ABSTRACT
Diverse chancelloriids from two sections of the Kaili Biota (Cambrian Wuliuan Stage) in Guizhou Province, China, are systematically described. A total of 25 complete individuals were collected from calcareous silty mudstones of the Cambrian Kaili Formation and are assigned to 3 genera and 6 species, including Archiasterella anchoriformis sp. nov., Chancelloria zhaoi sp. nov., C. eros, Allonnia erjiensis, Al. phrixothrix, and Al. sp. The new species Ar. anchoriformis with sclerites characterised by a large angle between two marginal-lateral rays and an obvious apical tuft represents the first unambiguous Archiasterella scleritome in South China. The C. zhaoi is dominated by a series of bilaterally symmetrical, rosette-like sclerites that composed of five or six lateral rays and a central ray. Moreover, based on a careful survey of the spatial-temporal distribution of chancelloriids (including both scleritome and isolated sclerite fossils) in South China, two flourishing ages, though may be related to preservational bias, of this metazoan group are recognised: 1) a significant diversification in the upper Stage 2 to Stage 3 and 2) a thriving in the Wuliuan Stage.
Introduction
Chancelloriids are a group of enigmatic animals that likely represent a stem group Eumetazoa (Janussen et al. Citation2002; Bengtson and Collins Citation2015; Yun et al. Citation2021). This animal is characterised by a radially symmetrical body, a unique orifice, a sessile lifestyle, and a flexible integument covered with dozens of composite hollow sclerites (Bengtson and Hou Citation2001; Janussen et al. Citation2002; Randell et al. Citation2005; Moore et al. Citation2014; Bengtson and Collins Citation2015; Yun et al. Citation2018, Citation2019a) that were formed with fibrous aragonites (James and Klappa Citation1983; Bengtson et al. Citation1990; Mehl Citation1996, Citation1998; Porter Citation2008; Yun et al. Citation2021).
The first appearance of chancelloriids may be traced back to Cambrian Precambrian–Cambrian transition (Zhu et al. Citation2017). From then on, they have experienced modest diversification and eventually went extinct in the late Cambrian (e.g. Mostler and Mosleh-Yazdi Citation1976; Bengtson and Hou Citation2001; Janussen et al. Citation2002; Paterson et al. Citation2016; Hong et al. Citation2016; Yun et al. Citation2019a). Isolated sclerites of chancelloriids are universally reported in early and middle Cambrian ‘SSFs’ (Small Shelly Fossils), while scleritomes and complete specimens have been recorded in the Lagerstätten due to extraordinary taphonomic events (e.g. Utah: Rigby Citation1978; Janussen et al. Citation2002; Canada: Randell et al. Citation2005; Harvey and Butterfield Citation2011; Bengtson and Collins Citation2015; South China: Chen and Zhou Citation1997; Hou et al. Citation1999; Bengtson and Hou Citation2001; Zhao et al. Citation2005; Zhao et al. Citation2018). However, the exact phylogenetic position of this animal group is still to be discovered.
Articulated chancelloriids had a cosmopolitan distribution in Konservat-Lagerstätten ranging from Cambrian Stage 3 to Drumian (Chen and Zhou Citation1997; Bengtson and Hou Citation2001; Bengtson and Collins Citation2015; Paterson et al. Citation2016; Cong et al. Citation2018; Yun et al. Citation2018; Zhao et al. Citation2018, p. 2019a). Based mainly on the structure, density, arrangement and symmetry of sclerites, 14 species and 5 genera have been distinguished. Almost all species of the genus Allonnia are characterised by sclerites composed of two lateral rays and one ascending ray. Whereas in the genus Archiasterella, the number of lateral rays is not less than three in sclerites. Chancelloria can be confidently discriminated from other genera by the presence of the central ray in a sclerite (Bengtson and Collins Citation2015; Yun et al. Citation2018, Citation2019a). In addition to the common types mentioned, the sclerites of Nidelric and Dimidia are formed with single ray and double rays, respectively (Hou et al. Citation2014; Zhao et al. Citation2018; Yun et al. Citation2019a; Yun et al. Citation2023).
In Kaili Biota, chancelloriids were only cursorily reported and not fully illustrated (Zhao et al. Citation2005, Citation2011). Based on new materials and restudy on the reported specimens, the underrated diversity and taxonomy of the Kaili chancelloriids are revealed. In addition to the previously reported C. eros and a poorly preserved Allonnia sp. (misinterpreted as Archiasterella sp.), there are four species of three genera discovered herein. With these Wuliuan taxa, the biogeographic and biostratigraphic distribution patterns of chancelloriids in South China are systematically summarised.
Geological setting, materials and methods
The Kaili Formation is a clastic shelf succession belongs to the narrow Jiangnan Slope basin of the south China during the Cambrian period (Yuan et al. Citation2002; Lin Citation2009; Gaines et al. Citation2011) and crops out in eastern Guizhou Province. Palaeogeographically, the Cambrian Kaili Biota is located along the southeastern margin of the Yangtze Platform (, 1B). It is perfectly exposed near the Balang village, Jianhe County of Guizhou, where the Kaili Formation at the Wuliu-Zengjiayan section () was recently ratified as the GSSP (Global Stratotype Section and Point) for the Cambrian Miaolingian Series and Wuliuian Stage (Zhao et al. Citation2019).
Figure 1. Location and geological setting of the studied area. A, map showing the location of the research area (red star) in the Yangtze Platform of South China. B, lithofacies reconstruction of South China during the Cambrian Miaolingian (after Feng et al. Citation2004; Zhang et al. Citation2008) with the studied area indicated by a red star. C, geological map of the Balang area and location of the Wuliu-Zengjiayan, Miaobanpo sections and the Jinyinshan quarry (modified from Zhao et al. Citation2019). D, stratigraphical column of the Kaili Formation at the Miaobanpo section, with fossil horizons indicated (after Yang et al. Citation2016).
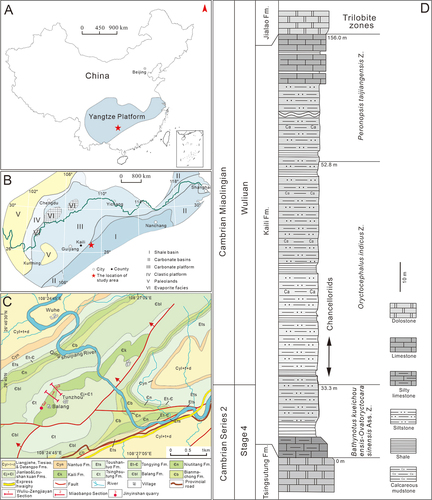
The Kaili Formation in the Wuliu-Zengjiayan section and the adjacent Miaobanpo section conformably overlies the Tsinghsutung Formation (a succession of thinly bedded dolostones) and is overlain by the Jialao Formation (a mixture of siliciclastic and carbonate rocks). The thickness of the formation varies between sections, 214 m in the Wuliu-Zengjiayan section and 156 m in the Miaobanpo section (, 1D) where its lowermost part is cut by a small fault (Zhao et al. Citation2005). Lithologically, the Kaili Formation is subdivided into three parts, the lower part comprises thinly bedded limestones with shale interbeds and belongs to Cambrian Stage 4, the middle part consists of calcareous silty mudstones and shales , belonging to the early of the Cambrian Wuliuian (within the trilobite Oryctocephalus indicus zone; Zhao et al. Citation2011, Zhao et al. Citation2019), and the upper part consists of bedded limestone layers grading into grainy dolostones (). The middle part of the Kaili Formation contains a Burgess Shale-type Lagerstätte that known as the Kaili Biota. The biota occurs slightly above the FAD of Oryctocephalus indicus, its species diversity and abundance are dominated by panarthropods, other major groups include algae, sponges, chancelloriids, cnidarians, ‘worms’, medusiform fossils, brachiopods, molluscs, echinoderms and various problematic fossils (Zhao et al. Citation2005, 2011).
A total of 25 chancelloriid fossils in this study were collected from the calcareous silty mudstone beds in the middle part of Kaili Formation at the Miaobanpo section and the nearby Jinyinshan quarry (), corresponding to the lower part of the trilobite Oryctocephalus indicus Zone (). There are 20 specimens from the Miaobanpo section, and the rest of 5 were from the Jinyinshan quarry.
Most chancelloriid is preserved as pyritised scleritomes (, 7B, 7D, 7E; 8) and only one specimen is as carbonaceous compression (). Some fossils sometimes occur in small clusters (, 6A, 8A) in which individual vary from size and shape, possibly suggesting in situ preservation. Some individuals, in addition, are attached onto brachiopods (, 4B), spherical structures (; less than 1 mm in diameter, spherical or sub-spherical, black, irregularly arranged in string, hollow in the centre and containing miniature quartz crystals and iron-stained powders (Lin et al. Citation2006).), hyolithoids (), and even other chancelloriids (, 8A). This is not the only case that chancelloriids attached to other benthos, some Burgess specimens, were also revealed as attaching to sponges and brachiopods (Bengtson and Collins Citation2015).
Figure 2. Terminology of chancelloriid sclerites. The lower right corner shows the schematic diagram of angle measurement between two marginal-lateral rays (modified from Bengtson and Collins Citation2015 and Yun Citation2019).
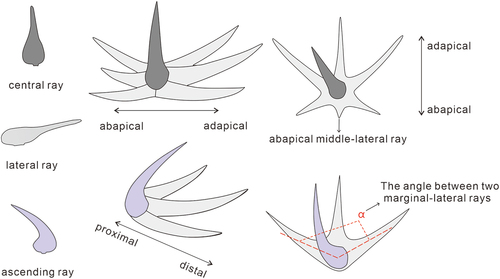
Figure 3. Archiasterella anchoriformis sp. nov. from the Kaili Biota. A, the fossil sketch drawing of B, dotted lines represent the indistinctive sclerites. B, the holotype scleritome (MBP-38); the white arrow points to an ascending ray of the sclerites. C, the sclerites of B (white arrow) and the proximal part of sclerites slightly swells. D, overall morphology, specimen no. MBP-43. E, overall morphology, specimen no. MBP-34. F, middle part of the body, specimen no. MBP-44. G, the apical tuft of MBP-43, is the countpart of D. H, J-K, the sclerites of D, E and F respectively, white arrow points to the ascending rays. I, the collapse of rays of D. Scale bars: A-B, D-F = 2 mm; C, G-H, J-K = 0.5 mm; I = 0.2 mm.

Figure 4. Archiasterella anchoriformis sp. nov. from the Kaili Biota. A, specimen no. JYS-1074. B, JYS-1074-1 of A (the lower left), 2 + 1A sclerites is present (red frame). C, JYS-1074-2 of A (the upper right). D, the specimen preserved from upper direction of A (the lower right), uncertain its type. E, detail of B (position marked by white frame). Scale bars: A = 5 mm; B-C = 2 mm; D = 1 mm; E = 0.5 mm.

All specimens illustrated here are housed in College of Resources and Environmental Engineering, Guizhou University, Guiyang, China. Some were prepared using a needle under a binocular microscope, for revealing parts covered by sediments. Digital photographs were taken by using a Canon 5D Mark Ⅳ camera with auxiliary light to increase the colour contrast of the morphological structures. Images and line drawings were made by using Coreldraw 19, Adobe Photoshop 2015.5 and image J software.
The morphological terms of the chancelloriid scleritome in Bengtson and Collins (Citation2015) and sclerite formula in Yun et al. (Citation2019b) are followed when describing materials. Structure of the sclerite is formulated as m+ nC and m+ nA, where m is the number of lateral rays, n is the number of C (central rays, only in genus Chancelloria) or A (ascending rays, applying to genera Archiasterella and Allonnia) (Yun et al. Citation2019b). MBP and JYS in specimen catalogue number indicate that the fossils came from the Miaobanpo section and the Jinyinshan quarry, respectively; and A and B denote part and counterpart of the fossil.
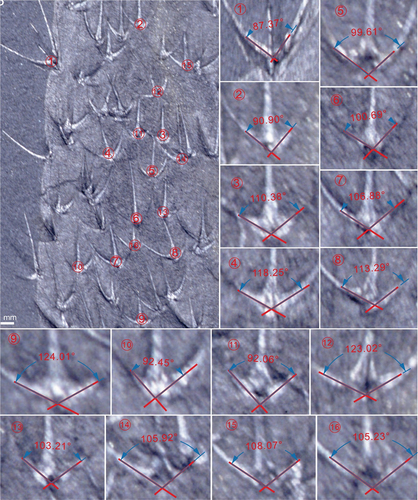
Additionally, the angle between lateral rays of the sclerites is an important feature in the classification of chancelloriids. In order to avoid unreliable results, three criteria are suggested in the measurement (): 1) the site for measurement should be close to the proximal part of lateral rays; 2) measuring angles between the central axis of the two lateral rays; 3) taking the mean values of the angles.
Figure 5. Chancelloria zhaoi sp. nov. from Kaili Biota. A, the fossil sketch drawing of B, dotted lines represent the uncertainty of sclerites. B, the holotype scleritome (MBP-42-1). C, apical tuft and orifice of B, the white arrow point to a stoutly composite sclerites. D, the paratype scleritome (MBP-46-3). E, 6 + 1C and 5 + 1C sclerites of C. zhaoi sp. nov., the length of ‘adr’ (adapical lateral ray) is longer than ‘abr’ (abapical lateral ray). Scale bars: A-B, D = 1 mm; C = 0.5 mm.

Systematic palaeontology
Phylum and Class uncertain
Order Chancelloriida Walcott Citation1920
Family Chancelloriidae Walcott Citation1920
Genus Archiasterella Sdzuy Citation1969
Type Species
Archiasterella pentactina Sdzuy Citation1969 (based on isolated sclerites).
Other representative species
Ar. tetraspina Vassiljeva and Sayutina 1993; Ar. fletchergryllus Randell et al. Citation2005; Ar. coriacea Bengtson and Collins Citation2015.
Archiasterella anchoriformis sp. nov.
Derivation of name
From the Latin ‘anchora’ (anchor) and ‘formis’ (shape), in reference to the shape of sclerites closely resembles an anchor.
Holotype
MBP-38A, B, from the Wuliuan Stage of Cambrian, Kaili Formation, Miaobanpo section, Kaili, Guizhou Province, China, housed in Institute of Palaeontology, College of Resources and Environmental Engineering, Guizhou University.
Paratypes
MBP-43A, B, from the same fossil locality as above.
Other Materials
MBP-34, , MBP-44A, B, and MBP-50, from the Miaobanpo section; JYS-1072A, B, JYS-1074 A, B-1, and JYS-1074 A, B-2, from the Jinyinshan quarry of the Kaili Biota.
Diagnosis
Species of Archiasterella having a spindle-shaped body with 3 + 1A sclerites and rare 2 + 1A sclerites, which are characterised by a bilateral symmetry, and the angle between two marginal-lateral rays is usually more than 120°. The apical orifice is surrounded by densely distributed slender, spine-shaped sclerites that form a teepee-like apical tuft.
Description
The overall shape of the holotype (, 3B) is spindle-shaped, ca 15.4 mm high, with a flat apical part (ca 2.6 mm wide), an inflated middle part (5.7 mm maximum width), and a narrow basal end (ca 1 mm wide). The apical orifice is surrounded by a dozen of sclerites consisting of modified spine-shaped rays (0.5–1.7 mm long, ca 0.025 mm wide), and the contracted basal end anchors to a brachiopod fragment. The sclerite formula of the specimen is 3 + 1A, including three lateral rays and one ascending rays (as indicated by the white arrow in ). All rays apically directed, and the lateral rays close the body surface while ascending rays curved away from the body surface. Remains of the basal part of the ascending ray are preserved. The proximal part of sclerites slightly swells and the distal part keeps original size (, ca 0.09 mm width). The middle-lateral rays are straight, while the two marginal-lateral rays curve towards each other within the plane formed by lateral rays. Therefore, the sclerites is bilaterally symmetrical and look like an anchor. At least 35 sclerites are irregularly arranged on the body. The separation of sclerites is inhomogeneous, ranging from 0.6 to 1.4 mm; the orientation of some sclerites in different positions of the body are a little deviate from the apical part. There is no obvious size variation of the sclerites throughout the scleritome.
The paratype MBP-43 () is 36.5 mm high, showing a ‘lamp bulb-shaped’ outline that includes a widest upper part (ca 16.5 mm wide) and a cylindrical lower part (ca 11.2 mm) with a rounded basal end. The counterpart of this specimen reveals a complete apical tuft composed of densest spine-shaped sclerites (, ca 6.1 mm high and 7.6 mm wide) when compared to other chancelloriids. The distal parts of these spine-shaped sclerites (about 0.1 mm in diameter) are closely converged to the central axis of the body and firmly shelter the apical orifice. The sclerites near the apical area (at least 3.4 mm in length and 0.3 mm in width of middle lateral ray) are bigger than the sclerites in the lower part (at least 1.4 mm length and 0.2 mm width of middle lateral ray). The marginal-lateral rays in this specimen form a 137° angle between each other ().
The specimen MBP-34 is of medium size, 28 mm high and 13.8 mm wide, (). The density of spine-shaped sclerites of apical tuft is between MBP-43 and MBP-38 (holotype). The basal part is incomplete and covered by sediments. The size and irregular arrangement pattern of sclerites is similar to that of the holotype. At the proximal part of the composite sclerites (the contact position of adjacent rays), four rays (three lateral rays and one broken ascending ray) in the plane can be obviously observed ().
A similar characteristic of sclerites can be seen in the incomplete specimen (MBP-44, ), which is 22.5 mm high and 8.4 mm wide. The angle between marginal-lateral rays of the sclerites is about 120°(), larger than that of the holotype. Size and arrangement of sclerites are also irregular.
The specimen JYS-1074-1 () is preserved in the large slab JYS-1074 (, 54 × 35 mm2), which was buried together with other individuals, including the specimen JYS-1074-2 (, see below) and a specimen originally compressed from top ().
The specimen JYS-1074-1 is of the smallest size of Ar. anchoriformis sp. nov., which has a baglike body 10.1 mm high and 5.5 mm wide (at the upper part of the body). The widths of the apex and the contractive basal end are about 4.0 mm and 1.4 mm, respectively. Like other specimens, it has a conspicuous apical tuft, a series of 3 + 1A sclerites with large angle between the marginal-lateral rays (ca 130°, ). The basal end is attached to a brachiopod fragment. The sclerites at the lower part of the body are smaller, more densely arranged than the upper part (from the lower to upper part, the ray length is from 0.8 mm to 1.3 mm and the intervals of sclerites from 0.5 mm to 1.4 mm). A few small 2 + 1A sclerites are scattered on the body margin (the lateral rays form an angle of 34°–53°).
The specimen JYS-1074-2 () has a cylindrical body with a slightly contracted middle part. The height is 17.9 mm, the maximum width is 5.2 mm, the minimum width 2.98 mm (at the middle part). Dozens of sclerites are irregularly arranged in the scleritome. The sclerites are discerned as two forms: 3 + 1A and rare 2 + 1A.
Comparison. In accordance with the typical sclerite formula (lateral rays + one ascending ray, the number of lateral rays is mostly three or more), this species belongs to the genus Archiasterella (Yun et al. Citation2019b). There are at least five other species characterised by 3 + 1A sclerites, including Ar. tetraspina (= ?Ar. tetractina Duan Citation1984), Ar. cometensis Moore et al., 2019, Ar. uncinata Moore et al., 2019, Ar. hirundo Bengtson in Bengtson et al. Citation1990, and Ar. coriacea Bengtson and Collins Citation2015. Although this genus is widely distributed all over the world, most of them are represented by the isolated sclerites instead of complete individuals. To date, except for the Ar. anchoriformis described herein, only Ar. coriacea was established based on exceptionally preserved scleritome materials, and it has peculiar integument with a scaly surface pattern and an inconspicuous apical tuft (Bengtson and Collins Citation2015). As mentioned above, the average angle between the marginal-lateral rays in the sclerites of Ar. anchoriformis is is generally greater than that of Ar. coriacea (see Annex , , , ). Moreover, the apical tuft formed by dozens of single-rayed sclerites is more conspicuous in Ar. anchoriformis.
Genus Chancelloria Walcott Citation1920
Type Species
Chancelloria eros Walcott Citation1920 from the Burgess Shale (based on scleritome).
Other representative species
C. cruceana Rusconi Citation1954; C. pentacta Rigby Citation1978; C. irregularis Qian Citation1989; C. australilonga Yun et al. Citation2019a.
Chancelloria zhaoi sp. nov.
()
Derivation of name
To commemorate professor Zhao Yuanlong of Guizhou University for discovering and extraordinary contributions on the Kaili Biota.
Holotype
MBP-42, from the Wuliuan Stage of Cambrian, Kaili Formation, Miaobanpo section, Kaili, Guizhou Province, China, housed in the Institute of Palaeontology, College of Resources and Environmental Engineering, Guizhou University.
Paratype
MBP-46, from the Miaobanpo section of the Kaili Biota.
Diagnosis
Saclike body mainly composed of 5 + 1C and 6 + 1C sclerites with typical bilateral symmetry. The lengths of adapical and abapical lateral rays are significantly different in the same sclerite. Generally, the size and density of sclerites are almost unchanged throughout the scleritome, and the arrangement is irregular. The apical tuft is conspicuous.
Description
The holotype (MBP-42, ) has a saclike body about 11.2 mm high. The abapical part tapers down to a blunt end and the apex is encircled by a series of sparsely distributed slender single-rayed sclerites (). The rays of these sclerites are around 0.04 mm in diameter while the thickest one is up to 0.2 mm (, marked by a white arrow). The width of both ends of the body is similar, and the widest part is at the upper part (at least 4.2 mm wide). The sclerites of C. zhaoi sp. nov. are bilaterally symmetrical and can be classified into two structures: 6 + 1C and 5 + 1C (), in which the central rays are mostly broken off and not preserved. In the 6 + 1C sclerites, abapical lateral rays and adapical lateral rays are both three in number. The shortest ray in a sclerite is the abapical middle-lateral ray. The structure of the 5 + 1C sclerites is similar to that of the 6 + 1C sclerites in a lacking of a abapical middle-lateral ray. The density and size of sclerites are almost unchanged throughout the whole body.
The paratype MBP-46 () is incomplete, with a hemispherical body that is 3.6–6.5 mm wide. In this scleritome, a small number of composite sclerites are recognisable, which reveal a 5 + 1C structure and a bilateral symmetry.
Comparison
To date, the species of Chancelloria based on scleritome materials include C. eros Walcott Citation1920, C. cruceana Rusconi Citation1954, C. pentacta Rigby Citation1978, and C. australilonga Yun et al. Citation2019a (Walcott Citation1920; Rusconi Citation1954; Rigby Citation1978; Beresi and Rigby Citation2013; Bengtson and Collins Citation2015; Yun et al. Citation2019a). These species, and C. zhaoi described herein, all have 5 + 1C and 6 + 1C sclerites while the proportion and detailed structures of these sclerites vary. As described above, the sclerites of C. zhaoi have two groups of lateral rays (three adapical and two or three abapical lateral rays) with nearly opposite orientations. The conspicuous apical tuft and sclerites formed by three groups of lateral rays in different directions (including two subparallel, remarkably elongate adapical rays) and a bilateral symmetry characterise C. australilonga (Yun et al. Citation2019a). The C. pentacta has two adapical lateral rays, two horizontal lateral rays and an abapical lateral ray, in which the horizontal lateral rays extend outward with ray tips curving slightly upwards (Rigby Citation1978). There are various types of sclerites in C. eros (including 5–8 + 1C and 3–4 + 0), most of which are asymmetrical or radial symmetrical (Walcott Citation1920; Bengtson and Collins Citation2015). With regard to C. cruceana, the general structure of the sclerites is similar to that of C. zhaoi, however, the complete scleritome of this species is not preserved and there were only 6 + 1C sclerites ever discovered. In addition, the bilateral C. irregularis Qian Citation1989 based on isolated sclerites is similar to some of the sclerites in C. zhaoi, however, it is impractical to reconstruct the arrangement pattern of the former.
Chancelloria eros Walcott Citation1920
()
1920 Chancelloria eros; Walcott, partim; pls 86:2, 2a, 2b, ?2c, 88:1 f, non pl. 88:1,1a-e.
1973 Chancelloria eros; Goryansky, p. 43.
1978 Chancelloria eros; Rigby, pl. 2, 3.
2002 Chancelloria eros; Janussen et al., .
2005 ?Chancelloria cf. eros; Randell et al., .
2015 Chancelloria eros; Bengtson and Collins, –11, 13–14.
Materials
A slab MBP-41 (36 × 13 mm2, ) from the Miaobanpo section of the Kaili Biota, containing an association of 4 specimens (MBP-41-1, MBP-41-2, MBP-41-3, and MBP-41-4).
Description
The specimen MBP-41-1 (Bottom of ) has a cylindrical body that is about 16.7 mm high and 6.4 mm wide. Both ends are rounded, and the minimum width is 2.5 mm. More than 50 sclerites are irregularly arranged, while the density is generally uniform on the body wall. Sclerites in the studied specimens are mainly 6 + 1C in structure (), with a few 5 + 1C () and 7+1C (), and others poorly-preserved. They are almost all asymmetrical and less common in radial symmetry, and all rays are slightly curved.
The specimen MBP-41-2 () is adjacent to the upper right margin of MBP-41-1, and the size of body is about a third of the latter. The sclerites are mainly 6 + 1C forms, and their sizes gradually decrease downward the body (except for the sclerite pointed by the white arrow whose length is up to 1.7 mm).
Figure 6. Chancelloria eros from the Kaili Biota. A, including four specimens of Chancelloria eros, specimen no. MBP-41. The specimen with the largest size is MBP-41-1. B, the second specimen (MBP-41-2) from the bottom of A, the white arrow indicates that the sclerite has a longer lateral ray. C, the third specimen (MBP-41-3) from the bottom of A, the white arrow stands for the possibility of stalk structure, and the red arrow indicates that the basal part of MBP-41-3 probably anchored to the top of MBP-41-2. D, the fourth specimen (MBP-41-4) from the bottom of A. E-G, 6 + 1C, 5 + 1C and 7 + 1C sclerites of the first C. eros from the bottom of A. Scale bars: A = 5 mm; B-D = 2 mm; E-G = 0.5 mm.

Figure 7. Allonnia from Kaili Biota. A-D, F, Allonnia erjiensis. A, the fossil sketch drawing of B, dotted lines represent the uncertainty of sclerites. B, MBP-35. C, MBP-45. D, MBP-37. E, G, Allonnia phrixothrix, specimen no. MBP-40. F, detail of D (position marked by white frame). G, integument detail of E; ‘lr’, lateral ray; ‘asr’, ascending ray. Scale bars: A-E = 2 mm; F, G = 0.1 mm.

The scleritome (MBP-41-3) () shows a mushroom-shaped body that is the smallest (2.48 mm high and 0.95 mm wide at its broadest area) when compared to other specimens in the slab MBP-41 (). The body is divided into two parts: 1) a narrow basal part (no more than 0.15 mm in width) and probably representing a contracted stalk (indicated by a white arrow in ) probably anchored to the top of another specimen (MBP-41-2, indicated by a red arrow in ), and 2) a bulged upper part with a series of composite sclerites.
The specimen MBP-41-4 () at the upper end of the slab also has a small body size and a similar association of 6 + 1C, 7 + 1C and 5 + 1C sclerites. But the length of the rays is relatively longer (up to 3.1 mm long) than that of the MBP-41-2.
Comparison
With indistinctive apical tuft and diverse forms of sclerites that are dominated by asymmetrical or radial symmetrical of structures, C. eros is distinctive from other Chancelloria species.
Genus Allonnia Doré and Reid Citation1965
Type Species
Allonnia tripodophora Doré and Reid Citation1965 (based on isolated sclerites).
Other representative species
Al. phrixothrix Bengtson and Hou Citation2001; Al. tintinopsis Bengtson and Collins Citation2015; Al. erjiensis Yun et al. Citation2018; Al. nuda Cong et al. Citation2018; Al. tenuis Zhao et al. Citation2018.
Allonnia erjiensis Yun et al. Citation2018
2018 Allonnia erjiensis sp. nov; Yun et al., figs. 1
2019 Allonnia erjiensis; Yun, figs. 4.2D, 4.3E, 4.4 H-J, 9.6
Materials
MBP-35, MBP-45, MBP-37A, B, and MBP-48, from the Miaobanpo section; JYS-874, from the Jinyinshan quarry of the Kaili Biota.
Description
The specimen MBP-35 (, 7B) is slim rod-shaped, 19.9 mm high. The body was divided into equal two parts. The lower part forms a cylindrical stalk, 10.1 mm long and 1.4 mm wide, with dense and short sclerites. The upper part is spindle-shaped, 9.8 mm long and 4.8 mm broad at the widest area, with sclerites not denser than the lower part but composed of long-conical rays. The apical tuft is not preserved. The base is attached to some spherical structures (whether they are organism remains or sedimentary structures is still to be discussed). The 2 + 1A sclerites are predominated on the body surface, with ascending rays visible only in the marginal parts of the body due to their outward-pointing position (Yun et al. Citation2018) (, 7B). The length of the rays increases upward from 1.4 mm to 6.9 mm.
The specimen MBP-45 () shows a slightly twisted and club-shaped body, about 10.6 mm high. The body width gradually increases from the base (0.6 mm wide) to the apex (ca 2.8 mm wide). The basal end anchored to a hyolithid conch, yet the upper part is incompletely preserved. The size change of sclerites is less evident than that of MBP-35, with ray length between 0.6 mm and 1.8 mm. Moreover, this specimen is dark grey in colour, indicating the preservation of carbonaceous films. The sclerites are 2 + 1A in structure, with ascending rays remarkably protruding out the body surface.
The specimen MBP-37 () shows a narrow, incomplete body, which is 10.8 mm high and 1.8 mm wide. The basal end attaches to a dark spherical structure (similar to MBP-35). The sclerites of the upper part are longer than those of the lower part, and the length of rays ranges from 3.7 mm to 7.3 mm. In consideration of the poor preservation, the sclerite structure is not well revealed, however, some ascending rays are discernable.
Comparison
The sclerites of Al. erjiensis is the densest of all known Allonnia species, and covers the whole body surface. Other species, such as Al. nuda, Al. tenuis, Al. phrixothrix, Al. tintinopsis, usually have seldom sclerites in the basal area (Bengtson and Hou Citation2001; Bengtson and Collins Citation2015; Cong et al. Citation2018; Zhao et al. Citation2018). Compared to the holotype, specimens described herein have a distinct basal end attaching to a hyolithid conch, and the body width is much narrower though the general heights are similar (ca 10 mm −15 mm) (Yun et al. Citation2018; Yun Citation2019).
Allonnia phrixothrix Bengtson and Hou Citation2001
()
2001 Allonnia phrixothrix sp. nov.; Bengtson and Hou, figs. 1, 2, 3, 4, 5, 7, 8, 11
2002 Allonnia junyuani sp. nov.; Janussen, Steiner and Zhu, figs. 5, 6.2
2019 Allonnia phrixothrix; Yun, figs. 4.2A, C, 4.3A–D, F, G, 4.4 A–G, 6.1, 8.1, 9.5
Material
MBP-40 from the Miaobanpo section of the Kaili Biota.
Description
shows an incomplete specimen, with a cylindrical, short body, 4.9 cm in high and 2.1 cm wide in the middle section. The upper part is ringed by sclerites composed of long rays and the apical tuft is not revealed. The lower part is flat. The soft integument is well preserved in reddish colour. There are obvious granular ornamentations (about 0.06 mm in diameter) on the integument (). Dozens of sclerites with a uniform size, shape and density are irregularly arranged in the scleritome. Each of sclerites has two lateral and one ascending rays (2 + 1A), and the rays are about 1.5 to 2.2 mm long and 0.08 mm wide. The lateral rays form a mutual angle of about 40°-50° and curve slightly towards each other, and the ascending ray leaves a pit or a tiny proximal part that were broken off due to the taphonomic processes.
Comparison
The sclerites of MBP-40 are strictly limited to the 2 + 1A form. The size, density, and structure of these sclerites are all in accordance with the holotype, except for the apical tuft.
Allonnia sp.
()
Material
MBP-47-1, MBP-47-2, MBP-47-3, MBP-49, MBP-39, from the Miaobanpo section of the Kaili Biota.
Description
Three incomplete specimens are preserved on the slab MBP-47 (25 × 19 mm2, ). The specimen MBP-47-1 (at the bottom right corner of ) shows a short, ellipsoidal body, 7.7 mm high and 3.3–4.1 mm wide. The sclerite structure is discerned as 2 + 1A and (possibly) 3 + 1A, with lateral rays about 0.7–1.6 mm long (the mutual angle of lateral rays is 35–60°). Some single-rayed sclerites are densely arranged parallel to each other, forming an apical tuft (about 1.6 mm long and 1.3 mm wide) (). The lower part of the body is not well preserved.
The specimen MBP-47-2 in the middle part of has a conical shape, a height of 5.6 mm and a width (upper part of the body) of 1.9 mm. The scleritome is poorly preserved, but the lateral rays of 2 + 1A sclerites are discernable. The length of the rays dramatically increase from the lower to upper part (1.2 mm to 5.1 mm). The lower part contracts abapically to a basal end that probably attaches to the upper part of another specimen (MBP-47-3) (). The specimen MBP-47 in the upper left part of is even more poorly preserved that only a few of 2 + 1A sclerites with remains of the ascending rays can be observed ().
Other specimens with probable 2 + 1A sclerites are shown in (MBP-39) and (MBP-49). They have similar body shape and size (about 7–8 mm). The sclerite rays on the upper part of the body are slender than that in the three specimens described above.
Discussion
A diverse chancelloriid assemblage in the Kaili Biota
Well-preserved fossil specimens reveal a relatively high taxonomic diversity of chancelloriid assemblage in the Kaili Biota (). Totally three genera and six species are recognised, including Archiasterella anchoriformis sp. nov., Chancelloria zhaoi sp. nov., C. eros, Allonnia erjiensis, Al. phrixothrix, and Al. sp. At the genus level of abundance, the assemblage is dominated by Allonnia (44% of the total), of which Al. erjiensis and Al. sp. occupy 45.45% of the whole genus, followed by Al. phrixothrix. In addition, Archiasterella, which accounts for a small proportion of the three genera and probably has a narrow geographic distribution, has become the most abundant type in the Kaili Biota at the species level (32% of the total). The abundance of Chancelloria is modest by comparison (24% of the total), and C. eros and C. zhaoi account for 66.7% of 33.3% of the whole genus, respectively.
Figure 8. Allonnia sp. from Kaili Biota. A, three specimens in different direction, specimen no. MBP-47. The white box indicates the basal part of MBP-47-2 probably anchored to the top of MBP-47-3. B, apical tuft of the lower right corner of A. C, sclerites detail of the upper left corner of A. D, MBP-39. E, MBP-49; ‘lr’, lateral ray; ‘asr’, ascending ray. Scale bars: A = 5 mm; B, C = 0.2 mm; D, E = 2 mm.

Figure 9. The reconstruction of chancelloriids in Kaili Biota (By Liu X). The labels 1 to 5 indicate Allonnia erjiensis, Chancelloria eros, C. zhaoi sp. nov., Archiasterella anchoriformis sp. nov., and Al. phrixothrix, respectively.

Previously, well-preserved complete scleritomes of Chancelloria were only discovered in the Burgess Shale (Walcott Citation1920; Bengtson and Collins Citation2015), Little Bear Biota (Butterfield and Nicholas Citation1996), and Wheeler Formation (Janussen et al. Citation2002) of Laurentia and Emu Bay Shale of East Gondwana (Yun et al. Citation2019a). The scleritomes of Archiasterella are even rarer and were only reported from the Burgess Shale (Bengtson and Collins Citation2015) and Sekwi Formation (Randell et al. Citation2005) of Laurentia. With regard to the Chengjiang and Guanshan biotas of South China, there are only Allonnia, Dimidia, and Nidelric ever discovered (Bengtson and Hou Citation2001; Hou et al. Citation2014; Cong et al. Citation2018; Zhao et al. Citation2018; Yun et al. Citation2018, Yun et al. Citation2023). Therefore, Ar. anchoriformis, C. zhaoi, and C. eros of the Kaili Biota represent the first occurrence of complete bodies of Chancelloria and Archiasterella in South China.
Spatial-temporal distribution of chancelloriids in South China
Chancelloriids were a globally distributed group which had lasted at least 45 million years (Cambrian Fortunian to Jiangshanian stages) (Yun Citation2019). In South China, almost all common chancelloriid genera were discovered, including Chancelloria, Allonnia, Archiasterella, Dimidia, Nidelric, and Cambrothyra (see the supplemental material. Yun Citation2019; Yun et al. Citation2019a). The earliest chancelloriids of South China were likely from the Kuanchuanpu Formation of the Fortunian Stage in southern Shaanxi Province, which are classified into Chancelloria and Cambrothyra (Fu Citation1983; Yang et al. Citation1983; Steiner et al. Citation2004). The sclerites of Chancelloria (associated with small shelly fossils) were discovered in a number of slightly younger localities such as Hubei (Qian et al. Citation1979; Qian Citation1989), Sichuan (Yin et al. Citation1980; Li Citation1999), and Yunnan (Luo et al. Citation1982; Moore et al. Citation2014) provinces. From late Stage 2 on, chancelloriids experienced a significant diversification, with abundant sclerites and scleritomes (of at least six genera) distributed across the Yangtze block (Qian Citation1989; Qian et al. Citation1999). While after the Wuliuan Stage, chancelloriids appeared to suffer a setback and were in serious decline with only few records from Huaqiao Formation of Hunan Province (Zhu et al. Citation2003), and they apparently became extinct after Cambrian the Guzhangian Stage.
In summary, two flourishing ages of chancelloriids in South China are distinguished, though they may be related to preservational bias. The first one (Flourishing Age I) is from late Stage 2 to middle stage 3, and the second one (Flourishing Age II) is the Wuliuan Stage ().
Figure 10. Distribution of the chancelloriids in South China.The stratigraphical distribution, shows two flourishing ages of genus-level chancelloriids. References for different formation: Kuanchuanpu Fm. = Fu (Citation1983), Yang et al. (Citation1983) and Steiner et al. (Citation2004); Huangshandong Fm. = Qian et al. (Citation1979) and Qian (Citation1989); Maidiping Fm. = Yin et al. (Citation1980), Qian (Citation1989), Li (Citation1999), Feng et al. (Citation2022); Zhongyicun to Dahai Mbs. of Zhujiaqing Fm. and Shiyantou Fm. = Luo et al. (Citation1982), Parkhaev and Demidenko (Citation2010), Moore et al. (Citation2014); Middle Jiulaodong Fm. = Yin et al. (Citation1980) and Qian (Citation1989); Xihaoping Fm. = Qian and Zhang (Citation1983), Duan (Citation1984), Wang and Xu (Citation1987), Qian (Citation1989), Ding et al. (Citation1990), Li et al. (Citation2004), Steiner et al. (Citation2004), Moore et al. (Citation2010), Yang et al. (Citation2015), Zhang et al. (Citation2021) ; Lower Shuijingtuo Fm. = Li et al. (Citation2004) and Yang et al. (Citation2015); Qingjiang Biota = Fu et al. (Citation2019); Chengjiang Biota = Bengtson and Hou (Citation2001), Janussen et al. (Citation2002), Hou et al. (Citation2014), Yun et al. (Citation2018), Cong et al. (Citation2018), Sun et al. (Citation2020), Yun et al. (Citation2022); Upper Jiulaodong Fm. = Yin et al. (Citation1980); Middle Guojiaba Fm. = Yun et al. (Citation2021) ; Lower Mingxinsi Fm. = Qian (Citation1989); Guanshan Biota = Hu et al. (Citation2010, Citation2013), Chen et al. (Citation2019), Zhao et al. (Citation2018); Mufushan Fm. = Zhang (Citation1995); Balang Fm. = Liu and Lei (Citation2013); Kaili Biota = Zhao et al. (Citation1999, Citation2005, Citation2011), Harvey et al. (Citation2012), this study; Upper Damao Gp. = Jiang and Huang (Citation1986), Damao Group is distributed in Hainan and likely near to the Yangtze Block in the Cambrian; Huaqiao Fm. = Zhu et al. (Citation2003).
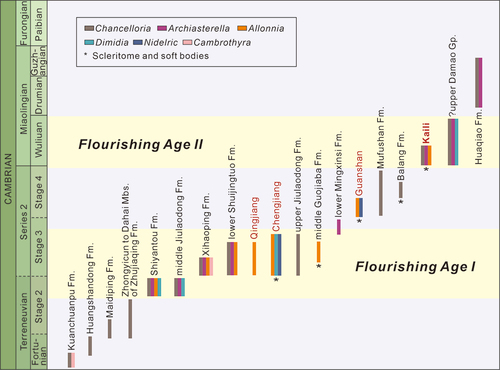
The Flourishing Age I is characterised by a rapid diversification of Archiasterella, Allonnia, Dimidia, and Nidelric following the widespread of Chancelloria. The genera Allonnia, Archiasterella, and Dimidia first appeared in the upper part of the Shiyantou Formation (eastern Yunnan) and middle member of Jiulaodong Formation (central Sichuan) of late Stage 2 (Luo et al. Citation1982; Qian Citation1989; Qian et al. Citation2002; Parkhaev and Demidenko Citation2010; Moore et al. Citation2014). As one of the globally distributed chancelloriids, Allonnia plays an important role in the diversity and abundance of chancelloriids from the well-known Chengjiang Biota. Dimidia, characterised by double-rayed sclerites, was previously regarded as a junior synonym of Allonnia, however, is discovered from scleritome fossils from the Chengjiang Biota (Yun et al. Citation2023). The genus Nidelric, dominated by single-rayed sclerites, is only discovered in the Chengjiang and Guanshan biotas of South China (Hou et al. Citation2014; Zhao et al. Citation2018).
The Flourishing Age II represents another thriving of diversity of the chancelloriid group. Although Cambrothyra and Nidelric likely went extinct after Stage 4, there were at least four genera, including Chancelloria, Allonnia, Dimidia, and Archasterella, flourished in the chancelloriid communities represented by the Kaili Biota (Zhao et al. Citation1999, Zhao et al. Citation2005, Zhao et al. Citation2011; Harvey et al. Citation2012; this study) and the upper Damao Group of Hainan Province (likely near to the Yangtze Block in the Cambrian Period; Jiang and Huang Citation1986).
Conclusions
New fossil materials from the Kaili Biota of South China have increased our knowledge on the diversity of chancelloriids during the Wuliuan Stage of Cambrian. Totally three genera and six species are recognised, in which the new species Archiasterella anchoriformis sp. nov. represents the only known complete body of the genus Archiasterella in South China. A careful survey on the spatial-temporal distribution of genus-level chancelloriids in South China reveals two flourishing ages of the group. The Flourishing Age I is late Stage 2 to Stage 3 that represented by the Chengjiang Biota and the Flourishing Age II is the Wuliuan Stage that represented by the Kaili Biota described herein. These flourishing ages can not only shed light on the biostratigraphic correlation of chancelloriid-bearing successions throughout South China, but also potentially reflect an evolutionary history.
Supplemental Material
Download MS Excel (11.9 KB)Acknowledgments
We thank the constructive suggestions from reviewers and the editor, which conduces to improve the quality of papers.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2023.2212382
Additional information
Funding
References
- Bengtson S, Collins D. 2015. Chancelloriids of the Cambrian Burgess Shale. Palaeontol Electron. 18:1–67.
- Bengtson S, Conway MS, Cooper BJ, Jell PA, Runnegar BN. 1990. Early Cambrian fossils from South Australia. Mem Assoc Australas Palaeontol. 9:1–368.
- Bengtson S, Hou XG. 2001. The integument of Cambrian chancelloriids. Acta Palaeontol Pol. 46(1):1–22.
- Beresi MS, Rigby JK. 2013. Middle Cambrian protospongiid sponges and chancelloriids from the Precordillera of Mendoza Province, western Argentina. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 268(3):259–274. doi:10.1127/0077-7749/2013/0329.
- Butterfield NJ, Nicholas CJ. 1996. Burgess Shale-type preservation of both non-mineralizing and ‘shelly’ Cambrian organisms from the Mackenzie Mountains, northwestern Canada. J Paleontol. 70(6):893–899. doi:10.1017/S0022336000038579.
- Chen FY, Zhang ZF, Betts MJ, Zhang ZL, Liu F. 2019. First report on Guanshan Biota (Cambrian Stage 4) at the stratotype area of Wulongqing Formation in Malong County, Eastern Yunnan, China. Geoscience Frontiers. 10(4):1459–1476. doi:10.1016/j.gsf.2018.09.010.
- Chen JY, Zhou G. 1997. Biology of the Chengjiang fauna. In: Chen J, Cheng YN, Van Iten HV, editors. The Cambrian Explosion and the Fossil Record. Bull Natl Mus Nat Sci. p. 11–105.
- Cong PY, Harvey THP, Williams M, Siveter DJ, Siveter DJ, Gabbott SE, Li YJ, Wei F, Hou XG. 2018. Naked chancelloriids from the lower Cambrian of China show evidence for sponge-type growth. Proc R Soc B. 285(1881):1–7. doi:10.1098/rspb.2018.0296.
- Ding LF, Qin HB, Li Y. 1990. Study on the boundary between Sinian and Cambrian in the south of Shaanxi Province, China. Professional Papers of Stratigraphy and Palaeontology. Beijing: Geological Publishing House, Vol. 23: 96–130. Chinese
- Doré F, Reid RE. 1965. Allonnia tripodophora nov. gen., nov. sp., nouvelle Eponge du Cambrien inférieur de Carteret (Manche) [Allonnia tripodophora nov. gen., nov. sp., new sponge of Carteret (Manche) in Lower Cambrian]. C.R. Sommaire Séances Soc. Géol. Fr. 20–21. French
- Duan CH. 1984. Small shelly fossils from the Lower Cambrian Xihaoping Formation in the Shennongjia District, Hubei Province—hyoliths and fossil skeletons of unknown affinities. Bulletin of the Tianjin Institute of Geology and Mineral Resources, Chinese. Academy of Geological Sciences. 7. 143–188. Chinese
- Feng Q, Pan B, Yang AH, Lu M, Li GX. 2022. Biostratigraphy of the Small Shelly Fossils from the Upper Maidiping Formation (Terreneuvian) at the Fandian Section, Sichuan Province, South China. Frontiers in Earth Science. 10:1–13. doi:10.3389/feart.2022.922439.
- Feng, ZZ, Peng YM, Jin ZK, Bao ZD. 2004. Lithofacies paleogeography of the Cambrian and Ordovician in China. Beijing: Petroleum Industry Press. Chapter 5, Cambrian and Ordovician lithofacies palaeogeography in South China; p. Chinese
- Fu JH. 1983. Discovery of small shelly fossils from Liangshan Country, Hanzhong Area and their Stratigraphic sighificance. Scientia Geol Sin. 4. 414–417. Chinese
- Fu DJ, Tong GH, Dai T, Liu W, Yang YN, Zhang Y, Cui LH, Li LY, Yun H, Wu Y, Sun A, Liu C, Pei WR, Gaines RR, Zhang XL. 2019. The Qingjiang biota—A Burgess Shale–type fossil Lagerstätte from the early Cambrian of South China. Sci. 363(6433):1338–1342. doi:10.1126/science.aau8800.
- Gaines RR, Mering JA, Zhao YL, Peng J. 2011. Stratigraphic and microfacies analysis of the Kaili Formation, a candidate GSSP for the Cambrian Series 2–Series 3 boundary. Palaeogeography, Palaeoclimatology, Palaeoecology. 311(3–4):171–183. doi:10.1016/j.palaeo.2011.08.018.
- Goryansky V.Yu. 1973. O neobkhodimosti isklyucheniya roda Chancelloria Walcott iz tipa gubok [On the necessity of excluding the genus Chancelloria Walcott from the sponge phylum]. Trudy Instituta Geologii I Geofiziki Sibirskoye Otdeieniye. 49. 39–44. Russian
- Harvey THP, Butterfield NJ. 2011. Great Canadian Lagerstätten 2. Macro- and Microfossils of the Mount Cap Formation (Early and Middle Cambrian, Northwest Territories). Geol Can. 38:165–173.
- Harvey THP, Ortega-Hernandez J, Lin J-P., Yuanlong Z., Butterfield NJ. 2012. Burgess Shale-type microfossils from the middle Cambrian Kaili Formation, Guizhou Province, China. Acta Palaeontologica Polonica. 57(2):423–436. doi:10.4202/app.2011.0028.
- Hong J, Choh SJ, Lee DJ. 2016. Distribution of chancelloriids in a middle Cambrian carbonate platform deposit, Taebaek Group, Korea. Acta Geologica Sinica English Edition. 90(3):783–795. doi:10.1111/1755-6724.12722.
- Hou XG, Jan Bergstöm, Wang HF, Feng XH, Chen AL.1999, The Chengjiang Fanua: exceptionally well-preserved animals from 530 million years ago. Yunnan Science and Technology Press Chinese.
- Hou XG, Williams M, Siveter DJ, Siveter D J., Gabbott S., Holwell D., Harvey T.H.P. 2014. A chancelloriid-like metazoan from the early Cambrian Chengjiang Lagerstätte, China. Sci Rep. 4(1):1–6. doi:10.1038/srep07340.
- Hu SX, Zhu MY, Luo HL, Steiner M, Zhao FC, Li GX, Liu Q, Zhang ZF. 2013. [Guanshan Biota]. Kunming: Yunnan Science and Technology Press. Chapter 5 (Biodiversity of the Guanshan Biota). p. 49–170. Chinese
- Hu SX, Zhu MY, Steiner M, Luo HL, Zhao FC, Liu Q. 2010. Biodiversity and taphonomy of the Early Cambrian Guanshan biota, eastern Yunnan. Science China Earth Sciences. 53(12):1765–1773. doi:10.1007/s11430-010-4086-9.
- James NP, Klappa CF. 1983. Petrogenesis of Early Cambrian reef limestones, Labrador, Canada. J sediment petrol. 53:1051–1096.
- Janussen D, Steiner M, Maoyan Z. 2002. New well preserved scleritomes of chancelloridae from the early Cambrian Yuanshan Formation (Chengjiang, China) and the middle Cambrian Wheeler Shale (Utah, USA) and paleobiological implications. J Paleontol. 76(4):596–606. doi:10.1666/0022-3360(2002)0760596:NWPSOC2.0.CO;2.
- Jiang ZW, Huang ZB. 1986. Middle Cambrian small shelly fanua in Yaxian country, Hainan Island. Geol Rev. 32(4):317–324.
- Lin JP. 2009. Review of the depositional environment of the Kaili Formation (Cambrian Series 2-3 boundary interval: China). Mem Assoc Australas Palaeontol. 37:131–149.
- Li GX. 1999. Early Cambrian Chancelloriids from Emei, Sichuan Province, SW China. Acta Palaeontol Pol. 38(2): 238–247. Chinese
- Lin JP, Scott AC, Li CW, Wu HJ, Ausich WI, Zhao YL, Hwu YK. 2006. Silicified egg clusters from a Middle Cambrian Burgess Shale type deposit, Guizhou, South China. Geo. 34(12):1037–1040.
- Liu Q, Lei QP. 2013. Discovery of an exceptionally preserved fossil assemblage in the Balang Formation (Cambrian Series 2, Stage 4) in Hunan, China. Alcheringa: An Australasian Journal of Palaeontology. 37(2):269–271. doi:10.1080/03115518.2013.764663.
- Li GX, Zhu MY, Michael S., Qian Y. 2004. Skeletal faunas from the Qiongzhusian of southern Shaanxi: biodiversity and lithofacies-biofacies links in the Lower Cambrian carbonate settings . Prog Nat Sci. 14(1):91–96. doi:10.1080/10020070412331343201.
- Luo HL, Jiang ZW, Wu XC, Song XL, Ou YL. 1982. The Sinian Cambrian Boundary in Eastern Yunnan, China. Yunnan Institute of Geological Sciences. 265. Chinese
- Mehl D. 1996. Organization and microstructure of the chancelloriid skeleton: implications for the biomineralization of the Chancelloriidae. Bull Inst Océanogr Monaco. 14:377–385.
- Mehl D. 1998. Porifera and Chancelloriidae from the Middle Cambrian of the Georgina Basin, Australia. Palaeontol. 41:1153–1182.
- Moore JL, Li GX, Porter SM., Lane, P. 2014. Chancelloriid sclerites from the Lower Cambrian (Meishucunian) of eastern Yunnan, China, and the early history of the group. Palaeontology. 57(4):833–878. doi:10.1111/pala.12090.
- Moore JL, Porter SM, Steiner M, Li GX. 2010. Cambrothyra ampulliformis, an unusual coeloscleritophoran from the Lower Cambrian of Shaanxi Province, China. J Paleontol. 84(6):1040–1060. doi:10.1666/09-091.1.
- Moore JL, Porter SM, Webster M, Maloof AC., Alvaro, J. 2021. Chancelloriid sclerites from the Dyeran–Delamaran (‘Lower–Middle’ Cambrian) boundary interval of the Pioche–Caliente region, Nevada, USA. Papers in Palaeontology. 7(1):1–59. doi:10.1002/spp2.1274.
- Mostler H, Mosleh-Yazdi A. 1976. Neue Poriferen aus oberkambrischen Gesteinen der Milaformation im Elburzgebirge (Iran) [New porifers from Upper Cambrian rocks of the Milaformation in the Elburz Mountains]. Geo Paläont Mitt Innsbruck. 5. 1–36. German
- Parkhaev PY, Demidenko YE. 2010. Zooproblematica and Mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontol J. 44:883–1161.
- Paterson JR, García-Bellido DC, Jago JB, Gehling JG, Lee MSY, Edgecombe GD. 2016. The Emu Bay Shale Konservat-Lagerstätte: a view of Cambrian life from East Gondwana. J Geol Soc London. 173(1):1–11. doi:10.1144/jgs2015-083.
- Porter SM. 2008. Skeletal microstructure indicates chancelloriids and halkieriids are closely related. Palaeontology. 51(4):865–879. doi:10.1111/j.1475-4983.2008.00792.x.
- Qian Y. 1989. Early Cambrian small shelly fossils of China with special reference to the Precambrian–Cambrian boundary (Stratigraphy and palaeontology of systemic boundaries in China: precambrian–Cambrian boundary. In: Nanjing: Nanjing University Publishing House.
- Qian Y, Chen ME, Chen YY. 1979. Hyolithid s and other small shelly fossils from the lower Cambrian Huangshandong Formation in the eastern part of the Yangtze Gorge. Acta Palaeontol Sin. 18(3): 207–232. Chinese
- Qian Y, Chen MG, He TG, Zhu MY. 1999. Taxonomy and Biostratigraphy of Small Crust Fossils in China. Beijing: Science Press. Chinese.
- Qian Y, Zhang SB. 1983. Small shelly fossils from the Xihaoping member of the Tongying Formation in Fangxian county of Hubei Province and their stratigraphical significance. Acta Palaeontol Sin. 22: 1 84–147. Chinese
- Qian Y, Zhu MY, Li GX, Jiang ZW, Heyo VI. 2002. A supplemental Precambrian Cambrian boundary global stratotype section in SW China. Acta Palaeontol Sin. 41(1):19–26.
- Randell RD, Lieberman BS, Hasiotis ST, Pope MC. 2005. New chancelloriids from the Early Cambrian Sekwi Formation with a comment on chancelloriid affinities. J Paleontol. 79(5):987–996. doi:10.1666/0022-3360(2005)079[0987:NCFTEC]2.0.CO;2.
- Rigby JK. 1978. Porifera of the Middle Cambrian Wheeler Shale, from the Wheeler Amphitheater, House Range, in western Utah. J Paleontol. 52:1325–1345.
- Rusconi C. 1954. Nuevas especies cámbricas del cerro Aspero, Mendoza [New Cambrian species from the Cerro Áspero, Mendoza]. Boletín Paleontoloógico de Buenos Aires. 29. 1–2. Spanish
- Sdzuy K. 1969. Unter- und mittelkambrische Porifera (Chancelloriida und Hexactinellida) [Lower and Middle Cambrian Porifera (Chancelloriida and Hexactinellida)]. Paläontologische Zeitschrift. 43(3–4): 115–147. German 10.1007/BF02987647
- Steiner M, Li GX, Qian Y, Zhu MY. 2004. Lower Cambrian small shelly fossils of northern Sichuan and southern Shaanxi (China), and their biostratigraphic importance. Geobios. 37(2):259–275. doi:10.1016/j.geobios.2003.08.001.
- Sun HJ, Zhao FC, Steiner M, Li GX, Na L, Pan B, Yin ZJ, Zeng H, Iten HV, Zhu MY. 2020. Skeletal faunas of the lower Cambrian Yu’anshan Formation, eastern Yunnan, China: metazoan diversity and community structure during the Cambrian Age 3. Palaeogeogr Palaeoclimatol Palaeoecol. 542:1–12. doi:10.1016/j.palaeo.2019.109580.
- Walcott CD. 1920. Cambrian geology and paleontology IV: 6 Middle Cambrian Spongiae. Smithson Misc Collect. 67:261–364.
- Wang MZ, Xu AD. 1987. [The establishment of lower Cambrian Huoshaodian Formation and its significance in stratigraphy, Zhenba, Southern Shanxi]. J Changchun Coll Geol. 17: 3 249–264. Chinese
- Yang XH, He YX, Deng SH. 1983. [On the Sinian-Cambrian boundary and the small shelly fossil assemblages in Nanjing area, Sichuan]. Bull Chengdu Inst Geol M R Chinese Acad. 4(2): 91–110. Chinese
- Yang B, Steiner M, Keupp H. 2015. Early Cambrian palaeobiogeography of the Zhenba–Fangxian Block (South China): independent terrane or part of the Yangtze Platform? Gondwana Res. 28(4):1543–1565. doi:10.1016/j.gr.2014.09.020.
- Yang YN, Zhao YL, Zhang XL. 2016. Fossil priapulid Ottoia from the Kaili biota (Cambrian Series 3) of South China. Journal of Systematic Palaeontology. 14(6):527–543. doi:10.1080/14772019.2015.1077900.
- Yin JC, Ding LF, He TG, Li SL, Shen LJ. 1980. The palaeontology and Sedimentary environment of the sinian system in Emei-Ganluo area, Sichuan. Chengdu: printed in the People’s Republic of China. Chapter 2. Characteristics of Biological Combination. 35–54. Chinese
- Yuan JL, Zhao YL, Li Y, Huang YZ. 2002. Trilobite fauna of Kaili Formation (uppermost Lower Cambrian Middle Cambrian) from Southeastern Guizou, South China. Shanghai: Science and Technology of Shanghai Press. Chapter 5, Sedimentary environment of Kaili Formation, burial science and paleoecology of trilobites; p. 44–48. Chinese.
- Yun H. 2019. Taxonomy and Evolution of the Cambrian Animal Chancelloriids [dissertation]. Shannxi: Northwest University. Chinese.
- Yun H, Brock GA, Zhang XL, Li LY, Garcia-Bellido DC, Paterson JR. 2019a. A new chancelloriid from the Emu Bay Shale (Cambrian Stage 4) of South Australia. Journal of Systematic Palaeontology. 17(13):857–867. doi:10.1080/14772019.2018.1496952.
- Yun H, Luo C, Chang C, Li LY, Reitner J, Zhang XL. 2022. Adaptive specialization of a unique sponge body from the Cambrian Qingjiang Biota. Proc Royal Soc B. 289(1976):1–9.
- Yun H, Reitner J, Zhang XL. 2023. Body reconstruction, taxonomy, and biostratigraphy of a ‘problematic’ chancelloriid. PalZ. doi:10.1007/s12542-023-00660-y.
- Yun H, Zhang XL, Brock GA, Li LY, Li GX. 2021. Biomineralization of the Cambrian chancelloriids. Geology. 49(6):623–628. doi:10.1130/G48428.1.
- Yun H, Zhang XL, Li LY. 2018. Chancelloriid Allonnia erjiensis sp. nov. from the Chengjiang Lagerstätte of South China. Journal of Systematic Palaeontology. 16(5):435–444. doi:10.1080/14772019.2017.1311380.
- Yun H, Zhang XL, Li LY, Pan B, Li GX, Brock GA. 2019b. Chancelloriid sclerites from the lowermost Cambrian of North China and discussion of sclerite taxonomy. Geobios. 53(2):65–75. doi:10.1016/j.geobios.2019.02.001.
- Zhang SG. 1995. New knowledge of the age of the beds with small shelly fossils in the Nanking Hills. J Stratigr. 19: 2 144–148. Chinese
- Zhang XL, Liu W, Zhao YL. 2008. Cambrian Burgess Shale-type Lagerstätten in South China: distribution and significance. Gondwana Res. 14(1–2):255–262. doi:10.1016/j.gr.2007.06.008.
- Zhang ZL, Pour MG, Popov LE, Holmer LE, Chen FY, Chen YL, Brock GA, Zhang ZF. 2021. The oldest Cambrian trilobite brachiopod association in South China. Gondwana Research. 89:147–167. doi:10.1016/j.gr.2020.08.009.
- Zhao J, Li G-B., Selden PA. 2018. New well-preserved scleritomes of Chancelloriida from early Cambrian Guanshan Biota, eastern Yunnan, China. J Paleontol. 92(6):955–971. doi:10.1017/jpa.2018.43.
- Zhao YL, Yuan JL, Loren EB, Guo QJ, Peng J, Yin LM, Yang XL, Peng SC, Wang CJ, Robert RG, et al. 2019. Global Standard Stratotype-Section and Point (GSSP) for the conterminous base of the Miaolingian Series and Wuliuan Stage (Cambrian) at Balang, Jianhe, Guizhou, China. J Stratigr. 42(2):165–184.
- Zhao YL, Yuan JL, Zhu MY, Yang RD, Guo QJ Qian Y, Huang YZ, Pan Y. 1999. A progress report on research on the early middle Cambrian Kaili biota, Guizhou. Acta Palaeontol Sin. 38: 1 1–14. Chinese
- Zhao YL, Zhu MY, Babcock LE, Peng J. 2011. The Kaili Biota—Marine organisms from 508 million years ago. Guiyang, Guizhou Science and Technology Press. Chapter 5. Systematic Paleontology. 68–76. Chinese
- Zhao YL, Zhu MY, Babcock LE, Yuan JL, Parsley RL, Peng J, Yang XL, Wang Y. 2005. Kaili Biota: a Taphonomic Window on Diversification of Metazoans from the Basal Middle Cambrian: Guizhou, China. Acta Geol Sin. 79(6):751–765. doi:10.1111/j.1755-6724.2005.tb00928.x.
- Zhu CF, Dong XP, Chen ME. 2003. Chancelloriids from Late Middle Cambrian of Huayuan, Hunan, South China. Acta Sci Nat Univ Pekin. 39: 5 716–726. Chinese
- Zhu MY, Zhuravlev AY, Wood RA, Zhao FC, Sukhov SS. 2017. A deep root for the Cambrian explosion: implications of new bio- and chemostratigraphy from the siberian platform. Geology. 45(5):459–462. doi:10.1130/G38865.1.
Annex
Figure A1. Archiasterella anchoriformis of Kaili Biota (MBP-38). The average angle of this specimen is 119.9°.

Figure A2. Archiasterella anchoriformis of Kaili Biota (MBP-43). The average angle of this specimen is 127.7°.

Figure A3. Archiasterella anchoriformis of Kaili Biota (JYS-1074-1). The average angle of this specimen is 130.8°.

Figure A4. Ar. coriacea of Burgess Shale (based on Bengtson and Collins, 2015). The average angle of this specimen is 105.1°.