
ABSTRACT
The cap-shaped shells of Triplicatella are known almost exclusively from small shelly fossil assemblages with articulated specimens showing unequivocally that they represent the operculum of a hyolith. Abundant specimens of Triplicatella opimus from the fine-grained shales of the Chengjiang Lagerstätte of South China with soft-part preservation are documented herein. The soft tissues, including the feeding apparatus and complex digestive system, in T. opimus strongly suggest that Triplicatella was a deposit feeder. The digestive tract of T. opimus consists of two limbs, a spiral loop folded into a chevron-like structure and a slightly recurved to straight anal tube, which are preserved as reddish-black traces enriched in iron. The new anatomical information obtained from T. opimus in the Chengjiang Biota suggests an intermediate stage in the development of the characteristic folded gut of orthothecid hyoliths. The new anatomical information reported here shows that Triplicatella is one of the best-preserved early members of the Orthothecida and promotes our understanding of the general anatomy and evolution of the Hyolitha.
Introduction
Hyoliths are a group of extinct Palaeozoic marine animals, which possessed distinct conical calcareous shells and range in age from the early Cambrian through to the Permian (Malinky and Yochelson Citation2007). The group was one of the first bilaterians to acquire shells and rapidly became one of the most abundant and cosmopolitan biomineralizing animals in Cambrian strata. Hyoliths are subdivided into two distinct groups, i.e. Hyolithida and Orthothecida, based on a number of morphological differences (Marek and Yochelson Citation1976). Typical hyolithids have a four-part external skeleton, including a conical conch with a ligula, an externally fitting operculum, and a pair of curved helens that resemble oars (Runnegar et al. Citation1975; Babcock and Robison Citation1988; Martí Mus and Bergström Citation2005; Martí Mus Citation2016). Orthothecids on the other hand only have two parts, a conch without a ligula and a retractable operculum (Marek and Yochelson Citation1964, Citation1976; Runnegar et al. Citation1975; Martí Mus et al. Citation2014; Skovsted et al. Citation2014).
Well-preserved hyoliths with articulated opercula are abundant in the Chengjiang Lagerstätte. Seven species of hyoliths have been previously recorded; however, these species are difficult to define based on previous descriptions (Hou et al. Citation1999, Citation2017). After re-examination of the hyoliths in the Chengjiang biota, we find that specimens referred to ‘Linevitus opimus’ Yu 1974 (Hou et al. Citation1999, Citation2017), that are normally articulated with their cap-shaped opercula, show a high morphological similarity with Triplicatella Conway Morris in Bengtson et al. Citation1990. Both taxa possess a nearly flat, subtriangular operculum without cardinal processes or clavicles and thus should be assigned to this genus (Liu et al. Citation2020).
The opercula and conch of Triplicatella were previously only known as components of small shelly fossil assemblages from Cambrian Series 2, Stages 3–4, (Bengtson et al. Citation1990; Skovsted et al. Citation2004, Citation2014; Skovsted and Peel Citation2007; Kouchinsky et al. Citation2015). Triplicatella was first reported by Bengtson et al. Citation1990 as a problematic, operculum-shaped fossil from the Cambrian of South Australia. Later different species of Triplicatella were subsequently discovered and described from small shelly fossil assemblages from North and North-East Greenland, western Newfoundland, Siberia and North China (Skovsted et al. Citation2004; Skovsted and Peel Citation2007; Kouchinsky et al. Citation2015; Pan et al. Citation2019). Due to the lack of a co-occurring conch, Triplicatella remained a problematic fossil (Skovsted et al. Citation2004), until Skovsted et al. (Citation2014) described articulated conchs and opercula of Triplicatella disdoma Conway Morris in Bengtson et al. Citation1990 in an assemblage of SSFs from South Australia and showed that Triplicatella was closely related to orthothecid hyoliths.
Triplicatella opimus (Yu, Citation1974) has a rapidly and evenly expanding conch with a subtriangular cross section. The conch of T. opimus appears strongly mineralised and all specimens show signs of brittle deformation although most specimens retain substantial topography and are partly filled with sediment. Until recently, no soft anatomy had been reported for T. opimus, but with the exquisite Konservat-preservation mode in the Chengjiang biota, specimens of Triplicatella, from fine-grained shales, are found with extraordinarily preserved soft parts, including a tentaculate feeding organ (Liu et al. Citation2020), digestive tracts and muscle scars (herein). Soft parts are preserved as reddish-brown imprints, predominantly associated with the operculum and distributed throughout the conch. Reports of orthothecid hyoliths with preserved soft parts are rare and typically limited to the gut (Thoral Citation1935; Marek Citation1967; Runnegar et al. Citation1975; Kruse Citation1996; Malinky Citation2003; Devaere et al. Citation2014; Sun et al. Citation2016; Berg-Madsen et al. Citation2018). Based on the morphological details recently reported (Liu et al. Citation2020) and the new details herein, it is clear that the material of Triplicatella from the Chengjiang biota is of significant value not only regarding the systematic affinity of Triplicatella, but also for revealing the first direct evidence of the soft anatomy of Cambrian orthothecids.
Material and methods
More than 1000 hyolith specimens have been collected from yellowish-green or greyish-green mudstone from eight localities of the Chengjiang Lagerstätte (Fu et al. Citation2014), distributed on both sides of Dianchi Lake, Kunming, eastern Yunnan, China, by the working team of the Early Life Institute of Northwest University (Liu et al. Citation2020). Most specimens of Triplicatella opimus examined here were derived from the Yu'anshan member and collected from sections at Erjie town on the west bank of Dianchi Lake, where the hyoliths exhibit exceptional preservation of soft tissues. In our material (see Supplementary material), about 241 specimens from eight sections can be definitely referred to Triplicatella opimus, including some clusters with over 30 individuals. Of these, 81 specimens of T. opimus are preserved with soft tissues, 45 conchs of T. opimus have the body cavity preserved and 15 with the digestive tract preserved. In addition, 63 specimens preserve soft parts associated with the operculum and 2 specimens preserve the neck like connection between conch and operculum (Liu et al. Citation2020; see Supplementary material).
All specimens are deposited in the Early Life Institute, Northwest University, Xi’an, China (Prefix: ELI). Fossils were examined under a binocular Zeiss Zoom Stereomicroscope and photographed with a stereophotographic Zeiss Smart Zoom 5, with different illuminations for particular views when high contrast images were required. Some uncoated specimens were analysed by a FEI Quanta 650 scanning electron microscope, Backscatter Scanning Electron Microscope (BSEM) and environmental mode-Energy Dispersive X-ray spectrometry (EDS) system to examine microstructures of fossils and taphonomic features, at the State Key Laboratory of Continental Dynamics, Northwest University, Xi’an. To understand the preservation of the fossils and their chemical composition, selected specimens were also examined by non-destructive Micro X-ray Fluorescence (μ-XRF). Some specimens were photographed under fluorescence Stereomicrograph Nikon SMZ1500 system at the Department of Geology, Northwest University, to improve the colour contrast of the fossilised soft tissues to the remaining skeletal hosts. Some selected specimens were investigated using Mirco-CT scans to observe the internal structures hidden inside three-dimensionally preserved conchs, the data were visualised in VG Studio Max 2.0 and Drishiti 2.4. Figures were prepared using Corel Draw 2018 with the rendered macrophotographs using ADOBE PHOTOSHOP CS4.
Systematic palaeontology
Phylum uncertain
Class Hyolitha Marek, 1963
Order Orthothecida Marek, Citation1966
Family Triplicatellidae fam. nov.
Zoobank id F6622763-6098-4F76-8AF8-64A93AB9978B
Type genus
Triplicatella Conway Morris in Bengtson et al. Citation1990.
Genera included
Type genus, Holoplicatella Clausen & Alvaro, 2006, Paratriplicatella Pan, Skovsted, Sun & Li, Citation2019.
Diagnosis
Orthothecid hyoliths with oval to rectangular to subtriangular conch cross section and a planar or pyramidal operculum with dorsal and/or ventral folds but lacking cardinal processes.
Discussion
Triplicatellidae differs from other orthothecid families by the lack of differentiated cardinal processes in the operculum. The cross section of the conch is highly variable from oval to subrectangular to subtriangular and a dorsal carina may be developed. The operculum is planar or laterally convex in a pyramidal shape and has dorsal and/or ventral folds in variable configurations that characterise individual genera and species.
Many Cambrian orthothecids are poorly known, with descriptions often restricted to the conch, while the operculum, that seems to be much more important for classification (Pan et al. Citation2019), is frequently lacking. However, it is clear that the morphological disparity of Cambrian hyoliths is much greater than among younger forms, which historically formed the basis of the current classification (Marek Citation1967; Malinky and Yochelson Citation2007). One important feature are the clavicles, that are characteristic features of hyolithid opercula but supposedly lacking in orthothecids (Malinky and Yochelson Citation2007). However, many Cambrian hyoliths seem to combine clavicle-like features with otherwise orthothecid-like morphologies (Malinky & Skovsted Citation2004; Skovsted et al. Citation2004). This is, for example, the case for Paratriplicatella from North China, which has an internal ridge of tubular clavicle-like structures on the inner dorsal rim of an operculum, otherwise the operculum is indistinguishable from Triplicatella (Pan et al. Citation2019). It is possible that clavicle-like features evolved separately in different groups of hyoliths during the early Cambrian and that such simple traits are of limited value for higher level classification. Consequently, the classification of hyoliths is currently in a state of flux, and the relationships of Triplicatellidae to other orthothecid families are difficult to evaluate without a major revision of the entire Order, which is outside the scope of the present paper.
Genus Triplicatella Conway Morris in Bengtson et al. Citation1990()
Table 1. Recognised species and occurrences of the genus Triplicatella.
Type species
Triplicatella disdoma Conway Morris, in Bengtson et al. Citation1990, pp. 231–232. figs 157, 158. Holotype: UNEL1848, Curramulka, Parara limestones, lower Cambrian of Australia.
Diagnosis
Bilaterally symmetrical operculum, transversely oval to subtriangular in outline. One to four weakly to strongly developed folds on the dorsal margin, sometimes accompanied with poorly developed ventral marginal folds or invaginations. No ornamentation on operculum or sometimes with weakly defined concentric growth lines. The internal surface of the operculum is smooth without cardinal processes or clavicles, but occasionally a deep median depression exists in the sub-central part. The conch of Triplicatella is elongate, cone-shaped and gently curved, with a rounded to subtriangular cross-section matching the shape of the operculum and with occasional transverse septa.
Remarks
This genus was first described by Bengtson et al. (Citation1990) as an operculum-like fossil of uncertain affinity from South Australia. Some researchers interpreted Triplicatella as the earliest known chiton, (Class Polyplacophora; Yates et al. Citation1992) and Conway Morris (Citation1993) suggested that it represented a halkieriid shell. A steinkern of Triplicatella with associated conch and operculum from Australia was illustrated by Demidenko (in Gravestock et al. Citation2001, pl. 10, ), which indicated that Triplicatella was probably a hyolith operculum. Unfortunately, this specimen was never described. Skovsted et al. (Citation2004) also suggested Triplicatella was an operculum of a hyolith, in accordance with the original interpretation (Bengtson et al. Citation1990), due to the similarity in shape and the possible muscle scars on the interior surface (Skovsted et al. Citation2004). This interpretation was then strengthened with the full description of Triplicatella from Australia as conchs preserved as internal moulds of a hyolith with the operculum in place at the apertural end (Skovsted et al. Citation2014). The elongate cone-shaped and gently curved conch with a rounded to subtriangular cross-section, combined with the planar operculum lacking clavicles and cardinal processes, clearly indicates that Triplicatella can be placed in the Orthothecida (Skovsted et al. Citation2014).
The morphological features of Triplicatella, in particular the distinct series of three folds that typically occur on one side of operculum, and occasionally on both sides, clearly distinguish the genus from other taxa. The genus Sysoievia Marek, Malinky and Geyer, 1997 from Morocco, was proposed for hyolith specimens that possessed an orthoconic conch with a flat, semicircular operculum ornamented by transverse growth lines. Sysoievia was considered to be similar to Triplicatella (Marek et al. Citation1997a); however, Sysoievia differs from Triplicatella in having a butterfly-like pair of cardinal processes, and by lacking prominent folds on the dorsal margin of the operculum. The genus Paratriplicatella from North China possesses a slightly dorsally curved conch with a triangular cross-section, a convex triangular operculum with a fold on the dorsal side and two divergent folds on the ventral side. It was also recently considered broadly similar to Triplicatella (Pan et al. Citation2019). The most noticeable feature of Paratriplicatella, that differentiates this genus from Triplicatella, is the strongly developed ridge of tubular clavicle-like structures on the inner dorsal rim of the operculum. Holoplicatella from Spain (Clausen & Alvaro Citation2006) displays a prominent notch on one side, and two widely divergent folds on the other side of the operculum. The presence of these folds does invoke a resemblance to Triplicatella, but the Spanish taxon differs in the oval shape and high degree of convexity. Holoplicatella was originally described as a potential sclerite of a larger animal (Clausen and Álvaro Citation2006) but despite the poor preservation and limited material, the general morphology of Holoplicatella suggests that this taxon represents the operculum of a hyolith closely related to Triplicatella.
Stratigraphic and geographic range
Lower Cambrian (Stages 3–4) of South Australia, South China, the Anabar Uplift northern Siberia, North Greenland, North−East Greenland, western Newfoundland and North China.
Triplicatella opimus (Yu Citation1974)()
Figure 1. Triplicatella opimus from the Chengjiang Biota, Yunnan Province, China. a. the populous occurrence of Triplicatella opimus on a slab, specimen no. ELI H-168. b. a complete specimen with operculum filled with sediment preserved three-dimensionally, showing a deep central longitudinal carina on the dorsum, specimen no. ELI H-183A. c. dorsal view of an incomplete specimen showing the two shorter, divergent dorsal folds on the operculum marked by a box, specimen no. ELI H-155. d. interior of flat subtriangular operculum with two divergent folds on the dorsal side, and a weak fold adorned at the frontier margins pointed by white arrows, specimen no. ELI H-181B. e. showing the trapezoid-shaped apex marked by a box, specimen no. ELI H-121A. f. two dorsally compacted specimens with operculum, overview of specimens with well-preserved conchs and opercula, specimen no. ELI H-185A. g. Close-up view of the operculum with two divergent folds on the dorsum pointed by white arrows, specimen no. ELI H-185A. h. the flat venter of conch, specimen no. ELI SJZ- H-1565

Figure 2. Morphology of Triplicatella opimus from the Chengjiang Biota. a-d, h cap-shaped opercula of T. opimus. c-d, h, opercula preserved with the arcuate structures pointed by white arrows. a, specimen no. ELI H-192A. b, specimen no. ELI H-181B. c, specimen no.ELI H-191B. d, specimen no.ELI H-132. H, specimen no.ELI H-181B. e. dorsum of Triplicatella opimus associated with operculum, specimen no. ELI H-196A. f. the imprint of the dorsum, specimen no. ELI H-115. g. venter overlapped the operculum ELI H-183. i. venter of T. opimus, specimen no. SJZ-H-1693. j. some individual of T. opimus preserved dorsally and ventrally in an accumulation, pointed by red arrows, specimen no. ELI H-197A

1974 Linevitus opimus sp. nov.; Yu Citation1974, p. 111–112, fig. 44.16–17.
1999 Linevitus opimus Yu, Citation1974; Hou et al. p. 84, fig. 111.
2003 Linevitus opimus Yu, Citation1974; Chen et al. p. 302, plate I, .
2005 Linevitus opimus Yu, Citation1974; Zhao et al. p. 760, plate I, fig. 13.
2017Linevitus opimus Yu, Citation1974; Hou et al. p. 108–109, fig. 16.4–5.
2020 Triplicatella opimus (Yu, Citation1974); Liu et al. , 8.
Holotype
Specimen 2271, a cone-shaped conch (Yu Citation1974, fig. 44.16–17), from the top of the Qiongzhusi (Chiungchussu) Formation, Lower Cambrian, in Yaoying, Wuding county, Yunnan province, China.
Material
In our collection, there are 241 specimens that can be referred to Triplicatella opimus. Of these, 63 represent articulated specimens while 178 are disarticulated conchs, with some isolated opercula.
Diagnosis
Orthothecid hyolith with broad, cone-shaped conch with subtriangular cross-section; dorsum with two crests separated by deep carina tapering from apex to aperture; ventral side appears flattened and slightly convex with a curved ventral margin; apex with internal septa in larger specimens. Retractable opercula subtriangular to heart-shaped with initial shell shifted posteriorly, situated at about one-third of posterior-anterior distance, lacking both cardinal processes and clavicles; three symmetrical, posteriorly diverging (dorsal) folds; a single wide but very weak anterior (ventral) fold; two weakly expressed lateral ribs radiating from the initial shell towards anterolateral corners. External surface of conch and operculum with concentric growth lines; internal surface smooth with imprint of central muscle attachment area in operculum. Anterior unmineralised feeding organs constructed by two radial arms and an uncertain number of fine tentacles surrounding mouth; extendable pharynx connected to central muscle attachment area of operculum; posterior neck connecting operculum with conch. U-shaped, slightly undulating to folded digestive tract.
Description
The rapidly and evenly expanding conch of Triplicatella opimus, is cone-shaped (),)), with length ranging from 3 to 28 mm and width ranging from 2 to 14 mm ()). Conchs are strongly mineralised and all specimens show signs of brittle deformation. Most specimens retain substantial topography and are partly filled with sediment (), ), ), )). The cross section of the conch was subtriangular. The dorsum bears a deep central longitudinal groove or carina, separating two rounded crests. The ventral side of the conch is flat or slightly convex. Ventral apertural margin with a rounded anterior edge, reminiscent of a ligula-like extension (), )). Some specimens preserved in ventral view show a weakly developed marginal carina, probably due to ventral folding during compaction ()). Closely spaced growth lines superimposed on both ventral and dorsal sides of the conch. The apical portion of the conch often appears horseshoe-shaped after breaking along simple transverse septa sealing off the original apex ()).
Figure 3. Bivariate plots of conchs and opercula of Triplicatella opimus from the Chengjiang Lagerstätte, South China. a. plots of the length and width of conchs of Triplicatella opimus. b. plots of the length and width of opercula of Triplicatella opimus.
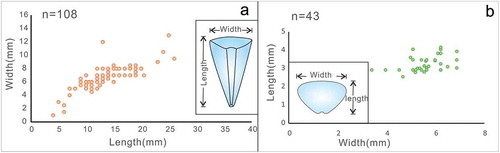
Figure 4. Soft-parts and connective tissues associated with the opercula and conch in Triplicatella opimus. a- cspecimen no. ELI H-176A, a. Specimen showing the neck-like band connecting the dorsal margin of the operculum and the internal region of the apertural conch. b. sketch of a. c. fluorescence microscope analysis of the soft-tissues preserved on the operculum. d. The iron elemental mapping of g, the soft tissues enriched in iron. e, g.Part and counterpart, ventral view of the 3-dimensionally preserved connective tissues that have been detached and are now preserved between the operculum and conch, specimen no. ELI H-129A and B. f. sketch drawing of eand g
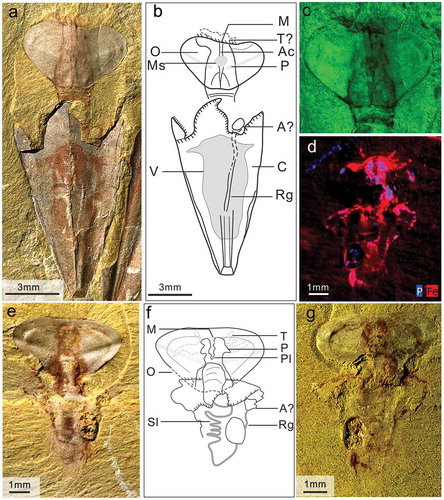
Figure 5. Soft-parts on the conch of Triplicatella opimus. a. a strongly compressed specimen with operculum, showing imprints of the spiral loop-shaped gut and recurved straight intestine, specimen no. ELI H-131A. b. the sketch of a. c. a dorsal interior impression of conch preserved with recurved gut and distal distended portions, specimen no. ELI H-140. d. internal mould preserved with the spiral portion of gut, specimen no. ELI H-12B. e. counterpart of d, note the negative impression of the spiral looped gut and are curved gut imprint, specimen no. ELI H-12A. f. dorsal view of the recurved gut on the partial preserved conch, specimen no. ELI H-186B

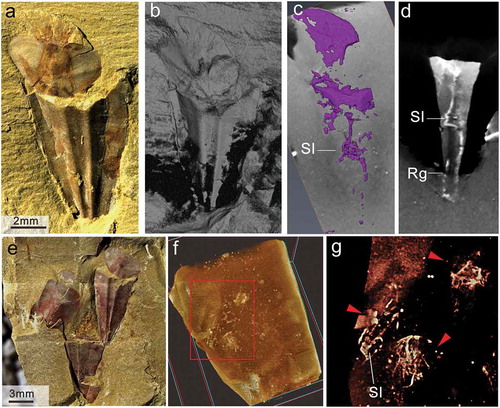
Figure 6. Micro-CT set-up and fine details revealed on the surface of the specimen Triplicatella opimus. a. Light microscopy (macrophotography), conjoined conch and operculum with extended ribbon-like tentacles, specimen no. ELI H-113. b. The same specimen as in a, but shown in a longitudinal X-ray section produced by micro CT. c-d. Fine details of the internal structures revealed with micro-CT about the specimen in a, dorsal view showing the gut tracts inside the conch. e. General views of the studied material by light microscopy, showing three T. opimus hyoliths individuals (red arrowheads) on a slab. Boxed area is shown enlarged in f, specimen no. ELI H-179. f-g. close up of the e, showing the fine details about the soft tissues like digestive tracts and muscle scars, revealed inside the conch of hyoliths produced by micro CT
The operculum is flat, subtriangular and cap-shaped (), )), with width ranging from 3 to 6.6 mm and length ranging from 2.9 to 4.1 mm ()), matching the apertural outline of the conical conch. The apex or initial shell is indistinct, but when observable appears to be shifted posteriorly (dorsally), to a position about one-third of the entire posterior-anterior distance. There are no cardinal processes or clavicles on the internal surface. Three shorter, divergent folds are present on the posterior (dorsal) side of the operculum ()). The low, wide central fold is separated from the two stronger lateral folds by two deep invaginations. A single, wide and poorly expressed central fold adorns the anterior (ventral) margin (), )). Two broad lateral ridges converge from the apex and run towards the widest lateral margin of the operculum ()). Ornament of fine growth lines is present on the surface along the margin of operculum.
Some specimens of Triplicatella opimus in the Chengjiang biota preserve exquisite imprints of soft tissue (,,). The soft parts are preserved in the form of reddish-brown stained patches on the conch or on the internal surface of the operculum. Soft parts representing the tentaculate feeding organ were described by Liu et al. (Citation2020) and additional soft parts representing the neck-like connection of the conch and operculum, the mantle cavity, muscle scars and the alimentary canal are described separately below.
Remarks
Triplicatella opimus (Yu Citation1974), from the Chengjiang biota of South China, has been previously described as a hyolithid, Linevitus opimus Yu, Citation1974 (Hou et al. Citation1999, Citation2017; Chen Citation2004; Zhao et al. Citation2005). The species has a cap-shaped operculum and a subtriangular cross section of the conch with a flattened venter, which is reminiscent of hyolithid hyoliths. But, it lacks both cardinal processes and clavicles in the opercula, and the arc-shaped margin of the ventral side of the conch aperture is not a ligula, but appears to match the curved ventral outline of the operculum like the prominent dorsal crests match the dorsal folds of the operculum. No remains of helens associated with Triplicatella could be observed in the exceptionally preserved biota. Abundant opercula are preserved directly associated with the conchs, and in many cases, the posterior of the operculum appears to be partially covered by the anterior margin of the conch (),)), which indicate that the operculum could be partly withdrawn inside the conch.
Thus, there are no characteristic features of the species which suggest inclusion in the Hyolithida. Based on a comparison of the morphology of the flattened cap-shaped opercula with ventral and dorsal folds but without cardinal processes or clavicles, we consider this taxon to be most closely comparable to the orthothecid genus Triplicatella. Until now, five species belonging to the genus Triplicatella have been described, including T. disdoma Conway Morris in Bengtson et al. Citation1990 (type species), T. sinuosa Skovsted, Peel, and Atkins, Citation2004, T. peltata Skovsted, Peel, and Atkins, Citation2004, T. papilo Kouchinsky in Kouchinsky et al. Citation2015 and Triplicatella xinjia Pan, Skovsted, Sun Li, Citation2019. The Chengjiang species possesses three weak dorsal folds, two low ridges towards the lateral margins of the operculum ()) and a single, wide and quite indistinct ventral fold (),)). However, the more weakly developed folds on the opercula when compared to other species known exclusively from acid residues may be due to the flattening of specimens during compaction, characteristic of this Konservat Lagerstätte (Zhu et al. Citation2005; Han et al. Citation2006). Regardless, the morphology of the operculum with a single ventral fold and three short dorsal folds clearly distinguish T. opimus from T. disdoma, T. sinuosa, T. papilo and T. xinjia all of which have three ventral folds and one to four much more prominent dorsal folds. Triplicatella opimus is more similar to T. peltata from Greenland in having a distinct triangular cross section and subdued folds. However, T. peltata has a single prominent dorsal fold unlike the three dorsal folds in T. opimus where the central fold is less pronounced than the lateral folds. The conch of the Chengjiang species shows a distinct median carina forming two crests on the dorsal side (),)) while the conch of T. disdoma from Australia is characterised by an curved and slowly expanding conch with a rounded, subtriangular cross-section (Skovsted et al. Citation2014), which is similar to the morphology of T. xinjia from North China (Pan et al. Citation2019). The conch is not known for any other described species of Triplicatella. Compared with T. disdoma and T. xinjia, the conch of T. opimus is more rapidly expanding with a relatively wider and shorter cone-shape, and with two crests and a deep carina on the dorsal side. The visible differences indicate that the Chengjiang species represent a separate species of Triplicatella, despite the fact that this genus is previously only known from SSF assemblages.
There is some morphological resemblance in conch morphology, particularly with the presence of two crests and a deep carina (), )), between T. opimus and ‘Hyolithes’ conularioides Tate, Citation1892 from Australia (Bengtson et al. Citation1990) and Antarctica (Wrona Citation2003). ‘Hyolithes’ conularioides in these records were preserved as internal moulds of the conch, with a convex dorsum with a median carina separated by longitudinal crests and a flat venter. It is worth noting that the apex of ‘Hyolithes’ conularioides has been recorded with a bulbous protoconch (Bengtson et al. Citation1990), and Wrona (Citation2003) described the apex as being separated by a septum-like wall, which is reminiscent of the horseshoe-shaped apical septa in the conch of T. opimus ()). However, the exact relationship between T. opimus and ‘Hyolithes’ conularioides cannot be evaluated as the operculum of the latter is not known.
The operculum of Triplicatella could probably be withdrawn a short distance into the conch, because the measurement of the operculum is smaller than the apertural diameter of the conch, as is typical for orthothecids (Skovsted et al. Citation2014). Although the conch shape of most specimens of T. opimus from Chengjiang is distorted by compaction, abundant specimens show partial overlapping of the posterior part of the operculum by the dorsal margin of the conch (), )). This probably indicates that the operculum of T. opimus could be withdrawn inside the conch.
In some aspects, Triplicatella opimus is similar to Paratriplicatella shangwanensis Pan, Skovsted, Sun and Li, Citation2019 from North China. These similarities include the convex dorsum and a flat venter with the short ligula-like extension on the venter slightly beyond the dorsal aperture and similar lateral ribs connecting the lateral margins of the opercula to the central area. However, T. opimus lacks the tubular clavicle-like structures along the arched margin on the dorsal rim on the operculum, that are characteristic of P. shangwanensis.
Occurrence
The Yu’anshan Member (Eoredlichia Zone) of the upper part of Heilinpu (formerly Qiongzhusi) Formation, Early Cambrian, Stage 3, Yunnan Province, South China.
Soft parts of Triplicatella opimus
Multiple specimens of Triplicatella opimus preserve imprints of the soft anatomy as red-brownish stains (,,). Soft parts of the operculum relating to the tentaculate feeding organ and central cylindrical mass (pharynx) (),)) were described by Liu et al. Citation2020. Arcuate structures in the form of red imprints at the dorsal margin of the operculum and following the growth lines along the lateral margins of the operculum were interpreted as the margins of shell secreting epithelia (Liu et al. Citation2020). These structures are also preserved in specimens illustrated here ()).
The connection of operculum and conch
In some specimens of Triplicatella opimus, soft parts in the form of a neck-like projection are present between the operculum and conch, showing the connection of the skeletal parts of the hyolith (); see also Liu et al. Citation2020, ). The neck-like structure extends from the posterior of the operculum to the anterior of the conch ()) and sometimes preserves obscure annulations. In one specimen, the neck-like structure is strong and thick (about 1 mm in width (approximately representing one-third of the width of the operculum) and 2.5 mm in length) with the width approximately matching the width of the central cylindrical mass of the operculum ()). In another specimen, the neck-like structure attached to faintly preserved soft parts without associated conch, apparently replicating the size and shape of the operculum, and an extended mass with vague spiral impressions (). We interpret these structures to represent detached soft parts of the secretory tissues of the conch margin and the visceral mass, including parts of the intestine. This specimen shows that the soft tissues of the hyolith conch were strongly connected to the operculum in the central position and could be displaced when the conch and operculum were separated ()). Compared with the hyolithids where the conch and operculum are connected through muscles attaching near the base of the cardinal process (Dzik Citation1978), the neck-like structure of Triplicatella opimus, we suggest, functioned as an extendable attachment organ between the conch and operculum in orthothecids like Triplicatella, which possess a flattened operculum without cardinal process.
The digestive system
Under a light microscope, most specimens of Triplicatella opimus from the Chengjiang biota clearly preserve a conical reddish impression inside the conch which probably represents the body cavity of the hyolith (). These red or dark stained areas in the conch centre are often located about one-third of the distance from the conch aperture, but red or yellow stained areas are also sometimes directly associated with the apertural region of the conch. These highly variable structures are not distinctive and preclude direct identification of their biological origin (),),)). We suggest that these areas represent a partly decomposed visceral mass but without differentiated organs.
The exquisite preservation of the Chengjiang biota suggests that the digestive tract of orthothecid hyolith may be preserved as thin imprints on the internal surface of some conchs. Triplicatella opimus bears a narrow U-shaped intestine, forming spiral loops displayed on the internal surface of the ventral side (),)) and a slightly recurved to straight intestine in a dorsal position inside the conch ()). The convoluted ventral digestive tract with a spiral loop folded into a chevron- or accordion-like structure, is preserved as a faint reddish-black tract enriched in Fe ()). The chevron-shaped gut usually appears near the left lateral margin of the visceral cavity in the conch. On the dorsal side, the recurved straight intestine, sometimes slightly bent, appears to be aligned with the right margin of the visceral cavity.
A complete digestive system is distinctly preserved in a single specimen where the soft parts are preserved without the conch ()). The spiral loops are ca. 3 mm in length with a posterior width of approximately 1.5 mm ()), and the straight gut is about 4 mm in length. A series of at least five arcuate loops can be identified in different specimens ()). The recurved distal part of the intestine is present as a straight to slightly undulating and partly distended band following the longitudinal axis of the conch and appear to end in an asymmetrically placed anus ()).
The gently sinuous to straight part of the gut was regarded as an anal tube on the dorsal side of the conch in a series of papers (Thoral Citation1935; Runnegar et al. Citation1975; Kruse Citation1996; Malinky Citation2003; Devaere et al. Citation2014; Berg-Madsen et al. Citation2018). The record of both anal tube and folded gut preserved in a single specimen is rare in our material, and the structures are usually separated between part and counterpart by mud infilling ()). This indicates that the original location of the two parts of the guts was not in the same plane inside of the Triplicatella conch.
Additional detailed information on the digestive system was observed under Micro-CT ()). In some of the best-preserved specimens of Triplicatella opimus that were infilled with sediment, fine internal structures could be resolved in the body cavity, which could not be otherwise observed on the surface of the fossil slab, highlighting the differential preservation of the soft tissues and the shell. The gut inside the conch can be clearly separated from the matrix due to the differential density, caused by the pyritisation of soft parts in the Chengjiang biota ()). The internal digestive tract in the best preserved specimen is composed of a short spiral looped part with just two arcuate loops, located in the central part of the conch, and a partially preserved recurved-anal tube ()). Another specimen preserves a cluster of three T. opimus individuals ()). The three individual specimens show association of conch and operculum (two specimens preserved dorsally, one preserved ventrally with the operculum beneath the conch), and the conchs are preserved as internal moulds infilled with mud. Through Micro-CT, anatomical structures including parts of the guts and other soft tissue can be observed (as indicated by red arrows in )). One individual on the left side preserves a partial digestive tract with a spiral looped gut (about 3 arcuate loops) ()). The other two specimens preserved obscure soft tissue, probably representing the decomposed visceral mass inside the conch.
Discussion
Systematic implications
Triplicatella has been studied for 30 years, but previous studies were limited to specimens preserved as small shelly fossils and the majority of publications focused purely on systematic descriptions because only the cap-shaped opercula were preserved (Bengtson et al. Citation1990; Skovsted et al. Citation2004, Citation2014; Skovsted and Peel Citation2007; Kouchinsky et al. Citation2015; Pan et al. Citation2019). Triplicatella disdoma was not demonstrated with certainty as an orthothecid hyolith until 2014, through specimens discovered in South Australia preserved as internal moulds of associated conch and opercula (Skovsted et al. Citation2014). The discovery of Triplicatella in the Chengjiang biota extends the distribution of Triplicatella to South China, and also represents the first discovery of Triplicatella from crack-out collections in shale. The digestive system of Triplicatella consisting of a folded gut and a recurved straight intestine (anal tube) reported herein, are characteristic of Orthothecid hyoliths (Thoral Citation1935; Marek Citation1967; Runnegar et al. Citation1975; Kruse Citation1996; Malinky Citation2003; Devaere et al. Citation2014; Sun et al. Citation2016; Berg-Madsen et al. Citation2018) and confirms the systematic position of Triplicatella among orthothecid hyoliths. The soft tissues exquisitely preserved in T. opimus from the Chengjiang biota, including the tentaculate feeding organ and pharynx associated with the operculum (Liu et al. Citation2020), the neck-like connection of conch and operculum as well as the folded gut provides a comprehensive understanding of the genus and adds new information on the anatomy of Orthothecida.
Taphonomy
For hyoliths, as a group of extinct animals, the reconstruction of the soft parts is a necessary step towards understanding both their anatomy and evolution. However, the discovery of hyoliths with associated soft parts is rare, with only a few published references of these recorded from the Cambrian, Ordovician and Devonian (English and Babcock Citation2010; Devaere et al. Citation2014; Martí Mus Citation2016; Sun et al. Citation2016; Moysiuk et al. Citation2017; Berg-Madsen et al. Citation2018, for references).
The specimens of hyoliths from the Chengjiang biota were all cracked out from fine-grained yellowish-green mudstone. However, the fossils of T. opimus documented here are preserved in three dimensions due to sedimentary infilling, and specimens always retain a thin coat of clay matrix inside the conchs. Many specimens preserve the operculum intimately associated with the conch, and it may be found either partly inside or just in front of the conch aperture. The opercula are often slightly displaced from the conch and in most cases appear as imprints of internal moulds (,,,). The opercula are rotated to a nearly horizontal position, invariably with the dorsal side closest to the conch, indicating that the main attachment of the operculum to the conch was along this part of the margin (,,,,) and this is confirmed by the position of the neck-like tissues connecting the skeletal parts. Some clusters of T. opimus show a preferential orientation (), )) probably reflecting biological concentration on the seafloor. The soft parts (consisting of the digestive tract, the body cavity, muscle scars and occasionally feeding organ) were exquisitely preserved in these specimens (,,,,)). These observations reasonably suggest that these animals were buried alive and in situ or experienced only gentle transportation prior to deposition (Hou et al. Citation2004).
Figure 7. Taphonomic details of Triplicatella opimus from the Chengjiang. a, two conjoined individuals with reddish-brown tinges, specimen no. ELI H-176B. b-c, The elemental maps of Si, Fe, P about the specimen in a, Overview of the high iron and phosphorus concentration on the soft tissues of Triplicatella opimus. d-f, The elemental maps of S, Fe, P of the specimen at Figure 5 c, showing the high iron and phosphorus concentration on the gut of T. opimus, specimen no. ELI H-140. g-h, Scanning electron micrograph (SEM) images of the soft tissues in Triplicatella opimus preserved with pyrite framboids
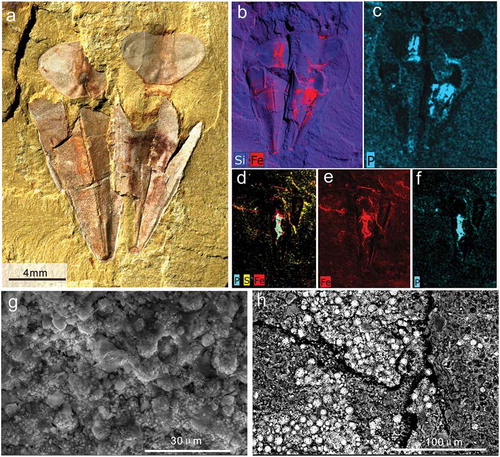
The mechanisms and processes of taphonomy in the Chengjiang Lagerstätte are still controversial despite being widely discussed (Briggs Citation2003; Butterfield Citation2003; Gabbott et al. Citation2004; Han et al. Citation2006). Soft parts of T. opimus are preserved as reddish-brown-stained impressions on the sediment surface ()), which is a common fossilisation mode for nonmineralized organisms in the Chengjiang Lagerstätte (Gabbott et al. Citation2004; Zhu et al. Citation2005; Forchielli et al. Citation2014). Analyses reveal no distinction in composition between the surface of shells and the surrounding matrix in SEM–EDX mapping ()). However, the soft parts of T. opimus show a higher concentration of Fe and P, with a slight increase in S ()), likely indicating the presence of pyrite. The low levels of sulphur are thought to be a consequence of weathering or diagenesis (Littke et al. Citation1991; Moses and Herman Citation1991; Zhu et al. Citation2005). Both framboids and octahedral crystals precipitated as pyrite have been found on the remains of the soft parts in T. opimus ()). Framboidal microarchitecture in secondary electron (SEM) images shows microcrystals (ca. 5 μm) densely arranged on the soft tissues (including feeding organ (Liu et al. Citation2020, ,7a-d), muscle scars, body cavity, and guts) of T. opimus, ()). Labile tissues such as these have been considered as the most reactive tissues related to microbial decay (Gabbott et al. Citation2004). The high concentration of iron on the surface of the fossils is most likely the result of the subsequent weathering of this pyrite.
Comparison of hyolith digestive systems
The digestive systems of hyoliths are different between the two orders, Orthothecida and Hyolithida. The guts of Hyolithida are generally considered to be simple, U-shaped and are mainly known from specimens preserved as compression fossils in shale from Konservat-Lagerstätten (Meshkova and Sysoev Citation1981; Babcock and Robison Citation1988; Houbrick et al. Citation1988; Butterfield Citation2003; English and Babcock Citation2010; Moysiuk et al. Citation2017). However, the digestive system of Orthothecida has a characteristic sinuously folded gut with a straight anal tube and is often preserved in three dimensions (Thoral Citation1935; Kruse Citation1996; Devaere et al. Citation2014; Berg-Madsen et al. Citation2018). The three dimensionality of the orthothecid digestive tract has been interpreted as being the result of phosphatic preservation, possibly due to partly phosphatic ingested sediments (Devaere et al. Citation2014; Berg-Madsen et al. Citation2018) and presumably caused by microbially mediated phosphatisation (Devaere et al. Citation2014). Herein, the intestine of Triplicatella is preserved by means of a red mineral film containing Fe-oxides, a result of weathering after diagenetic pyritisation. The reason why the guts of orthothecids from the Chengjiang Biota differ from specimens in previous reports, is probably due to different preservational modes. All material in our collection comes from mudstone in the Yuanshan Member where all fossils have been subjected to high levels of compression (Han et al. Citation2006). Lacking early cementation, substantial decay occurred prior to mineralisation, that resulted in the imprint-preservation mode for the soft parts in the Chengjiang Biota (Briggs Citation2003).
Although the digestive tracts of T. opimus are only partially preserved in the available specimens, the arrangement of the spiral loops is closely comparable to the spiral proximal portion of the intestine of orthothecid hyoliths described previously (Thoral Citation1935; Kruse Citation1996; Devaere et al. Citation2014; Berg-Madsen et al. Citation2018). Three different configurations of the digestive tract were observed by Devaere et al. (Citation2014) in the Orthothecid Conotheca subcurvata Yu, Citation1974 from France: ‘(1) anal tube and gut parallel, straight to slightly undulating; (2) anal tube straight and loosely folded gut; and (3) anal tube straight and gut straight with local zigzag folds’ (Devaere et al. Citation2014, p. 1). Devaere et al. (Citation2014) measured the diameter of the conch of C. subcurvata (micro-sized hyoliths preserved the guts, with the size about 100 − 300 μm in diameter) to investigate whether the digestive tract changed morphology in relation to growth, concluding that the number of intestinal loops increased with the growth of the conch (Devaere et al. Citation2014, fig. 10). Recently, Berg-Madsen et al. (Citation2018) documented the orthothecid hyolith Circotheca johnstrupi (the most complete specimen showed that the conch was about 42 mm in length and the apertural diameter was 7 mm) from the lower Cambrian of Bornholm with a highly convoluted digestive tract folded into a chevron-like structure. Considering the large diameter and remarkably uniform size of the folds, this morphology suggests an adult individual, lending support to the proposal by Devaere et al. (Citation2014). Until now, publications regarding the digestive system of orthothecid hyoliths found after Cambrian Stage 3, predominantly show a sinuously looped (zigzag) intestine, coupled with a straight, tapering rectum (Thoral Citation1935; Runnegar et al. Citation1975; Kruse Citation1996; Berg-Madsen et al. Citation2018). However, most specimens in our material of Triplicatella opimus from the Chengjiang Lagerstätte, which preserve the intestine, with the operculum size ranging 5–6 mm (width) and 2.9–3.2 mm (length) and conch size ranging 9–15 mm (length) and 5–7 mm (width) (see Supplementary material), show that the guts have a regular morphology consisting of a straight anal tube and a loosely folded gut ((e-g),5,8). According to Devaere et al. (Citation2014), regardless of the size, this configuration may represent an intermediate stage in the development in orthothecid hyoliths. The loosely folded digestive tract of orthothecids is rarely reported, and the gut of T. opimus here is broadly similar to that described by Devaere et al. (Citation2014) from the Terrenuvian C. subcurvata. Although the conch size for considering the ontology of hyoliths cannot be excluded either due to the relatively small number of orthothecid specimens with preserved digestive tracts. The more loosely folded gut in T. opimus from Cambrian Stage 3, compared with the more tightly folded guts of later orthothecids, could be interpreted as evidence for support of the increased complexity of orthothecid guts with time as suggested by Devaere et al. (Citation2014).
The feeding strategy of Triplicatella (Orthothecid)
Despite the spate of recent discoveries (e.g. Moysiuk et al. Citation2017; Sun et al. Citation2018; Liu et al. Citation2020), the feeding strategy of the extinct Hyolitha remains controversial. Historically speaking, the most convincing arguments regarding the feeding strategy of hyoliths have been derived from the morphology of the digestive system (Thoral Citation1935; Butterfield Citation2001, Citation2003; Malinky Citation2003; Martí Mus Citation2016). The digestive tract of hyolithids was thin, relatively long and shows a flexible U-shaped, loose configuration which is often regarded as characteristic of filter or suspension feeders that ingested soft particles (Babcock and Robison Citation1988; Butterfield Citation2001, Citation2003; Martí Mus Citation2016). As discussed above, this contrasts sharply with the sediment-filled, tightly packed and convoluted guts observed in orthothecids (Kruse 1996; Devaere et al. Citation2014; Berg-Madsen et al. Citation2018). Marek et al. 1997b discussed the differences in functional morphology between hyolithids and orthothecids based on experimental flume studies. The morphology of the hyolithid conch together with the possession of helens appeared to strengthen the hydrodynamic stability of the individual from ambient currents (Marek et al. Citation1997b). The extension of the ligula and the associated conical shape of the operculum in hyolithids increased flow velocity, focusing the amount of suspended material, favouring hyolithids as suspension feeders, an effect which was not observed for orthothecid conchs with retractable opercula (Marek et al. Citation1997b).
Recently, however, the documentation of the hyolithid Haplophrentis from the Burgess Shale possessing a simple U-shaped digestive tract and an extendable gullwing-shaped, tentacle-bearing organ has added substantially to the feeding strategy debate. The presence of this tentacular feeding organ provided further evidence to support an epibenthic, suspension feeding habit for hyolithids (Moysiuk et al. Citation2017). The distinct morphological differences between the digestive system of hyolithids and orthothecids, however, cannot be ignored and most likely reflect different feeding strategies.
The typical gut for a deposit-feeding strategy was discussed by Lopez et al. (Citation1989), who demonstrated that a straight tube in which little axial mixing occurs is probably the most common digestive tract in animals that are deposit feeders. Although a clear distinction between deposit- and suspension-feeding strategies based on digestive systems may not be feasible (Taghon et al. Citation1980), deposit feeders (as sediment microbial strippers) usually increase the surface area of the gut, through, for example, extensive folding, to increase the rate of absorption (Fenchel Citation1970; Lopez et al. Citation1989). With the discoveries of the sediment-filled guts in several orthothecid taxa, evidence for the feeding strategy of the orthothecid hyoliths generally point towards deposit feeding.
Triplicatella was previously thought to be related to orthothecid hyoliths (Skovsted et al. Citation2004, Citation2014). Recently, the tentacular feeding apparatus of Triplicatella opimus from the Chengjiang biota with a tuft-like clustered configuration of tentacles was regarded as a specialised feeding organ, used for collecting food particles directly from the sediment (Liu et al. Citation2020). The exceptionally preserved digestive system documented herein provides new anatomical evidence to reconstruct the internal structure of Triplicatella. The gut of Triplicatella opimus forms spiral loops folded into a loose chevron-like structure with a slightly recurved to straight anal tube. As discussed above, a chevron-like gut is strong evidence that the organ was adapted to increase absorption of nutrients mixed with sedimentary particles. The new information on the anatomy of T. opimus demonstrates unequivocally that T. opimus was an orthothecid hyolith and a deposit feeder. The morphology of the broad flattened ventral surface on the conch and the lack of helens suggests that Triplicatella reclined on the seafloor (Liu et al. Citation2020), probably indicating a surface microbe feeding mode of life as defined by Lopez et al. (Citation1989). The accordion-shape of the gut in T. opimus, we argue, also helps to explain the discrepant arrangement of the tentacles compared with Haplophrentis, that possessed a feeding apparatus adapted to a suspension-feeding strategy.
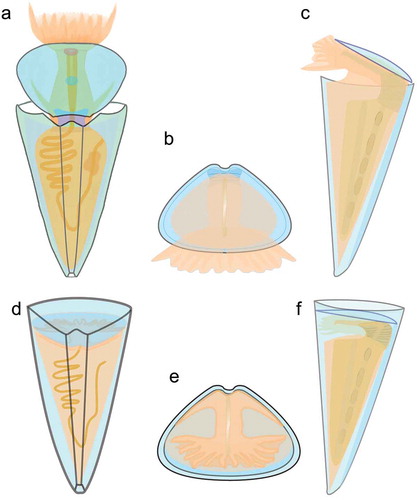
Based on the new information obtained recently on the morphology of Triplicatella opimus (Liu et al. Citation2020 and herein), a reconstruction of the genus, showing both the external morphology and known internal characters, is given in . This reconstruction of Triplicatella may serve as a model for the ancestral morphology of orthothecids and as a starting point for further research on the evolution of the typical orthothecid body plan as defined by Marek (Citation1966).
Figure 8. Anatomical reconstruction of Triplicatella opimus from the Chengjiang Lagerstätte. a, d. A dorsal profile. b, e. A frontal profile, with the operculum. c, f. A left lateral profile
Conclusions
The hyolith Triplicatella opimus is systematically revised for the first time based on exceptionally preserved specimens in mudstone from the Chengjiang Biota, South China. The material includes complete specimens of Triplicatella opimus (conch associated with operculum) and preserved soft parts and adds further evidence to support the classification of Triplicatella as an orthothecid hyolith. Through a series of technical methods, new internal anatomical structures in Triplicatella were revealed and provides significant new evidence concerning the general reconstruction of hyolith anatomy, especially concerning the digestive system of orthothecid hyoliths. The gut of T. opimus as described here consists of a spiral loop folded into a chevron-like structure and a slightly recurved to straight anal tube. The gut is preserved as a reddish-brown trace enriched in iron, which conforms to the principal mode of preservation of nonmineralized tissues in the Chengjiang Lagerstätten. These observations, in combination with the previous records of characteristic orthothecid features and a specialised feeding organ, indicate that T. opimus was a deposit feeder. The anatomical features of T. opimus from the Chengjiang biota will allow us to achieve a complete descriptive basis for the anatomical morphology of orthothecid hyoliths, indicating the possibility that Triplicatella could be used as a model for the ancestral morphology of orthothecids.
Data accessibility
All the specimens dealt with in this paper are deposited in the Early Life Institute and Department of Geology (http://geology.nwu.edu.cn/). Correspondence and requests for materials should be addressed to ZZF ([email protected]).
supplemental_material-revised.xlsx
Download MS Excel (29.5 KB)Acknowledgments
We warmly thank Drs. Zhang, X.L. Han, J. and Liu, J.N. at Northwest University of Xi’an for their kind help with the field trip and suggestions. Thanks are also due to Zhai J.P. and Cheng M.R. for fossil preparation and Sun J. for Micro-CT technical assistance in the Shaanxi Key Laboratory of Early Life and Environments, Northwest University of Xi’an. This work was supported from the National Natural Science Foundation of China [grants 41890844, 41425008, 41621003 and 41720104002 to ZZF, 41772002 to CBS] and Overseas Expertise Introduction Centre for Discipline Innovation [111 Center: D17013] for the continuous fossil collections of Xi’an group is sincerely acknowledged. This work was also funded by the Swedish Research Council to CBS [VR2016-04610] and TPT [VR2017-05183] and Young Thousand Talents Plan of China to TPT [41720104002]. LF is supported by the China Scholarship Council [No. 201906970019] for a one-year research stay with CBS at the Swedish Museum of Natural History. Many thanks to the two anonymous referees for constructive suggestions that have greatly improved the manuscript.
Disclosure statement
We declare that we do not have any commercial or associated interest that represents a conflict of interest in connection with the work submitted.
Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
Related Research Data
References
- ForchielliA, Steiner M, Kasbohm J, Hu S, Keupp H. 2014. Taphonomic traits of clay-hosted early Cambrian Burgess Shale-type fossil Lagerstätten in South China. Palaeogeogr Palaeoclimatol Palaeoecol. 398:59–85. doi:https://doi.org/10.1016/j.palaeo.2013.08.001.
- KouchinskyA, Bengtson S, Clausen S, Vendrasco MJ. 2015. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontol Pol. 60(2):421–513.
- EnglishAM, Babcock LE. 2010. Census of the Indian Springs Lagerstätte, Poleta Formation (Cambrian), western Nevada, USA. Palaeogeogr Palaeoclimatol Palaeoecol. 295(1–2):236–244. doi:https://doi.org/10.1016/j.palaeo.2010.05.041.
- YatesAM, Gowlett-Holmes KL, Mchenry BJ. 1992. Triplicatella disdoma Conway Morris, 1990, reinterpreted as the earliest known polyplacophoran. J Malacol Soc Aus. 13:71.
- PanB, Skovsted CB, Sun HJ, Li GX. 2019. Early Cambrian Hyoliths from the North China Platform and their biostratigraphic and palaeogeographic significance. Alcheringa. 43:351–380. doi:https://doi.org/10.1080/03115518.2019.1577492.
- RunnegarB, Pojeta J, Morris NJ, Taylor JD, Taylor ME, Mcclung G. 1975. Biology of the hyolitha. Lethaia. 8(2):181–191. doi:https://doi.org/10.1111/j.1502-3931.1975.tb01311.x.
- SkovstedCB, Peel JS. 2007. Small shelly fossils from the argillaceous facies of the Lower Cambrian Forteau Formation of western Newfoundland. Acta Palaeontol Pol. 52(4):729–748.
- SkovstedCB, Peel JS, Atkins CJ. 2004. The problematic fossil Triplicatella from the early Cambrian of Greenland, Canada, and Siberia. Can J Earth Sci. 41(11):1273–1283. doi:https://doi.org/10.1139/e04-066.
- SkovstedCB, Topper TP, Betts MJ, Brock GA. 2014. Associated conchs and opercula of Triplicatella disdoma (Hyolitha) from the early Cambrian of South Australia. Alcheringa Aus J Palaeontol. 38(1):148–153. doi:https://doi.org/10.1080/03115518.2014.843381.
- MosesCO, Herman JS. 1991. Pyrite oxidation at circumneutral pH. Geochim Cosmochim Acta. 55(2):471–482. doi:https://doi.org/10.1016/0016-7037(91)90005-P.
- BriggsDEG. 2003. The role of decay and mineralization in the preservation of soft-bodied fossils. Annu Rev Earth Planet Sci. 31(1):275–301. doi:https://doi.org/10.1146/annurev.earth.31.100901.144746.
- GravestockDI, Alexander EM, Demidenko Y-E, et al. 2001. Geological setting. In Alexander E M, Jago J B, Rozanov A Y, et al editors. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Transaction of the Palaeontological Institute 282; pp. 1–344.
- FuDJ, Zhang XL, Budd GE, Liu W, Pan XY. 2014. Ontogeny and dimorphism of Isoxys auritus (Arthropoda) from the early Cambrian Chengjiang biota, South China. Gondwana Res. 25(3):975–982. doi:https://doi.org/10.1016/j.gr.2013.06.007.
- LopezG, Taghon GL, Levinton JS. 1989. Ecology of marine deposit feeders. Lecture notes on coastal and estuarine studies (USA). p. 1–319.
- TaghonGL, Nowell AR, Jumars PA. 1980. Induction of suspension feeding in spionid polychaetes by high particulate fluxes. Science. 210(4469):562–564. doi:https://doi.org/10.1126/science.210.4469.562.
- SunHJ, Babcock LE, Peng J, Zhao YL. 2016. Three-dimensionally preserved digestive systems of two Cambrian hyolithides (Hyolitha). Bull Geosci. 91(1):51–56. doi:https://doi.org/10.3140/bull.geosci.1590.
- DzikJ. 1978. Larval development of hyolithids. Lethaia. 11(4):293–299. doi:https://doi.org/10.1111/j.1502-3931.1978.tb01884.x.
- HanJ, Shu DG, Zhang ZF, Liu J, Zhang XL, Yao Y. 2006. Preliminary notes on soft-bodied fossil concentrations from the Early Cambrian Chengjiang deposits. Chin Sci Bull. 51(20):565–574. doi:https://doi.org/10.1007/s11434-005-2151-0.
- MoysiukJ, Smith MR, Caron JB. 2017. Hyoliths are Palaeozoic lophophorates. Nature. 541:394–397. doi:https://doi.org/10.1038/nature20804.
- MalinkyJM. 2003. Ordovician and Silurian hyoliths and gastropods reassigned from the Hyolitha from the Girvan district, Scotland. J Paleontol. 77(4):625–645. doi:https://doi.org/10.1017/S0022336000044383.
- MalinkyJM, Yochelson EL. 2007. On the systematic position of the Hyolitha (Kingdom Animalia). Memoir Assoc Aus Paleontol. 34:521.
- ChenJY. 2004. The dawn of animal world. Nanjing (China): Jiangsu Science and Technology Press. in Chinese with English summary; p. 1–366.
- DevaereL, Clausen S, Álvaro JJ, Peel JS, Vachard D. 2014. Terreneuvian orthothecid (Hyolitha) digestive tracts from northern Montagne Noire, France; taphonomic, ontogenetic and phylogenetic implications. PLoS One. 9(2):e88583. doi:https://doi.org/10.1371/journal.pone.0088583.
- MarekL. 1966. New hyolithid genera from the Ordovician of Bohemia. Casopis Narodniho Muzea. 153:89–92.
- MarekL. 1967. The class Hyolitha in the Caradoc of Bohemia. Sborník Geologickych Ved, Rada Paleontologie. 9:51–113.
- MarekL, Malinky JM, Geyer G. 1997a. Middle Cambrian fossils from Tizi n’Tichka, the High Atlas, Morocco. Part 2. Hyolitha. J Paleontol. 71(4):638–656. doi:https://doi.org/10.1017/S0022336000040129.
- MarekL, Parsley RL, Galle A. 1997b. Functional morphology of hyoliths based on flume studies. Vestník Ceského Geologického Ústavu. 72(4):277–283.
- MarekL, Yochelson EL. 1964. Paleozoic mollusk: hyolithes. Science. 146(3652):1674–1675. doi:https://doi.org/10.1126/science.146.3652.1674.
- MarekL, Yochelson EL. 1976. Aspects of the biology of Hyolitha (Mollusca). Lethaia. 9(1):65–82. doi:https://doi.org/10.1111/j.1502-3931.1976.tb00952.x.
- BabcockLE, Robison RA 1988. Taxonomy and paleobiology of some Middle Cambrian Scenella (Cnidaria) and Hyolithids (Mollusca) from western North America. The University of Kansas Paleontological Contributions, Paper 121, 1–22.
- Liu F, Skovsted CB, Topper TP, Zhang ZFShu DG. 2020. Are hyoliths Palaeozoic lophophorates?. National Science Review. 7(2):453–469. doi:https://doi.org/10.1093/nsr/nwz161.
- Martí MusM. 2016. A hyolithid with preserved soft parts from the Ordovician Fezouata Konservat-Lagerstätte of Morocco. Palaeogeogr Palaeoclimatol Palaeoecol. 460:122–129. doi:https://doi.org/10.1016/j.palaeo.2016.04.048.
- Martí MusM, Bergström J. 2005. The morphology of hyolithids and its functional implications. Palaeontology. 48:1139–1167. doi:https://doi.org/10.1111/j.1475-4983.2005.00511.x
- Martí MusM, Jeppsson L, Malinky JM. 2014. A complete reconstruction of the hyolithid skeleton. J Paleontol. 88:160–170. doi:https://doi.org/10.1666/13-038
- ThoralM 1935. Contribution à l’étude paléontologique de l’ordovicien inférieur de la Montagne Noire et révision sommaire de la faune cambrienne de la Montagne Noire, par Marcel Thoral, … la Charit.
- Malinky JM,Skovsted CB. 2004. Hyoliths and small shelly fossils from the lower Cambrian of North-East Greenland. Acta Palaeontologica Polonica. 49(4):551–578.
- ZhuMY, Babcock LE, Steiner M. 2005. Fossilization modes in the Chengjiang Lagerstätte (Cambrian of China): testing the roles of organic preservation and diagenetic alteration in exceptional preservation. Palaeogeogr Palaeoclimatol Palaeoecol. 220(1–2):31–46. doi:https://doi.org/10.1016/j.palaeo.2003.03.001.
- ButterfieldNJ. 2001. Cambrian food webs. 40–43. In Briggs DEG. and Crowther PR, editors. Palaeobiology II, a synthesis. Oxford: Blackwell Scientific; pp. 583.
- ButterfieldNJ. 2003. Exceptional fossil preservation and the Cambrian explosion. Integr Comp Biol. 43:166–177. doi:https://doi.org/10.1093/icb/43.1.166.
- MeshkovaNP, Sysoev VA. 1981. Nakhodka slepkov pishchevapitel’nogo apparata nizhnekembrieskikh khiolitov [Discovery of traces of the digestive system in Lower Cambrian hyoliths]”. Institut Geologii I Geofiziki Sibirskogo Otdeleniya AN SSSR, Trudy. 481:82–85.
- KrusePD. 1996. Hyolith guts in the Cambrian of northern Australia‐turning hyolithomorphs upside down. Lethaia. 29(3):213–217. doi:https://doi.org/10.1111/j.1502-3931.1996.tb01651.x.
- LittkeR, Klussmann U, Krooss B, Leythaeuser D. 1991. Quantification of loss of calcite, pyrite, and organic matter due to weathering of Toarcian black shales and effects on kerogen and bitumen characteristics. Geochim Cosmochim Acta. 55(11):3369–3378. doi:https://doi.org/10.1016/0016-7037(91)90494-P.
- TateR. 1892. The Cambrian fossils of South Australia. Transac Royal Soc South Aus. 15:183–189.
- WronaR. 2003. Early Cambrian molluscs from glacial erratics of King George Island, west Antarctica. Polish Polar Res. 24(3–4):181–216.
- HoubrickRS, Stürmer W, Yochelson EL. 1988. Rare mollusca from the Lower Devonian Hunsrück slate of southern Germany. Lethaia. 21(4):395–402. doi:https://doi.org/10.1111/j.1502-3931.1988.tb01768.x.
- BengtsonS, Conway Morris S, Cooper BJ, Jell PA, Runnegar BN. 1990. Early Cambrian fossils from South Australia. Memoir Assoc Aus Paleontol. 9:1–364.
- ClausenS, Álvaro JJ. 2006. Skeletonized microfossils from the Lower-Middle Cambrian transition of the Cantabrian Mountains, northern Spain. Acta Palaeontol Pol. 51(2):223–248.
- Conway MorrisS. 1993. The fossil record and the early evolution of the Metazoa. Nature. 361(6409):219–225. doi:https://doi.org/10.1038/361219a0.
- GabbottSE, Hou XG, Norry MJ, Siveter DJ. 2004. Preservation of Early Cambrian animals of the Chengjiang biota. Geology. 32(10):901–904. doi:https://doi.org/10.1130/G20640.1.
- Sun HJ, Smith MR, Zeng H, Zhao FC, Li GX,Zhu MY. 2018. Hyoliths with pedicles illuminate the origin of the brachiopod body plan. Proceedings of the Royal Society B: Biological Sciences, 285(1887): 20181780. doi: https://doi.org/10.1098/rspb.2018.1780
- FenchelT. 1970. Studies on the decomposition of organic detritus derived from the turtle grass Thalassia testudinum 1. Limnol Oceanogr. 15(1):14–20. doi:https://doi.org/10.4319/lo.1970.15.1.0014.
- Berg-MadsenV, Valent M, Ebbestad JOR. 2018. An orthothecid hyolith with a digestive tract from the early Cambrian of Bornholm, Denmark. GFF. 140(1):25–37. doi:https://doi.org/10.1080/11035897.2018.1432680.
- YuW. 1974. Cambrian hyolithids. In: Handbook of Stratigraphy and palaeontology of Southwest China, editor. Nanjing Institute of Geology and Paleontology. Beijing: Academia Sinica, Science Press; p. 111–112.
- Hou XG, Siveter DJ, Siveter DJ, Aldridge RJ, Cong PY, Gabbott SE, Ma XY, Purnell MA, Williams M. 2017. The Cambrian fossils of Chengjiang, China: the flowering of early animal life. New York: Wiley‐Blackwel. p. 1–316.
- HouXG, Aldridge RJ, Bergström J, Siveter DJ, Siveter DJ, Feng X. 2004. The Cambrian fossils of Chengjiang, China: the flowering of early animal life. Oxford: Blackwell Publishing, p. 233.
- HouXG, Bergström J, Wang HF, Feng XH, Chen AL. 1999. The Chengjiang Fauna: exceptionally well-preserved animals from 530 million years ago. Kunming: Yunnan Science and Technology Press. In Chinese with English summary; p. 1–80.
- ChenXY, Zhao YL, Wang PL. 2003. Preliminary research on hyolithids from the Kaili Biota, Guizhou. Acta Micropalaeontologica Sinica. 21(3):296–302.
- ZhaoYL, Zhu MY, Babcock LE, Yuan JL, Parsley RL, Peng J,Yang XL, Wang Y. 2005. Kaili Biota: a taphonomic window on diversification of metazoans from the Basal Middle Cambrian: Guizhou, China. Acta Geologica Sinica. 79(6):751–765. doi:https://doi.org/10.1111/j.1755-6724.2005.tb00928.x.