
ABSTRACT
Knowledge of Neogene fish diversity in Taiwan is extremely limited. In this paper, we present a collection of 1716 fish otoliths recovered from the late Miocene Tapu Formation. The abundance and density of otoliths vary across the sites. Although the preservation of samples is considerably limited, our sample coverage is sufficient and reveals the presence of at least 34 otolith-based taxa belonging to 13 families. Four new species are introduced: Larimichthys koae sp. nov., Nibea chaoi sp. nov., Taosciaena jiangi sp. nov., and T. hui sp. nov. Among the earliest fossil records, this collection features a remarkable abundance of Larimichthys spp. The assemblage is dominated by otoliths of Sciaenidae, Gobiidae and Soleidae, and it is indicative of a coastal shallow-water palaeoenvironment with muddy to sandy bottoms, perhaps adjacent to a river mouth. We hypothesise that the differences in the taxonomic composition between the fossil assemblage and modern fauna are mainly chronological and evolutionary, and only a moderate turnover of certain lineages has occurred since the Miocene. Our study provides a unique ‘window’ to a rarely visited fossil fish community that exclusively documented by fossil otolith data, and reveals a past coastal fish diversity that otherwise could not be recognised.
Introduction
Fish diversity and composition play important roles in structuring modern marine communities. Therefore, understanding their roles in the geological past can provide insights into how fish have shaped ancient and modern marine diversity patterns (Renema et al. Citation2008). Fish diversity in the tropical–subtropical west Pacific is regarded as one of the critical biodiversity hotspots; however, little is known about the historical process that structured the modern fish fauna often because lacking adequate, relevant fossil records (Lin et al. Citation2019a). Fossil fish otoliths provide a unique opportunity for investigating this research topic due to their high abundance and nearly ubiquitous occurrence both in time and space (Nolf Citation1985, Citation2013). For example, the ancient Kuroshio Current potentially shaped the fish fauna of the Pleistocene (Mitsui et al. Citation2021).
However, fossil otolith records are yet to be fully explored, particularly those in the tropical and subtropical Indo-west Pacific, where systematic research has rarely been conducted (Nolf Citation2013; Lin et al. Citation2019a). Related otolith studies are currently restricted to Japan; although numerous papers have been published, the corresponding studies appear to have been small in scope, as each paper has reported a single horizon or outcrop. More recent papers include, but are not limited to, those on Neogene to Pleistocene assemblages (Miyata et al. Citation2018; Ohe and Watanabe Citation2018; Ohe et al. Citation2020; Mitsui et al. Citation2021); a review of Plio-Pleistocene Myctophidae otoliths (Schwarzhans and Ohe Citation2019); and a paper on the early Oligocene assemblage in Kyushu (Schwarzhans et al. Citation2017). This last paper can be regarded as the only Palaeogene record from the entire west Pacific. An additional report from the Pliocene Philippines by Van Hinsbergh and Helwerda (Citation2019) is available.
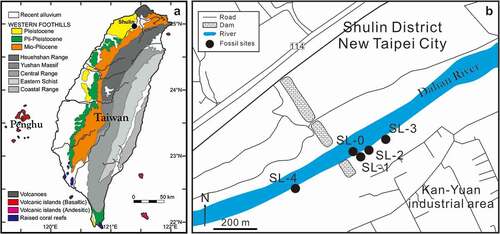
In subtropical Taiwan, studies on fossil otoliths are in the initial stage, and systematic sampling of otoliths is yet to be conducted (Lin et al. Citation2019a). Lin et al. (Citation2018b) documented some early Pleistocene otoliths from southwestern Taiwan, but only surface collection was performed. Subsequently, a similar museum collection from the Chochen–Tsailiao locality, southern Taiwan, was reported (Lin et al. Citation2019a). The materials in this collection only provided an approximate age; the exact horizons were not lodged during sampling. Data on the Neogene otoliths of Taiwan were reported by Lin (Citation2010b); he first mentioned a small outcrop along the Dahan River () and reported approximately 50 surface-collected otoliths with many erroneous identifications. The formation that yielded otoliths in the study by Lin (Citation2010b) was erroneously indicated as being the younger Erchiu Formation, and the same was indicated in the study by Lin et al. (Citation2019a); these errors must be corrected (see below, Geological setting). Further quantitative sampling from the same area was undertaken by one of the authors of the current paper (CHL) between 2013 and 2020. This new collection along with that of Lin (Citation2010b) provided abundant material, which is described here. The present paper aims to understand the Neogene subtropical coastal fish community with very limited knowledge so far from Taiwan and demonstrates its potential to reveal the geological structuring of fish diversity in the west Pacific.
Figure 1. Maps showing the sampling sites. a, geological map of Taiwan with the highlighted Shulin area (black circle); the map was modified after Lin et al. (Citation2019a). b, sampling sites.
Geological setting
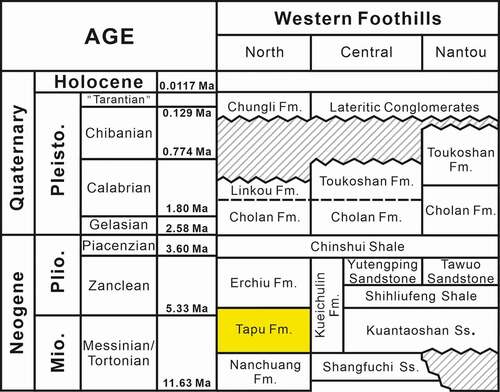
The exposed strata in the study area are the late Miocene Nanchuang Formation and Tapu Formation (). The Nanchuang Formation is composed of thin-layered sandstone–shale interbeds with thick-layered whitish sandstones and thin coalbeds, which are deposited in fluvial and estuarine bay environments (Pan Citation2011). In the Nanchuang Formation, several marked basaltic lava flows, volcanic breccia, and tuff layers of the Chiaopanshan Volcanic Stage are intercalated (Tsao et al. Citation1992). The younger Tapu Formation, equivalent to the lower part of the Kueichulin Formation in the central Western Foothills, is composed of bioturbated thick-layered massive greenish sandstones with thin-layered sandstone–shale interbeds, which are deposited in littoral and shallow marine environments during the transgression stage (Pan Citation2011; Chen and Yu Citation2016). In the sampling area, a thick basaltic tuff layer of c. 2 m was presented between the Nanchuang Formation and Tapu Formation, and the layer formed a cliff along the right bank of the Dahan River (Appendix 1). Equivalent strata of this basaltic tuff were observed in the adjacent area, whose K–Ar ages have been dated as 8–10 Ma (Tsao et al. Citation1992). The samples for this study were collected from the lowest part of the Tapu Formation immediately above the basaltic tuff, and the age was recognised as late Miocene ().
Figure 2. Stratigraphic correlation of the Western Foothills, Taiwan (modified after Chen and Yu Citation2016). The studied formation, the Tapu Formation, is indicated in yellow.
Other associated fossils
The associated marine fossils included both vertebrate and invertebrate specimens. The vertebrate specimens were confined to fish teeth (both teleost and elasmobranch teeth, Lin et al. unpub. data). Various types of mollusc were the most notable, along with sea urchins (possibly Sinaechinocyamus sp.), corals (Dendrophyllia sp., Ribas-Deulofeu and Lin in review), and barnacles. Foraminifera specimens were poorly preserved. All associated marine fossils were maintained, but they are not further discussed here.
Materials and methods
Sampling
The fossil sites are located in the Kan-Yuan industrial area, Shulin, New Taipei City (). Although the sites are all restricted in extent, they have been known to amateur collectors for years. The exposures are located along the right bank of the Dahan River and were high above the water level (10–20 m) during the field work. They were adjacent to an abandoned dam that was washed away by a typhoon in 2004, and since then, the flooded area, including the fossil sites, has been exposed. Bulk sediment samples and surface hand-picked samples were collected. Descriptions and coordinates of each locality are presented.
Site SL-0
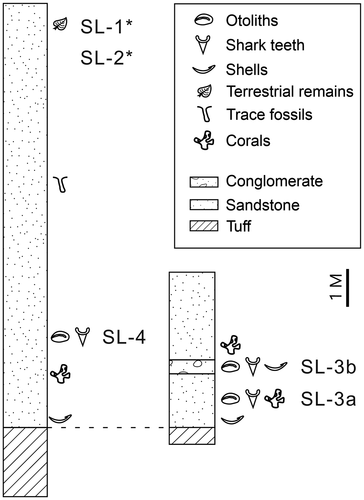
The site was approximately 100 m downstream from the dam. The GPS coordinates were 24°57’38.1“N, 121°23’01.9”E (). This site was the most accessible and yielded the most fish otoliths, shark teeth, and shells. However, the entire site was flushed away in the rainy seasons of 2015–2016. Surface specimens were collected by CHL in 2010 (Lin Citation2010b), and a large bulk sample comprising approximately 23 kg of sediment was collected by CHL in 2013.
Site SL-1
The site is located immediately behind SL-0. Its GPS coordinates are 24°57’37.27’’N, 121°23’00.75’’E (). Approximately 6 kg of sediment was sieved from the site. The sediment contained fragments of coal but no otoliths or other marine fossils. The site correlates to the horizon with terrestrial remains, as shown in .
Figure 3. Measured section of the outcrops in the study area. Sampling horizons from each site are indicated. Asterisks indicate lateral extension of the same horizon.
Site SL-2
This site is located approximately 30 m from SL-1. Its GPS coordinates are 24°57’38.44’’N, 121°23’02.23’’E (). Sediment weighing approximately 5 kg was sieved. The site had an identical horizon to SL-1, and no otoliths were obtained from this site.
Site SL-3 (SL-3a/3b)
This site is located approximately 200 m downstream from the dam. Its GPS coordinates are 24°57’40.25’’N, 121°23’05.18’’E (). The site is located on top of a small bluff of 8-m height along the river (Appendix 1). Two distinct fossil layers were recognised; the lower sampling layer (SL-3a) was immediately below a conglomerate horizon (mixed with tuffaceous material) and just above the basal tuff, and the upper layer (SL-3b) was within the conglomerate horizon (). The lower layer SL-3a was a lateral equivalent of SL-4, whereas the upper layer SL-3b of conglomerate contained numerous recrystallised molluscs and gravel of various sizes, suggesting sediment reworking and transport. This conglomerate horizon was locally restricted and was not found in SL-4 (). The basal tuff did not contain any fossils. Seven and five bulk samples from SL-3a and SL-3b, respectively, were collected by CHL in 2019–2020 ().
Table 1. Summary of samples
Site SL-4
This site is located approximately 220 m upstream from the dam. Its GPS coordinates are 24°57’32.82’’N, 121°22’50.75’’E (). The site is in a small open area approximately 20-m long, and the horizon yielding fossils is the widest among all the sites (Appendix 1). A measured section of the site is illustrated in . Twenty-one bulk samples weighing c. 213 kg were collected by CHL in 2020 ().
Surface collection
Additional specimens were collected from the surface of exposures (above sites) during several field surveys. Surface specimens were not localised and were collected by several collectors. These are included in the description but are not further analysed in diversity analyses.
Otolith extraction, images, and identification
Bulk sediments were first dried and weighed. Each sample contained approximately 10 kg of sediment, except for the large sample obtained from the SL-0 site (). The sediment was then soaked in water before being sieved through a 500-μm mesh screen. All otoliths above this 500-μm screen were hand-picked under a stereomicroscope (SMZ1270, Nikon Instruments Inc., Tokyo, Japan), and stacking images were taken. For incomplete otolith, specimens of at least one-half complete were analysed further.
To aid otolith identification, relevant literature such as Ohe (Citation1985), Yu and Shen (Citation1987), Schwarzhans (Citation1993, Citation1999), Iizuka and Katayama (Citation2008), Lin and Chang (Citation2012), and Nolf (Citation2013) were consulted. In addition, recent otolith reference collections including those deposited at the National Museum of Marine Biology and Aquarium (NMMBA and Global Sciaenidae Conservation Network, GSCN, https://croakers.biodiv.tw) and our in-house laboratory collection (under registration code CHLOL) were directly compared. Whenever possible, otoliths were identified at the species level. All the materials are stored in the Biodiversity Research Museum, Academia Sinica, Taiwan (BRMAS, http://museum.biodiv.tw/eng), and all imaged specimens are stored under the registration code ASIZF.
Diversity analysis
Subsequent analyses involved bulk samples. The density of otoliths in the sediments was calculated as the otolith count divided by the dry sediment weight (kg) for each sample, and the preservation status was roughly indicated by the identification rate (number of identified specimens divided by total number of otoliths in a sample). These are illustrated in box plots. Sample-based rarefaction and extrapolation with 95% confidence intervals were calculated using EstimateS software (Colwell Citation2013) and the rank abundance of identified taxa in each site are presented to indicate the taxonomic composition.
Systematic palaeontology
A list of taxa and their abundance recovered from the collection are presented in . Otoliths were identified as belonging to 13 families and 34 taxa. The classification scheme followed that of Nelson et al. (Citation2016). For the authorships of higher taxa, the method of Van Der Laan et al. (Citation2014) was employed, whereas authorships for genus and species, Fricke et al. (Citation2021) were followed. In the descriptions, the otolith refers to the sagittal (saccular) otolith unless otherwise indicated (i.e. lapillus (utricular otolith) in Ariidae). The general otolith terminology followed that of Nolf (Citation1985, Citation2013), and for utricular otolith of Ariidae, those of Ohe (Citation2006) and Aguilera et al. (Citation2013) were followed. The synonymy list, if any, is restricted to relevant records from Taiwan. Based on published images, related otolith fossils from Japan are only listed here with a “?” in front to indicate their tentative nature and are subject to detailed revisions in the future.
Table 2. List of otolith-based fish taxa from the late Miocene Shulin, northern Taiwan
Figure 4. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–f, Ariosoma sp.; a, SL-3a, ASIZF 0100001; b–d, SL-0, ASIZF 0100002–04; e, f, SL-4, ASIZF 0100005; e, ventral view. g, h, Clupeidae indet., SL-4, ASIZF 0100006; g, ventral view. i–k, Thryssa sp.; i, SL-3b, ASIZF 0100007; j, k, SL-0, ASIZF 0100008.
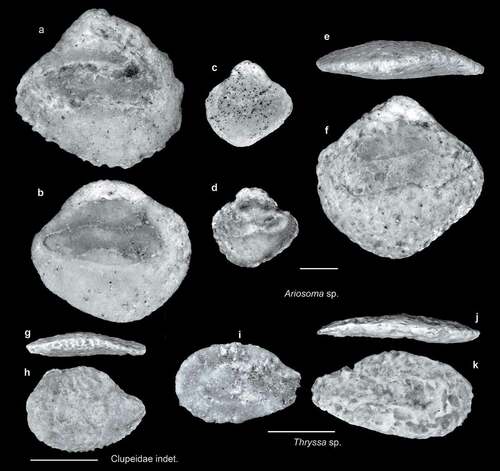
Figure 5. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–g, Arius sp., lapilli; a, b, SL-3b, ASIZF 0100009; c–g, SL-0, ASIZF 0100010–11; a, c, g, dorsal views; b, e, f, ventral views. h–j, Netuma sp., lapilli, SL-0, ASIZF 0100012; h, dorsal view; j, ventral view. k, l, Ostorhinchus sp., SL-0, ASIZF 0100013; k, ventral view. m, n, Apogonidae indet., SL-0, ASIZF 0100014–15. o, p, Sillago sp., SL-3a, ASIZF 0100016; o, ventral view.

Figure 6. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views. a–f, Parachaeturichthys sp., ASIZF 01000017–22; a, d, SL-4; b, SL-0; c, e, SL-3a; f, SL-3b. g–m, Gobiidae indet. 1, ASIZF 01000023–29; g, i, m, SL-4; h, SL-0; j, l, SL-3b; k, SL-3a.
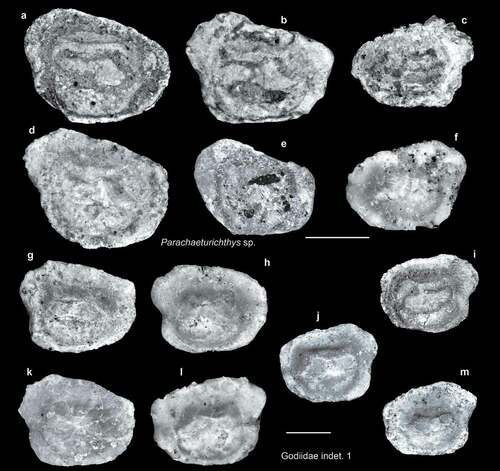
Figure 7. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views. a, b, Gobiidae indet. 2, ASIZF 01000030–31; a, SL-3b; b, SL-4. c, d, Gobiidae indet. 3, SL-4, ASIZF 01000032–33. e, f, Gobiidae indet. 4, ASIZF 01000034–35; e, SL-3b; f, SL-3a. g, Gobiidae indet. 5, SL-4, ASIZF 01000036. h–l, Pardachirus sp. 1, ASIZF 01000037–41; h, SL-0; i–k, SL-4; l, SL-3b.

Figure 8. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views. a–c, Pardachirus sp. 2, ASIZF 01000042–44; a, b, SL-4; c, SL-0. d, e, Tarphops oligolepis (Bleeker, 1858), ASIZF 01000045–46; d, SL-4; e, SL-0. f, g, Cynoglossus sp., ASIZF 01000047–48; f, SL-3a; g, SL-4.
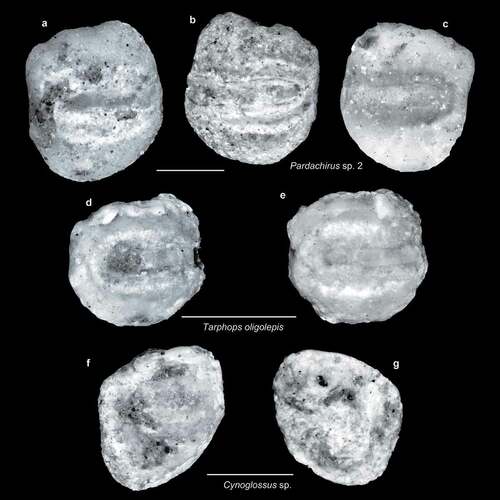
Figure 9. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–c, Uranoscopus sp.; a, SL-0, ASIZF 01000049; b, c, SL-4, ASIZF 01000050; b, ventral view. d–f, Pomadasys sp.; d, SL-3a, ASIZF 01000051; e, f, SL-4, ASIZF 01000052; e, ventral view. g–i, Argyrosomus sp., SL-3b, ASIZF 01000088; h, ventral view; i, anterior view.
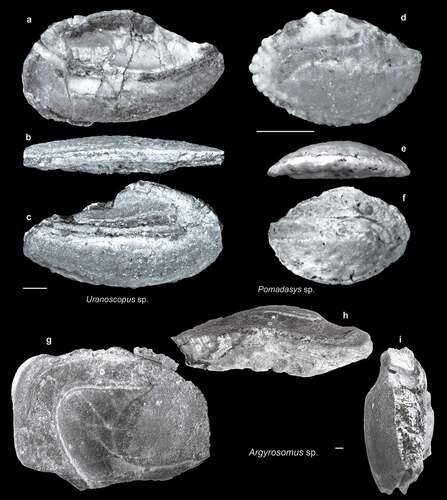
Figure 10. Fish otoliths from the late Miocene northern Taiwan and recent comparative specimens. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–e, Larimichthys crocea (Richardson, 1846); a, b, recent otoliths, sampled from Miaoli, Taiwan; a, 3 March 2020, 261.01 mm SL, CHLOL 000780; b, 2 March 2020, 191.93 mm SL, CHLOL 000772; c, SL-3b, ASIZF 01000053; d, e, SL-0, ASIZF 01000054; d, ventral view. f–i, Larimichthys polyactis (Bleeker, 1877); f, recent otoliths, sampled from Miaoli, Taiwan, 2 March 2020, 139.40 mm SL, CHLOL 000778; g–i, SL-0, g, ASIZF 01000055; h, i, ASIZF 01000056; h, ventral view.
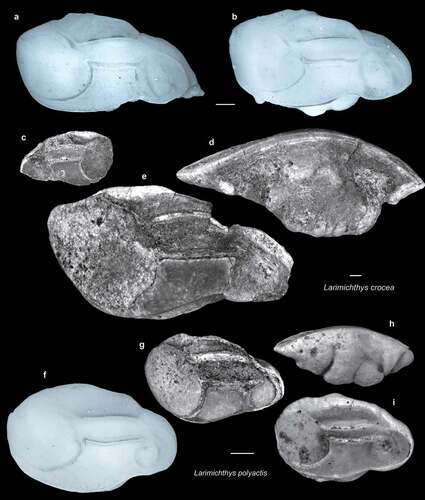
Figure 11. Fish otoliths from the late Miocene northern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–e, Larimichthys koae sp. nov., SL-0; a–c, holotype, ASIZF 01000057; a, ventral view; c, anterior view; d, e, paratypes, ASIZF 01000058–59. f–j, Miichthys miiuy (Basilewsky, 1855); f, g, SL-0, ASIZF 01000060; f, ventral view; h, SL-4, ASIZF 01000061; i, j, SL-0, ASIZF 01000062; i, ventral view.
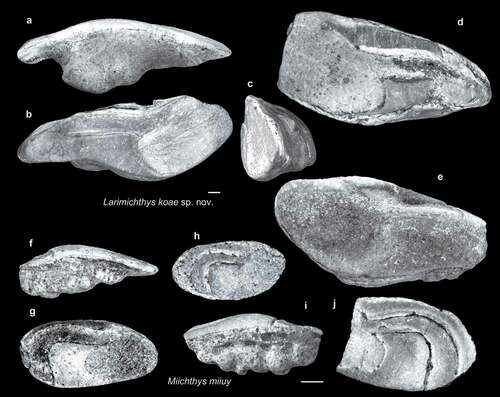
Figure 12. Fish otoliths from the late Miocene northern Taiwan and recent comparative specimens. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–i, Nibea chaoi sp. nov.; a–f, paratypes; a, b, SL-3a, ASIZF 01000063; a, ventral view; c, d, SL-3b, ASIZF 01000064; c, ventral view; e, f, SL-0, ASIZF 01000065–66; g–i, holotype, SL-3a, ASIZF 01000067; g, ventral view; i, anterior view. j–l, Nibea albiflora (Richardson, 1846); j, k, SL-3b, ASIZF 01000068; k, ventral view; l, SL-3a, ASIZF 01000069. m, n, Protonibea diacanthus (Lacepède, 1802), recent otoliths; m, sampled from Miaoli, Taiwan, 20 March 2019, 252.69 mm SL, GSCN 000914; n, sampled from Changhua, Taiwan, 3 August 2019, 352.11 mm SL, GSCN 002094. o, p, Protonibea aff. diacanthus (Lacepède, 1802), SL-3a, ASIZF 01000070; p, ventral view.
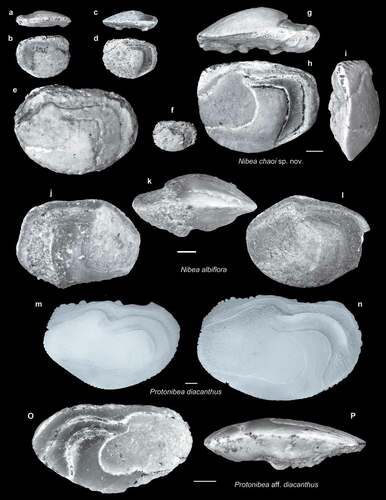
Figure 13. Fish otoliths from the late Miocene northern Taiwan. Scale bar = 1 mm. Images are inner views unless otherwise indicated. a–l, Taosciaena hui sp. nov.; a, b, paratype, SL-3a, ASIZF 01000071; a, ventral view; c–e, holotype, SL-3a, ASIZF 01000072; c, ventral view; d, anterior view; f–l, paratypes; f, SL-4, ASIZF 01000073; g, SL-3a, ASIZF 01000074; h–k, SL-4; h, i, ASIZF 01000075; h, ventral view; j, k, ASIZF 01000076; j, ventral view; l, SL-3a, ASIZF 01000077.

Figure 14. Fish otoliths from the late Miocene northern Taiwan. Scale bar = 1 mm. Images are inner views unless otherwise indicated. a–p, Taosciaena jiangi sp. nov.; a, b, paratypes; a, SL-3a, ASIZF 01000078; b, SL-0, ASIZF 01000079; c, d, holotype, SL-0, ASIZF 01000080; c, ventral view; e–p, paratypes; e, f, SL-0, ASIZF 01000081; e, ventral view; g, SL-3a, ASIZF 01000082; h, SL-4, ASIZF 01000083; i–k, SL-3a, ASIZF 01000084; i, ventral view; k, anterior view; l, m, SL-0, ASIZF 01000085; l, ventral view; n, o, SL-0, ASIZF 01000086; n, ventral view; p, SL-4, ASIZF 01000087.

Order Anguilliformes Regan, 1909
Family Congridae Kaup, 1856
Genus Ariosoma Swainson, 1838
Ariosoma sp. (–f)
2018b Ariosoma sp.; Lin et al., pl. 1, fig. 7 [Taiwan].
Remarks
The otoliths of Ariosoma are adequately recognisable on the basis of their distinct sulcus type (Schwarzhans Citation2019a, pl. 2). Twenty Ariosoma otoliths of various sizes are found in the bulk samples. Their appearance is very similar to that of A. anagoides (Schwarzhans Citation2019a, pl. 2, fig. 2), but our specimens differ from those in the recent collections in that they have a more compact shape and a somewhat more downwards-extended ventral area. Given these features, our otoliths likely belong to an undescribed fossil species, but is left in open nomenclature because of poor preservation.
Order Clupeiformes Bleeker, 1859
Family Engraulidae Gill, 1861
Genus Thryssa Cuvier, 1829
Thryssa sp. (–k)
Remarks
Six otoliths show a reasonable resemblance to those of the recent Thryssa (Lin and Chang Citation2012, pl. 71), but all our specimens are poorly preserved, not allowing more precise assignment.
Family Clupeidae Cuvier, 1816
Clupeidae indet. (, h)
Remarks
Only a single clupeid otolith is found in the collection. Similar to the aforementioned Thryssa otoliths, it is considerably worn and can be assigned to the family level only.
Order Siluriformes Cuvier, 1817
Family Ariidae Bleeker, 1858
Genus Arius Valenciennes, 1840
Arius sp. (–g)
2010b Pennahia argentata; Lin, pl. 17, figs 4, 5 [Taiwan].
2018b Ariidae indet.; Lin et al., pl. 1, figs 8, 9, non fig. 10 (Plicofollis) [Taiwan].
Remarks
The lapilli (utricular otoliths) of Ariidae are robust and thick and can be readily recognised by their triangular shape and smooth margins; therefore, ariid lapilli have higher taxonomic value than do their sagittae, which are small, thin and fragile. Their dorsal face is usually marked by an elevated, central hump (umbo), which is surrounded by visible concentric growth rings. The ventral face is gently convex, and a mesial shallow depression is present on the mesial rim. Three genera are found in Taiwan – Arius, Netuma and Plicofollis – and they can be distinguished based on their distinct lapilli (Ohe Citation2006). The lapilli of Arius are characterised by a strong antero-mesial projection, a marked mesial inward curve, a more gently curved (convex) posterior rim, a concave anterior rim, and a flat mesial rim. The lapilli of Netuma and Plicofollis are similar; they both have straight anterior and posterior rims and an angled mesial rim. However, the lapilli of Netuma have an obtuse postero-mesial rim and a more protruding distal part, and their lapillus width (Lw) is longer than their lapillus length (Ll; see also Ohe Citation2000, pl. 2, fig. 5, as Arius thalassinus), whereas in Plicofollis, a notable angle is present in the postero-mesial rim, and the Lw is approximately the same as Ll (Lin et al. Citation2018b, pl. 1, fig. 10). The preservation of the specimens does not allow further taxonomic assignment; moreover, the taxonomy of the recent Ariidae in Taiwan is yet to be fully explored (Tseng and Lin unpub. data).
Genus Netuma Bleeker, 1858
Netuma sp. (–j)
Remarks
See under Arius sp.
Order Kurtiformes Jordan, 1923
Family Apogonidae Günther, 185
Genus Ostorhinchus Lacepède, 1802
Ostorhinchus sp. (, l)
Remarks
A large, adult apogonid otolith characterised by an elongated shape with a tapering posterior part and a long ostium that is approximately twice as long as the cauda is allocated to the genus Ostorhinchus. More than 20 species of Ostorhinchus are known in Taiwan, and recent comparative references are lacking for many of these species (Lin and Chang Citation2012, labelled as Apogon therein).
Apogonidae indet. (, n)
Remarks
Otoliths of Apogonidae are uncommon in our collection. They are easily recognisable as an apogonid but are often represented as juvenile individuals without diagnostic morphology enabling clear identification. The reference specimens of Apogonidae are insufficient, and otoliths of closely related genera and/or species exhibit similar features, precluding their clear identification.
Order Gobiiformes Günther, 1880
Family Gobiidae Cuvier, 1816
Genus Parachaeturichthys Bleeker, 1874
Parachaeturichthys sp. (–f)
2018b Gobiidae indet.; Lin et al., pl. 2, figs. 1, 2 [Taiwan].
Remarks
The otoliths of this taxon have a large dorsal area that protrudes enormously in the postero-dorsal direction. This enlarged part is a distinct feature that can be seen in the otoliths of Amblychaeturichthys, Chaeturichthys and Parachaeturichthys, but the size of our fossil specimens and the shape of the dorsal rim are similar to only the otoliths of Parachaeturichthys (Ohe Citation1985, p. 132, 133; Ohe et al. Citation2020, fig. 7; Iizuka and Katayama Citation2008, p. 145).
Gobiidae indet. 1–5 (–m, 7a–g)
Remarks
Although not abundant, gobiid otoliths are not uncommon in our material. However, approximately 250 gobiid species are found in Taiwan, and many closely related species exhibit almost identical otolith morphology. Considering that an extensive gobiid otolith reference collection in related areas is lacking and that the specimens available are generally poorly preserved, we decided to take a rather conservative taxonomic approach and restricted the identification of the specimens to otolith morphotypes. Gobiidae indet. 1 represents a group of otoliths that are considered very generalised in morphology; it has a rectangular shape with a slightly enlarged postero-dorsal area but not one as large as that of Parachaeturichthys (see above). A notable notch is present in the middle of the posterior rim. Both Gobiidae indet. 2 and 3 have square otoliths; Gobiidae indet. 2 has a swollen collicular crest on the lower side of the cauda, and its otolith size is usually much larger than that of Gobiidae indet. 3. In Gobiidae indet. 4, the dorsal rim of the otoliths is strongly angled with its pointed tip immediately behind the midline of the otolith. Finally, a single, small otolith with an oblique dorsal rim represents yet another group, namely Gobiidae indet. 5. The trapezoid otolith is shaped by this dorsal rim, and several recent genera superficially share this feature, for instance, Clariger (Ohe et al. Citation2020, fig. 8), Exyrias (Rivaton and Bourret Citation1999, pl. 79, figs 11, 12 but see Lin and Chang Citation2012, pl. 124), and Gymnogobius (Ohe et al. fig. 7).
Order Pleuronectiformes Bleeker, 1859
Family Paralichthyidae Regan, 1910
Genus Tarphops Jordan & Snyder, 1900
Tarphops oligolepis (Bleeker, 1858) (, e)
?1977 Chascanopseta cf. C. lugubris; Ohe, fig. 16 [Japan].
?1981 Solea sp. indet. Round type; Ohe, pl. 9, fig. 19 [Japan].
?1983 Chascanopseta cf. C. lugubris; Ohe, pl. 14, fig. 4 [Japan].
?1983 Engyprosopon ui; Ohe, pl. 14, fig. 6 (upside down figure) [Japan].
Remarks
Three small otoliths with a square shape are assigned to Tarphops oligolepis on the basis of the oblique corners on both the antero-ventral and postero-ventral rims, a nearly straight posterior rim, and a slightly elevated antero-dorsal part. The sulcus is somewhat narrow (), almost reaching the anterior rim, and is filled by a single straight colliculum that does not divide it into the ostium and cauda (Schwarzhans Citation1999, figs 240–243). Previous Neogene materials from Japan require extensive revision if a reliable taxonomic conclusion is to be made.
Family Soleidae Bonaparte, 1833
Genus Pardachirus Günther, 1862
Pardachirus sp. 1 & 2 (–l, 8a–c)
Remarks
Otoliths of Pardachirus are the most abundant among the soleids; they are characterised by small otolith size, an elevated and enlarged antero-dorsal part, and a deep but somewhat pointed ventral area. All the margins are smooth except the dorsal margin, which may be slightly undulated. The sulcus is only slightly divided into an elongate ostium and an oval cauda (Schwarzhans Citation1999, figs 872–875). In the examined material, the specimens can be separated into two plausible species based on the shape of the posterior rim; it is concave and may form a notch in Pardachirus sp. 1, whereas a rather straight posterior rim is present in the second taxon. The ventral area of Pardachirus sp. 1 is also shorter than the second taxon. The otolith reported as Solea sp. (Ohe Citation1983) from the Pliocene Hijikata Formation, Japan, very likely belongs to Pardachirus.
Family Cynoglossidae Jordan, 1888
Genus Cynoglossus Hamilton, 1822
Cynoglossus sp. (, g)
Remarks
The otoliths of Cynoglossus are triangular with an oblique dorsal rim, largely curved anterior and ventral rims, and straight posterior rim; they have a sulcus with a narrow but rod-like ostium and a wide, somewhat triangular cauda. These features are sufficiently derived and can be recognised at the genus level in our material, but the preservation status of the specimens, especially in terms of the sulcus, does not allow a more precise assignment. A Cynoglossus otolith reported by Lin et al. (Citation2019a, fig. 4q, r) from the Plio-Pleistocene Tasiliao River bed, southern Taiwan, appears to represent yet another species.
Order Trachiniformes Rafinesque, 1810
Family Uranoscopidae Bonaparte, 1831
Genus Uranoscopus Jordan & Snyder, 1900
Uranoscopus sp. (–c)
2010b Uranoscopus sp.; Lin, pl. 18, fig. 5 [Taiwan].
Remarks
Three large otoliths from the collection can be allocated to Uranoscopus on the basis of their centrally located, slender, long, and upwards-bent sulcus. All otoliths are broken in some parts. Shape variation of the otoliths, especially the dorsal rim, within species has been reported (Lin Citation2010a; Schwarzhans Citation2019b), and recent taxonomic studies in this group have revealed great species diversity in the west Pacific (Fricke Citation2018; Vilasri et al. Citation2019), indicating that a more extensive reference of Uranoscopidae otoliths is required. The much thicker Uranoscopus otoliths reported by Lin et al. (Citation2018b, pl. 2, figs 3, 4) from the Pleistocene Niubu locality, southern Taiwan, appear to belong to a different species.
Order Perciformes Bleeker, 1859
Suborder Percoidei Bleeker, 1859
Family Haemulidae Gill, 1885
Genus Pomadasys Lacepède, 1802
Pomadasys sp. (–f)
Remarks
Except for one large but broken specimen, all our Pomadasys otoliths are small juveniles. The key features for identifying these otoliths are the configuration of their caudal part (Lin et al. Citation2013), but these parts are often too eroded in our specimens.
Order Acanthuriformes sensu (Nelson et al. Citation2016)
Suborder Sciaenoidei sensu Betancur-R. et al., 2013
Family Sciaenidae Cuvier, 1829
Genus Argyrosomus De la Pylaie, 1835 (–i)
Remarks
One extremely large (Otolith length = 25.64 mm) but broken specimen is assigned to Argyrosomus. The otolith is bullet-shaped though the anterior part is largely lost. The sulcus is most similar to Argyrosomus, with a large ostium that widens in the postero-ventral part and a long and strongly curved cauda. Another key feature is the straight and vertical posterior rim of the otolith immediately after a marked postero-dorsal angle (Schwarzhans Citation1993, figs 251–267).
Genus Larimichthys Jordan & Starks, 1905
Remarks
The genus Larimichthys is characterised by massive otoliths with a much derived sulcus type. The huge and rounded ostium almost reaches the anterior rim of the otolith, and the peculiar cauda is separated into a wide, straight, and rod-like anterior part with an oval posterior part having an elevated caudal colliculum in the centre. The morphology of this caudal colliculum and its space between the cristae are crucial features for identification at the species level (L. crocea and L. polyactis): (1) the caudal colliculum is very rounded in L. polyactis, whereas it has an oblique cutting rim on the dorsal part in L. crocea; and (2) the spaces between the caudal colliculum and crista superior (as a posterior depression found by Ohe Citation2011) as well as the crista inferior (as anterior depression found by Ohe Citation2011) are much wider in L. crocea (Schwarzhans Citation1993, figs 368–371; Ohe Citation2011, fig. 11a–d; , b, f). Schwarzhans (Citation1993) suggested that the presence of a protruding posterior tip on the otoliths of L. crocea can be used to distinguish the two species. However, our extensive reference material reveals that this feature is not consistent among all the examined L. crocea otoliths because some individuals appear to be more blunt, as seen in L. polyactis individuals (), although a posterior tip has never been found in L. polyactis. Although the otoliths of Larimichthys are among the most abundant in our samples and are readily distinguished at the genus level, the preservation status does not allow confident identification in many cases; these are assigned Larimichthys sp. At least three taxa are identified – L. crocea, L. koae sp. nov., and L. polyactis – and these are among the earliest records of Larimichthys.
Larimichthys crocea (Richardson, 1846) (–e)
2010b Larimichthys crocea (Richardson, 1846); Lin, pl. 18, fig. 3, non fig. 2 [Taiwan].
Remarks
These otoliths can be reasonably assigned to recent L. crocea, but they are more or less eroded on the surface of the sulcus. Further remarks are provided under the genus.
Larimichthys koae sp. nov. (–e)
2010b Larimichthys crocea (Richardson, 1846); Lin, pl. 18, fig. 2 [Taiwan].
Derivation of name
This species is named after Mrs. Meng-Chen Ko, who accompanied the author (CHL) to the field for the first time in 2010 and from whom the author (CHL) has received continuous support in life, work, and in many other ways.
Type material
Holotype: a left otolith, ASIZF 01000057 (–c); six paratypes, of which two have been figured, ASIZF 01000058–59 (, e). Type locality and horizon: Tapu Formation at site SL-0. Dimensions of the holotype: Length = 17.29 mm, height = 6.70 mm, thickness = 5.98 mm.
Diagnosis
OL/OH = 2.64; OL/OT = 2.88. Extremely elongate otoliths up to 20 mm length. Inner face convex; outer face with large postcentral umbo. Dorsal rim flat, shallow, anteriorly high. Ventral rim concave in the middle. Ostium expands upwards anteriorly. Cauda extremely long with rounded tip.
Description
The otoliths of this new species are extremely elongated. The deepest part is in their anterior half. The posterior part gradually becomes narrow, with a tapering tip. Almost no dorsal area is present above the crista superior. A larger ventral area is restricted to the space between the posterior rim of the ostium and anterior rim of the downturned cauda. A notable constriction on the ventral rim immediately below the downturned part of the cauda is observed. The margins are smooth. The inner face is convex. The outer face is very thick, with a massive swollen postcentral located umbo, and this contributes to much of the otolith thickness. The sulcus is of a typical Larimichthys type (see under genus). The ostium is huge relative to the entire otolith, and it is directed forwards and reaches the anterior rim. The spaces between the caudal colliculum and cristae appear to be more similar to those in L. crocea than those in other species, though all specimens have some abrasion on the surface of the cauda.
Discussion
The otoliths of L. koae sp. nov. are differentiated from other congeners by their extremely slender shape, large ostium, and nearly absent dorsal area.
Occurrence
Currently, only known from the late Miocene lower Tapu Formation at Shulin, Taiwan.
Larimichthys polyactis (Bleeker, 1877) (–i)
Remarks
See remarks under the genus.
Genus Miichthys Lin, 1938
Miichthys miiuy (Basilewsky, 1855) (–j)
?1974 (Sciaenidarum) kiyoharai n. sp.; Ohe, fig. 1 [Japan].
?1976 Paleosciaena kiyoharai (Ohe); Ohe, p. 82 [Japan].
2018b Miichthys miiuy; Lin et al., pl. 3, figs. 6, 7 [Taiwan].
Remarks
Three otoliths are identified as belonging to this species. They closely resemble those of the recent M. miiuy because they have an elongated shape with a blunt anterior rim (Schwarzhans Citation1993, figs 239, 240; Ohe Citation2011, fig. 12). The posterior rim varies; it may be largely blunt or pointed with a notch on the upper part. A massive postcentral umbo exists on the outer face. The ostium is huge, occupying approximately more than half of the otolith length, and it runs downwards towards the anterior rim. The space between the posterior rim of the ostium and anterior rim of the downturned cauda is large. A single otolith described by Ohe (as (Sciaenidarum) kiyoharai in 1974 and as Paleosciaena kiyoharai in 1976) from the early Miocene Yakuoji Formation and several further otoliths from the Mizunami Group (as Paleosciaena mizunamiensis in Ohe Citation1976) are similar to the otoliths of M. miiuy, but the Japanese specimens appear to have a shorter ostium and a much narrower distance between the posterior rim of the ostium and the downturned part of the cauda. The so-called Paleosciaena with two true fossil species may be a genus closely related to Miichthys. However, Schwarzhans (Citation1993, p. 12) listed P. mizunamiensis as a doubtful species and was followed as such in Nolf (Citation2013, p. 190). We conclude that a detailed taxonomic work on this particular taxon is required.
Genus Nibea Jordan & Thompson, 1911
Remarks
Otoliths of Nibea form a well-defined morphological group among other sciaenids. They are bullet-shaped with a vertical, straight posterior rim and a somewhat blunt anterior rim. The antero-dorsal rim is often oblique and oriented downwards anteriorly, and a large angle of approximately 90° exists on the postero-dorsal rim. The inner face is convex and considerably so in some species, whereas a massive postcentral located umbo exists on the outer face. The cauda is horizontal and narrow in the anterior part and widens gradually and then, turns strongly downwards with an angle of 90°, with a tip pointed anteriorly. Two species, N. albiflora and N. chaoi sp. nov., have been identified in our material.
Nibea albiflora (Richardson, 1846) (–l)
1976 Nibea albiflora Richardson; Ohe, pl. 24, figs. 1, 3 [Japan].
1981 Nibea albiflora Richardson; Ohe, pl. 6, fig. 2 [Japan].
2018b Nibea albiflora; Lin et al., pl. 3, fig. 8 [Taiwan].
2019a Nibea albiflora; Lin et al., fig. 5n–p [Taiwan].
Remarks
See remarks under the genus and further discussion under N. chaoi sp. nov.
Nibea chaoi sp. nov. (–i)
?1976 Nibea diacanthus Richardson; Ohe, pl. 24, figs. 4–10 [Japan].
?1977 Nibea diacanthus Lacepede; Ohe, fig. 10 [Japan].
?1981 Nibea diacanthus Lacépède; Ohe, pl. 6, fig. 1 [Japan].
2010b Nibea semifascata Chu, Lo & Wu, 1963; Lin, pl. 17, fig. 1, 2 [Taiwan].
2010b Nibea sp.; Lin, pl. 17, fig. 6, second specimen from the left, non others [Taiwan].
2019a Protonibea diacanthus; Lin et al., fig. 5e [Taiwan].
2019a Nibea soldado; Lin et al., fig. 5f, g [Taiwan].
Derivation of name
This species is named after Prof. Ning Labbish Chao (NMMBA), who has contributed so much globally to the systematics of Sciaenidae and built up the associated GSCN collection at the NMMBA and who is also a mentor to the author (CHL).
Type material
Holotype: a right otolith, ASIZF 01000067 (–l); 57 paratypes, of which four have been figured, ASIZF 01000063–66 (–f). Type locality and horizon: Tapu Formation at site SL-3a. Dimensions of the holotype: Length = 7.26 mm, height = 5.46 mm, thickness = 2.83 mm.
Diagnosis
OL/OH = 1.30–1.45; OL/OT = 3.08–3.30. Oval to square otoliths up to 7.2 mm length. Inner face convex; outer face slightly concave. Dorsal rim flat, shallow, expanded anteriorly. Ventral rim gently curved. Ostium triangular, wide post-ventrally, narrow anteriorly. Cauda long, anterior half horizontal and posterior half strongly downwards bent.
Description
The otoliths are oval to square with a stronger angle on the postero-dorsal part. The antero-dorsal part is extended antero-dorsally and forms a triangular dorsal area above the ostium. This feature can be observed in all size ranges. The ventral rim is gently curved. The margins are smooth. The inner face is convex, and the outer face is flat or slightly concave, without an umbo. The thickest part is in the posterior portion of the otolith. The sulcus is of a typical Nibea type (see remarks under genus). The ostium is large, triangular in shape, and completely filled by a colliculum. The postero-ventral part of the ostium is enlarged backwards, and slight constrictions are present in the middle of both cristae. The cauda is narrow and horizontally straight in the anterior half, widening posteriorly after turning downwards at 90°. The very posterior tip of the cauda is pointed forwards.
Discussion
On the basis of the peculiar enlargement of the antero-dorsal area, the otoliths of this new species have been assigned to Protonibea diacanthus in many previous studies (see the synonym list). Although the enlargement of the antero-dorsal area in N. chaoi sp. nov. is very similar to that of P. diacanthus, the cauda is of a typical Nibea type (see remarks under genus). Our extensive recent otolith collections (i.e. GSCN, NMMBA, and CHLOL) reveal that regardless of its size, the width of the cauda in P. diacanthus remains constant and is not as wide as that of Nibea before its posterior end, and it turns downwards more gently (without a 90° angle). These two features are specific to Protonibea otoliths (, n). The otoliths of N. chaoi sp. nov. also share many similarities with those of N. albiflora and N. soldado, but the antero-dorsal rim is sharply oblique in N. albiflora, whereas the shape is more compact with a much shorter ostium in N. soldado (Schwarzhans Citation1993, fig. 309). The otoliths of N. albiflora are also characterised by an anteriorly stronger convex inner face and a much thicker profile than the new species.
Occurrence
Taiwan: late Miocene lower Tapu Formation at Shulin and not localised Plio-Pleistocene riverbed of the Tsailiao River; ?Japan: Pliocene Kakegawa Group.
Genus Protonibea Trewavas, 1971
Protonibea aff. diacanthus (Lacepède, 1802) (, p)
Remarks
Two otoliths share many similar features with P. diacanthus otoliths, as they have a largely triangular shape with a deep ventral area, a large ostium, and a narrow cauda with a gentle downturned angle and that only widens at its posterior tip. On the outer face of smaller recent specimens, a tiny pointed angle is often present in the middle of the dorsal margin (Schwarzhans Citation1993, figs 293, 294; Ohe Citation2011, fig. 18), whereas the dorsal margin varies from smooth to undulated in large otoliths (Ohe Citation2011, fig. 19; Lin and Chang Citation2012, pl. 41, erroneously as Argyrosomus japonicus).
Genus Taosciaena gen. nov.
Type species
Taosciaena jiangi sp. nov.
Derivation of name
The genus is named after the Taiwanese palaeontologist Prof. Hsi-Jen Tao in combination with the genus name Sciaena. The name is in honour of her outstanding contributions to palaeontology and comparative anatomy studies in Taiwan and her role as a mentor to the author (CHL). The gender is feminine.
Diagnosis
The otoliths of Taosciaena are oval to elliptic in shape. The highest part of the otolith is in the antero-dorsal part. The margins are smooth. The otoliths are massive with moderate thickness (OL/OT = 2.27–3.26). The inner face is strongly convex, and the outer face is moderately convex, with or without a weak postcentral umbo. The sulcus is of a typical sciaenid type, with a wide and rounded ostium and a long but strongly bent cauda. The horizontal part of the cauda is as long as the downturned posterior part. The cauda ends very slightly below the ostial crista inferior, and the downturned part of the cauda is slightly widened. The space between the posterior rim of the ostium and downturned part of the cauda is wide.
Discussion
The otoliths of this new genus have a distinct combination of features that are similar in part to those of several extant genera, such as Collichthys, Larimichthys, and Sonorolux (see Schwarzhans Citation1993, figs 368–377). The high but very short ostium, markedly downturned caudal tip, and flat dorsal rim are most similar to that of Larimichthys (a west Pacific genus) than any other known genera (, b, f). In particular, the morphology of the otoliths of T. hui sp. nov. (see below) likely represents transitional morphology between that of Larimichthys and that of Taosciaena, suggesting a close relationship between the two genera. Because the otoliths of extant genera of Sciaenidae are adequately known (Schwarzhans Citation1993; Nolf Citation2013), and given the low possibility of finding fish skeletal fossils in the study region (Lin et al. Citation2019a), we therefore establish an otolith-based fossil genus Taosciaena.
Occurrence
Currently only known from the late Miocene lower Tapu Formation at Shulin, Taiwan.
Composition
Taosciaena jiangi and T. hui.
Taosciaena hui sp. nov. (–l)
Derivation of name
It is named in honour of the late Prof. Chung-Hung Hu in recognition of his fundamental contributions to palaeontology in Taiwan.
Type material
Holotype: a right otolith, ASIZF 01000072 (–e); 14 paratypes, of which six have been figured, ASIZF 01000071, 73–77 (, b, f–l). Type locality and horizon: Tapu Formation at site SL-3a. Dimensions of the holotype: Length = 6.37 mm, height = 4.49 mm, thickness = 2.34 mm.
Diagnosis
OL/OH = 1.31–1.48; OL/OT = 2.51–3.01. Oblong otoliths up to 6.4 mm length. Inner face convex; outer face convex with large postcentral umbo. Dorsal rim oblique, elevated anteriorly. Ventral rim gently curved. Ostium rounded, wide and short. Cauda long, horizontal anteriorly and strongly downwards bent posteriorly.
Description
The otoliths are oblong. Both the anterior and posterior rims are blunt, but the anterior one is much larger. The dorsal rim shows a mild slope that is highest anteriorly, and all other rims curve gently. The margins are smooth. Both the inner and outer faces are convex, this is most evident in the outer face which is very thickset. An umbo is present on the posterior part of the outer face. A general description of the sulcus is provided under the genus. The ostium is huge, very high relative to the otolith height, and directed downwards, reaching the anterior rim. The horizontal part of the cauda is longer than the curved posterior part, and these two parts turn through an angle of approximately 90°. The anterior part (crista inferior) of the downturned part of the cauda is slightly grooved. The space between the posterior rim of the ostium and downturned part of the cauda is wide.
Discussion
This new species is uncommon in the collection. See below for a discussion of the comparison of the more abundant congeners, T. jiangi sp. nov.
Occurrence
Currently, only known from the late Miocene lower Tapu Formation at Shulin, Taiwan.
Taosciaena jiangi sp. nov. (–p)
2010b Nibea albiflora (Richardson, 1846); Lin: pl. 17, fig. 3 [Taiwan].
2010b Nibea sp.; Lin: pl. 17, figs. 6 (non second specimen from the left), 7 [Taiwan].
Derivation of name
This species is named after Mr. Chao-Ping Jiang (Taipei, Taiwan), a friend and enthusiastic fossil lover, who was among the first to discover the localities described in this study and who kindly guided us there.
Type material
Holotype: a right otolith, ASIZF 01000080 (, d); 421 paratypes, of which nine have been figured, ASIZF 01000078–79, 81–87 (, b, e–p). Type locality and horizon: Tapu Formation at site SL-0. Dimensions of the holotype: Length = 5.11 mm, height = 3.71 mm, thickness = 2.14 mm.
Diagnosis
OL/OH = 1.16–1.41; OL/OT = 2.27–3.26. Oblong to rounded otoliths up to 5.5 mm length. Inner face convex; outer face convex with slightly raised postcentral umbo. Dorsal flat, slightly elevated anteriorly. Ventral rim gently curved. Ostium rounded, wide and short. Cauda long, anterior half horizontal; posterior half strongly downwards bent and its crista inferior significantly grooved.
Description
The otoliths of the species are massive and robust, with a thickset ventral portion. Ontogenetic variation of the otolith shape is evident; it is elliptic in the larger (adult) specimens (OH/OL = 70%–75%), but more rounded in the smaller ones (OH/OL = 85%). The margins are smooth. In large specimens, the dorsal rim has a mild slope and is highest anteriorly; other rims are rounded and gently curved. Both the inner and outer faces are convex, but this is most expressed in the inner face. The thickest part of the inner face is located in the middle of the otolith, whereas in the outer face, it is slightly shifted posteriorly, often forming a weak postcentral umbo. The outer face convexity is most evident in the adults; it is rather flat in smaller specimens. The ostium is very high, and its anterior rim occupies almost the entire anterior rim of the otolith. The connection between the ostium and the cauda is marked by a strong constriction on the cristae, but the crista inferior is slightly behind the superior one, causing a somewhat ventrally oriented appearance in the ostium shape. The horizontal part of the cauda is as long as the curved posterior part, and these two parts turn with an angle slightly less than 90°. The anterior part (crista inferior) of the downturned part of the cauda is significantly deep and grooved, providing a cliff-like structure to the crista inferior. This feature is also observed in the juvenile specimens. The space between the posterior rim of the ostium and the downturned part of the cauda is wide.
Discussion
The smaller specimens of T. jiangi sp. nov. are usually poorly preserved, and therefore, our description is mainly based on adult specimens. The otoliths of T. jiangi have much in common with the congeneric species T. hui sp. nov. (see above), but T. jiangi differs from the latter species in the following features: (1) the outline is less elongated, and the antero-dorsal part is flatter; (2) the distance between the posterior rim of the ostium and downturned part of the cauda is smaller; (3) the downturned part of the cauda is longer and anteriorly curved at its tip with an angle of less than 90°; and (4) the anterior rim of the downturned cauda is deeper and groove-like.
Occurrence
Currently, only known from the late Miocene lower Tapu Formation at Shulin, Taiwan.
Order Spariformes Bleeker, 1876
Family Sillaginidae Richardson, 1846
Genus Sillago Cuvier, 1816
Sillago sp. (, p)
Remarks
One strongly eroded specimen is available. The sulcus is long and horizontal across the entire upper part of the otolith, a feature that has been adequately recognised in Sillago otoliths.
Results
Abundance and preservation
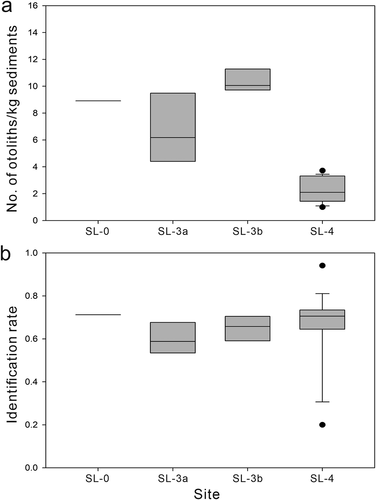
A total of 1716 otolith specimens were recovered from both bulk (n = 1551) and surface (n = 165) samples (). The abundance of otoliths was not evenly distributed in the sediments, with bulk samples from site SL-4 yielding the lowest average density of otoliths (c. 2 kg−1 sediment) and SL-3b yielding the highest (10.41 kg−1; , ). The average density of otoliths across all bulk samples was 4.3 kg−1. The preservation of the otoliths in bulk samples was generally poor, with high numbers of unidentifiable otoliths (546 of 1551 otoliths). Most specimens were fragile in nature, and many were found to be broken into fragments after preparation; they had severe abrasion, leaching, and weathering. In most of the bulk samples, the identification rate to any specific taxonomic level was approximately 60%, although it varied considerably in site SL-4 (). Only seven unidentified otoliths (of 165) were encountered in the surface samples.
Figure 15. Differences in average density of otoliths (a) and identification rate (b) for the sampled sites. Bulk samples from the same site are grouped, and samples containing fewer than 10 otoliths are excluded. Box plots with lower (25th percentile), median, and upper (75th percentile) boundaries, whiskers of 10th and 90th percentiles, and outliers (solid circle) outside of 10th and 90th percentiles are presented.
Diversity and composition
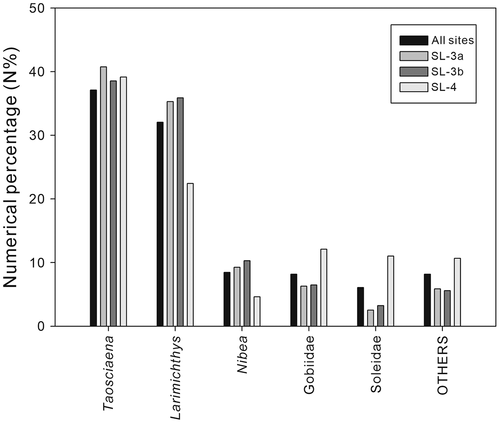
The diversities in surface and bulk samples were different (). In the surface samples, three families (Ariidae, Uranoscopidae, and Sciaenidae) and 15 taxa were recognised, whereas all families and most taxa (except Netuma sp., Argyrosomus sp., and P. aff. diacanthus) of the entire collection could be found in the bulk samples. Rarefaction and extrapolation curves for bulk samples indicated that our sampling effort nearly reached the maximum coverage, even for the less sampled sites SL-3a and SL-3b (). Accounting for more than 65% of the total otolith count, the sciaenids dominate across all assemblages by their taxonomic composition and abundance. Five main taxa belonging to three families, Scieanidae, Gobiidae, and Soleidae, comprised more than 90% in all assemblages (). Samples from sites SL-3a and SL-3b exhibited similar patterns, whereas otoliths of Gobiidae and Soleidae were more abundant than those of Nibea in site SL-4 (). Moreover, a higher proportion of otoliths belonging to other taxa was observed in SL-4 ().
Figure 16. Sample-based rarefaction and extrapolation curves with 95% confidence intervals. Reference samples are indicated by solid circles, rarefaction by solid lines, and extrapolation by dashed lines.
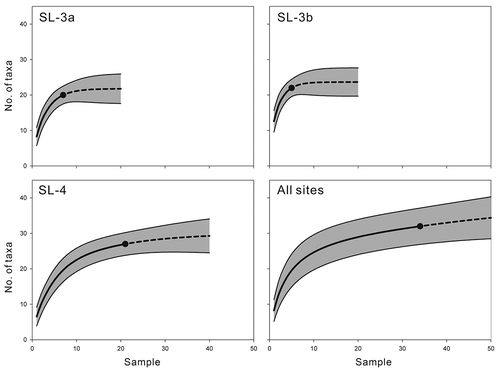
Figure 17. Rank abundance of otolith taxa in sites at Shulin.
Discussion
Taphonomic considerations and sample coverage
Otolith density has not been routinely measured in previous studies. Although several approximate values have usually been obtained from a single large bulk sample per site, these may be difficult to evaluate statistically without replicates (e.g. Lin et al. Citation2017a). The otolith counts in our samples were within the range of averages compared with typical clastic sedimentary environments, although the density was markedly low in the SL-4 samples (, ). For instance, Stringer et al. (Citation2016, Citation2020) reported a density of approximately 5 kg−1 in late Cretaceous assemblages from North America. In some cases, the density can be extremely high, representing a high otolith concentration (e.g. 44.52 kg−1 in Schwarzhans and Stringer Citation2020). Moreover, Lin et al. (Citation2019b) and Heard et al. (Citation2020) reported higher densities of 44.5 kg−1 and 20 kg−1 in coral and algal reefs, respectively. It seems likely that otoliths are not distributed evenly, even across a restricted area, as in our sample sites and that the sedimentation rate is a significant controlling factor (, SL-3a/3b vs SL-4). Consequently, sufficient sampling coverage would be required.
The majority of our otoliths were eroded; particularly, on the surface and margins, they were abraded or consolidated with tiny sediment grains. This was because of the high quartz content and poorly sorted grains in the sediments (Appendix 2) as well as subaerial weathering and acidic leaching. This preservation status impeded taxonomic work considerably, with a high proportion of otoliths being unidentifiable ().
Despite the varied otolith density and poor preservation, our sampling can be considered sufficient, as demonstrated by the rarefaction curves (). At least 34 taxa were represented in our overall 34 bulk samples, and the extrapolation curves revealed that the Shulin assemblage would contain c. 35 taxa, with an additional 16 samples (up to a total of 50 bulk samples of approximately 500 kg of sediment; ). The difference in the taxonomic diversity between the bulk and surface samples should be noted, however. While the bulk samples likely exhibited natural community structures (cf. Schwarzhans Citation2013; Lin et al. Citation2016, Citation2017b, Citation2018a), the surface samples contained marginally less than half of the entire composition and, inevitably, of taxa with large otoliths, such as the Sciaenidae and Ariidae (), were overestimated. This is natural because they are the largest and easiest to be found by surface sampling.
Ecological representation and inferred palaeoenvironment
The study of 1716 otoliths from the Tapu Formation in the Shulin area revealed the presence of at least 34 teleost taxa, among which four are newly described species (). The assemblage is not particularly diverse but is numerically dominated by three families: Sciaenidae, Gobiidae, and Soleidae, which comprise more than 90% of the total otolith count in all samples and account for more than half of the taxa identified (20 of 34; ). These are typical coastal elements demersally inhibiting shallow water environments with flat bottoms. Neritic epipelagic taxa including Engraulidae and Clupeidae were found but in a very marginal proportion, and deeper water taxa were totally absent. Furthermore, the occurrence of some Ariidae otoliths indicates the possible presence of a rather large river system or an environment close to a river mouth.
Although only at the family level, the high numbers of Sciaenidae, Gobiidae, and Soleidae were roughly similar to the fish composition found in modern western Taiwan (Kuo and Shao Citation1999; Shao et al. Citation2002; Shao Citation2021), suggesting closely related environments. On the basis of a few otoliths from the same area, Lin (Citation2010b) first interpreted the palaeoenvironment as being reflective of a shallow coastal sea. The sedimentary environment of the Tapu Formation is generally littoral to the offshore transition zone (Pan et al. Citation2015; Chen and Yu Citation2016). These lines of evidence and the otoliths described herein, along with the associated marine fossils (see above, Other associated fossils), allow us to infer that in the late Miocene, the Shulin area was a coastal and estuary palaeoenvironment, particularly a shallow water environment with gentle topography covered by muddy or sandy bottoms.
Otolith fossil record of Sciaenidae in the west Pacific
The most remarkable result of the present collection is the dominance of sciaenid otoliths. Otolith-based fossil records for Sciaenidae have been particularly abundant worldwide and provide a more complete picture in terms of palaeodiversity and distribution relative to skeletal records (Aguilera et al. Citation2016; Bannikov et al. Citation2009, Citation2018; Přikryl et al. Citation2021). The presence of Larimichthys koae sp. nov., Taosciaena hui sp. nov., and T. jiangi sp. nov., and the earliest records of Larimichthys spp. based on otoliths reveal a high diversity in the Larimichthys-related species group (Schwarzhans Citation1993) during the late Miocene in western Taiwan. This result agrees well with the findings of the molecular study, which indicated a divergent time for Larimichthys in the late Miocene–early Pliocene (Lo et al. Citation2015). Based on fossil otolith data, it seems very likely that this specific lineage experienced species radiation precedingly unnoticed.
At generic level, however, the sciaenid composition in Shulin is distinct. The modern fish fauna of western Taiwan includes large numbers of small Johnius and Pennahia (Shao et al. Citation2002), both of which are absent in Shulin. Otoliths of Johnius are known from the Pliocene of Japan (Ohe Citation1976, Citation1981), and those of Pennahia are known from the Pleistocene (Ohe Citation1976). These fossil records are younger than the molecular results, which suggest the first occurrence early in the late Miocene and coeval to Nibea and Larimichthys (Lo et al. Citation2015). The Taiwanese records of Johnius and Pennahia only date back to late Pleistocene (Lin et al. Citation2018b, Citation2019a), but this could be a sampling artefact. We expect that more fossils from various horizons would help develop a more complete view of the evolutionary history of these taxa.
In addition to the extinct Taosciaena, it appears that Nibea and Larimichthys have been important taxa in the subtropical west Pacific fish fauna since the late Miocene. Nevertheless, Larimichthys would have an abundance only similar to that in the modern fish fauna, if this fauna was pristine and not so heavily fished, even to a point that the genus is nearly extinct in the wild today (Liu and Mitcheson Citation2008). In the 1980s, a fishery collapse occurred in stocks of Larimichthys, particularly L. crocea, and they are now mainly obtained through aquaculture. The dramatic contrast between the fossil assemblages and modern communities is not the result of natural extinction but reflects the anthropogenic impact in this particular case.
Conclusion
The present collection is among the first to describe Neogene otoliths from Taiwan in detail. Although identification is limited due to the poor preservation and differing densities of otoliths in the sediments from different sites, our sampling of more than 1700 otoliths revealed an assemblage of 34 taxa belonging to 13 families. The dominant taxa suggest a coastal, nearshore palaeoenvironment with flat shallow muddy or sandy bottoms and perhaps a location near to a river mouth. The general composition is similar to the present-day fish fauna in the west of Taiwan but very different at genus and species levels. This suggests a mild turnover at certain lineages (Larimichthys-related group). We provide unique ‘snapshots’ of a coastal fish community in the rarely explored late Miocene subtropical northwest Pacific and, possibly exclusively with fossil otolith data, reveal the past fish diversity that otherwise cannot be recognised. Although it would be too early to evaluate Taiwanese fish fauna in time and space at this stage, our results clearly demonstrate the potential of a larger scale biogeographical study. We thus expect future works comparing coeval assemblages from the entire west Pacific, such as with the better understood Kakegawa Group fauna from Japan (Ohe Citation1974, Citation1977, Citation1981, Citation1983).
Supplemental_Fig.2_.tif
Download TIFF Image (15.1 MB)Suppl._Fig._1.tif
Download TIFF Image (38 MB)Acknowledgments
It is our great pleasure to thank all the people who have helped in the field: Chao-Ping Jiang, Ching-Tung Hsiung, Ching-Yuan Lee, Chia-Fong Chen, Shih-Chuan Chen, Chih-Kai Chuang, Shih-Wei Lee (National Museum of Marine Science and Technology), Meng-Chen Ko, Li-You Lin, Li-Li Lin, Li-Man Lin, Junko Kurosaki, and the members of Marine Paleontology Lab, BRCAS (https://otolithlin.biodiv.tw/). We are grateful to Hsi-Jen Tao (National Taiwan University) for providing fossil specimens, Chih-Wei Chang (Marine Ecology and Conservation Research Center, National Academy of Marine Research, Taiwan) for providing access to the recent otolith collection, Mitsui Shota (Tokyo University of Marine Science and Technology) for providing necessary Japanese literature, and Yen-Chun Wang for preparing the measured section of the sites (). This manuscript has been improved by the revisions of Angela Girone, Werner Schwarzhans, Dirk Nolf, and an anonymous reviewer.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Suppplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
Related Research Data
References
- Aguilera OA, Moraes-Santos H, Costa S, Ohe F, Jaramillo C, Nogueira A. 2013. Ariid sea catfishes from the coeval pirabas (Northeastern Brazil), cantaure, castillo (Northwestern Venezuela), and castilletes (North Colombia) formations (early miocene), with description of three new species. Swiss J Palaeontol. 132:45–68. doi:https://doi.org/10.1007/s13358-013-0052-4.
- Aguilera OA, Schwarzhans W, Béarez P. 2016. Otoliths of the sciaenidae from the neogene of tropical America. Palaeo Ichthyo. 14:7–90.
- Bannikov AF, Carnevale G, Landini W. 2009. A new early miocene genus of the family sciaenidae (teleostei, perciformes) from the eastern Paratethys. C R Palevol. 8(6):535–544. doi:https://doi.org/10.1016/j.crpv.2009.03.001.
- Bannikov AF, Schwarzhans W, Carnevale G. 2018. Neogene paratethyan croakers (teleostei, sciaenidae). Riv Ital Paleontol S. 124(3):535–571.
- Chen W-S, Yu N-T. 2016. Geology of the Western Foothills. In: Chen W-S, editor. An introduction to the geology of Taiwan. Taipei. Chinese: Geological Society of Taiwan; p. 53–90.
- Colwell RK 2013. EstimateS: statistical estimation of species richness and shared species from samples. version 7.5. [ accessed 2021 Feb 2]. http://purl.oclc.org/estimates
- Fricke R. 2018. Two new species of stargazers of the genus. Uranoscopus. 4476(1):157–167. (Teleostei: Uranoscopidae) from the western Pacific Ocean. Zootaxa.
- Fricke R, Eschmeyer WN, Van Der Laan R 2021. Eschmeyer’s catalog of fishes: genera, species, references. [ accessed 2021 Feb 28]. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
- Heard J, Tung W-C, Pei Y-D, Lin T-H, Lin C-H, Akamatsu T, Wen Colin K-C. 2020. Coastal development threatens Datan area supporting greatest fish diversity at taoyuan algal reef, northwestern Taiwan. Aquat Conserv Mar Freshw Ecosyst. published online 2020 Nov 16. doi:https://doi.org/10.1002/aqc.3477.
- Iizuka K, Katayama S. 2008. Otolith morphology of teleost fishes of Japan. Bull Fish Res Agency. 25:1–222.
- Kuo S-R, Shao K-T. 1999. Species composition of fish in the Coastal Zones of the Tsengwen Estuary, with descriptions of five new records from Taiwan. Zool Stud. 38(4):391–404.
- Leprieur F, Descombes P, Gaboriau T, Cowman PF, Parravicini V, Kulbicki M, Melian CJ, De Santana CN, Heine C, Mouillot D, et al. 2016. Plate tectonics drive tropical reef biodiversity dynamics. Nat Commun. 7(1):11461. doi:https://doi.org/10.1038/ncomms11461.
- Lin C-H. 2010a. Study of uranoscopidae otolith morphology from Da-si, Yi-lan County. J National Taiwan Mus. 63(2):17–29. Chinese.
- Lin C-H 2010b. Morphology of otolith of living and fossil fishes from Taiwan. [master’s thesis]. Taipei (TW): National Taiwan University. Chinese.
- Lin C-H, Chang C-W. 2012. Otolith atlas of Taiwan fishes. Pingtung: National Museum of Marine Biology and Aquarium.
- Lin C-H, Chiang Y-P, Tuset VM, Lombarte A, Girone A. 2018a. Late quaternary to recent diversity of fish otoliths from the Red Sea, central Mediterranean, and NE Atlantic sea bottoms. Geobios. 51(4):335–358. doi:https://doi.org/10.1016/j.geobios.2018.06.002.
- Lin C-H, Chien C-W, Lee S-W, Chang C-W. 2019a. Fossil fishes of Taiwan: a review and prospection. Hist Biol. 1–11. published online 2019 Dec 10. doi:https://doi.org/10.1080/08912963.2019.1698563.
- Lin C-H, De Gracia B, Pierotti MER, Andrews AH, Griswold K, O’Dea A. 2019b. Reconstructing reef fish communities using fish otoliths in coral reef sediments. PLoS ONE. 14(6):e0218413. doi:https://doi.org/10.1371/journal.pone.0218413.
- Lin C-H, Girone A, Nolf D. 2016. Fish otolith assemblages from recent NE Atlantic sea bottoms: a comparative study of palaeoecology. Palaeogeogr Palaeoclimatol Palaeoecol. 446:98–107. doi:https://doi.org/10.1016/j.palaeo.2016.01.022.
- Lin C-H, Li K-T, Chang C-W. 2013. Identification of pomadasys species (pisces, haemulidae) from an archaeological midden site in Nankuanli East (Taiwan), based on otolith morphology. Raffles Bull Zool. 61(1):293–302.
- Lin C-H, Nolf D, Steurbaut E, Girone A. 2017a. Fish otoliths from the lutetian of the Aquitaine Basin (SW France), a breakthrough in the knowledge of the European Eocene ichthyofauna. J Syst Palaeontol. 15(11):879–907. doi:https://doi.org/10.1080/14772019.2016.1246112.
- Lin C-H, Taviani M, Angeletti L, Girone A, Nolf D. 2017b. Fish otoliths in superficial sediments of the Mediterranean Sea. Palaeogeogr Palaeoclimatol Palaeoecol. 471:134–143. doi:https://doi.org/10.1016/j.palaeo.2016.12.050.
- Lin C-H, Wang L-C, Wang C-H, Chang C-W. 2018b. Common early pleistocene fish otoliths from Niubu in Chia-Yi County, southwestern Taiwan. J National Taiwan Mus. 71: 47–68. Chinese.
- Liu M, Mitcheson Y. 2008. Profile of a fishery collapse: why mariculture failed to save the large yellow croaker. Fish Fish. 9(3):219–242. doi:https://doi.org/10.1111/j.1467-2979.2008.00278.x.
- Lo P-C, Liu S-H, Chao NL, Nunoo FKE, Mok H-K, Chen W-J. 2015. A multi-gene dataset reveals a tropical New World origin and early miocene diversification of croakers (perciformes: sciaenidae). Mol Phylogenet Evol. 88:132–143. doi:https://doi.org/10.1016/j.ympev.2015.03.025.
- Mitsui S, Taru H, Ohe F, Lin C-H, Strüssmann CA. 2021. Fossil fish otoliths from the chibanian miyata formation, kanagawa prefecture, Japan, with comments on the paleoenvironment. Geobios. published online 2021 Feb 4, 64:47–63. doi:https://doi.org/10.1016/j.geobios.2020.11.003.
- Miyata S, Kawano S, Ando Y, Hattori S. 2018. The fish otolith and teeth from the lower miocene akeyo formation, mizunami group at the construction site of Mizunami-Kita Junior High School in Mizunami City, Gifu, Japan. Bull Mizunami Foss Mus. 44:: 31–38. Japanese.
- Nelson JS, Grande TC, Wilson MVH. 2016. Fishes of the World. 5th ed. Hoboken (NJ): John Wiley & Sons.
- Nolf D. 1985. Otolithi piscium. In: Schultze H, editor. Handbook of paleoichthyology. Vol. 10. Stuttgart: Gustav Fischer Verlag.
- Nolf D. 2013. The diversity of fish otoliths, past and present. Brussels: the Royal Belgian Institute of Natural Sciences.
- Ohe F. 1974. Fossil sciaenoid otolith (bonyfish’s ear-stone) of the miocene yakuoji formation, Isshi Group, Mie Pref., Central Japan. Kaseki No Tomo. 12: 4–11. Japanese.
- Ohe F. 1976. Fish otoliths of sciaenid fishes from the tertiary and quaternary formations, Tokai district, central Japan and their paleontological vicissitudes. Mizunami Foss Mus Bull. 3: 73–97. Japanese.
- Ohe F. 1977. Otoliths from dainichi sand and hosoya tuffaceous beds of Kakegawa Group (Pliocene), Shizuoka Prefecture, Central Japan. Kaseki No Tomo. 16:: 13–19. Japanese.
- Ohe F. 1981. Fish-otoliths from the dainichi sand and the fosoya tuffaceous members of the Pliocene Kakegawa Group, Shizuoka Prefecture, Central Japan. Bull Senior High School, Aichi Univ. 8:: 125–194. Japanese.
- Ohe F. 1983. On the otoliths of deep water fishes from pliocene hijikata mud formation exposed in the Southern part of Kakegawa City, Shizuoka Prefecture, Central Japan. Bull Senior High School, Aichi Univ. 10:: 1–54. Japanese.
- Ohe F. 1985. Marine fish-otoliths of Japan. Kariya: The Senior High School Attached to the Aichi University of Education. Japanese.
- Ohe F. 2000. Otoliths of three species belonging to family ariidae from Eeast China Sea. Kaseki No Tomo. 47:: 35–42. Japanese.
- Ohe F. 2006. Skulls and otoliths of eleven sea catfishes (family ariidae) from Malaysia and one species related to them from the East China Sea. Nat Environ Sci Res. 19:11–28.
- Ohe F. 2011. Otolith figures of Scieanid fish inhibiting the waters around Japan, with the comments for them from the geohistorical background. Kaseki No Tomo. 56:: 32–39. Japanese.
- Ohe F, Ando Y, Ugai H, Hirose K, Kawano S. 2020. An otolith assemblage of the pleistocene ogushi formation, Itsuwa-machi, Amakusa City, Kumamoto Prefecture, Japan. Bull Goshoura Cretac Mus. 21:: 1–20. Japanese.
- Ohe F, Watanabe A. 2018. An otolith of alaska pollock theragra chalcogramma from the middle pleistocene wakimoto formation, Oga Peninsula, Akita Prefecture. Annu Akita Prefectural Mus. 43:: 13–20. Japanese.
- Pan T-Y 2011. A study on sedimentary environments of nanchuang formation to yangmei formation along the Dahan River section, northwestern Taiwan. [master’s thesis]. Taoyuan (TW): National Central University. Chinese.
- Pan T-Y, Lin AT-S, Chi W-R. 2015. Paleoenvironments of the evolving pliocene to early pleistocene foreland basin in northwestern Taiwan: an example from the Dahan River section. Island Arc. 24(3):317–341. doi:https://doi.org/10.1111/iar.12113.
- Přikryl T, Brzobohatý R, Carnevale G. 2021. Skeletal remains with otoliths in situ of the miocene croaker trewasciaena cf. kokeni (teleostei, sciaenidae) from the pannonian of the Vienna Basin. Bull Geosci. 96(1):19–28. doi:https://doi.org/10.3140/bull.geosci.1813.
- Renema W, Bellwood DR, Braga JC, Bromfield K, Hall R, Johnson KG, Lunt P, Meyer CP, Mcmonagle LB, Morley RJ, et al. 2008. Hopping hotspots: global shifts in marine biodiversity. Science. 321(5889):654–657. doi:https://doi.org/10.1126/science.1155674.
- Rivaton J, Bourret P. 1999. Les otolithes des poissons de l’Indo-Pacifique. Nouméa: Documents scientifiques et techniques, Institut de recherche pour le développement.
- Schwarzhans W. 1993. A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). München: Verlag F. Pfeil.
- Schwarzhans W. 1999. A comparative morphological treatise of recent and fossil otoliths of the order Pleuronectiformes. München: Verlag F. Pfeil.
- Schwarzhans W. 2013. Otoliths from dredges in the Gulf of Guinea and off the Azores –an actuo-paleontological case study. Palaeo Ichthyologica. 13:7–40.
- Schwarzhans W. 2019a. A comparative morphological study of recent otoliths of the congridae, muraenesocidae, nettastomatidae and colocongridae (anguilliformes). Memorie della Società Italiana di Scienze Naturali e del Museo di Storia Naturale di Milano. 46:327–354.
- Schwarzhans W. 2019b. A comparative morphological study of recent otoliths of the so-called trachinoidei. Memorie della Società Italiana di Scienze Naturali e del Museo di Storia Naturale di Milano. 46:371–388.
- Schwarzhans W, Ohe F. 2019. Lanternfish otoliths (teleostei, myctophidae) from the pliocene and pleistocene of Japan. Riv Ital Paleontol S. 125(2):355–400.
- Schwarzhans W, Ohe F, Ando Y. 2017. An early oligocene fish-fauna from Japan reconstructed from otoliths. Zitteliana. 90:3–26.
- Schwarzhans W, Stringer GL. 2020. Fish otoliths from the late maastrichtian kemp clay (Texas, USA) and the early danian clayton formation (Arkansas, USA) and an assessment of extinction and survival of teleost lineages across the K-Pg boundary based on otoliths. Riv Ital Paleontol S. 126(2):395–446.
- Shao K-T 2021. Taiwan fish database. [ accessed 2021 Feb 15]. http://fishdb.sinica.edu.tw
- Shao K-T, Hsieh L-Y, Wu -Y-Y, Wu C-Y. 2002. Taxonomic and distributional databases of fishes in Taiwan. Environ Biol Fishes. 65(2):235–240. doi:https://doi.org/10.1023/A:1020092500050.
- Stringer G, Oman L, Badger B. 2016. Woodbury formation (campanian) in New Jersey yields largest known otolith assemblage of teleostean fishes in North America. Proc Natl Acad Sci. 165: 15–36. Philadelphia.
- Stringer G, Schwarzhans W, Phillips G, Lambert R. 2020. Highly diversified late cretaceous fish assemblage revealed by otoliths (ripley formation and owl creek formation, northeast Mississippi, USA). Riv Ital Paleontol S. 126:111–155.
- Tsao S, Hong E, Song S-R, Chu H-T, Chung S-H. 1992. The absolute age of the boundary between nanchuang and kueichulin formations. Special Publication of the Central Geological Survey, MOEA. 6:223–234.
- Yu L-C, Shen S-C. 1987. Study on sciaenoid fishes from the adjacent waters around Taiwan. Annu Taiwan Mus. 30:65–134.
- Van Der Laan R, Eschmeyer WN, Fricke R. 2014. Family-group names of recent fishes. Zootaxa. 3882(1):1–230. doi:https://doi.org/10.11646/zootaxa.3882.1.1.
- Van Hinsbergh VWM, Helwerda RA. 2019. Fish otoliths from the cabarruyan piacenzian-gelasian fauna found in the Philippines. Zootaxa. 4563(3):401–443. doi:https://doi.org/10.11646/zootaxa.4563.3.1.
- Vilasri V, Ho H-C, Kawai T, Gomon MF. 2019. A new stargazer, ichthyscopus pollicaris (perciformes: uranoscopidae), from East Asia. Zootaxa. 4702(1):49–59. doi:https://doi.org/10.11646/zootaxa.4702.1.10.
Appendix
Appendix 1. Aerial photographs of the sampling sites. A–C, SL-3a/3b. D, SL-4; the layer with fossils (dashed lines) and lateral extension of SL-1 and SL-2 are indicated.
Appendix 2. Sieved sediments from SL-4 under amicroscope. A, an otolith of Taosciaena jiangi sp. nov. is visible (white circle). B, quartz grains.