
ABSTRACT
The Daniels collection of fossil birds from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK) contains multiple specimens of the Psittacopedidae and Zygodactylidae, which are here for the first time studied in detail. The Psittacopedidae include Parapsittacopes bergdahli, Psittacomimus eos, gen. et sp. nov., ?Psittacopes occidentalis, sp. nov., and at least one further unnamed species. The Zygodactylidae comprise Primozygodactylus cf. danielsi, Primoscens cf. minutus, Primoscens carolinae, sp. nov., as well as several unnamed species, which are represented by fragmentary remains. A very small zygodactylid-like species, which was previously identified as P. minutus, is described as Minutornis primoscenoides, gen. et sp. nov. A phylogenetic analysis showed the Zygodactylidae and Psittacopedidae to be stem group representatives of the Passeriformes (passerines), but the exact interrelationships of the fossil taxa are poorly resolved. In particular, no compelling character evidence supports a monophyletic Zygodactylidae, with distinctive features of these birds being likely to be plesiomorphic for Psittacopasseres or also found in crown group Passeriformes. In the majority rule consensus tree of our analysis, Minutornis is the sister taxon of a clade including the Zygodactylidae and Passeriformes, whereas the affinities of the Psittacopedidae were insufficiently resolvedhttp://zoobank.org/urn:lsid:zoobank.org:pub:81800349-5DE9-468B-81D7-6A5664526DF1.
Introduction
Current molecular analyses congruently support a clade termed Psittacopasseres (sensu Sangster et al. Citation2022), which includes the Psittaciformes (parrots) and Passeriformes (passerines). Whereas parrots are rather short-legged birds with zygodactyl feet (in which the fourth toe is reversed), passerines have long legs and anisodactyl feet (in which three toes direct forwards), and these two morphologically very different extant taxa were not considered sister taxa by earlier authors.
The recognition of Psittacopasseres had a major impact on the interpretation of some fossil taxa with zygodactyl feet, which are now considered to be zygodactyl stem group representatives of the Passeriformes (Mayr Citation2015; Ksepka et al. Citation2019, Citation2022). The best represented of these fossils belong to the Psittacopedidae and Zygodactylidae.
The Psittacopedidae includes rather short-legged birds with a parrot-like tarsometatarsus. The taxon was initially erected for Psittacopes lepidus from the latest early or earliest middle Eocene of Messel in Germany (Mayr and Daniels Citation1998). This species is known from two skeletons that show only few osteological details. However, Mayr and Daniels (Citation1998) also commented on similar birds from the early Eocene London Clay of Walton-on-the-Naze, which are represented by multiple three-dimensionally preserved bones. At this time, all of these fossils were in private collections, which prevented their formal description. One of the specimens, from the collection of the late Paul Bergdahl, was recently acquired by the Senckenberg Research Institute Frankfurt and was described as Parapsittacopes bergdahli (Mayr Citation2020). However, most of the London Clay fossils mentioned by Mayr and Daniels (Citation1998) were in the collection of Michael Daniels, which is now in the National Museums Scotland.
The Psittacopedidae were initially hypothesised to be stem group representatives of the Psittaciformes (Mayr and Daniels Citation1998), but in analyses by Mayr (Citation2015, Citation2020) the taxon Psittacopes was recovered at the base of a clade formed by the Zygodactylidae and Passeriformes (Mayr Citation2015). These analyses grouped Psittacopes with the Palaeogene taxa Morsoravis, Pumiliornis, and Eocuculus. Eocuculus was first described as a putative cuckoo (Cuculiformes) from the early Oligocene of North America (Chandler Citation1999), but tentative records exist from the early Oligocene of France (Mayr Citation2006, Citation2008a). The tiny, long-beaked Pumiliornis occurs in Messel (Mayr Citation1999, Citation2008a; Mayr and Wilde Citation2014), with a tentative record having been described from the early Eocene Nanjemoy Formation of Virginia, USA (Mayr et al. Citation2022). Morsoravis was initially established for species from the early Eocene Fur Formation of Denmark (Bertelli et al. Citation2010; Mayr Citation2011), but a putative record was also found in the North American Green River Formation (Grande Citation2013). In an analysis by Ksepka et al. (Citation2019), Psittacopes resulted in a clade that also included Pumiliornis, Eocuculus, Morsoravis, and Eofringillirostrum (the latter includes two species from the Green River Formation and Messel, respectively; Ksepka et al. Citation2019).
The taxon Zygodactylidae was established by Brodkorb (Citation1971) for Zygodactylus ignotus and Z. grivensis, which were then only known from fragmentary leg bones from the early Miocene of Germany and the middle Miocene of France, respectively (Ballmann Citation1969a; Citationb). Meanwhile, complete skeletons of zygodactylids were identified in various Eocene fossils sites of Europe and North America. These small and long-legged birds are particularly species-rich in Messel, where six species of the taxon Primozygodactylus have so far been reported (Mayr Citation1998; Mayr and Zelenkov Citation2009, Citation2017a). From the North American Green River Formation, two zygodactylid species were described as Eozygodactylus americanus and Zygodactylus grandei (Weidig Citation2010; Smith et al. Citation2018). Fragmentary remains of zygodactylids were also found in the early Eocene Nanjemoy Formation in Virginia (USA; Mayr et al. Citation2022). An initially misclassified species from the London Clay of the Isle of Sheppey (Essex, UK) was described as Primoscens minutus based on an incomplete carpometacarpus (Harrison and Walker Citation1977). Mayr (Citation2008b) reported a well-preserved skeleton of Zygodactylus from the early Oligocene of France, which was described as Z. luberonensis, and a species from the early Oligocene of North America was classified as Z. ochlurus (Hieronymus et al. Citation2019).
The affinities of Zygodactylus were considered unknown in the original description (contrary to the statement of Smith et al. Citation2018, p. 3; Ballmann Citation1969a; Citationb did not propose passeriform affinities, but discussed similarities to the Piciformes [woodpeckers and allies] and the Psittaciformes). Primoscens was assigned to the Passeriformes by Harrison and Walker (Citation1977). Mayr (Citation1998, Citation2004) hypothesised that zygodactylids (his ‘primoscenids’) belong to the Piciformes, which was also assumed by Simpson and Cracraft (Citation1981) for Zygodactylus. However, the recognition of a sister group relationship of the Psittaciformes and Passeriformes, as well as new data on the skeletal morphology of zygodactylids, led to the hypothesis that these fossil birds are in fact stem group representatives of the Passeriformes (Mayr Citation2008b, Citation2015).
Again, the London Clay of Walton-on-the-Naze yielded well-preserved zygodactylid fossils. Some of these specimens, from the Daniels collection, were briefly mentioned and figured by Mayr (Citation1998, Citation2009, Citation2022), but none has so far been described in detail. In the following, we revisit the Psittacopedidae and the Zygodactylidae from the Daniels collection; because both taxa are successive sister taxa of crown group Passeriformes in current analyses (Mayr Citation2015; Ksepka et al. Citation2019, Citation2020), they are treated together in the present study. In addition to the description of four new species, we report new material of previously described taxa and include all of the well-represented London Clay species of the Psittacopedidae and Zygodactylidae in a phylogenetic analysis.
Material and methods
The fossils are deposited in the Staatliche Naturwissenschaftliche Sammlungen Bayerns – Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany (SNSB-BSPG), the Denver Museum of Natural History, Colorado, USA (DM), the Geological Museum of the University of Copenhagen, Denmark (MGUH), the Natural History Museum, London, UK (NHMUK), the National Museums Scotland, Edinburgh, UK (NMS), the Senckenberg Research Institute Frankfurt, Germany (SMF), and the National Museum of Natural History, Smithsonian Institution, Washington D.C. (USNM).
A phylogenetic analysis was performed on the basis of the emended character matrix of Mayr (Citation2020; see Electronic Supplementary Material ESM1 and ESM2 for character descriptions and character matrix). The analysis was run with the heuristic search modus of NONA 2.0 (Goloboff Citation1993) through the WINCLADA 1.00.08 interface (Nixon Citation2002), using the commands hold 10,000, mult*1000, and hold/10. Bootstrap support values were calculated with 1000 replicates, ten searches holding ten trees per replicate, and TBR branch swapping without max*. The trees were rooted with the anseriform Anhimidae. Two characters were scored as additive. Tree length (L), consistency index (CI), and retention index (RI) were calculated.
Systematic palaeontology
Aves Linnaeus, 1758
Psittacopasseres sensu Sangster et al. (Citation2022)
Psittacopedidae Mayr, Citation2015
Included genera
Psittacopes Mayr and Daniels, Citation1998; Parapsittacopes Mayr, Citation2020; Psittacomimus, gen. nov.
Emended diagnosis
Characterised by dorsoventrally deep and short beak with large nostrils; humerus with well-delimited tuberculum supracondylare dorsale; short tarsometatarsus with large trochlea accessoria that is separated by a furrow from the trochlea metatarsi IV; sulcus between foramen vasculare distale and incisura intertrochlearis lateralis on the dorsal surface of the tarsometatarsus.
Psittacomimus, gen. nov.
Type species
Psittacomimus eos, sp. nov.
Differential diagnosis
Differs from Psittacopes in that the carpometacarpus has a curved os metacarpale minus and a wide spatium intermetacarpale, in the presence of a fossa between the processus pisiformis and the os metacarpale minus, and in that the tarsometatarsus has a proportionally wider shaft. Distinguished from Parapsittacopes in that the coracoid has a shorter processus procoracoideus, the scapula has a more pointed acromion, the distal end of the radius lacks a distoventrally protruding tuberculum aponeurosis ventralis, and in that the os metatarsale I has a proportionally longer processus articularis tarsometatarsalis.
Etymology
From μῖμος (mimos, Gr.): imitator and Psittacus, a genus of parrots, in reference to the similarity to psittaciform birds.
Psittacomimus eos, sp. nov.
Holotype
NMS.Z.2021.40.38 (); partial skeleton including skull, left quadrate, several vertebrae, left coracoid, proximal and distal ends of left humerus, fragmentary distal end of right humerus, proximal end of left ulna, right radius, right carpometacarpus, fragmentary distal portion of left carpometacarpus, pelvis, proximal end of right femur, right tibiotarsus, partial right tarsometatarsus, proximal end of left tarsometatarsus, pedal phalanges), collected in 1991 by M. Daniels (original collector’s number WN 91711).
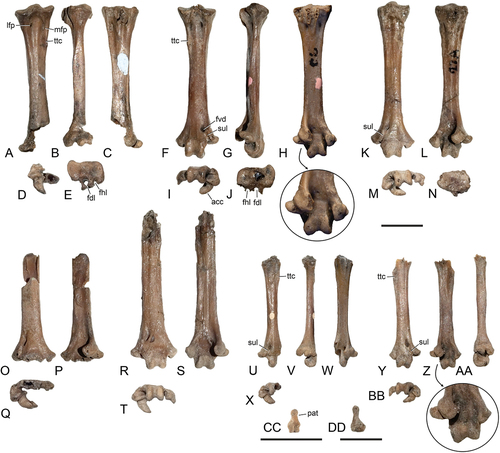
Figure 1. Specimens of Psittacomimus eos, gen. et sp. nov. from the London Clay of Walton-on-the-Naze (Essex, UK). (A) Holotype, NMS.Z.2021.40.38 (the two images on the left show two views of the block of matrix containing the skull). (B) NMS.Z.2021.40.39. (C) NMS.Z.2021.40.40. (D) Tentatively referred specimen NMS.Z.2021.40.41. (E) Tentatively referred specimen NMS.Z.2021.40.42. Abbreviations: ubk, upper beak; mdb, mandible; pel, pelvis; tbt, right tibiotarsus. The scale bar equals 5 mm.

Diagnosis
As for genus.
Etymology
The species epithet refers to the Eocene age of the fossils.
Type locality and horizon
Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation (previously Division A2; Rayner et al. Citation2009; Aldiss Citation2012); early Eocene (early Ypresian, 54.6‒55 million years ago; Collinson et al. Citation2016).
Referred specimens
NMS.Z.2021.40.39 (); partial skeleton including several vertebrae, furcula, both coracoids, both scapulae, proximal end of right humerus, partial pelvis, proximal portion of right femur, distal end of left tibiotarsus, left tarsometatarsus, distal end of right tarsometatarsus, several pedal phalanges), collected in 1985 by M. Daniels (original collector’s number WN 85506); NMS.Z.2021.40.40 (); right tarsometatarsus and pedal phalanx), collected in 1989 by M. Daniels (original collector’s number WN 89627A).
Tentatively referred specimens
NMS.Z.2021.40.41 (); distal portion of right tarsometatarsus lacking trochlea metatarsi III), collected in 1980 by M. Daniels (original collector’s number WN 80282A); NMS.Z.2021.40.42 (); right tarsometatarsus lacking proximal end and pedal phalanx), collected in 1990 by M. Daniels (original collector’s number WN 90642A).
Measurements (maximum length, in mm)
NMS.Z.2021.40.38: Skull, 35.0; left coracoid, 21.5; radius, 30.5; right carpometacarpus, 18.6; tibiotarsus length as preserved, 35.0, estimated total length, ~36; right tarsometatarsus length as preserved 21.7. NMS.Z.2021.40.39: Left tarsometatarsus, 19.1. NMS.Z.2021.40.40: Right tarsometatarsus, 18.6. NMS.Z.2021.40.42: Right tarsometatarsus length as preserved, 18.3.
Taxonomic remarks
The tarsometatarsus of NMS.Z.2021.40.42 lacks the proximal end, but appears to be longer than that of the holotype and may therefore belong to a different species. Because of this possible occurrence of two large psittacopedid species in Walton-on-the-Naze, the fragmentary specimen NMS.Z.2021.40.41 is likewise only tentatively referred to the new species.
Description and comparisons
The osteology of this species was already outlined by Mayr and Daniels (Citation1998; their ‘species A’), and some of the more general remarks are not repeated here. The holotype includes a nearly complete skull, which is still partially embedded in matrix (). In its overall proportions it resembles the skull of Parapsittacopes bergdahli, and as in the latter species the internarial bar of the upper beak widens caudally. Also as in P. bergdahli (), the caudal nasal bar of the beak is situated in a rostral position. As a consequence, there is an unusually large distance between the caudal nasal bar and the rostral end of the neurocranium. The nostril is very large and has an angled rather than a rounded caudodorsal portion; in its relative size and shape, it long resembles the nostrils of the charadriiform Thinocoridae, whereas the nostrils of most extant arboreal land birds are proportionally smaller and have a more rounded caudal portion. The rostrum maxillae is very short. The septum interorbitale is extensively ossified and the fossa temporalis moderately deep, but other morphologically informative details of the neurocranium (of which mainly the dorsolateral surface is exposed in the fossil) cannot be discerned.
Figure 2. Skull, selected vertebrae, and pelvis of the Psittacopedidae from Walton-on-the-Naze (Essex, UK). (A, B) Skull of Psittacomimus eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38). (C) Fragmentary mandible of Ps. eos (holotype, NMS.Z.2021.40.38). (D) Skull of Parapsittacopes bergdahli (holotype, SMF Av 653). (E‒H) left quadrate of Ps. eos (holotype, NMS.Z.2021.40.38) in (E) lateral, (F) caudal, (G) medial, and (H) ventral view. (I) Atlas of Ps. eos (holotype, NMS.Z.2021.40.38). (J, K) Axis (cranial view) of (J) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44) and (K) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46). (L, M) Third cervical vertebra (dorsal view) of (L) Pa. bergdahli (holotype, SMF Av 653) and (M) Ps. eos (holotype, NMS.Z.2021.40.38). (N) Thoracic vertebra of Ps. eos (NMS.Z.2021.40.39). (O, P) Pelvis (left lateral view) of Ps. eos (holotype, NMS.Z.2021.40.38), in (P) the surrounding matrix was digitally removed. Abbreviations: cdl, condylus lateralis; cpo, capitulum oticum; cps, capitulum squamosum; cnb, caudal nasal bar; fac, foramen acetabuli; fii, foramen ilioischiadicum; icf, incisura fossae; inb, internarial bar; nos, nostril; ntc, notch; pac, processus articularis caudalis; plc, pleurocoel; pnf, pneumatic foramina; psp, processus spinosus; pvt, processus ventralis; scp, caudal portion of scapula; tbt, right tibiotarsus; zgc, zygapophysis cranialis. The scale bars equal 5 mm.

As far as comparisons are possible, the quadratum () corresponds well to that of P. bergdahli. As in the latter species, the tip of the processus oticus exhibits a large pneumatic opening on its caudal surface. The condylus pterygoideus is broken.
A fragment of a mandibular ramus is situated next to the skull. In contrast to Psittacopes, it does not exhibit a large fenestra mandibulae ()).
The holotype and the referred specimen NMS.Z.2021.40.39 include several vertebrae. The atlas exhibits an open incisura fossae and has a well-developed processus ventralis ()). The axis is craniocaudally long and has a long dens. The vertebra here identified as the third thoracic one has a longer zygapophysis cranialis than the corresponding vertebra of P. bergdahli; there is a small notch on the left side of the corpus, which is absent on the right side ()). As in Parapsittacopes, the thoracic vertebrae ()) are pleurocoelous, that is, they bear deep fossae on the lateral surfaces of the body (Mayr Citation2021).
The coracoid () is distinguished from that of P. bergdahli () in that the processus procoracoideus is distinctly shorter. Unlike in P. bergdahli, the shaft of the bone exhibits a shallow incisura nervi supracoracoidei (NMS.Z.2021.40.39; )).
Figure 3. Pectoral girdle and wing bones of the Psittacopedidae from Walton-on-the-Naze (Essex, UK). (A‒C) Psittacomimus eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), left coracoid in (A) dorsal, (B) medial, and (C) ventral view. (D) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), left coracoid in dorsal view. (E‒G) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44) (holotype, NMS.Z.2021.40.38), left coracoid in (E) dorsal, (F) medial, and (G) ventral view. (H, I) Parapsittacopes bergdahli (NMS.Z.2021.40.43), left coracoid in (H) dorsal and (I) ventral view. (J, K) Pa. bergdahli (holotype, SMF Av 653), right coracoid in (J) ventral and (K) dorsal view. (L‒N) Right scapula (Medial view) of (L) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), (M) Pa. bergdahli (holotype, SMF Av 653), and (N) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44). (O, P) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), proximal end of left humerus in (O) caudal and (P) cranial view. (Q, R) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), distal ends of (Q) right and (R) left humerus in cranial view. (S) Pa. bergdahli (holotype, SMF Av 653), distal end of left humerus in cranial view. (T, U) Pa. bergdahli (NMS.Z.2021.40.43), left humerus in (T) cranial and (U) caudal view. (V, W) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), proximal end of left humerus (V: caudal view) and distal end of right humerus (W: cranial view). (X) Pa. bergdahli (holotype, SMF Av 653), proximal end of left ulna in cranial view. (Y) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), proximal end of left ulna in cranial view. (Z‒BB) distal ends of (Z) right and (AA, BB) left ulna (ventral view) of (Z, AA) Pa. bergdahli (Z: NMS.Z.2021.40.43; AA: holotype, SMF Av 653) and (BB) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44). (CC, DD) Pa. bergdahli, distal end of right radius in ventral view (CC: holotype, SMF Av 653 [erroneously considered to be from the left side by Mayr 2002: fig. 5Q]; DD: NMS.Z.2021.40.43). (EE) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), distal end of right radius in ventral view. (FF) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46), distal end of left radius in ventral view. (GG) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), distal end of right radius in ventral view. (HH, II) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), right carpometacarpus in (HH) ventral and (II) dorsal view. (JJ, KK) Pa. bergdahli (holotype, SMF Av 653), proximal end of left carpometacarpus in (JJ) ventral and (KK) dorsal view. (LL, MM) Pa. bergdahli (NMS.Z.2021.40.43), proximal portion of left carpometacarpus in (LL) ventral and (MM) dorsal view. (NN, OO) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), right carpometacarpus in (NN) ventral and (OO) dorsal view. Abbreviations: acr, acromion; cdd, condylus dorsalis; ctd, cotyla dorsalis; ctv, cotyla ventralis; fos, fossa between processus pisiformis and os metacarpale minus; hkp, hook-like process; ins, incisura nervi supracoracoidei; pim, processus intermetacarpalis; pis, processus pisiformis; ppc, processus procoracoideus; tav, tuberculum aponeurosis ventralis; tbc, tuberculum carpale; tbd, tuberculum dorsale; tsd, tuberculum supracondylare dorsale. The scale bar equals 5 mm.
![Figure 3. Pectoral girdle and wing bones of the Psittacopedidae from Walton-on-the-Naze (Essex, UK). (A‒C) Psittacomimus eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), left coracoid in (A) dorsal, (B) medial, and (C) ventral view. (D) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), left coracoid in dorsal view. (E‒G) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44) (holotype, NMS.Z.2021.40.38), left coracoid in (E) dorsal, (F) medial, and (G) ventral view. (H, I) Parapsittacopes bergdahli (NMS.Z.2021.40.43), left coracoid in (H) dorsal and (I) ventral view. (J, K) Pa. bergdahli (holotype, SMF Av 653), right coracoid in (J) ventral and (K) dorsal view. (L‒N) Right scapula (Medial view) of (L) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), (M) Pa. bergdahli (holotype, SMF Av 653), and (N) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44). (O, P) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), proximal end of left humerus in (O) caudal and (P) cranial view. (Q, R) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), distal ends of (Q) right and (R) left humerus in cranial view. (S) Pa. bergdahli (holotype, SMF Av 653), distal end of left humerus in cranial view. (T, U) Pa. bergdahli (NMS.Z.2021.40.43), left humerus in (T) cranial and (U) caudal view. (V, W) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), proximal end of left humerus (V: caudal view) and distal end of right humerus (W: cranial view). (X) Pa. bergdahli (holotype, SMF Av 653), proximal end of left ulna in cranial view. (Y) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), proximal end of left ulna in cranial view. (Z‒BB) distal ends of (Z) right and (AA, BB) left ulna (ventral view) of (Z, AA) Pa. bergdahli (Z: NMS.Z.2021.40.43; AA: holotype, SMF Av 653) and (BB) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44). (CC, DD) Pa. bergdahli, distal end of right radius in ventral view (CC: holotype, SMF Av 653 [erroneously considered to be from the left side by Mayr 2002: fig. 5Q]; DD: NMS.Z.2021.40.43). (EE) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), distal end of right radius in ventral view. (FF) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46), distal end of left radius in ventral view. (GG) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), distal end of right radius in ventral view. (HH, II) ?P. occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), right carpometacarpus in (HH) ventral and (II) dorsal view. (JJ, KK) Pa. bergdahli (holotype, SMF Av 653), proximal end of left carpometacarpus in (JJ) ventral and (KK) dorsal view. (LL, MM) Pa. bergdahli (NMS.Z.2021.40.43), proximal portion of left carpometacarpus in (LL) ventral and (MM) dorsal view. (NN, OO) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), right carpometacarpus in (NN) ventral and (OO) dorsal view. Abbreviations: acr, acromion; cdd, condylus dorsalis; ctd, cotyla dorsalis; ctv, cotyla ventralis; fos, fossa between processus pisiformis and os metacarpale minus; hkp, hook-like process; ins, incisura nervi supracoracoidei; pim, processus intermetacarpalis; pis, processus pisiformis; ppc, processus procoracoideus; tav, tuberculum aponeurosis ventralis; tbc, tuberculum carpale; tbd, tuberculum dorsale; tsd, tuberculum supracondylare dorsale. The scale bar equals 5 mm.](/cms/asset/afda8e32-feb9-40d7-a5b3-bfcc3e6794ea/ghbi_a_2141629_f0003_oc.jpg)
The scapula is long and slender, with a long and pointed acromion ()). The U-shaped furcula is well preserved in specimen NMS.Z.2021.40.39 ()). The extremitas omalis forms a long processus acromialis and bears a small facies articularis acrocoracoidea. The extremitas sternalis exhibits a small apophysis furculae.
Only fragments of the humerus are present in the specimens (). The proximal end has a small tuberculum dorsale. There are no pneumatic openings at the bottom of the fossa pneumotricipitalis. The distal end of the bone bears a small but well-defined tuberculum supracondylare dorsale; as in P. bergdahli this tubercle is continuous with a transverse attachment scar for the musculus extensor carpi radialis on the cranial surface of the bone (this scar is more pronounced on the distal end of the left humerus of the holotype).
The ulna is likewise poorly preserved in the fossils. The proximal end ()) corresponds well with the proximal ulna of P. bergdahli ()), even though the long cotyla dorsalis has a more rounded proximal margin. As in P. bergdahli, an oblique ridge extends distally from the cotyla dorsalis. The fragmentary distal end is preserved in NMS.Z.2021.40.39.
The distal end of the radius forms a small, hook-like process ((GG)), which is somewhat smaller than in Parapsittacopes ((CC), (DD)); unlike in the latter, the tuberculum aponeurosis ventralis is not strongly developed and distoventrally protruding.
In contrast to Psittacopes, the carpometacarpus ((NN), (OO)) has a bowed os metacarpale minus, so that the spatium intermetacarpale is much wider. Unlike in Psittacopes, there is furthermore a fossa at the proximal end of the os metacarpale minus; the cranial margin of this fossa is bordered by a ridge from the processus pisiformis to the os metacarpale minus. The os metacarpale majus forms a very small processus intermetacarpalis.
The pelvis of the holotype is situated on the block of matrix, which contains the skull (). Even though a significant portion of the left side is present, its preservation does not allow the recognition of many osteological details. The exposed parts show a close resemblance to the pelvis of P. bergdahli.
As far as comparisons are possible, the femur resembles that of P. bergdahli. The crista trochanteris is poorly developed and there is a large fossa, which extends over much of the caudal surface of the proximal end.
The long tibiotarsus of the holotype is attached to the block of matrix that also includes the skull (); its proximal end is broken. Both condyles of the distal end have an equal proximodistal depth, but the condylus medialis is mediolaterally much narrower than the condylus lateralis. The sulcus extensorius is bridged by a pons supratendineus.
The tarsometatarsus is almost complete in the referred specimen NMS.Z.2021.40.39 (). The hypotarsus bears a nearly closed canal for the tendon of musculus flexor digitorum longus and a sulcus for that of musculus flexor hallucis longus. The foramina vascularia proximalia are widely spaced and are similar in size (in the holotype, the medial foramen is somewhat larger than the lateral one and situated slightly farther distally). The tuberositas musculi tibialis cranialis has an ovate outline and is located near the medial margin of the tarsometatarsus. The foramen vasculare distale is fairly large. On the dorsal surface of the bone there is a sulcus between the foramen vasculare distale and the incisura intertrochlearis lateralis, which represents the dorsally open canalis interosseus distalis. The trochlea metatarsi II forms a plantar projection. The trochlea metatarsi III is mediolaterally very wide and has a somewhat asymmetric outline, with the lateral trochlear rim protruding slightly farther distally than the medial one; the trochlear furrow is moderately deep. The trochlea accessoria of the trochlea metatarsi IV is well developed and separated from the main trochlea by a furrow.
Figure 4. Leg bones of the Psittacopedidae from Walton-on-the-Naze (Essex, UK). (A‒E) Psittacomimus eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38), partial right tarsometatarsus in (A) dorsal, (B) lateral, (C) plantar, (D) distal, and (E) proximal view. (F‒J) Ps. eos (NMS.Z.2021.40.39), left tarsometatarsus in (F) dorsal, (G) lateral, (H) plantar, (I) distal, and (J) proximal view; the arrow indicates an enlarged detail of the distal end. (K‒N) Ps. eos (NMS.Z.2021.40.40), right tarsometatarsus in (K) dorsal, (L) plantar, (M) distal, and (N) proximal view. (O‒Q) Ps. eos, tentatively referred specimen NMS.Z.2021.40.41, distal portion of right tarsometatarsus in (O) dorsal, (P) plantar, and (Q) distal view. (R‒T) Ps. eos, tentatively referred specimen NMS.Z.2021.40.42, partial right tarsometatarsus in (R) dorsal, (S) plantar, and (T) distal view. (U‒X) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), partial right tarsometatarsus in (U) dorsal, (V) lateral, (W) plantar, and (X) distal view. (Y‒BB) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46), left tarsometatarsus (Y) dorsal, (Z) plantar, (AA) lateral, and (BB) distal view; the arrow indicates an enlarged detail of the distal end. (CC, DD) Os metatarsale I of Parapsittacopes bergdahli (CC: holotype, SMF Av 653) and Ps. eos (DD: holotype, NMS.Z.2021.40.38). Abbreviations: acc, trochlea accessoria; fdl, hypotarsal sulcus/canal for tendon of musculus flexor digitorum longus; fhl, hypotarsal sulcus/canal for tendon of musculus flexor hallucis longus; fvd, foramen vasculare distale; lfp, lateral foramen vasculare proximale; mfp, medial foramen vasculare proximale; pat, processus articularis tarsometatarsalis; sul, sulcus between foramen vasculare distale and incisura intertrochlearis lateralis; ttc, tuberositas musculi tibialis cranialis. The scale bars equal 5 mm.
The os metatarsale I ()) has a very slender processus articularis tarsometatarsalis; with regard to this feature it is distinguished from the corresponding ossicle of P. bergdahli ()). Several pedal phalanges are preserved in the holotype and specimen NMS.Z.2021.40.39, and these have similar proportions to the phalanges of P. bergdahli. As in P. bergdahli, the tuberculum flexorium of the ungual phalanges is very poorly developed.
Parapsittacopes bergdahli Mayr, Citation2020
Referred specimen
NMS.Z.2021.40.43 (); left coracoid, caudal portion of scapula, left humerus, proximal end of right ulna, distal ends of both ulnae, distal end of right radius, proximal end of right carpometacarpus, os carpi ulnare, phalanx proximalis digiti majoris), collected in 1994 by M. Daniels (original collector’s number WN 94811).
Figure 5. Specimens of small species of the Psittacopedidae from the London Clay of Walton-on-the-Naze (Essex, UK). (A) Parapsittacopes bergdahli Mayr, Citation2020 (NMS.Z.2021.40.43). (B) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46). (C) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44). The scale bar equals 5 mm.

Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Left coracoid, 16.9; left humerus, 21.9.
Remarks
The holotype of Parapsittacopes bergdahli is a partial skeleton from Walton-on-the-Naze, which was described by Mayr (Citation2020). NMS.Z.2021.40.43 is assigned to this species, because the distal end of the radius exhibits a hook-like process. Except for the os carpi ulnare, the new specimen does not include skeletal elements that are not preserved in the holotype. In the new specimens, the coracoid and humerus are entire and allow reliable length measurements, whereas both bones are incomplete in the holotype. The carpometacarpus exhibits a small processus intermetacarpalis of similar size to that of Psittacomimus eos. As in the latter species, the proximal end of the os metacarpale minus meets the carpometacarpus at a steeper angle than in Psittacopes, which indicates a wider spatium intermetacarpale.
?Psittacopes Mayr and Daniels, Citation1998
?Psittacopes occidentalis, sp. nov.
Holotype
NMS.Z.2021.40.44 (); partial skeleton including several vertebrae, furcula, right scapula, left and partial right coracoids, proximal end of left humerus and distal end of right humerus, distal portion of left ulna, partial radii, both carpometacarpi, proximal end of left femur, partial right tarsometatarsus, pedal phalanges), collected in 1986 by M. Daniels (original collector’s number WN 86540).
Differential diagnosis
Distinguished from Psittacopes lepidus in a proportionally longer carpometacarpus (carpometacarpus and tarsometatarsus measuring 13.2 and 13.6 mm, respectively, versus 10.4 and ~13.3 mm in P. lepidus; Mayr and Daniels Citation1998). Differs from Parapsittacopes bergdahli in that the scapula is shorter (), the coracoid has a smaller processus acrocoracoideus, the distal end of the radius does not form a hook-like projection ()), and the carpometacarpus lacks a processus intermetacarpalis and a ridge from the processus pisiformis to the os metacarpale minus. Differs from Psittacomimus eos in a much smaller size and a straight os metacarpale minus.
Etymology
From occidentalis (Lat.): Western, in reference to the geographical location of the London Clay to the west of Messel in Germany (which is the type locality of Psittacopes lepidus).
Type locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Left coracoid, 16.8; carpometacarpus, 13.2 (left), 13.2 (right); phalanx proximalis digiti majoris, 6.5; phalanx distalis digiti majoris, 5.3; tarsometatarsus, length as preserved, 13.6.
Remarks
This is ‘species B’ of Mayr and Daniels (Citation1998).
Description and comparisons
The coracoid closely resembles that of Parapsittacopes bergdahli in its overall shape, but the facies articularis is proportionally smaller than in the latter species. As in P. bergdahli, the processus procoracoideus is very long. The scapula is shorter than in P. bergdahli. The extremitates omales of the furcula are proportionally narrower than in Psittacomimus eos.
The humerus ()) corresponds well to that of P. bergdahli in the morphology of the proximal and distal ends. Only the distal portion of the ulna is preserved in the holotype, which resembles the distal ulna of P. bergdahli. Unlike in P. bergdahli, the distal end of the radius does not form a hook-like projection and the tuberculum aponeurosis ventralis is not strongly developed and distoventrally protruding.
As in Psittacopes lepidus, but unlike in Psittacomimus eos, the carpometacarpus has a straight os metacarpale minus and a narrow spatium intermetacarpale (as detailed above, the spatium intermetacarpale also appears to have been wider in Parapsittacopes). Furthermore, there is no fossa between the processus pisiformis and the os metacarpale minus. In contrast to Psittacomimus and Parapsittacopes, a processus intermetacarpalis is absent.
The proximal end of the femur resembles that of P. bergdahli. The shaft of the tarsometatarsus is more slender than in Psittacomimus eos. As in the latter species, there is a sulcus on the dorsal surface of the distal tarsometatarsus, between the foramen vasculare distale and the incisura intertrochlearis lateralis. The trochlea accessoria of the trochlea metatarsi IV closely resembles that of Psittacomimus and Parapsittacopes. The pedal phalanges correspond to those of P. bergdahli in their proportions.
Psittacopedidae, gen. et sp. indet. A
Referred specimen
NMS.Z.2021.40.45 (distal end of right tibiotarsus, proximal portion of right tarsometatarsus lacking distal end), collected in 1996 by M. Daniels (original collector’s number WN 96926).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Remarks
This fragmentary specimen was figured by Mayr and Daniels (Citation1998: text-figs. 5J and 7B, D). Unlike in Psittacomimus eos, the hypotarsus exhibits a closed canal for the tendon of musculus flexor hallucis longus (the tendon of musculus flexor digitorum longus was situated in a sulcus). Because the hypotarsus of Parapsittacopes bergdahli is unknown, there exists a possibility that NMS.Z.2021.40.45 belongs to this species. However, the distal end of the tibiotarsus and the proximal tarsometatarsus are also unknown from the similar-sized () ?Psittacopes occidentalis, sp. nov. and the unnamed species mentioned below, so that a definitive identification of NMS.Z.2021.40.45 is not possible.
Table 1. Length measurements (in mm) of major limb bones of the Psittacopedidae and Zygodactylidae (from Mayr Citation1998; Citation2017a, Citation2020; Mayr and Daniels Citation1998; Mayr and Zelenkov Citation2009; Smith et al. Citation2018; this study). Species in bold are from the London Clay of Walton-on-the-Naze.
Psittacopedidae, gen. et sp. indet. B
Referred specimen
NMS.Z.2021.40.46 (); partial skeleton including jugal bar, a few vertebrae, both scapulae, proximal portion of right humerus, distal end of left radius, proximal and distal portions of left carpometacarpus, fragmentary synsacrum, and left tarsometatarsus lacking proximal end), collected in 1996 by M. Daniels (original collector’s number WN 96949).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (in mm)
Tarsometatarsus, length as preserved, 13.4.
Description and comparisons
This specimen represents ‘species C’ of Mayr and Daniels (Citation1998). Unlike in Parapsittacopes bergdahli, the distal end of the radius does not form a hook-like process and the tuberculum aponeurosis ventralis is not strongly developed and distoventrally protruding ()). NMS.Z.2021.40.46 differs from the holotype of ?Psittacopes occidentalis, sp. nov. in that the axis is proportionally larger, with a longer and more strongly protruding processus articularis caudalis ()), and the tarsometatarsus has a somewhat wider shaft ()). As in the much larger Psittacomimus eos, there is a sulcus on the dorsal surface of the distal tarsometatarsus, between the foramen vasculare distale and the incisura intertrochlearis lateralis. The trochlea metatarsi II is proportionally smaller than in P. eos. Even though NMS.Z.2021.40.46 is likely to represent a new species, we considered it inappropriate to formally name it owing to the fragmentary preservation of the only known specimen.
Parapasseres Mayr, Citation2015
Zygodactylidae Ballmann, Citation1969a
Remarks
The species of the Zygodactylidae exhibit a distinctive morphology, which distinguishes them from other Palaeogene birds and all extant avian taxa. Shared features include a long tarsometatarsus with a characteristic morphology of the distal end and a carpometacarpus with a well-developed processus intermetacarpalis. However, some of the shared similarities are likely to be plesiomorphic for the Psittacopasseres, and this is particularly true for the morphology of the distal end of the tarsometatarsus, which closely resembles the distal tarsometatarsus of the Psittacopedidae and crown group Psittaciformes (Citation2015).
Primozygodactylus Mayr, Citation1998
Emended diagnosis
Spina externa of sternum blade-like; extremitas omalis of furcula widened into a subtriangular expansion; humerus with dorsal margin of distal end bearing a marked, edge-like prominence; crista bicipitalis (humerus) forming a distinct convexity; cotyla dorsalis of ulna reaching farther distally than cotyla ventralis; femur longer than humerus; hypotarsus enclosing two canals for the tendons of musculus flexor digitorum longus and musculus flexor hallucis longus; tarsometatarsus shaft with crista plantaris lateralis; trochlea metatarsi II deflected plantarly and forming a plantar projection; medial margin of accessory trochlea of trochlea metatarsi IV (‘Sehnenhalter’ sensu Ballmann Citation1969a; Citationb; Mayr Citation1998) not reaching midline of trochlea metatarsi III.
Primozygodactylus cf. danielsi Mayr, Citation1998
Referred specimens
NMS.Z.2021.40.47 (); partial skeleton including rostral portion of mandible, partial furcula, extremitas omalis of left coracoid, both humeri, left ulna, fragments of both radii, both carpometacarpi, phalanges proximales digiti majoris; fragments of pelvis, right femur, left tibiotarsus, complete right tarsometatarsus, partial left tarsometatarsus, and a few pedal phalanges), collected in 1988 by M. Daniels (original collector’s number WN 88583); NMS.Z.2021.40.48 (); partial skeleton including right scapula, extremitas sternalis of furcula, cranial portion of sternum, right humerus, left ulna, proximal end of left carpometacarpus, distal portion of left tarsometatarsus, and a few pedal phalanges), collected in 1989 by M. Daniels (original collector’s number WN 89609); NMS.Z.2021.40.49 (); right humerus, right scapula, distal end of right ulna, distal portion of right radius, and fragmentary proximal and distal portions of right carpometacarpus), collected in 1983 by M. Daniels (original collector’s number WN 83477); NMS.2021.40.50 (); proximal and distal ends of left tarsometatarsus), collected in 1981 by M. Daniels (original collector’s number WN 81317); NMS.2021.40.51 (); both quadrates, distal end of left ulna, proximal portion of left carpometacarpus, phalanx proximalis digiti majoris, proximal portion of left femur, and other bone fragments), collected in 1980 by M. Daniels (original collector’s number WN 80282B); SMF Av 660 (distal portion of right tarsometatarsus and pedal phalanx), collected in 1981 by P. Bergdahl (original collector’s number BC 8102).
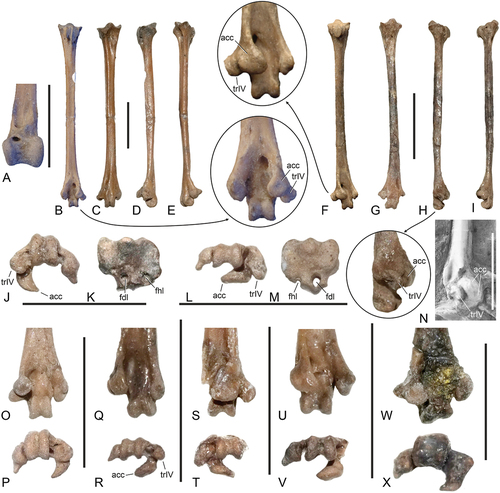
Figure 6. Zygodactylid fossils from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). Major bones are shown to illustrate the different sizes of the species and the completeness of the specimens, which are separated by dashed lines. (A) Primozygodactylus cf. danielsi (NMS.2021.40.47). (B) P. cf. danielsi (NMS.2021.40.48; cranial portion of sternum not shown). (C) P. cf. danielsi (NMS.2021.40.49). (D) P. cf. danielsi (NMS.2021.40.50). (E) P. cf. danielsi (NMS.2021.40.51). (F) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54). (G) Pr. carolinae, sp. nov. (NMS.2021.40.55). (H) Pr. carolinae, sp. nov. (NMS.2021.40.56). (I) cf. Pr. carolinae, sp. nov. (NMS.2021.40.57). (J) Primoscens sp. A (NMS.2021.40.58). (K) Primoscens sp. B (NMS.2021.40.59). (L) Primoscens cf. minutus (NMS.2021.40.53). (M) Zygodactylidae, gen. et sp. indet. A (NMS.2021.40.60). (N) Zygodactylidae, gen. et sp. indet. B (NMS.2021.40.61). (O) Minutornis primoscenoides, gen. et sp. nov. (holotype, NMS.2021.40.62); the sternum is not figured. The scale bar equals 5 mm.

Figure 7. Cranial elements of the Zygodactylidae from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). (A, B) Primozygodactylus cf. danielsi (NMS.2021.40.47), rostral portion of mandible in (A) dorsal and (B) left lateral view. (C, D) Zygodactylidae, gen. et sp. indet. B (NMS.2021.40.61), rostral portion of upper beak in (C) dorsal and (D) left lateral view. (E‒L) P. cf. danielsi (NMS.2021.40.51), (E‒H) left and (I‒L) right quadrate in (E, I) lateral, (F, J) caudal, (G, K) medial, and (H, L) ventral view. (M‒P) Zygodactylidae, gen. et sp. indet. B (NMS.2021.40.61), right quadrate in (M) lateral, (N) caudal, (O) medial, and (P) ventral view. Abbreviations: cdc, condylus caudalis; cdl, condylus lateralis; cdm, condylus medialis; cdp, condylus pterygoideus; cpo, capitulum oticum; cps, capitulum squamosum; fos, fossa; pnf, pneumatic foramina; tsc, tuberculum subcapitulare. The scale bars equal 5 mm.

Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
NMS.2021.40.47: Right humerus, 16.6; left humerus, 16.7; left ulna, 18.4; right carpometacarpus, 9.5; left carpometacarpus, 9.5; right femur, 17.8; left tibiotarsus, length as preserved, 27.2; right tarsometatarsus, 20.0. NMS.2021.40.48: Right Humerus, 15.4; left ulna, 17.9. NMS.2021.40.49: Right humerus, 15.8.
Remarks
Primozygodactylus danielsi was described by Mayr (Citation1998) based on compression fossils from Messel in Germany. The specimens from Walton-on-the-Naze are about 5 million years older than those from Messel, so that it is likely that they belong to a different species. However, currently it is not possible to unambiguously differentiate them from P. danielsi, with which the fossils from Walton-on-the-Naze correspond well in their bone dimensions (; the slightly longer carpometacarpus may be due to the fact that the exact length of this bone is difficult to measure in the articulated skeletons from Messel).
Description and comparisons
The skeletal morphology of the taxon Primozygodactylus was described in detail in previous studies (Mayr Citation1998; Mayr and Zelenkov Citation2009, Citation2017a), and the following description mainly focuses on features that are not visible in the skeletons from Messel. If not indicated otherwise, the description is based on specimen NMS.2021.40.47.
The quadrate (; NMS.2021.40.51) closely resembles that of Parapsittacopes bergdahli. The bone exhibits a large pneumatic opening on the caudal surface of the processus oticus. The processus oticus itself has a broad tip, with widely spaced capitula and a shallow incisura intercapitularis. A small tuberculum subcapitulare is situated ventral to the capitulum squamosum. There is a distinct fossa on the caudal surface of the processus oticus. The dorsal portion of the tip of the condylus lateralis forms a lip-like projection. The condylus pterygoideus is well-defined and narrow. The condylus caudalis shows little ventral prominence.
The mandible () is long and narrow, with a moderately long symphysis. In its proportions it is similar to the mandible of Acrocephalus warblers (Passeriformes, Sylviidae).
The extremitas omalis of the coracoid is fairly straight and has a rounded tip (). It is similar in its shape to the extremitas omalis of Zygodactylus (), whereas the coracoid of Primoscens and Primozygodactylus has a hook-like extremitas omalis. The facies articularis scapularis is shallow. The processus procoracoideus is broken in NMS.2021.40.47, so that its shape cannot be determined. This process was considered to be greatly reduced in the Messel specimens of P. danielsi (Mayr Citation1998, Citation2017a).
Figure 8. Pectoral girdle bones of the Zygodactylidae from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). (A, B) Primozygodactylus cf. danielsi (NMS.2021.40.47), extremitas omalis of left coracoid in (A) dorsal and (B) ventral view. (C, D) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54), left coracoid in (C) dorsal and (D) ventral view. (E, F) Minutornis primoscenoides, gen. et sp. nov. (holotype, NMS.2021.40.62), left coracoid in (E) dorsal and (F) ventral view. (G) Zygodactylidae, gen. et sp. indet. B (NMS.2021.40.61), left coracoid in dorsal view. (H) Zygodactylus luberonensis from the early Oligocene of France (SMF Av 519), right coracoid in dorsal view; coated with ammonium chloride. (I) P. cf. danielsi (NMS.2021.40.48), right scapula in lateral view. (J) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), partial right scapula in lateral view. (K, L) M. primoscenoides (holotype, NMS.2021.40.62), (K) extremitas sternalis and (L) scapus of furcula. (M) P. cf. danielsi (NMS.2021.40.47), scapus of furcula. (N, O) P. cf. danielsi (NMS.2021.40.48), extremitas sternalis of furcula in (N) cranial and (O) lateral view. (P, Q) Zygodactylidae, gen. et sp. indet. A (NMS.2021.40.60), (P) extremitas sternalis and (Q) scapus of furcula. (R, S) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), cranial portion of sternum in (R) ventral and (S) right lateral view. (T‒V) P. cf. danielsi (NMS.Z.2021.40.48), cranial portion of sternum in (T) right lateral, (U) dorsal, and (V) ventral view. (W‒Y) M. primoscenoides (holotype, NMS.2021.40.62), sternum in (W) left lateral, (X) dorsal, and (Y) ventrolateral view. Abbreviations: acr, acromion; apf, apophysis furculae; exo, extremitas omalis; mpr, medial projection of extremitas sternalis; pac, processus acrocoracoideus; ppc, processus procoracoideus; spe, spina externa. The scale bars equal 5 mm.

The acromion of the scapula ()) is long and slightly bipartite. The corpus of the bone is narrow and only slightly angled at its caudal end.
The furcula () has a widened extremitas omalis of subtriangular shape, which is wider than in Zygodactylus ()). The apophysis furculae is long and blade-like (NMS.2021.40.47; ).
The cranial portion of the sternum is preserved in NMS.Z.2021.40.48 (). As in the Messel specimens of P. danielsi, the spina externa is blade-like.
The humerus () corresponds to that of the Primozygodactylus specimens from Messel. As in the latter, the crista bicipitalis forms a small bulge. The fossa pneumotricipitalis lacks pneumatic openings. The tuberculum dorsale is very small and proportionally smaller than in Primoscens. On the distal end of the bone, there is a marked, edge-like dorsal prominence (NMS.2021.40.49). A transverse ridge on the cranial surface of this projection serves for the attachment of musculus extensor carpi radialis. The processus flexorius is moderately developed and strongly distally projecting.
Figure 9. Wing bones of the Zygodactylidae from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). (A) Primozygodactylus cf. danielsi (NMS.2021.40.49), right humerus in cranial view; the arrow indicates an enlarged detail of the distal end. (B, C) P. cf. danielsi (NMS.2021.40.47), (B) right and (C) left humerus in (B) caudal and (C) cranial view. (D‒F) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54), right humerus in (D) cranial and (E) caudal view, (F) left humerus in cranial view; the arrow indicates an enlarged detail of the distal end. (G, H) Minutornis primoscenoides, gen. et sp. nov. (holotype, NMS.2021.40.62), (G) left humerus in cranial view, (H) right humerus in caudal view; the arrow indicates an enlarged detail of the distal end. (I, J) P. cf. danielsi (NMS.2021.40.47), left ulna in (I) cranial and (J) ventral view; the arrow indicates an enlarged detail of the proximal end. (K, L) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), left ulna in (K) cranial and (L) ventral view; the arrow indicates an enlarged detail of the proximal end. (M) Pr. cf. minutus (NMS.2021.40.47), proximal end of right ulna in cranial view; the arrow indicates an enlarged detail. (N, O) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), right (N: ventral view) and partial left (O: cranial view) radius. (P) P. cf. danielsi (NMS.2021.40.47), distal portion of right radius (ventral view) and proximal end of ?right radius. (Q) Primoscens sp. B (NMS.2021.40.59), distal and proximal portion of right radius. (R) P. cf. danielsi (NMS.2021.40.48), proximal end of ?right radius. (S‒V) P. cf. danielsi (NMS.2021.40.47), (S, T) left and (U, V) right carpometacarpus in (S, U) ventral and (T, V) dorsal view; the arrow indicates an enlarged detail of the proximal end. (W‒Z) M. primoscenoides, gen. et sp. nov. (holotype, NMS.2021.40.62), (W, X) left and (Y, Z) right carpometacarpus in (W, Y) ventral and (X, Z) dorsal view; the arrows indicate enlarged details of the proximal end. (AA, BB) Primoscens cf. minutus (NMS.2021.40.53), left carpometacarpus in (AA) ventral and (BB) dorsal view. (CC, DD) Pr. minutus (holotype; NHMUK A 4681), right carpometacarpus in (CC) ventral and (DD) dorsal view; the arrow indicates an enlarged detail of the proximal end. (EE) P. cf. danielsi (NMS.2021.40.47), right phalanx proximalis digiti majoris in ventral view. (FF) M. primoscenoides, gen. et sp. nov. (holotype, NMS.2021.40.62), left phalanx proximalis digiti majoris in ventral view. Abbreviations: ctd, cotyla dorsalis; ctv, cotyla ventralis; flx, processus flexorius; fst, fossa supratrochlearis; olc, olecranon; oma, os metacarpale alulare; pcd, processus cotylaris dorsalis; pim, processus intermetacarpalis; prc, small process at proximal end of sulcus tendinosus; prj, projection of proximal end of radius; psd, processus supracondylaris dorsalis; ptm, protuberantia metacarpalis; rdg, ridge from processus pisiformis to os metacarpale minus; tbc, tubercle dorsal to condylus dorsalis; tbd, tuberculum dorsale; trs, transition of caudal rim of trochlea carpalis into os metacarpale minus. The scale bars equal 5 mm.

The ulna () is longer than the humerus. The well-developed olecranon is not as narrow as in crown group Passeriformes. The cotyla dorsalis reaches proximally well beyond the cotyla ventralis. The proximal end of the radius () bears a well-defined projection, which also occurs in Primoscens.
The carpometacarpus () has a distinctive morphology, which characterises zygodactylids. The dorsal portion of the trochlea carpalis is proximally drawn-out; its ventral portion is craniocaudally narrow. A distinct, narrow fossa supratrochlearis runs across the dorsal portion of the trochlea carpalis. As in crown group Passeriformes, there is a shallow fossa between the processus pisiformis and the proximal end of the os metacarpale minus. The processus intermetacarpalis is well developed and contacts the os metacarpale minus, with which it does not fuse (contrary to crown group Passeriformes). The os metacarpale minus protrudes slightly farther distally than the os metacarpale majus. The distal end of the sulcus tendinosus is overhung by a small, broadly convex projection. The carpometacarpus of Primozygodactylus differs from that of Zygodactylus (as exemplified by Z. luberonensis; see Mayr Citation2008b) in that the bone is craniocaudally narrower and the processus intermetacarpalis smaller.
The phalanx proximalis digiti majoris () is craniocaudally narrow and lacks a processus internus indicis. Its shape resembles the corresponding phalanx of Zygodactylus luberonensis.
NMS.Z.2021.40.47 includes various fragments of the pelvis (). The synsacrum is nearly completely, and the long and slender processus terminales ischii of the ischium are also preserved. The shape of the latter resembles the processus terminalis ischii of ‘Zygodactylus’ grandei from the North American Green River Formation (Smith et al. Citation2018: fig. 5).
The femur () is an elongate and narrow bone. The proximal end lacks a well-developed crista trochanteris.
The tibiotarsus is the longest limb element (). The distal end corresponds well with the tibiotarsus of Zygodactylus (Ballmann Citation1969a). The sulcus extensorius is situated near the medial margin of the distal end of the bone (), whereas it is more centrally located in crown group Passeriformes. The condylus medialis is narrower than the condylus lateralis.
Figure 10. Tibiotarsus and tarsometatarsi of the Zygodactylidae from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). (A) Primozygodactylus cf. danielsi (NMS.2021.40.47), distal end of left tibiotarsus in cranial view. (B‒E) P. cf. danielsi (NMS.2021.40.47), right tarsometatarsus in (B) plantar, (C) dorsal, (D) lateral, and (E) medial view; the arrow indicates an enlarged detail of the distal end. (F‒I) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54), left tarsometatarsus in (F) plantar, (G) dorsal, (H) lateral, and (I) medial view; the arrows indicate enlarged details of the distal end. (J, K) P. cf. danielsi (NMS.2021.40.47), right tarsometatarsus, (J) distal end in distal view and (K) proximal end in proximal view. (L, M) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), left tarsometatarsus, (L) distal end in distal view and (M) proximal end in proximal view. (N) Zygodactylus luberonensis from the early Oligocene of France (SMF Av 519), distal end of left tarsometatarsus in lateroplantar view; ultraviolet-induced fluorescence photographs. (O, P) P. cf. danielsi (NMS.2021.40.50), left tarsometatarsus (NMS.2021.40.50) in (O) plantar and (P) distal view. (Q, R) Zygodactylidae, gen. et sp. indet. A (NMS.2021.40.60), left tarsometatarsus (NMS.2021.40.60) in (Q) plantar and (R) distal view. (S, T) Primoscens sp. A (NMS.2021.40.58), right tarsometatarsus (NMS.2021.40.58) in (S) plantar and (T) distal view. (U, V) Primoscens sp. B (NMS.2021.40.59), left tarsometatarsus (NMS.2021.40.59) in (U) plantar and (V) distal view. (W, X) Zygodactylidae, gen. et sp. indet. B (NMS.2021.40.61), left tarsometatarsus (NMS.2021.40.61) in (W) plantar and (X) distal view. Abbreviations: acc, trochlea accessoria; fdl, hypotarsal sulcus/canal for tendon of musculus flexor digitorum longus; fhl, hypotarsal sulcus/canal for tendon of musculus flexor hallucis longus; trIV, trochlea metatarsi IV proper. The scale bars equal 5 mm.
The greatly elongated tarsometatarsus () has a distinctive morphology. The foramina vascularia are small and the medial foramen is situated slightly proximal to the lateral one. Unlike in crown group Passeriformes, there is no ossified arcus extensorius and the impressiones retinaculi extensorii are likewise hardly visible. The hypotarsus encloses two canals for the tendons of musculus flexor digitorum longus and musculus flexor hallucis longus. The eminentia intercotylaris is prominent. Apart from the smaller accessory trochlea of the trochlea metatarsi IV, the distal end of the bone closely resembles the distal tarsometatarsus of Zygodactylus. The foramen vasculare distale is large and its dorsal opening is situated in a deep fossa. The trochlea metatarsi II is plantarly deflected and bears a distinct plantar projection. The trochlea metatarsi IV forms an accessory trochlea, which is delimited by a furrow from the main portion of the trochlea. Unlike in Zygodactylus, this accessory trochlea does not reach farther distally than the trochlea metatarsi IV proper.
The three pedal phalanges preserved in NMS.Z.2021.40.47 are elongated as in the Messel specimens of Primozygodactylus danielsi and as in Zygodactylus luberonensis.
Primoscens Harrison and Walker, Citation1977
Taxonomic remarks
The taxon Primoscens was established by Harrison and Walker (Citation1977) for an incomplete carpometacarpus from the London Clay of the Isle of Sheppey (). The only species included in the taxon, Primoscens minutus, was assigned to the Passeriformes in the original description, but M. Daniels (in Feduccia Citation1999, p. 166) identified two fossils of P. minutus among the avian material from Walton-on-the-Naze and considered the species closely related to the birds that are assigned to Primozygodactylus in the present study. Mayr (Citation1998) concurred with this hypothesis and figured the more complete of these putative P. minutus fossils (Mayr Citation1998: pl. 7). However, this specimen, which is now catalogued as NMS.2021.40.62, is clearly distinguished from the Primoscens minutus holotype in the morphology of the carpometacarpus and is here described as a new taxon (see further below).
The second fossil from Walton-on-the-Naze that was assigned to P. minutus by Daniels (NMS.2021.40.53) is indeed very similar to this species in carpometacarpus morphology. This fossil is here tentatively assigned to P. minutus.
The Daniels collection includes several other fossils that are likely to belong to Primoscens. One of these is a partial skeleton (NMS.2021.40.54), which, apart from its smaller size (), shows a close resemblance to Eozygodactylus americanus from the North American Green River Formation. Weidig (Citation2010) differentiated the taxon Eozygodactylus from Primoscens on the basis of a larger size and the presence of a processus supracondylaris dorsalis on the humerus. However, size is an insufficient criterion to distinguish higher-level taxa and comparisons of the humerus morphology were made with the above-mentioned fossil (NMS.2021.40.62) that was erroneously referred to Primoscens. If our assignment of NMS.2021.40.54 to Primoscens is correct, it is likely that Eozygodactylus Weidig, Citation2010 is a junior synonym of Primoscens Harrison and Walker, Citation1977.
Primoscens cf. minutus Harrison and Walker, Citation1977
Referred specimen
NMS.2021.40.53 (; proximal end of right ulna, left carpometacarpus, and a few other bone fragments), collected in 1991 by M. Daniels (original collector’s number WN 91718).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Tarsometatarsus, 16.0.
Remarks
The carpometacarpus of this fossil () shows a close resemblance to that of the P. minutus holotype (). As in the latter, the transition of the caudal rim of the trochlea carpalis into the os metacarpale minus is abrupt and both structures meet at a steep angle. This configuration of the carpal trochlea distinguishes Primoscens from Primozygodactylus (). The processus intermetacarpalis is somewhat smaller than in the P. minutus holotype, which we attribute to individual variation.
The proximal end of the ulna () resembles that of the new Primoscens species described below (), but the caudal margin of the processus cotylaris dorsalis is more rounded in NMS.2021.40.53.
Primoscens carolinae, sp. nov.
Holotype
NMS.2021.40.54 (; partial skeleton including left coracoid, right scapula, fragmentary cranial portion of sternum, both humeri, left ulna, both radii, left femur, distal ends of tibiotarsi, right tarsometatarsus, a few pedal phalanges), collected in 1992 by M. Daniels (original collector’s number WN 92747).
Diagnosis
Coracoid with medial margin of extremitas sternalis forming marked projection; spina externa of sternum slightly bifurcated; humerus with well-developed process supracondylaris dorsalis; femur shorter than humerus; elevated, subcircular muscle attachment scar proximal to condylus dorsalis (humerus); hypotarsus enclosing canal for tendon of musculus flexor digitorum longus and bearing a small sulcus for tendon of musculus flexor hallucis longus; distal end of tarsometatarsus narrow in dorsoplantar direction, with accessory trochlea of trochlea metatarsi IV situated close to trochlea metatarsi III; medial margin of accessory trochlea reaching midline of trochlea metatarsi III.
Differential diagnosis
Distinguished from the above fossil assigned to Primoscens minutus (NMS.2021.40.53) in that the caudal margin of the processus cotylaris dorsalis of the ulna is less rounded and forms a distal hook-like projection. Differs from Eozygodactylus americanus and ‘Zygodactylus’ grandei ‒ the only other Eocene zygodactylids with a well-developed processus supracondylaris dorsalis (humerus) ‒ in its much smaller size (humerus length 12.7 mm versus 16.8 and 18.6 mm, respectively).
Etymology
The new taxon is dedicated to the memory of Caroline Daniels, daughter of Michael and Pamela Daniels, who was a friend of one of the authors (ACK) and helped to collect several fossils in her father’s collection.
Type locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Referred specimens
NMS.2021.40.55 (; distal end of right humerus and distal end of right ulna), collected in 1990 by M. Daniels (original collector’s number WN 90664); NMS.2021.40.56 (; cranial portion of left scapula and distal end of left tarsometatarsus), collected in 1992 by M. Daniels (original collector’s number WN 92722).
Tentatively referred specimen
NMS.2021.40.57 (partial femora), collected in 1976 by M. Daniels (original collector’s number WN 76137).
Measurements (maximum length, in mm)
NMS.2021.40.54: Right humerus, 12.7; left ulna, 13.1; left femur, 11.8; right tarsometatarsus, 14.7.
Taxonomic remarks
Because the carpometacarpus of Primoscens carolinae, sp. nov. is unknown, direct comparisons with the holotype of P. minutus are not possible. However, and as detailed below, the ulna of the new species is distinguished from the ulna of specimen NMS.2021.40.53, which we tentatively assign to P. minutus (and which includes a carpometacarpus that is very similar to the holotype of P. minutus). Therefore, we consider both specimens to be from different species, and it would hamper comparisons between the London Clay zygodactylids if the more complete specimen were left unnamed. We acknowledge some ambiguity in our decision to refer NMS.2021.40.53 to P. minutus and to describe NMS.2021.40.54 as a new species, but the great similarity of the carpometacarpus of the former specimen to that of P. minutus and the lack of this bone in the latter specimen leaves us no reasonable alternative. Only future discoveries of a carpometacarpus of P. carolinae will be able to corroborate or refute our taxonomic decision.
Description and comparisons
Unlike in Primozygodactylus, the extremitas omalis of the coracoid () has a hook-shaped outline. The tip of the processus procoracoideus is broken in the holotype. The facies articularis scapularis is shallow. The medial margin of the extremitas forms a marked projection.
The furcula is not preserved in the specimens. The scapula () appears to have been proportionally shorter than in Primozygodactylus and has a shorter acromion.
The cranial fragment of the sternum preserved in the holotype () exhibits a mediolaterally wide and slightly bifurcated spina externa. In its shape, the spina externa resembles that of ‘Zygodactylus’ grandei from the North American Green River Formation (Smith et al. Citation2018), whereas the spina externa of Primozygodactylus is narrow and blade-like.
The humerus () is proportionally stouter than in Primozygodactylus, and unlike in the latter, the crista bicipitalis does not form a convexity. The proximal and distal ends of the bone are proportionally larger than in Primozygodactylus. The crista deltopectoralis measures about one third of the humerus length and has a straight dorsal margin. The distal end of the bone is strikingly similar to the distal humerus of crown group Passeriformes, which is especially true for the presence of a strongly developed processus supracondylaris dorsalis. Furthermore as in crown group Passeriformes there is an elevated, subcircular muscle attachment scar proximal to the condylus dorsalis (in passerines, this attachment site forms a tubercle). The processus flexorius is strongly developed and distally projected. The elongate fossa musculi brachialis is situated close to the medial margin of the bone.
The ulna () is only slightly longer than the humerus. The proximal end of the bone has a somewhat shorter cotyla dorsalis than the ulna of Primozygodactylus, and the processus cotylaris dorsalis forms a small hook (whereas it is broadly rounded in Primozygodactylus). The presence of this hook also distinguishes the proximal ulna of P. carolinae from that of specimen NMS.2021.40.53, which is here classified as P. cf. minutus and in which the caudal margin of the processus cotylaris dorsalis is more rounded ()). Except for the somewhat more pointed tuberculum carpale, the distal end of the bone resembles the distal ulna of Primozygodactylus. The distal end the radius forms a small projection ()).
The femur is shorter than the humerus (as in ‘Z’. grandei) and has a wider distal end than that of Primozygodactylus, in which the portion lateral to the sulcus intercondylaris is narrower.
Only the distal end of the tibiotarsus is preserved in the fossils, which corresponds well to the distal tibiotarsus of Primozygodactylus. However, the sulcus extensorius is somewhat narrower than in the latter taxon.
The tarsometatarsus () resembles that of Primozygodactylus in its overall morphology, but exhibits some distinct differences in osteological details. The hypotarsus encloses a single canal for the tendon of musculus flexor digitorum longus, whereas the tendon of m. flexor hallucis longus ran along a sulcus. The shaft lacks a crista plantaris lateralis. The foramen vasculare distale is very large. The trochlea metatarsi II is proportionally smaller than that of Primozygodactylus. Unlike in the latter, the medial margin of the accessory trochlea of the trochlea metatarsi IV reaches the midline of the trochlea metatarsi III. The trochlear furrow of the trochlea metatarsi III is more pronounced than in Primozygodactylus.
The holotype includes a long ungual phalanx, which shows little curvature and has a very low tuberculum flexorium ().
Primoscens sp. A
Referred specimen
NMS.2021.40.58 (; right tarsometatarsus lacking trochlea metatarsi II), collected in 1983 by M. Daniels (original collector’s number WN 83440).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Tarsometatarsus, 16.0.
Remarks
As far as comparisons are possible, this specimen closely resembles the tarsometatarsus of Primoscens carolinae in its morphology, but is from a larger species.
Primoscens sp. B
Referred specimen
NMS.2021.40.59 (; a few vertebrae, fragmentary left coracoid, fragmentary radius, left tarsometatarsus, and pedal phalanges), collected in 1998 by M. Daniels (original collector’s number WN 98998).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Tarsometatarsus, 13.8.
Remarks
This tarsometatarsus shows a close resemblance to that of Primoscens carolinae, but is stouter and has a wider shaft (Compare ).
Zygodactylidae, gen. et sp. indet. A
Referred specimens
NMS.2021.40.60 (; partial skeleton including partial furcula, partial left scapula, distal end of left ulna, radius, phalanx proximalis digiti majoris, right femur, right tarsometatarsus lacking proximal end, and several pedal phalanges), collected in 1990 by M. Daniels (original collector’s number WN 90660).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Right femur, 10.8.
Remarks
These bones belong to a very small species, with the femur and tarsometatarsus being somewhat smaller than the corresponding bones of the holotype of Primoscens carolinae. The distal end of the tarsometatarsus () differs from that of P. carolinae () in that the trochlea accessoria is more plantarly deflected and the trochlea metatarsi IV proper is proportionally smaller.
The extremitas omalis of the furcula ()) is wider than in the new species described below ()), which prevents an assignment to this species. The shape of the extremitas omalis closely resembles that of Zygodactylus luberonensis ().
In size, NMS.2021.40.60 corresponds well to Primoscens minutus. However, the fossil does not include a carpometacarpus, so that there exists no basis for a referral to the latter species.
Zygodactylidae, gen. et sp. indet. B
Referred specimen
NMS.2021.40.61 (; partial skeleton including rostral portion of upper beak, partial right quadrate, left coracoid, left humerus in block of matrix that also contains other bones, left tarsometatarsus, and some pedal phalanges), collected in 1997 by M. Daniels (original collector’s number WN 97966).
Locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Left tarsometatarsus, 19.3.
Remarks
This specimen corresponds to Primozygodactylus cf. danielsi in its size, but the distal end of the tarsometatarsus is proportionally wider and the trochlea metatarsi II less plantarly deflected (Compare ). The coracoid () exhibits a long processus procoracoideus and a flange-like projection on the medial margin of the shaft similar to that reported for ‘Zygodactylus’ grandei by Smith et al. (Citation2018: fig. 4).
The shape of the upper beak () is similar to that of Primozygodactylus spp., as exemplified by the specimens from Messel (Mayr Citation1998, Citation2017a). The preserved morphology of the quadrate () likewise corresponds to that of Primozygodactylus (), but the caudal surface of the processus oticus exhibits smaller pneumatic openings.
Parapasseres incertae sedis
Minutornis, gen. nov.
Type species
Minutornis primoscenoides, sp. nov.
Differential diagnosis
A very small taxon that is distinguished from Primoscens (as exemplified by the holotype of Primoscens minutus) in that the os metacarpale alulare is proportionally longer, the transition of the caudal rim of the trochlea carpalis into the os metacarpale minus is less abrupt, a ridge from the processus pisiformis to the os metacarpale minus is absent, and the processus intermetacarpalis is somewhat smaller. Furthermore, it differs from Primoscens carolinae in that the spina externa (sternum) is more strongly bifurcated and forms two dorsally projected branches, and the humerus has a larger tuberculum dorsale and lacks an enlarged processus supracondylaris dorsalis. Distinguished from Primozygodactylus in that the spina externa is bifurcated (blade-like in Primozygodactylus), the extremitas omalis of the furcula is not greatly widened, the extremitas omalis of the coracoid is hook-shaped, and the humerus is stouter with a larger tuberculum dorsale. Compared with Eofringillirostrum, the processus procoracoideus of the new taxon is distinctly longer.
Etymology
From minutus (Lat.): tiny and ornis (Gr.): bird.
Remarks
This taxon was erroneously designated as Primoscens by Mayr (Citation1998), who assigned the holotype of the type species (NMS.2021.40.62) to Primoscens minutus. Weidig (Citation2010) and Smith et al. (Citation2018) likewise based some of their comparisons with Primoscens on NMS.2021.40.62.
Minutornis primoscenoides, gen. et sp. nov.
Holotype
NMS.2021.40.62 (; left coracoid, fragmentary furcula, partial sternum, both humeri, both carpometacarpi, phalanx proximalis digiti majoris), collected in 1987 by M. Daniels (original collector’s number WN 87558A).
Diagnosis
As for genus.
Etymology
The species epithet refers to the resemblance between the carpometacarpus of the new taxon and that of Primoscens, and the name of the new species alludes to the fact that the holotype was previously assigned to Primoscens minutus (Mayr Citation1998).
Type locality and horizon
Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm)
Left humerus, 10.9; right carpometacarpus, 7.3.
Description and comparisons
The coracoid () has a hook-shaped extremitas omalis. The tip of the long processus procoracoideus is expanded. The facies articularis scapularis is shallow. The medial margin of the extremitas sternalis is damaged, so that it is unknown whether there was a medial projection as in Primoscens. The processus lateralis is short.
The extremitas omalis of the furcula () is much narrower than in Primozygodactylus () and has an asymmetric, somewhat club-shaped outline. The extremitas sternalis () has a pathological morphology and appears to show a healed fracture.
The sternum () is more elongate than that of Primozygodactylus (as exemplified by complete sterna from Messel; see Mayr Citation1998). Unlike in Primozygodactylus, the spina externa is strongly bifurcated and forms two dorsally projected branches, which, in lateral view, give the spina a markedly kinked shape. The carina sterni is very deep.
The humerus () is somewhat stouter than that of Primozygodactylus and Primoscens, and the tuberculum dorsale is proportionally larger. The proximal portion of the proximal end is caudally deflected. The dorsal margin of the distal end forms a proximodistally deep tuberculum supracondylare dorsale. The distally protruding processus flexorius is prominent.
The carpometacarpus () compares well to that of the Primoscens minutus holotype () in its size, but there are various subtle morphological differences. In particular, the os metacarpale alulare of NMS.2021.40.62 is longer than in P. minutus, and the transition of the caudal rim of the trochlea carpalis into the os metacarpale minus is less abrupt. Unlike in the holotype of P. minutus, there is no ridge from the processus pisiformis to the os metacarpale minus. As in P. minutus, the processus intermetacarpalis is well-developed and reaches the os metacarpale minus. The dorsal margin of the os metacarpale majus bears a distinct protuberantia metacarpalis. Proximal to it, and at the proximal end of the sulcus tendinosus, there is a small, cranially directed process, which is better developed than in Primozygodactylus; this process is a derived characteristic of crown group Passeriformes (Mayr Citation1998, p. 46; Mayr and Manegold Citation2006). The os metacarpale minus has an undulated caudal margin.
The phalanx proximalis digiti majoris ((FF)) is craniocaudally wider than in Primozygodactylus.
Results of the phylogenetic analysis
The analysis of the character matrix in the Electronic Supplementary Material resulted in 84 most parsimonious trees. With regard to the critical fossil taxa, the strict consensus tree ()) showed a poor resolution. However, the analysis obtained a monophyletic Psittacopasseres and supported a clade including the Psittacopedidae, Zygodactylidae, and crown group Passeriformes.
Figure 11. (A) Strict consensus tree of 84 most parsimonious trees resulting from the phylogenetic analysis (L = 300, CI = 0.34, RI = 0.62); bootstrap support values are indicated next to the internodes. (B) Majority rule consensus tree; the numbers indicate the percentage of individual trees in which the respective nodes were present.
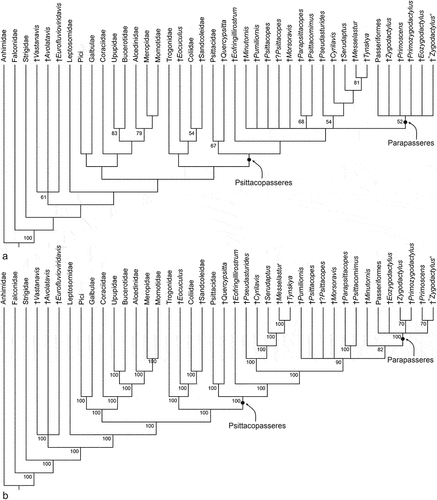
The majority rule consensus tree ()) was better resolved and recovered a clade including Pumiliornis, Morsoravis, Psittacopedidae, Zygodactylidae and crown group Passeriformes. Whereas the strict consensus tree placed the included zygodactylid taxa in a polytomy together with crown group Passeriformes, the majority rule consensus tree resulted in sister group relationships between Primozygodactylus and Zygodactylus and between Primoscens and ‘Zygodactylus’ grandei, respectively. Minutornis is the sister taxon of a clade including zygodactylids and crown group Passeriformes in the majority rule consensus tree.
However, we note that the position of the Halycyornithidae and Messelasturidae within Psittacopasseres does not conform to the results of other analyses (e.g. Mayr Citation2015; Ksepka et al. Citation2019, Citation2020) and is likely to be incorrect. Unlike in zygodactyl taxa of the Psittacopasseres, the trochlea accessoria of the Halycyornithidae and Messelasturidae is not separated by a furrow from the trochlea metatarsi IV and the coracoid exhibits a foramen nervi supracoracoidei.
Concerning the extant taxa, some results of our analysis likewise do not conform to well-supported tree topologies obtained from current sequence-based studies (e.g. Prum et al. Citation2015; Kuhl et al. Citation2021), which places a caveat on the affinities of the fossils. Instances of major incongruence concern the position of Psittacopasseres in a clade together with coraciiform and piciform birds (instead of being the sister taxon of the Falconidae), as well as a sister group relationship between the Coliiformes and Trogoniformes, and the nesting of the Coliiformes within a clade including coraciiform, piciform, and psittacopasserine birds.
Discussion
Comparison of the Psittacopedidae and Zygodactylidae with putatively related Palaeogene taxa
As detailed in the introduction, the three early Eocene taxa Morsoravis, Pumiliornis, Eofringillirostrum, as well as the early Oligocene Eocuculus, were associated with Psittacopes in recent analyses (Mayr Citation2015; Ksepka et al. Citation2019, Citation2020). These taxa show a high disparity in the shape of the beak, which is greatly elongated and narrow in Pumiliornis, thrush-like in Morsoravis, and conical and finch-like in Eofringillirostrum (the beak of Eocuculus is unknown). Our analysis provides some support for a clade including Pumiliornis, Morsoravis, and psittacopedids, but this clade was only recovered in the majority rule consensus tree.
Apart from the much longer beak, Pumiliornis shows an overall resemblance to the Psittacopedidae in its postcranial skeletal morphology, but lacks a pons supratendineus, which suggests different locomotory characteristics. Moreover, close comparisons are impeded by the fact that Pumiliornis is mainly known from compression fossils. A tarsometatarsus from the Nanjemoy Formation of Virginia, which was tentatively assigned to Pumiliornis by Mayr et al. (Citation2022), shows a close resemblance to the tarsometatarsus of ?Psittacopes occidentalis. However, it is uncertain whether this North American fossil () was correctly assigned to Pumiliornis (the tentative classification was mainly based on the absence of a pons supratendineus in a referred tibiotarsus), and it appears possible that the fossil is instead from the Psittacopedidae. Unlike in the fossils from Walton-on-the-Naze, there is no dorsally open sulcus between the foramen vasculare distale and the incisura intertrochlearis lateralis of Pumiliornis ()).
Figure 12. Comparison of major postcranial bones of the Psittacopedidae, Zygodactylidae, and possibly related Palaeogene taxa. (A, B) left coracoid (dorsal view) of the Zygodactylidae (A: Primozygodactylus cf. danielsi, NMS.2021.40.47; B: Primoscens carolinae, sp. nov., holotype, NMS.2021.40.54). (C, D) left coracoid (dorsal view) of the Psittacopedidae (C: ?Psittacopes occidentalis, sp. nov., holotype, NMS.Z.2021.40.44; D: Psittacomimus eos, gen. et sp. nov., holotype, NMS.Z.2021.40.38). (E) P. cf. danielsi (NMS.2021.40.49), right humerus in cranial view. (F) Parapsittacopes bergdahli (NMS.Z.2021.40.43), left humerus in cranial view. (G) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), left ulna in cranial view. (H) Pa. bergdahli (holotype, SMF Av 653), left ulna in cranial view. (I‒K) Right carpometacarpus (ventral view) of (I) P. cf. danielsi (NMS.2021.40.47), (J) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), and (K) Ps. eos, gen. et sp. nov. (holotype, NMS.Z.2021.40.38). (L‒R) Tarsometatarsi (dorsal view) of (L) P. cf. danielsi (NMS.2021.40.47; right side); (M) Psittacopedidae, gen. et sp. indet. B (NMS.Z.2021.40.46; left side); (N) Ps. eos (NMS.Z.2021.40.39; left side); (O) Morsoravis sedilis from the early Eocene Fur Formation in Denmark (holotype, MGUH 28930; left side); (P) Eocuculus cherpinae from the early Oligocene of Colorado, USA (DM 10682; right side, cast of holotype); (Q) Eocuculus cf. cherpinae from the early Oligocene of France (SMF Av 425; right side); and (R) Pumiliornis tessellatus from the early/middle Eocene of Messel (SMF-ME 2475B; left side). (S‒W) Left tarsometatarsus from the early Eocene Nanjemoy Formation of Virginia (USNM PAL 771592), which was tentatively referred to Pumiliornis by Mayr et al. (Citation2022), in (S) dorsal, (T) plantar, (U) lateral, (V) proximal, and (W) distal view. (X‒Z) ?Psittacopes occidentalis, sp. nov. (holotype, NMS.Z.2021.40.44), partial right tarsometatarsus in (X) dorsal, (Y) lateral, and (Z) plantar view. (AA‒CC) Ps. eos, gen. et sp. nov., distal end of right tarsometatarsus in plantar and distal view (AA, BB; NMS.Z.2021.40.39), proximal end of left tarsometatarsus in proximal view (CC: holotype, NMS.Z.2021.40.38). (DD‒FF) P. cf. danielsi (NMS.2021.40.47), distal end of right tarsometatarsus in (DD) plantar and (EE) distal view; (FF) proximal end in proximal view. (GG‒II) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), distal end of left tarsometatarsus in (GG) plantar and (HH) distal view; (II) proximal end in proximal view. Abbreviations: acc, trochlea accessoria; fdl, hypotarsal sulcus/canal for tendon of musculus flexor digitorum longus; fhl, hypotarsal sulcus/canal for tendon of musculus flexor hallucis longus; ttc, tuberositas musculi tibialis cranialis. The scale bars equal 5 mm.

The wings of Morsoravis are unknown, but the tarsometatarsus ()) has a proportionally smaller trochlea metatarsi III than that of psittacopedids (in contrast to psittacopedids, the trochlea metatarsi III is distinctly narrower than the tarsometatarsus shaft in Morsoravis), and the trochlea metatarsi IV does not exhibit the derived morphology found in the Psittacopedidae, in which it is shorter than in Morsoravis and more laterally projected. Unlike in the Psittacopedidae and Zygodactylidae, the trochlea metatarsi II of Morsoravis exhibits a distinct furrow on its medial surface (Bertelli et al. Citation2010; Mayr Citation2011). The morphology of the plantar surface of the tarsometatarsus of Morsoravis is unknown, but the position of the toes in the holotype of M. sedilis suggests the absence of fully zygodactyl feet and, hence, a large trochlea accessoria is also likely to be absent. The distinct differences in the morphology of the tarsometatarsus suggest that Morsoravis is outside a clade including psittacopedids and zygodactylids and place a caveat on the results of our phylogenetic analysis.
Our analysis does not support close affinities between Eocuculus and psittacopedids. The holotype of Eocuculus cherpinae, from the early Oligocene of North America, also differs from the London Clay psittacopedids in the morphology of the tarsometatarsus ()), which in E. cherpinae has a smaller foramen vasculare distale and a better defined (in dorsal view) trochlea metatarsi II that reaches farther distally. Still, the tarsometatarsi of psittacopedids and Eocuculus show some overall similarity in their shapes, and this is particularly true if comparisons are made with the specimen from the early Oligocene of France ()), which was referred to Eocuculus by Mayr (Citation2006). In this specimen there is a dorsally open sulcus on the distal end of the bone between the foramen vasculare distale and the incisura intertrochlearis lateralis. However, and not least due to the fact that the skeletal morphology of the E. cherpinae holotype is too poorly preserved for detailed comparisons, it may not be possible to unambiguously assign the better preserved French fossils to Eocuculus. Our analysis recovered Eocuculus as sister taxon of coliiform birds, but because these taxa were also associated with the Trogoniformes (which conflicts with all current sequence- and morphology-based phylogenies), this result is likely to be due to inadequate character sampling.
Eofringillirostrum is distinguished from psittacopedids and zygodactylids in the morphology of its legs, which are shorter than in zygodactylids and have longer and more pointed ungual phalanges than those of psittacopedids and zygodactylids; unlike in psittacopedids, the first phalanx of the second toe of Eofringillirostrum is shortened and the carpometacarpus exhibits a well-developed processus intermetacarpalis. However, these differences may be due to a specialised ecology of Eofringillirostrum and do not per se preclude close affinities to psittacopedids.
Apart from psittacopedids, the Daniels collection includes fossils of other ‘parrot’-like birds, and in addition to remains of the Halcyornithidae and Messelornithidae (Mayr Citation2022) there are various other undescribed species. We expect future studies of these fossils to contribute to a better understanding of the phylogenetic affinities of some of the above taxa and aim for their timely description, which will probably yield new data that improve our phylogeny.
Monophyly and interrelationships of the Zygodactylidae
A clade including the Zygodactylidae and crown group Passeriformes is well-supported and was termed Parapasseres (Mayr Citation2015). In addition to a very long and slender tarsometatarsus, the Zygodactylidae and crown group Passeriformes share distinctive derived characters of the carpometacarpus, which exhibits an enlarged processus intermetacarpalis, a protuberantia metacarpalis, and a cranially directed process proximal to this protuberantia.
Actually, the carpometacarpus of zygodactylids is so similar to that of crown group Passeriformes that it was erroneously assigned to the Passeriformes by Ballmann (Citation1969a), who referred only fragmentary leg bones to the Neogene species Zygodactylus ignotus and Z. grivensis (distal tibiotarsus and distal tarsometatarsus in the case of Z. ignotus from the early Miocene of Germany and distal tarsometatarsus in the case of Z. grivensis from the middle Miocene of France; Ballmann Citation1969a; Citationb). Mayr (Citation1998, p. 54) identified a possible carpometacarpus of Z. ignotus in the fossil material from Wintershof-West, the type locality of Z. ignotus, which is here for the first time figured (). This carpometacarpus was assigned to the Passeriformes by Ballmann (Citation1969a, p. 51; his ‘Species F’), but actually it closely resembles the carpometacarpus of Z. luberonensis (). As in the latter species, the processus intermetacarpalis of the Z. ignotus carpometacarpus is much better developed than that of the zygodactylids from Walton-on-the-Naze and, with regard to the development of this process, the carpometacarpus of Z. ignotus more closely corresponds to that of crown group Passeriformes (). The carpometacarpus of Primoscens minutus was likewise mistaken for that of a passerine by Harrison and Walker (Citation1977).
Figure 13. Comparison of selected bones of Primozygodactylus and Primoscens with those of Zygodactylus. (A‒C) Humerus of (A) Primozygodactylus cf. danielsi (NMS.2021.40.47; right side, cranial view); (B) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54; right side, caudal view); and (C) Zygodactylus luberonensis (SMF Av 519; right side, caudal view; coated with ammonium chloride). (D‒F) Carpometacarpus of (D, E) P. cf. danielsi (NMS.2021.40.47; D: ventral view, E: dorsal view); (F, G) Z. ignotus (SNSB-BSPG 18211; F: ventral view, G: dorsal view); and (H) Z. luberonensis (SMF Av 519; dorsal view, coated with ammonium chloride). (I‒K) Distal end of tarsometatarsus (plantar view) of (I) P. cf. danielsi (NMS.2021.40.47; right side); (J) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54; left side); and (K) Z. ignotus (SNSB-BSPG 18165; right side). Abbreviations: acc, trochlea accessoria (broken in Zygodactylus); fst, fossa supratrochlearis; pim, processus intermetacarpalis; prc, small process at proximal end of sulcus tendinosus; psd, processus supracondylaris dorsalis; ptm, protuberantia metacarpalis. The scale bars equal 5 mm; (G), (H), and (K) are not to scale.
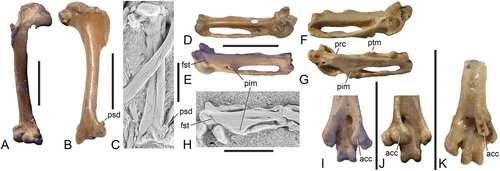
Figure 14. Comparison of selected bones of the Zygodactylidae and crown group Passeriformes. (A) Primoscens carolinae, sp. nov. (holotype, NMS.2021.40.54), distal end of right humerus in cranial view. (B) Sylvia borin (Passeriformes, Sylviidae), distal end of right humerus in cranial view. (C, D) Primozygodactylus cf. danielsi (NMS.2021.40.47), right carpometacarpus in (C) ventral and (D) dorsal view. (E, F) Saxicola rubicola (Passeriformes, Muscicapidae), right carpometacarpus in (E) ventral and (F) dorsal view. (G) P. cf. danielsi (NMS.2021.40.47), distal end of left tibiotarsus in cranial view. (H) Motacilla alba (Passeriformes, Motacillidae), distal end of left tibiotarsus in cranial view. (I, J) P. cf. danielsi (NMS.2021.40.47), distal end of left tarsometatarsus in (I) plantar and (J) distal view. (K, L) Pr. carolinae, sp. nov. (holotype, NMS.2021.40.54), distal end of right tarsometatarsus in (K) plantar and (L) distal view. (M, N) Zygodactylus grivensis (holotype, after Mayr Citation1998) from the middle Miocene of France, distal end of right tarsometatarsus in (M) plantar and (N) distal view. (O, P) Fringilla montifringilla (Passeriformes, Fringillidae), distal end of left tarsometatarsus in (O) plantar and (P) distal view. All drawings except those in (M) and (N) are by Michael Daniels. Abbreviations: acc, trochlea accessoria; pim, processus intermetacarpalis; psd, processus supracondylaris dorsalis; pst, pons supratendineus; tbc, tubercle dorsal to condylus dorsalis. Not to scale.
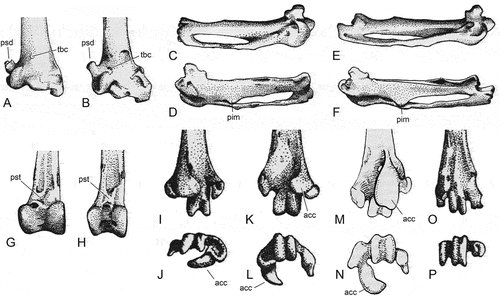
Although a clade including zygodactylids and crown group Passeriformes is well supported, the Zygodactylidae exhibit a high diversity in their skeletal morphology, and the monophyly of the taxon is poorly established. Even the two unambiguous parapasserine taxa from Walton-on-the-Naze ‒ Primoscens and Primozygodactylus ‒ are remarkably different in their skeletal morphology, and the disparate humerus and tarsometatarsus morphologies suggest different ecotypes.
The distal end of the humerus of Primoscens (as exemplified by P. carolinae, sp. nov.) shows a derived morphology, with a large processus supracondylaris dorsalis and a tubercle proximal to the condylus dorsalis (); these features are absent in Primozygodactylus, and among extant birds they are only found in the Passeriformes (a well-developed processus supracondylaris dorsalis is also present in Eozygodactylus americanus, ‘Zygodactylus’ grandei, and Zygodactylus luberonensis). Furthermore, Primoscens shares with crown group Passeriformes a small, hook-like projection on the distal end of the radius and an (albeit only slightly) bifurcated spina externa of the sternum. These two features are likewise absent in Primozygodactylus. Mayr (Citation2017b, p. 218) hinted at the possibility that zygodactylids are paraphyletic with respect to crown group Passeriformes and hypothesised that the fossils that are here assigned to Primoscens may be more closely related to crown group Passeriformes than is Primozygodactylus.
The majority rule consensus tree of our analysis supports a sister group relationship between Primozygodactylus and Zygodactylus. In both taxa the extremitas omalis of the coracoid has a rounded tip, whereas it is hook-like in Primoscens and crown-group Passeriformes. The tarsometatarsus of Primozygodactylus is also more similar to that of Zygodactylus in the morphology of the distal end, which is less compressed in dorsoplantar direction than in Primoscens and has a proportionally larger and more plantarly directed trochlea metatarsi II (). In plantar and distal view, the distal end of the tarsometatarsus of Primoscens resembles that of Psittacomimus, with the bone being more compressed in the dorsoplantar direction than in Primozygodactylus, and the trochlea accessoria of the trochlea metatarsi IV being situated closer to the trochlea metatarsi III (). Outgroup comparisons with the Psittacopedidae suggest that the morphology of the distal tarsometatarsus of Primoscens is plesiomorphic for the Zygodactylidae.
Eozygodactylus americanus and ‘Zygodactylus’ grandei from the Green River Formation were recovered in a clade together with the early Oligocene Zygodactylus luberonensis in the analysis of Smith et al. (Citation2018). However, we note that the morphology of the accessory trochlea of ‘Z’. grandei was misinterpreted by Smith et al. (Citation2018) and, unlike in Zygodactylus, it is not distally elongated and resembles the accessory trochlea of Primoscens carolinae in its relative size. As detailed above, it is possible that Eozygodactylus Weidig, Citation2010 is a junior synonym of Primoscens Harrison and Walker, Citation1977, and we consider it likely that ‘Zygodactylus’ grandei is a representative of one of these two taxa (the species was recovered as the sister taxon of Eozygodactylus americanus in the analysis of Smith et al. Citation2018 and resulted as the sister taxon of Primoscens carolinae in the majority consensus tree of our study).
In summary, early Eocene representatives of the Parapasseres exhibit a high degree of morphological homoplasy, which impedes a straightforward resolution of their interrelationships. The fossils that are here assigned to Primoscens show a striking similarity to crown group Passeriformes in the morphology of the humerus. By contrast, Primozygodactylus is more similar to crown group Passeriformes in features of the furcula and tarsometatarsus, whereas Minutornis shares a distinctly bifurcated spina externa with crown group Passeriformes, but is distinguished from other zygodactylids and extant passerines in the much narrower extremitas omalis of the furcula. The Neogene Zygodactylus adds to this confusing character mosaic and shows a close resemblance to crown group Passeriformes in the morphology of the carpometacarpus. Currently, no compelling character evidence exists in order to support a monophyletic Zygodactylidae, with distinctive characters of these birds being likely to be plesiomorphic for the Psittacopasseres (morphology of the distal end of the tarsometatarsus) or being also found in crown group Passeriformes (derived features of the carpometacarpus and long tarsometatarsus).
Mayr_and_Kitchener_electronic_supplementary_material.docx
Download MS Word (122.6 KB)Acknowledgments
We thank Sven Tränkner for taking some of the photographs (additional images were taken by GM). Comments from two anonymous reviewers improved the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2022.2141629.
References
- Aldiss DT. 2012. The stratigraphical framework for the Palaeogene successions of the London Basin, UK. Brit Geol Surv Open Rep. 12(4):1–87.
- Ballmann P. 1969a. Die Vögel aus der altburdigalen Spaltenfüllung von Wintershof (West) bei Eichstätt in Bayern. Zitteliana. 1:5–60.
- Ballmann P. 1969b. Les oiseaux miocènes de La Grive-Saint-Alban (Isère). Geobios. 2:157–204. doi:10.1016/S0016-6995(69)80005-7.
- Bertelli S, Lindow BEK, Dyke GJ, Chiappe LM. 2010. A well-preserved ‘charadriiform-like’ fossil bird from the Early Eocene Fur Formation of Denmark. Palaeontology. 53(3):507–531. doi:10.1111/j.1475-4983.2010.00950.x.
- Brodkorb P. 1971. Catalogue of fossil birds: part 4 (Columbiformes through Piciformes). Bull Florida State Mus nat hist. Biol Sci. 15:163–266.
- Chandler RM. 1999. Fossil birds of Florissant, Colorado: with a description of a new genus and species of cuckoo. Geol Res Div Techn Rep. NPS/NRGRD/GRDTR-99:49–53.
- Collinson ME, Adams NF, Manchester SR, Stull GW, Herrera F, Smith SY, Andrew MJ, Kenrick P, Sykes D. 2016. X-ray micro-computed tomography (Micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany. 94(9):697–711. doi:10.1139/cjb-2016-0078.
- Feduccia A. 1999. The origin and evolution of birds. 2nd ed. New Haven: Yale University Press.
- Goloboff PA. 1993. NONA version 2.0 [computer software]. S. M. de Tucumán: Published by the author.
- Grande L. 2013. The lost world of Fossil Lake. Snapshots from deep time. Chicago: University of Chicago Press.
- Harrison CJO, Walker CA. 1977. Birds of the British Lower Eocene. Tert Res Spec Pap. 3:1–52.
- Hieronymus TL, Waugh DA, Clarke JA. 2019. A new zygodactylid species indicates the persistence of stem passerines into the early Oligocene in North America. BMC Evol Biol. 19(1):3. doi:10.1186/s12862-018-1319-6.
- Ksepka DT, Grande L, Mayr G. 2019. Oldest finch-beaked birds reveal parallel ecological radiations in the earliest evolution of passerines. Curr Biol. 29(4):657–663. doi:10.1016/j.cub.2018.12.040.
- Kuhl H, Frankl-Vilches C, Bakker A, Mayr G, Nikolaus G, Boerno ST, Klages S, Timmermann B, Gahr M, Battistuzzi FU. 2021. An unbiased molecular approach using 3′-UTRs resolves the avian family-level tree of life. Mol Biol Evol. 38(1):108–127. doi:10.1093/molbev/msaa191.
- Mayr G. 1998. „Coraciiforme” und „piciforme” Kleinvögel aus dem Mittel-Eozän der Grube Messel (Hessen, Deutschland). Cour Forsch-Inst Senckenberg. 205:1–101.
- Mayr G. 1999. Pumiliornis tessellatus n. gen. n. sp., a new enigmatic bird from the middle Eocene of Grube Messel (Hessen, Germany). Cour Forsch-Inst Senckenberg. 216:75–83.
- Mayr G. 2004. The phylogenetic relationships of the early Tertiary Primoscenidae and Sylphornithidae and the sister taxon of crown group piciform birds. J Ornithol. 145:188–198. doi:10.1007/s10336-003-0018-1.
- Mayr G. 2006. A specimen of Eocuculus Chandler, 1999 (Aves, ?Cuculidae) from the early Oligocene of France. Geobios. 39(6):865–872. doi:10.1016/j.geobios.2005.10.007.
- Mayr G. 2008a. Pumiliornis tessellatus Mayr, 1999 revisited — new data on the osteology and possible phylogenetic affinities of an enigmatic Middle Eocene bird. Paläontologische Zeitschrift. 82(3):247–253. doi:10.1007/BF02988891.
- Mayr G. 2008b. Phylogenetic affinities of the enigmatic avian taxon Zygodactylus based on new material from the early Oligocene of France. J Syst Palaeontol. 6(3):333–344. doi:10.1017/S1477201907002398.
- Mayr G. 2009. Paleogene fossil birds. 1st ed. Heidelberg: Springer.
- Mayr G. 2011. On the osteology and phylogenetic affinities of Morsoravis sedilis (Aves) from the early Eocene Fur Formation of Denmark. Bulletin of the Geological Society of Denmark. 59:23–35. doi:10.37570/bgsd-2011-59-03.
- Mayr G. 2015. A reassessment of Eocene parrotlike fossils indicates a previously undetected radiation of zygodactyl stem group representatives of passerines (Passeriformes). Zool Scr. 44(6):587–602. doi:10.1111/zsc.12128.
- Mayr G. 2017a. New species of Primozygodactylus from Messel and the ecomorphology and evolutionary significance of early Eocene zygodactylid birds (Aves, Zygodactylidae). Historical Biology. 29(7):875–884. doi:10.1080/08912963.2016.1261135.
- Mayr G. 2017b. Avian evolution: the fossil record of birds and its paleobiological significance. Chichester: Wiley-Blackwell.
- Mayr G. 2020. A remarkably complete skeleton from the London Clay provides insights into the morphology and diversity of early Eocene zygodactyl near-passerine birds. Journal of Systematic Palaeontology. 18(22):1891–1906. doi:10.1080/14772019.2020.1862930.
- Mayr G. 2021. On the occurrence of lateral openings and fossae (pleurocoels) in the thoracic vertebrae of neornithine birds and their functional significance. Vertebr Zool. 71:453–463. doi:10.3897/vz.71.e71268.
- Mayr G. 2022. Paleogene fossil birds. 2nd ed. Cham (Switzerland): Springer.
- Mayr G, Daniels M. 1998. Eozäne Papageien von Messel (Hessen, Deutschland) und dem London Clay von Walton-on-the-Naze (Essex, England). Senckenbergiana lethaea. 78(1–2):157–177. doi:10.1007/BF03042767.
- Mayr G, De Pietri VL, Scofield RP. 2022. New bird remains from the early Eocene Nanjemoy Formation of Virginia (USA), including the first records of the Messelasturidae, Psittacopedidae, and Zygodactylidae from the Fisher/Sullivan site. Historical Biology. 34(2):322–334. doi:10.1080/08912963.2021.1910820.
- Mayr G, Manegold A. 2006. A small suboscine-like passeriform bird from the early Oligocene of France. Condor. 108(3):717–720. doi:10.1093/condor/108.3.717.
- Mayr G, Wilde V. 2014. Eocene fossil is earliest evidence of flower-visiting by birds. Biol Lett. 10(5):20140223. doi:10.1098/rsbl.2014.0223.
- Mayr G, Zelenkov N. 2009. New specimens of zygodactylid birds from the middle Eocene of Messel, with description of a new species of Primozygodactylus Mayr, 1998. Acta Palaeontologica Polonica. 54(1):15–20. doi:10.4202/app.2009.B103.
- Nixon KC. 2002. WinClada, version 1.00.08 [Computer software]. Ithaca (NY): Published by the author.
- Prum RO, Berv JS, Dornburg A, Field DJ, Townsend JP, Lemmon EM, Lemmon AR. 2015. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature. 526(7574):569–573. doi:10.1038/nature15697.
- Rayner D, Mitchell T, Rayner M, Clouter F. 2009. London Clay fossils of Kent and Essex. Rochester (Kent): Medway Fossil and Mineral Society.
- Sangster G, Braun EL, Johansson US, Kimball RT, Mayr G, Suh A. 2022. Phylogenetic definitions for 25 higher-level clade names of birds. Avian Res. 13:100027. doi:10.1016/j.avrs.2022.100027.
- Simpson SF, Cracraft J. 1981. The phylogenetic relationships of the Piciformes (Class Aves). Auk. 98:481–494.
- Smith NA, DeBee AM, Clarke JA. 2018. Systematics and phylogeny of the Zygodactylidae (Aves, Neognathae) with description of a new species from the early Eocene of Wyoming, USA. PeerJ. 6:e4950. doi:10.7717/peerj.4950.
- Weidig I. 2010. New birds from the lower Eocene Green River Formation, North America. Rec Austral Mus. 62(1):29–44. doi:10.3853/j.0067-1975.62.2010.1544.