
ABSTRACT
Ornithopod remains are reported from the Papo-Seco Formation (lower Barremian, Early Cretaceous) at Cape Espichel, south of Lisbon, in western central Portugal. The fossil material comes from the Boca do Chapim, Praia do Areia do Mastro and Praia do Guincho palaeontological sites. Some fossils are known from the 19th century and the beginning of the 20th century and other remains have been recently collected. Ornithopod and other vertebrate remains from the Papo-Seco Formation occur in limestones, marls, sandstones and conglomerates in a succession interpreted as being deposited in lagoonal and estuarine environments. A study of the available palaeontological material, including teeth and postcranial remains, suggests the presence of a large styracosternan-like iguanodontian, but does not allow confirmation of the occurrence of Iguanodon or Mantellisaurus. Ornithopod remains from the Papo-Seco Formation are provisionally assigned to Ornithopoda indet., Iguanodontia indet. and cf. Styracosterna indet., increasing the record of Portuguese ornithopods of early Barremian age.
Introduction
Vertebrate fossils from the Lower Cretaceous beds at Cape Espichel, south of Lisbon (Portugal), have been known since the 19th century. Dinosaur and crocodile teeth were first reported from the Papo-Seco Formation, at the Boca do Chapim site (Sauvage Citation1897-1998-1898). Subsequently, turtle remains and both crocodile and dinosaur teeth and bones have also been described from this site. Among dinosaurs, two fragmentary theropod teeth, initially referred to Megalosaurus superbus (Sauvage Citation1897-1898; Lapparent and Zbyszewski Citation1957), were later classified as Carnosauria incertae sedis (Galton Citation1994). Ruiz-Omeñaca and Canudo (2003) considered these teeth as belonging to Theropoda indet. Jaw fragments bearing teeth found at Boca do Chapim were originally regarded as crocodilian and identified as Suchosaurus girardi (Sauvage Citation1897-1898) and later referred to the spinosaurid theropod Baryonyx (Buffetaut Citation2007). More recently, Mateus et al. (Citation2011) have described cranial and postcranial remains from Praia das Aguncheiras (= Praia do Guincho; see Figueiredo et al. Citation2015) and attributed them to Baryonyx walkeri. These materials are further studied and have been described in a new species Iberospinus natarioi (Mateus and Estraviz-López, Citation2022) At Praia do Areia do Mastro, in the Boca do Chapim Formation (the formation above the Papo-Seco Formation, see ), microvertebrate remains (pycnodontid and crocodilian teeth, bone and eggshell fragments, and fish otoliths and scales) were reported by Rodrigues et al. (Citation2008).
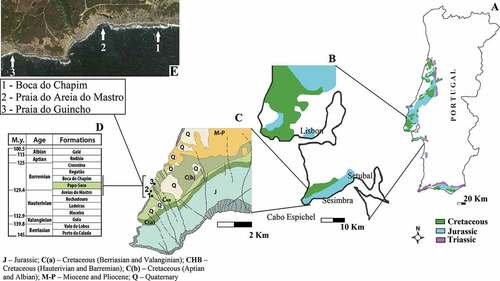
Figure 1. Regional geological map of the Cabo Espichel area with relevant location of the studied site and the limits between deposits of the Mesozoic cropping out in the area: A - Portuguese Mesozoic formations; B - Mesozoic formations of Lisbon and Setúbal peninsulas; C - Geology of Cabo Espichel area; D – Chronostratigraphic sequence of the Lower Cretaceous geological formations of Espichel, the Papo-Seco Formation is highlighted in green; E - Localization of the Paleontological sites Boca do Chapim, Praia do Areia do Mastro and Praia do Guincho in an image of Google Earth. Scales: A - 20 km; B - 10 km; C - 2 km.
Herbivorous dinosaur teeth from Boca do Chapim were identified by Sauvage Citation1897-1998-1898) as belonging to the ornithopod Iguanodon mantelli and to the sauropod Pleurocoelus valdensis. The sauropod teeth have subsequently been referred to Astrodon valdensis (Lapparent and Zbyszewski Citation1957; Galton 1981) or to Sauropoda indet. (Antunes and Mateus 2003). The validity of ‘Pleurocoelus valdensis’ is currently controversial (see Ruiz-Omeñaca and Canudo Citation2005; Upchurch et al. Citation2011); it is provisionally identified as a basal titanosauriform (Upchurch et al. Citation2011). Ornithopod remains from the Boca do Chapim site, including three teeth, three vertebrae and a fragment of femur, were attributed to Iguanodon mantelli by Lapparent and Zbyszewski (Citation1957) and later assigned to Iguanodon cf. atherfieldensis by Crespo (Citation2001) and to Iguanodon sp. by Antunes and Mateus (2003). These authors also mentioned one tooth and one deformed vertebral centrum kept in the collection of M.T. Antunes, which they attributed to Iguanodon sp. (Mateus and Antunes, 2003). This material was referred without any description, so we cannot confirm this taxonomic attribution.
In recent years, fieldwork carried out by a palaeontological team from the Centro Português de Geo-História e Pré-História (CPGP) has provided new fossils from the Papo-Seco Formation. At the Boca do Chapim site, a partial ornithopod skeleton, including postcranial (vertebral and appendicular) remains has been found (Figueiredo Citation2010, Citation2014). At the Praia do Areia do Mastro site, a maxillary fragment with teeth and isolated teeth of Iguanodon-like iguanodontians have been reported, together with other vertebrate remains belonging to a diverse fauna of actinopterygians, chelonians, crocodyliforms, pterosaurs, and theropod and sauropod dinosaurs (Figueiredo et al. Citation2015). At the Praia do Guincho site, a prezygapophysis of a vertebra was found that is described here for the first time. In addition, a natural cast of an ornithopod footprint has been discovered in this site (Figueiredo et al. Citation2017, Citation2021).
The purpose of this paper is to evaluate the ornithopod remains from the Papo-Seco Formation at Cabo Espichel (Portugal), including a review of fossils previously published from Boca do Chapim and the description of new material from three sites: Boca do Chapim, Praia do Areia do Mastro and Praia do Guincho. These data increase the current knowledge of Portuguese Early Cretaceous ornithopods.
Geological Setting
The Papo-Seco Formation outcrops between Boca do Chapim (in the south) and Praia do Guincho (in the north; ). They belong to the Sesimbra municipality and are located ~40 km south of Lisbon, in the SW side of the Setúbal Peninsula (coordinates: 38°26ʹ14.3”N, 9°12ʹ34.4”W) ().
The formal name of the Papo-Seco Formation was proposed by Rey (Citation1992). Names for this formation in older geological literature are “Grès marneux à grands sauriens” (Choffat Citation1904) and “Grès à Dinosauriens” (Rey Citation1972). Lapparent and Zbyszewski (Citation1957), and previously Sauvage Citation1897-1998-1898) and Choffat (Citation1904), regarded this unit and the fauna it contains as Aptian-Albian in age. The Papo-Seco Formation is currently attributed to the lower Barremian (Rey Citation1992; Manuppella et al. Citation1999). It is stratigraphically located between the Areia do Mastro Formation and the Boca do Chapim Formation (). Three palaeontological sites are known in the studied area: Boca do Chapim, Praia do Areia do Mastro and Praia do Guincho, which are located about 2 km north of Cabo Espichel, on the coastal cliffs of the Cabo Espichel anticline. The Papo-Seco Formation is 18.5 m-thick and composed of marls and green silty clays with traces of wood and gypsum, interbedded with sandstones; it displays a horizontal bedding (Manupella et al. Citation1999). Fossils of a diverse fauna from the Papo-Seco Formation mainly consist of vertebrate remains, found in the coarse sandstone layers, and invertebrates (bivalves, gastropods and ostracods) from the marls. The upper portion of the formation is composed of bioturbated calcareous silty layers with ostreid remains (Manupella et al. Citation1999). The sedimentological and palaeontological features generally point to a progression of an open estuarine environment to a lagoonal setting of low energy (Manupella et al. Citation1999; Rey et al. Citation2003; Figueiredo et al. Citation2015; Dinis et al. Citation2016, Citation2016, Citation2017, Citation2020a).
Figure 2. Lithostratigraphy of the Praia do Areia do Mastro site. A - Stratigraphic column and legend: 1 – sandstone; 2 – marl; 3 – limestone; 4 – lamination; 5 and 6 – vertebrates fossils (5 – footprints; 6 – bones and teeth); 7 – bivalves; 8 – gastropods. Codes: U – lithostratigraphic unit; T – thickness; B – bed reference; L – Lithology; F – fines; S – sand; G – gravel; AM – Areia do Mastro Formation; PS – Papo-Seco Formation. (Adapted from Figueiredo et al. Citation2022). B – Praia do Areia do Mastro outcrop, with the area of the Papo-Seco Formation on the stratigraphic column marked by a black rectangle.
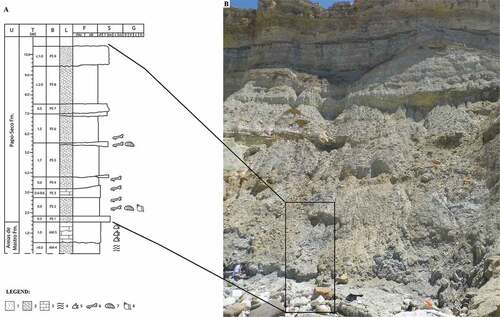
In the studied area, two geological formations occur: the Areia do Mastro and Papo-Seco formations. At the Boca do Chapim and Praia do Areia do Mastro sites, most of the ornithopod remains were discovered in the lowermost beds of the Papo-Seco Formation, which are overlain by two strata of thin yellowish sandstones. From the lower to the uppermost portion of the exposed succession of the Papo-Seco Formation, the deposits consist of grey marls, grey carbonate silts and lenses of fine sandstones, with fossils of gastropods and bivalves (). In the Praia do Areia do Mastro site, the discovery of vertebrate bones and teeth belonging to a diverse fauna, including crocodyliforms, pterosaurs, turtles and dinosaurs, including ornithopods, has previously been reported (Figueiredo et al. Citation2015, Citation2016, Citation2018, Citation2020b). The alluvial debris from the cliffs are found mostly in place without any serious transitional flow.
Figure 3. Isolated teeth of Ornithopoda indet. from the Papo-Seco Formation of Portugal. Dentary teeth MG8760 (A) and MG10 (B); maxillary tooth CPGP.1.03.3 (C). Scale: 2 cm.

Materials and methods
Ornithopod material from the Papo-Seco Formation consists of both cranial and postcranial remains from several individuals. Cranial remains are represented by isolated teeth; postcranial elements include vertebral remains and a fragmentary limb bone (see ) and a partial skeleton of disarticulated bones from the axial and appendicular regions (see Figueiredo et al. Citation2015, Citation2020b).
Table 1. Ornithopods from the Lower Cretaceous (Barremian) of the Papo Seco Formation - Cape Espichel, Portugal.
The ornithopod material reported here includes previous specimens and remains recently found (1999-2018). Old finds were described by Lapparent and Zbyszewski (Citation1957; see also Sauvage Citation1897-1898) and consist of nine fossils from Boca do Chapim. These remains are five isolated teeth, three vertebrae, a distal fragment of a femur, and six bone fragments; housed at the Geological Museum (MG) and in the National Museum of Natural History of Lisbon University (MNHNUL/MUHNAC). New specimens include a partial skeleton with disarticulated vertebral and appendicular remains from Boca do Chapim site (Figueiredo Citation2010), one tooth and one vertebra from Praia do Areia do Mastro site, and one vertebral fragment, a prezygapophysis, from Praia do Guincho site. All these materials are housed in the collection of the Portuguese Centre of Geohistory and Prehistory (CPGP). The study of the new fossils was conducted in the Laboratory of Archaeozoology and Palaeontology of the CPGP.
Institutional abbreviations
CPGP: Centro Português de Geo-História e Pré-História (Portuguese Centre of Geohistory and Prehistory); MG: Museu Geológico (Geological Museum); MNHNUL/MUHNAC: Museu Nacional de História Natural da Universidade de Lisboa (National Museum of Natural History of Lisbon University)/Museu Nacional de História Natural e da Ciência (National Museum of Natural History and Science)
Systematic palaeontology
Cranial remains
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Ornithopoda Marsh, 1881
Ornithopoda indet.
Material: MG10, MG8760, two very fragmentary and worn crowns of dentary teeth ( A and B).
Locality and horizon: Boca do Chapim locality; Papo-Seco Formation, lower Barremian.
Description: These crowns are very fragmentary and worn. MG10 () is a small fragment of a tooth that has a large central ridge on the labial surface and a marginal smaller ridge on the mesial margin. The lingual surface lacks ornamentation. MG8760 () is larger, but the tooth is deformed and badly preserved, its crown is 52.3 mm high and 15.9 mm mesiodistally wide, giving a height-to-width ratio of 3.28 (); it does not preserve any ornamentation.
Table 2. Measurements of the ornithopod teeth from the Papo-Seco Formation referred in this study. Abbreviations: LApex – Apex Length; Wapex - Apex Weight; CH - crown height; H-Wr -Height-to-width ratio; CBL - crown base length; MW - mesiodistally width; RCH/MW: ratio crown height/mesiodistally width.
Discussion: MG8760 () and MG 10 () were described and referred to Iguanodon mantelli by Lapparent and Zbyszewski (Citation1957) and latter attributed to Iguanodon sp. (Galton Citation1994). MG 10 preserves the characteristic marginal crests present on the teeth of ornithopod dinosaurs (Norman Citation1980, Citation2011; Araújo et al. Citation2011). The fragmentary and distorted nature of these teeth does not allow to assign them to a specific group of ornithopods; so, they are referred to Ornithopoda indet.
Material: CPGP.1.03.3, one isolated maxillary tooth ().
Locality and horizon: Praia do Areia do Mastro site; Papo-Seco Formation, lower Barremian.
Description: CPGP.1.03.3 is a fragmentary crown tooth, very worn, especially on the occlusal surface. It has no ornamentation. The distalmost surface is flat, making an angle with the lingual zone; the mesial area is curved (Figueiredo et al. Citation2015).
Discussion: This tooth was previously described by Figueiredo et al. (Citation2015) and referred to Ornithopoda indet. Its worn and fragmentary nature does not allow for a more accurate assignment.
Iguanodontia Dollo Citation1888
Iguanodontia indet.
Material: MG 4744, is an isolated maxillary tooth (). It was discovered in 1951 by Octávio da Veiga Ferreira and described by Lapparent and Zbyszewski (Citation1957).
Figure 4. Isolated teeth of Iguanodontia indet. from the Papo-Seco Formation of Portugal. Dentary teeth MNHNUL.DIN.002 (A-C) and MNHNUL.DIN.003 (D-G): A and D – labial surface; B and E - posterior surface; C and F- lingual surface. G – Illustration of Lapparent & Zbyzsewski (Citation1957) with complete tooth MNHNUL.DIN.003. Maxillary tooth MG4744 (H, I J): H - labial surface; I - lingual surface; J - posterior surface. Maxillary tooth CPGP.1.18.44 (L-N): L - labial surface; M - lingual surface; N - posterior surface. Maxillary tooth CPGP.1.01.1 (O-Q). O – labial surface; P - lingual surface; Q - posterior surface Abbreviations: d – lingulate denticles; eb – enamel border; irs – inter-ridge surface; mde – marginal denticles; Nc – neck; pr – primary ridge; sr – secondary ridges. Scale: 2 cm

Locality and horizon: Boca do Chapim; Papo Seco Formation, Barremian.
Description: MG4744 is a rather worn maxillary tooth with a crown in good condition. It is labiobucally narrow at the base of the crown; on the buccal side, it is thickly enameled and has a very large primary ridge, arising from the base, on the distal side; it has three small accessory ridges at the distal surface, two of them being bifurcated in the basal region of the tooth, and three more marginal ridges in the mesial surface. The three marginal ridges are shorter than the ones of the anterior surface. MG4744 has nine marginal denticles along the mesial margin and nine more denticles on the distal margin. At the posterior side, the marginal edge is curved. The lingual surface is slightly convex without any ornamentation.
Discussion: This maxillary tooth has labially a primary ridge that is displaced distally on the crown, multiple secondary ridges and marginal denticles that are mammillate, as typical in iguanodontians (Norman Citation2011, Citation2015). This tooth has ridges confluent with marginal denticles; the tooth crown is about 50% higher than mesiodistally wide ( – RCH/MW), this is a synapomorphy of Iguanodontia; (Weishampel and Heinrich Citation1992).
Material: CPGP.1.18.44, isolated tooth crown ().
Locality and horizon: Praia do Areia do Mastro; Papo Seco Formation, Barremian. Found in 2018 in the lower beds of the formation at the base of the cliff (layer PS3; ).
Description: CPGP.1.18.44 is an isolated right maxillary tooth crown. It is narrow at the base of the crown; in buccal view, it is thickly enamelled and has a very large primary ridge, arising from the base, on the anterior side; it has six small accessory ridges on the anterior surface and three in the posterior edge. The crown is heavily worn without the apex or marginal denticles, by tooth-to-tooth occlusion. From the posterior viewpoint, the marginal edge is curved. The lingual surface, which still conserves part of the matrix of the sediment from the layer where the specimen was found, is slightly convex and without any ornamentation.
Discussion: This tooth exhibits features similar to other teeth found in the Papo Seco Formation (see above) and is therefore assigned to Iguanodontia indet. It has a lozenge-shaped, in labial view and presents the primary ridge displaced distally and the tooth crown is almost 50% higher than mesiodistally wide ( – RCH/MW: the fact that this dental crown is heavily worn, without the apex, results in a ratio crown higher/mesiodistal width lower than it should be). Those are a synapomorphy of Iguanodontia (Weishampel and Heinrich Citation1992; Paul Citation2007).
Material: CPGP.1.99.7, a left maxilla fragment with five teeth (); CPGP.1.01.1, a tooth crown ().
Figure 5. Maxilla fragment of Iguanodontia indet. CPGP.1.99.7. A - lingual view; B - buccal view (inverted image); C - buccal view of crown 4; D - dorsal view of crown 4; E and E - buccal views of crown 5. Abbreviations: Form – foramina; m - mammilla; mde - marginal denticules; Pr - primary ridge; Sr - secondary crown ridges. Scales: A and B - 2 cm; C to F - 1 cm.

Locality and horizon: Praia do Areia do Mastro; Papo-Seco Formation, Barremian.
Description: The maxilla fragment (CPGP.1.99.7) is 9.8 cm long. It has three foramina, which circumscribe the alveolar parapet. The maxilla preserves five tooth crowns: three are complete (crowns 2, 4 and 5; 2 and 5 are beginning to erupt, see A and B) and two are broken (two of them with the crown completely above the alveolar parapet, one with the apex above and two still within the alveoli). An X-ray showed that it possesses one active and one replacement tooth per alveolar position and shows that the dental alveoli are wide and deep, revealing a long root (see Figueiredo et al. Citation2015), these are characters of Iguanodontia, described by Norman Citation2011. The teeth, including those of the maxilla and the isolated specimen CPGP.1.01.2, have a thickly enamelled labial surface of the crown. The teeth 4 and 5 () have a strongly denticulate margin, with eight denticules, in the distal margin, and eleven, in the mesial margin. The lingual surfaces of the teeth do not display any ornamentation. The crown has a very large primary ridge arising from the base and seven (four on the distal surface and three on the mesial surface) secondary ridges. Like the maxillary tooth of Brighstoneus simmondsi and others iguanodontidians, the teeth of CPGP.1.99.7 have mammillae on the marginal denticles approaching the apical region of the crown (Lockwood et al. Citation2021). On crown 4, the apical margin is mamillated, with seven small and spherical mammillae, reducing to two at the apex ( C, D). Crown 5 (an emergent crown) has the same number of mammillae, reducing to one at the apex.
The isolated tooth CPGP.1.01.2 () shows features similar to the third maxillary tooth of CPGP.1.99.7 (). It bears eight accessory ridges between the primary ridge and the marginal ridges (five between the primary ridge and the anterior marginal ridge and three between the primary ridge and the posterior marginal ridge). The lingual surface is slightly convex and without any ornamentation.
Discussion: This material from Praia do Areia do Mastro was previously described by Figueiredo et al. (Citation2015) and referred to Iguanodontidae indet. The validity of the family Iguanodontidae is the subject of current debate and its monophyly has been questioned (see e.g. Norman Citation2002; Paul Citation2007; Lockwood et al. Citation2021; Madzia et al. Citation2021 and references therein). The combination of characters in these teeth, such as one replacement tooth per tooth position, as identified in the X-ray on CPGP.1.99.7, the labial surface of the crown thickly enameled, strongly denticulate margin and with a very large primary ridge arising from the base of the crown, suggests that they belong to the clade Iguanodontia (Norman and Weishampel Citation2004; Paul Citation2007, 2011; Norman Citation2011; Citation2013). The features observed in the teeth resembled those of Iguanodon and Mantellisaurus, as described by Paul (Citation2007, 2011), Norman (Citation2011, Citation2013) and Verdú (Citation2017). They have a strongly denticulate margin and both mesial and distal sides excavated to accommodate adjacent teeth. A very large ridge arising from the base of the crown and extending to the apex dominates the labial surface. This primary ridge asymmetrically divides the enameled, labial surface of the crown. Distal to the primary ridge, the surface is narrow and smooth while the mesial surface is wider and has several faint accessory ridges, which extend parallel to the primary ridge; the teeth do not display any ornamentation on the lingual surface. The size of the maxillary fragment is more consistent with Mantellisaurus than with Iguanodon (Figueiredo et al., Citation2015), but there are no unequivocal diagnostic characters allowing to assign it to this taxon. Until the discovery of more complete material, CPGP.1.99.7 and CPGP.1.01.1 are provisionally assigned to Iguanodontia indet.
Styracosterna Sereno Citation1986
Cf. Styracosterna indet.
Material: MNHNUL.DIN.002; MNHNUL.DIN.003, two isolated dentary teeth (). These teeth were initially described and published by Sauvage Citation1897-1998-98) and then by Lapparent and Zbyszewski (Citation1957).
Locality and horizon: Boca do Chapim; Papo Seco Formation, Barremian.
Description: MNHNULDIN.002 () does not preserve the root; however, the apicobasal region is still well preserved. The crown is 51.4 mm high and 31.8 mm mesiodistally wide, the height-to-width ratio is 1.61 (). The crown is mesiodistally expanded with a wide, flattened lingual surface. The enamel is limited to the lingual side of the crown and to the marginal denticles. Both mesial and distal surfaces of the labial face are concave. A primary ridge runs along the entire height of the crown, widening at the base of the crown and is displaced slightly distally relative to the center of the lingual surface, demarcating two relatively deep sulci and dividing the surface in two unequal parts. Four shallow, secondary ridges, one located distally, and three mesially relative to the primary ridge, extend parallel to the primary one along the enameled surface. Denticles are present on both mesial and distal crown margins, with 17 along the distal margin and 11 along the mesial margin. Marginal denticles display mammillae and are graded in size, being smaller towards the base of the crown. The lingual surface of the tooth is broad and smooth.
The crown of MNHNULDIN.003 was complete at the time of the publication by Lapparent and Zbyszewski (Citation1957, pl. XII, fig. 12; see ). It is currently incomplete having lost its apical region (). The crown has a maximum length of 48.1 mm and a mesiodistal width of 30.7 mm (). The crown is mesiodistally expanded with a wide, flattened lingual surface. Neither ridge nor the apicobasal region are preserved. Part of the central root axis and the basal region is preserved. The crown widens at its base and is displaced slightly distally relative to the center of the lingual surface. Three denticles are present on the mesial margin, which are mammillate and graded in size. The lingual surface of the tooth is broad and smooth.
Discussion: Both dentary crowns exhibit a prominent ridge with subsidiary ridges on either side, a common morphology (clypeodont condition of Norman, 2014) observed in hypsilophodontians and iguanodontians (see Norman Citation2015; Dieudonné et al., 2020). Moreover, the primary ridge of the crowns is slightly distally offset and the marginal denticles on the sides of the crowns bear small mammillae, as is typical in Iguanodontia (Norman Citation2011, Citation2015). In unworn dentary teeth, such as MNHNUL.DIN.002, the crown margin is truncated and shows a distinct “shoulder” mesial to the tip of the crown formed at the intersection of the primary ridge with the crown margin, as occurs in Ankylopollexia (sensu Norman Citation2015). Also, the enameled crown face of the dentary tooth MNHNUL.DIN.002 is inclined posterodorsally, as has been described for Styracosterna (Norman Citation2015). Therefore, these dentary teeth recovered from the Papo Seco Formation exhibit iguanodontian characters and could belong to a member of Styracosterna.
Postcranial remains
Ornithopoda indet.
Material: CPGP.02.07.1, partial skeleton including disarticulated bones from the axial and appendicular skeleton of a single individual (). It consists of several restored remains: four vertebrae, five chevrons, two neural spines, a dorsal rib, one left ischium and one phalanx. Other materials are still in the restoration and preparation process. The first material (two chevrons) was discovered in 2007 and then an excavation of an area of 6 m2 was carried out between 2008 and 2012 (Figueiredo Citation2010, Citation2014). Associated fossils found in the same level belong to other vertebrates (theropod teeth, pterosaur phalanx, turtle shell fragment), bivalves and plants (wood debris) (see Figueiredo Citation2010; Citation2014; Figueiredo et al. Citation2018, 2020).
Figure 6. Partial skeleton CPGP.02.07.1 provisionally referred to Ornithopoda indet. Representation of some of the material already restored (A - one of the four caudal vertebrae, B - dorsal rib, C - neural spine, D - haemal arch, E – ischium, F – phalanx) and its localization on an ornithopod skeleton

Locality and horizon: Boca do Chapim site: Papo-Seco Formation, lower Barremian. The material was found in the basal marly grey bed of this formation (Figueiredo Citation2010). Based on taphonomic and sedimentological features, it is suggested that the ornithopod carcass was deposited in a low-energy environment or stagnant water (estuary/lagoon) and could have been exposed, at least partially, for some time.
Description: The CPGP.02.07.1 material as it is still in the process of preparation and study. These remains correspond to a large-sized individual. This is the most complete ornithopod material discovered to date in the Papo-Seco Formation. A caudal vertebra shows evidence of bite marks produced by conical teeth ().
Discussion: Without a detailed description of the specimen and its affinities, it is provisionally identified as Ornithopoda indet. (Figueiredo Citation2010). The relative size of the bones suggest that they belong to a large ornithopod.
Material: CPGP.3.13.5a, isolated prezygapophysis (). Found during fieldwork in 2013.
Figure 7. Vertebral remain (prezygapophysis) of Ornithopoda indet. CPGP.3.13.5a. A and C – lateral views; B – anterior view.

Locality and horizon: Praia do Guincho site; Papo-Seco Formation, lower Barremian. The fossil comes from the upper beds of the formation.
Description: It is a fragment of the articular facet of a prezygapophysis. The articular surface is concave. Maximum length 5.3 cm; width 2.3 cm.
Discussion: Based on its morphology and dimensions, this fragmentary element could correspond to a large ornithopod.
Material: MG 4739: fragment of a vertebral articular facet ( - E; MG17: fragment of a vertebra ( -F); MG 8761: fragment of a proximal part of a chevron ( -A) and MG 8759.3: three fragments of chevrons ( -B-8-D).
Figure 8. Indeterminate ornithopod bones: A – MG 8761; B to D – MG 8759.3; E – MG 4739; F – MG 17). G – Label of one of the unpublished specimens of Museu Geologico (MG 8761) with the identification “Iguanodon” and Boca do Chapim.

Locality and horizon: Boca do Chapim site; Papo Seco Formation, lower Barremian.
Description: MG 4739 is a fragment of an articular facet of a vertebral centrum, the articular surface is flat and is wider than it is high (11.2 cm wide and 9 cm high). MG 8761 is a proximal fragment of a chevron, MG 8759.3 consists of three fragments of chevrons, and MG 17 is a fragment of the top of the neural canal of a vertebra, it preserves the postzygapophysis and the base of the neural spine.
Discussion: This set of fragments of bones is unpublished and they are referred to in the inventory of the MG collection as Iguanodon remains from Boca do Chapim (). There is no other information about these fossils, they are in the same box and present the same type of fossilization as MG 4741 (- above), so they may have come from the same bed and eventually belong to the same individual. If they belong to the same individual as MG 4741, these remains could be referred to Iguanodontia indet.. However, the lack of information and their fragmentary condition does not allow a more precise taxonomic classification than Ornithopoda indet.
Figure 9. Axial and appendicular skeleton remains: 1 and 2 - caudal vertebrae of Iguanodontoidea indet.: 1 - MG4741 (above). MG 13.2 (middle) and MG 13.1 (down). A – cranial view; B – lateral view; C – caudal view; 2 - CPGP.1.14.18. A- cranial view; C- caudal view; B and D – lateral views. Abbreviations: C – centrum; fac – articular faces; nc – neural channel; tp – transverse process; 3 - Femur of Iguanodontoidea indet. MG4740 A – cranial view; B – caudal view; C – Medial view. Abbreviations: bu: buttress; ieg: intercondylar extensor groove; ifg: intercondylar flexor groove; lc: lateral condyle; mc: medial condyle.

Iguanodontia indet.
Material: MG 13.1, MG 13.2 and MG 4741, are three caudal vertebrae (); MG4740, is a distal end of a right femur (). MG 13.1-2, and MG 4740 show the same type of fossilization as MG 4739, so they may belong to the same individual; MG 4741 shows a different fossilization type (it has a minor degree of permineralization and recrystallization, less petrification and a different colour than the MG 13.1-2, and MG 4740), hence it might belong to a distinct individual and coming from another bed.
Locality and horizon: Boca do Chapim site; Papo-Seco Formation, lower Barremian.
Description: Only the centrum and the basal most part of the neural arch are preserved in the three vertebrae. MG4741 exhibits a spool-shaped and flat to shallowly concave centrum, with concave articular facets. MG 13.1 and MG13.2 does not exhibit a spool-shaped centrum but has concave articular facets. The centra of MG 13.1 and MG 13.2 are higher than wide and long, while in MG 4741 the centrum is longer than it is high and wide. MG4741 is somewhat deformed due to thaphonomic compression. The neural canal of these vertebrae seems to be narrow basally. MG 13.1 and MG13.2 come from the anterior part of the caudal region; MG4741 is a dorsal vertebra.
MG 4740 is a distal end of a large femur (maximum width: 24 cm). It is expanded, with the medial condyle being larger and more robust than the lateral condyle (). A prominent and vertical ridge extends on the lateral side of the lateral condyle. The intercondylar extensor groove is large, U-shaped and is enclosed by the lateral and medial edges of both condyles. Posteriorly, the intercondylar flexor groove is fully open but partially enclosed by the lateral inflation of the medial condyle ().
Discussion: Lapparent and Zbyszewski (Citation1957) described these remains and referred them to Iguanodon mantelli. The features observed in the femur are similar to those of iguanodontians as described by Norman (Citation2011): in cranial view, it shows a deep intercondylar extensor groove; in caudal view, it presents a deep intercondylar flexor groove ( – 3B) and in lateral view, it shows a strongly curved expansion of the articular surface, producing a buttress ( – 3C), so the remains are here assigned to Iguanodontia indet. The deep intercondylar flexor grove is a synapomorphy for Iguanodontia (Weishampel and Heinrich Citation1992). The dimensions of the caudal vertebrae are compatible with those of a large Iguanodon-like iguanodontian, but the fragmentary nature of the material and the absence of diagnostic characters do not allow to assign the remains to the genus Iguanodon.
Material: CPGP.1.14.18, anterior caudal vertebra (). Found during fieldwork in 2014.
Locality and horizon: Praia do Areia do Mastro site; Papo-Seco Formation, lower Barremian. The fossil comes from the upper beds of the formation.
Description: CPGP.1.14.18, this vertebra, fallen down from the cliff, preserves the centrum and part of the right transverse process. It is 56 mm long, 92 mm high and 73 mm wide. The centrum is amphiplatyan and forms a hexagonal cylinder. The articular surfaces are wider than high (67 mm wide and 54 mm high in the anterior surface, 65 mm wide and 57 mm high in the posterior surface). The neural canal is narrow.
Discussion: Features such as the hexagonal form of the articular surfaces of the vertebra, the amphicoelous condition of the centrum and its relative size are consistent with those observed in the caudal vertebrae of iguanodontian ornithopods (Norman Citation2004; Verdú Citation2017; Verdú et al. 2019).
Discussion
Iguanodontians are ornithopod dinosaurs of medium to large size, herbivores and primarily bipedal (Norman Citation2004). Iguanodontia consists of multiple genera and species, which were typical components of the Early Cretaceous dinosaur faunas, known throughout Europe, especially in Western Europe (e.g. Norman Citation2004; Citation2011; Weishampel et al. Citation2004; Pereda-Suberbiola et al. 2012; Verdú et al. Citation2017), but also in North America and Asia. Large iguanodontians from the Barremian of Europe are highly diverse and include the styracosternans Iguanodon bernissartensis, Iguanodon galvensis, Mantellisaurus atherfieldensis, Delapparentia turolensis, Morelladon beltrani, Magnamanus soriaensis, Portellsaurus beltrani and Brighstoneus simmondsi (see Norman Citation2011; Ruiz-Omeñaca Citation2011; Gasulla et al. Citation2015; Fuentes-Vidarte et al. Citation2016; Verdú et al. 2018; Santos-Cubedo et al. 2021; Lockwood et al. Citation2021 and references therein). All of them but Brighstoneus are represented by fossils in the Iberian (Spanish) Barremian sites. Of these taxa, only Iguanodon galvensis, Portellsaurus and Brighstoneus are known from the early Barremian, the first two from Spain (Teruel and Castellón, respectively) and the third from England (Isle of Wight). The temporal distribution of Magnamanus from Soria (Spain) is late Hauterivian-early Barremian. The relationships of these and other Wealden ornithopods are subject to debate. Some taxa, such as Iguanodon species, are included within the clade Iguanodontoidea, others are considered to be basal styracosternans, basal hadrosauriforms, or even basal hadrosauroids (see phylogenetic hypotheses of Norman Citation2015; Verdú et al. 2018; Lockwood et al. Citation2021; Santos-Cubedo et al. 2021).
The presence of Iguanodon in the lower Barremian Papo-Seco Formation, at Boca do Chapim site has been suggested by Lapparent and Zbyszewski (Citation1957). The occurrence of Mantellisaurus in the same geological unit and site has also been suggested (see Crespo Citation2001; Figueiredo et al. Citation2015). However, the available material does not allow us to recognize the presence of these taxa based on diagnostic features (see Norman Citation1980, 1986, Citation2011; Citation2012). Teeth found in the deposits of the Papo Seco Formation are the most informative elements of the ornithopod assemblage. These teeth exhibit common features of Iguanodontia (sensu Norman Citation2011; Citation2015; see Systematic Palaeontology). Furthermore, the morphology of the dentary teeth from Boca do Chapim site (MNHNULDIN.002 and tentatively MNHNULDIN.003) should be referred to as cf. Styracosterna indet.
In the Boca do chapim site and Praia do Areia do Mastro site, in the uppermost beds of the Areia do Mastro Formation (underlying the first beds of the Papo Seco Formation), tracks belonging to the ichnogenus Caririchnium have been described (Figueiredo et al. Citation2017, Citation2021, Citation2022). Following Leonardi (Citation1979) and Díaz-Martínez et al. (2015), this ichnogenus can be assigned to iguan odontian ornithopods; the producer of the tracks may be a basal member of Ankylopollexia or a basal Styracosterna. In the Praia do Guincho site, in the Papo Seco Formation, a natural footprint cast of an ornithopod was discovered in a small coarse-grained limestone block (Figueiredo et al. Citation2017). The cast is an average tridactyl footprint from a right pes and has a plantigrade and rounded form. It has short toes with blunt, rounded ends. This cast was also attributed to a Caririchnium (Figueiredo et al., Citation2022). The footprints of Areia do Mastro and Papo-Seco formations assigned to the ichnogenus Caririchnium complement the evidence of the ornithopod dinosaurs from these formations provided by bone fossil remains.
Conclusions
The outcrops of the Lower Cretaceous Papo Seco Formation at Cape Espichel (Lusitanian Basin, Portugal) have yielded to date an important set of fossil remains of aquatic, semiaquatic and terrestrial vertebrates, as well as bivalves and plants. The vertebrate fossil association, of early Barremian age, includes dinosaurs, crocodyliforms, pterosaurs, turtles and bony fish. Dinosaurs are represented by theropods, sauropods and ornithopods. The dinosaur remains could have originated from near inland river subenvironments and later transported to marine, estuarine and lagoon environments, where they finally were deposited.
Ornithopod fossils from the Papo Seco Formation have been discovered in three sites: Boca do Chapim (south), Praia do Areia do Mastro and Praia do Guincho (north). Most of the remains have been collected from the lower beds of the formation at the base of the cliffs. The material consists of cranial (i.e. maxilla fragment with teeth, isolated teeth) and postcranial (mainly vertebrae and other axial elements, a few appendicular bones) remains. Several different individuals are represented in the assemblage, the most complete being a partial skeleton (still undescribed) found at Boca do Chapim site.
The presence of Iguanodon and Mantellisaurus in the Papo Seco Formation of Portugal cannot be confirmed based on the available material, despite having been previously suggested in the literature. Ornithopod remains studied here are classified as Ornithopoda indeterminate, Iguanodontia indeterminate and cf. Styracosterna indeterminate. The tooth morphology and the relative size of the remains suggest that the presence of member of a Styracosterna is likely. Without ruling out that these taxa may be represented in the deposits of this formation, it seems necessary to discover more complete material in order to identify them. The new ornithopod material described in this work increases the record of this group of dinosaurs in the Iberian Early Cretaceous, and more specifically in the lower Barremian of Portugal.
Acknowledgements
This study was supported by CPGP (Portuguese Centre of Geohistory and Prehistory), Lisbon/Golegã through project PALEOGE-ORCI_17; by the FCT (Science and Technology Foundation), through projects UID/Multi/00073/2020 (Geosciences Center - Coimbra University) and UID/MAR/04292/2020 (MARE); and Ciência Viva – Agencia Nacional para a Cultura Científica e Tecnológica, which has financially supported part of the field work betwwen 2002 and 2018 through the program “Ocupação Científica dos Jovens nas Férias” (Scientific Occupation of Youngsters During Holidays), involving young people of the secondary school level. We would like to thank all the team members who have collaborated in the fieldwork. The research work of Ismar de Souza Carvalho was funded by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (Proc. E-26/200.828/2021, Brazil) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 303596/2016-3, Brazil).The research work of Xabier Pereda Suberbiola has been supported by the project CGL2017-85038-P of the Spanish Ministry of Science, Innovation and Universities and the European Regional Development Fund, and by the research group IT418-19 of the Basque Gouvernement.
The authors would also like to thank to the Museu Geológico (Lisbon), in particular to Miguel Ramalho and Jorge Sequeira and the Museu Nacional de História Natural (Lisbon), in particular to Liliana Póvoas, for the authorization to take the photos of the materials of these museums that illustrate this work.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Araújo R, Castanhinha R, Mateus O. 2011. Evolutionary major trends of ornithopod dinosaurs teeth. In: Calvo J, Porfiri JD, González Riga B, Dos Santos D, editors. Dinosaurios y Paleontologıa desde América Latina. Mendoza, Argentina: EDIUNC, Editorial de la Universidad Nacional de Cuyo; p. 25–31.
- Buffetaut E. 2007. The spinosaurid dinosaur Baryonyx (Saurischia, Theropoda) in the Early Cretaceous of Portugal. Geol Mag. 144(6):1021–1025. doi:10.1017/S0016756807003883.
- Choffat P. 1904. Le Cretacique dans l’Arrabida et dans la contrée de Ericeira. Comunicações dos Serviços Geológicos de Portugal. VI(7):1–55.
- Crespo EG. 2001. Paleo-Herpetofauna de Portugal. Museu Bocage. Lisboa.
- Dinis PA, Dinis JL, Mendes MM, Rey J, Pais J. 2016. Geochemistry and mineralogy of the Lower Cretaceous of the Lusitanian Basin (western Portugal): Deciphering palaeoclimates from weathering indices and integrated vegetational data. Comptes Rendus Geoscience. 348(2):139–149. doi:10.1016/j.crte.2015.09.003.
- Dollo L. 1888. Iguanodontidae et Camptonotidae. Comptes Rendus de Academie des Sciences. Paris. 106:775–777.
- Figueiredo SD. 2000. Classificação do dente n° 10 do Museu Geológico. Boletim do Centro Português de Geo-História e Pré-História. I(1):2–3.
- Figueiredo SD. 2008. Dinossauros de Portugal. Edições Cosmos. Chamusca.
- Figueiredo SD. 2010. Breve Notícia sobre a Descoberta de um Novo Dinossáurio Ornitópode no Cabo Espichel. Boletim do Centro Português de Geo-História e Pré-História. III(4):7–15.
- Figueiredo SD. 2014. Os Dinossáurios em Território Português: as espécies, as jazidas e os fósseis. Chiado Editora, Lisboa.
- Figueiredo SD, Bachtsevanidou Strantzali I, Gomes M, Rita P, Santos M. 2017. Preliminary Data of New Dinosaurs and Turtles Remains from the basal Deposits of the Papo-Seco Formation in Areia do Mastro’s Quarry (Cabo Espichel - Portugal) Arnava. VI. 117–130.
- Figueiredo SD, Carvalho CN, Cunha PP, Carvalho IS. 2021. New Dinosaur Tracks from the Lower Barremian of Portugal (Areia do Mastro Formation, Cape Espichel). Journal of Geoscience and Environment Protection. 9(1):84–96. doi:10.4236/gep.2021.91007.
- Figueiredo SD, Carvalho IS, Pereda-Suberbiola X, Cunha PP, Antunes V, Dıaz-Martínez I. 2022. New ornithopod footprints from the Areia do Mastro Formation (Lower Cretaceous), Espichel Cape (Portugal, Western Iberia) and their context in the Iberian ichnological ornithopod record. Cretaceous Research. 131:105069. doi:10.1016/j.cretres.2021.105069
- Figueiredo SD, Pereda-Suberbiola X, Cunha PP, Sousa F, Rosina P. 2020b. A brief approach about the Ornithopod dinosaurs from the Papo-Seco Formation (Cabo Espichel, western central Portugal) Boletim do Centro Português de Geo-História e Pré-História. 2(1):59–63.
- Figueiredo SD, Rosina P, Figuti L. 2015. Dinosaurs and other vertebrates from the Papo-Seco Formation (Lower Cretaceous) of southern Portugal. Journal of Iberian Geology. 41(3):301–314. doi:10.5209/rev_JIGE.2015.v41.n3.47828.
- Figueiredo SD, Rosina P, Strantzali I, Antunes V, Figueiredo S. 2020a. Paleoenvironmental Changing Approach on the Lower Cretaceous Sequences of Areia do Mastro (Cabo Espichel, Southern Portugal). J Environ Sci Eng A. 9:66–71. doi:10.17265/2162-5298/2020.02.003
- Figueiredo SD, Strantzali I, Rosina P, Gomes M. 2016. New Data about the Paleo Environment of the Papo-Seco Formation (Lower Cretaceous) of Southern Portugal. Journal of Environmental Science and Engineering A. 5:463–470.
- Figueiredo SD, Suberbiola XP, Cunha PP, Sousa MF, Gomes M 2018. Ornithopods dinosaurs from the Papo-Seco Formation (Cabo Espichel, western central Portugal). Abstract book of XVI Meeting of the European Association of Vertebrate Paleontologists. P. 71
- Fuentes-Vidarte C, Meijide M, Meijide F, Meijide M. 2016. Un nuevo dinosaurio estiracosterno (Ornithopoda: Ankylopollexia) del Cretácico Inferior de España. Spanish Journal of Palaeontology. 31(2):407–446. doi:10.7203/sjp.31.2.17163.
- Galton PM. 1994. Notes on Dinosauria and Pterodactylia from the Cretaceous of Portugal. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 194(2–3):253–267. doi:10.1127/njgpa/194/1994/253.
- Gasca JM, Canudo JI, Moreno-Azanza M. 2014. On the diversity of Iberian iguanodont dinosaurs: New fossils from the lower Barremian, Teruel province, Spain. Cretaceous Research. 50:264–272. doi:10.1016/j.cretres.2014.05.009
- Gasca JM, Moreno-Azanza JM, Ruiz-Omeñaca JI, Canudo JI. 2015. New material and phylogenetic position of the basal iguanodont dinosaur Delapparentia turolensis from the Barremian (Early Cretaceous) of Spain. Journal of Iberian Geology. 41(1):57–70. doi:10.5209/rev_JIGE.2015.v41.n1.48655.
- Gasulla JM, Escaso F, Narváez I, Ortega F, Sanz JL. 2015. A New Sail-Backed Styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PLoS ONE. 10(12):e0144167. doi:10.1371/journal.pone.0144167.
- Gasulla JM, Escaso F, Ortega F, Sanz JL. 2014. New hadrosauriform cranial remains from the Arcillas de Morella Formation (lower Aptian) of Morella, Spain. Cretaceous Research. 47:19–24. doi:10.1016/j.cretres.2013.10.004
- Lambe LM. 1902. On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous). Contributions to Canadian Paleontology. 3:25–81.
- Lapparent AF, Zbyszewski G. 1957. Les dinosauriens du Portugal. Memórias dos Serviços Geológicos de Portugal, Lisboa. 2: 1-63.
- Leonardi G. 1979. Nota preliminar sobre seis pistas de dinosaurios Ornithischia da Bacia do Rio do Peixe, em Souse, Paraiba, Brasil. Anais da Academia Brasileira de Ciências. 51:501–516.
- Lockwood JAF, Martill DM, Maidment SCR. 2021. A new hadrosauriform dinosaur from the Wessex Formation, Wealden Group (Early Cretaceous), of the Isle of Wight, southern England. Journal of Systematic Palaeontology. 19(12):847–888. doi:10.1080/14772019.2021.1978005.
- Madzia D, Arbour VM, Boyd CA, Farke AA, Cruzado-Caballero P, Evans DC. 2021. The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ. 9:e12362. doi:10.7717/peerj.12362
- Manupella G, Antunes MT, Pais J, Ramalho MM, Rey J. 1999. Notícia Explicativa da Carta Geológica de Setúbal. Serviços Geológicos de Portugal. 143.
- Marsh O. 1882. Classification of the Dinosauria. American Journal of Sciences. 3:81–86.
- Mateus O, Araújo R, Natário C, Castanhinha R. 2011. A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus. Zootaxa. 2827(1):54–68. doi:10.11646/zootaxa.2827.1.3.
- Mateus O, Estraviz-López D. 2022. A new theropod dinosaur from the early cretaceous (Barremian) of Cabo Espichel, Portugal: Implications for spinosaurid evolution. PLoS ONE. 17(2):e0262614. pone.0262614. doi:10.1371/journal.
- McDonald AT. 2012a. Phylogeny of basal iguanodonts (Dinosauria: Ornithischia): An update. PLoS ONE. 7(5):e36745. pone.0036745. doi:10.1371/journal.
- McDonald AT. 2012b. The status of Dollodon and other basal iguanodonts (Dinosauria: Ornithischia) from the Lower Cretaceous of Europe. Cretaceous Research. 33(1):1–6. doi:10.1016/j.cretres.2011.03.002.
- Norman DB. 1980. On the ornithischian dinosaur Iguanodon bernissartensis from the Lower Cretaceous of Bernissart (Belgium) Mémoires de L’Institut Royal des Sciences Naturalles de Belgique. 178. 1–105.
- Norman DB. 2002. On Asian ornithopods (Dinosauria: Ornithischia). 4. Redescription of Probactrosaurus gobiensis Rozhdestvensky, 1966. Zoological Journal of the Linnean Society (London). 136(1):113–144. doi:10.1046/j.1096-3642.2002.00027.x.
- Norman DB. 2004. Basal Iguanodontia. In: Weishampel DB, Osmólska H, Dodson P, editors. The Dinosauria. Berkeley: University of California Press; p. 413–437.
- Norman DB. 2010. A taxonomy of iguanodontians (Dinosauria: Ornithopoda) from the lower Wealden Group (Cretaceous: Valanginian) of southern England. Zootaxa. 2489(1):47–66. doi:10.11646/zootaxa.2489.1.3.
- Norman DB. 2011. The ornithopod dinosaurs. In: Batten D, editor. English Wealden fossils. The Paleontological Association. London; p. 407–475.
- Norman DB. 2012. Iguanodontian taxa (Dinosauria: Ornithischia) from the Lower Cretaceous of England and Belgium. In: Godefroit P, editor. Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Bloomington: Indiana University Press; p. 175–212.
- Norman DB. 2013. On the taxonomy and diversity of Wealden iguanodontian dinosaurs (Ornithischia: Ornithopoda). Revue de Paléobiologie. 32(2):385–404.
- Norman DB. 2015. On the history, osteology, and systematic position of the Wealden (Hastings group) dinosaur Hypselospinus fittoni (Iguanodontia: Styracosterna). Zool J Linn Soc. 173(1):92–189. doi:10.1111/zoj.12193.
- Norman DB, Weishampel DB. 2004. Basal Ornithopoda, in The Dinosauria. California: University of California Press; p. 395–412.
- Paul G. 2006. Horns and Beaks. Bloomington: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press; p. 69–77.
- Paul G. 2007. A revised taxonomy of the iguanodont dinosaur genera and species. Cretaceous Research. 29(2):192–216.
- Paul GS. 2008. A Revised taxonomy of the iguanodont dinosaur genera and species. Cretaceous Research. 29(2):192–216.
- Paul GS. 2012. Notes on the rising diversity of Iguanodont taxa, and Iguanodonts named after Darwin, Huxley, and evolutionary science. Actas de V Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Burgos. 123–133.
- Rey J. 1972. Recherches géologiques sur le Crétacé inférieur de l’Estremadura (Portugal). Lisbioa: SGP; p. 477. Memórias dos Serviços Geológicos de Portugal, 21
- Rey J. 1992. Les unités lithostratigraphiques du Crétacé inférieur de la région de Lisbonne. Comunicações dos Serviços Geológicos de Portugal. 78:103–124.
- Rey J, Graciansky PC, Jacquin TH. 2003. Les séquences de dépôt dans le Crétacé inférieur du Bassin Lusitanien. Comunicações do Instituto Geológico e Mineiro. 90:15–42.
- Rodrigues PC, Buscalioni ÁD, Santos VF, Fregenal-Martínez MA. 2008. Vertebrate microremains from the Lower Barremian at N of Cabo Espichel (Sesimbra, Portugal): historical review and preliminary results. In: Documents des Laboratoires de Géologie. Cognac (France). June 24th-28th 2008; p. 79–82.
- Ruiz-Omeñaca JI. 2011. Delapparentia turolensis nov. gen et sp., un nuevo dinosaurio iguanodontoideo (Ornithischia: Ornithopoda) en el Cretácico Inferior de Galve. Estudios geológicos. 67(1):83–110. doi:10.3989/egeol.40276.124.
- Ruiz-Omeñaca JI, Canudo JI. 2005. «Pleurocoelus» valdensis Lydekker, 1889 (Saurischia, Sauropoda) en el Cretácico inferior (Barremiense) de la Península Ibérica. Geogaceta. 38:43–46.
- Sánchez-Hernández B, Benton MJ, Naish D. 2007. Dinosaurs and other fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NE Spain. Palaeogeogr Palaeoclimatol Palaeoecol. 249(1–2):180–215. doi:10.1016/j.palaeo.2007.01.009.
- Sauvage H-E. 18971998. Vertébres fossils du Portugal. Contribuition à l’étude des Poisson et des Reptiles du Jurrassique et Crétacique. Direção dos Trabalhos Geológicos de Portugal, 98.
- Seeley HG 1888. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London, 43, 165–171; doi: 10.1098/rspl.1887.0117.
- Sereno PC. 1986. Phylogeny of the bird-hipped dinosaurs. Natl Geogr Res. 2:234–256.
- Sereno PC. 1997. The origin and evolution of dinosaurs. Annu Rev Earth Planet Sci. 25(1):435–489. doi:10.1146/annurev.earth.25.1.435.
- Sereno PC. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie. 210(1):41–83. doi:10.1127/njgpa/210/1998/41.
- Upchurch P, Mannion PD, Barrett PM. 2011. Sauropod dinosaurs. In: Batten DJ, editor. English Wealden Fossils. The Palaeontological Association Field Guide to Fossils Number 14. London: Wiley-Blackwell; p. 476–525.
- Verdú FJ, Royo-Torres R, Cobos A, Alcalá L. 2015. Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain). Cretaceous Research. 56:250e264. doi:10.1016/j.cretres.2015.05.010.
- Verdú FJ, Royo-Torres R, Cobos A, Alcalá L. 2017. New systematic and phylogenetic data about the early Barremian Iguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain. Hist Biol. doi:10.1080/08912963.2017.1287179
- Weishampel DB, Dodson P, Osmólska H. 2004. The Dinosauria. Berkeley: University of California Press.
- Weishampel DB, Heinrich RE. 1992. Systematics of hypsilophodontidae and basal Iguanodontia (dinosauria: Ornithopoda. Hist Biol. 6(3):159–184. doi:10.1080/10292389209380426.