In: Fortelius M, Kappelman J, Bernor RL, editors. Geology and paleontology of the Miocene Sinap Formation, Turkey. New York, NY: Columbia University Press; p. 141–163.
- Theocharopoulos KD. 2000. Late Oligocene-Middle Miocene Democricetodon, Spanocricetodon and Karydomys n. gen. from the eastern Mediterranean area. Gaia. 8:1–116.
- Topachevski VA. 1969. Fauna of the USSR, mammals, mole rats, Spalacidae. In: Academy of sciences of the USSR, editor. Fauna of the USSR, Mammals. Zoological Institute, New Series, 99, 1–150. New Delhi: Amerind Publishing Co Pvt. Ltd.
- Ünay E. 1978. Pliospalax primitivus n. sp. (Rodentia, Mammalia) and Anomalomys gaudryi from the Anchitherium fauna of Sarıçay. Bull Geol Society Turkey. 212:121–128.
- Ünay E. 1981. Middle and Upper Miocene rodents from the Bayraktepe section (Canakkale, Turkey). Proc Koninklijke Akademie van Wetenschappen B. 84:217–238.
- Ünay E. 1990. A new species of Pliospalax (Rodentia, Mammalia) from the Middle Miocene of Paşalar, Turkey. J Hum Evol. 19:445–453.
- Ünay E. 1996. On fossil Spalacidae (Rodentia). In: Bernor, R. L, Fahlbush, V, & Mittmann, HW, editors. The evolution of Western Eurasian Neogene mammal faunas. New York (NY): Columbia University Press; p. 246–252.
- Ünay E, Bruijn H de, Saraç G. 2003. A preliminary zonation of the continental Neogene of Anatolia based on rodents. In: Reumer JFW, & Wessels W. editors, Distribution and migration of Tertiary mammals in Eurasia. Deinsia 10, 539–548.
- Ünay E, Göktaş F. 1999. Late Early Miocene and Quaternary small mammals in the surroundings of Söke (Aydın). Preliminary results. Geol Bull Turkey, 43, 91–102.