
ABSTRACT
We described a new pair of incomplete mandibles from the Qingshuiying locality, Northwest China, and referred it to Eggysodon, E. lingwuensis sp. nov. The new species has a typical mosaic assemblage of features: dp1 (or p1) is present at adult stage, but the symphysis is posteriorly elongated and the diastema distance between the canine and dp1 (or p1) is further shortened. In terms of the evolutionary stage, E. lingwuensis sp. nov. is more advanced than E. osborni, but comparable to that of E. gaudryi. Together with other mammals from the Qingshuiying locality, we suggested that the strata-produced new material was deposited during the Early Oligocene. On the other hand, the continental fauna exchange during the Palaeogene between Europe and Asia had very scarce records. Eggysodon has four species from the Oligocene of Europe. This record definitely confirmed that there is a migration route between Central Europe and East Asia during the Early Oligocene.
Introduction
Yang and Zhou (Citation1956) and Hu (Citation1959) reported mammalian remains, including Cyclomylus, Archaeotherium, Eumeryx, Schizotherium, and Paraceratherium from the Qingshuiying locality of Ningdong Town, Ningxia, Northwest China. These genera are typical endemic groups from East Asia, and mainly from Northwest China and Mongolia. Cyclomylus and Eumeryx are two common genera from Northwest China, and also typical genera from Ulantatalian in the Early Oligocene (age of East Asia land mammals) (Wang et al. Citation2019). In addition, Schizotherium is from the Late Eocene, and Paraceratherium is found in the Early to Late Oligocene strata (Wang et al. Citation2019; Deng et al. Citation2021). This fauna assemblage makes the age of this locality debatable.
Eggysodon is a famous rhinoceros from the Oligocene Europe, with four species (Roman Citation1911; Stehlin Citation1930; Uhlig Citation1999; Antoine et al. Citation2011). This genus is a small-sized rhinoceros, characterised by the large canine and short distance between the canine and lower cheek teeth (Qiu and Wang Citation1999). Bai and Wang (Citation2012) reported a rhinoceros from the Late Eocene of Mongolia and assigned an independent genus Proeggysodon, indicative of a primitive form of Eggysodon. As for now, there are five genera are included in Eggysodontidae, Proeggysodon, Eggysodon, Ilianodon, Guangnanodon and Allacerops, and the last genus is from Central Asia and East Asia (Qiu and Wang Citation2007; Bai and Wang Citation2012). There has been no fossil record of Eggysodon from Asia until this work. We described an incomplete mandible attributed to Eggysodon, Eggysodon lingwuensis sp. nov. This new material adds to our knowledge of the mandible evolutionary tendency of Eggysodon, and also provides important evidence to infer the age of this locality produced new material.
Material and method
The new material is a pair of incomplete rhinoceros mandible found in yellow sandstone, with horizontal or trough cross-bedding, in the Qingshuiying locality of Ningxia, Northwest of China. Thin silty layers are sandwiched between these beddings of sandstone, and the gypsum horizon is occasionally present. A boulder bed is present in the upper part of the section. This locality and the bed in which the new mandible was found are the same as those mentioned by Yang and Zhou (Citation1956) and Hu (Citation1959). We will describe this new material and compare it with other contemporary rhinoceros, such as Ilianodon, Proeggysodon, Guangnanodon, and Allacerops. In addition to the published specimens of Eggysodon and Allacerops, materials of Ilianodon, Proeggysodon, and Guangnanodon from China are observed directly. In the Discussion section, the contents include inferences of the evolutionary tendency and age of stratigraphy.
In this study, terms of mandible and teeth follow the work of Qiu and Wang (Citation2007), and protocols of measure of teeth follow the criteria by Guerin (Citation1980). The lower cheek teeth is called with prefix ‘p’ for the lower premolar and ‘m’ for the lower molar.
Systematic paleontology
Order Perissodactyla Owen, Citation1848
Family Hyracodontidae Cope, Citation1879
Genus Eggysodon Roman, Citation1910
Type species: Eggysodon osborni (Schlosser, Citation1902)
Included species: Eggysodon gaudryi (Rames, Citation1886), Eggysodon pomeli (Roman, Citation1911), Eggysodon reichenaui (Deninger, Citation1903).
Stratigraphy and locality: Oligocene, China, Germany, Switzerland, and France.
Diagnosis
(revised from Uhlig Citation1999): Small-sized rhinoceros. Dentition formula: ?.1.4.3/2.1.4.3. On upper cheek teeth, the antecrochet is weak; the constriction of both the protocone and hypocone is obvious at the basal position; the lingual cingulum is present at the entrance of the median valley, and the labial cingulum is not continuous. On M3, the ectoloph and metaloph are fused, and the metacone is reduced. P2 is submolariform, but P3 and P4 are submolariform to paramolariform (based on criteria by Heissig, Citation1989). On the upper premolars, the lingual cingulum is developed and continuous, and the labial cingulum is always present but not continuous. The lingual cingulid of the lower cheek teeth is always interrupted around the metaconid. The lower canine is vertical, and larger than the incisors in size. The mandibular corpus is lower, and its ventral side is gradually upturning at position anterior to p4.
Eggysodon lingwuensis sp. nov.
Etymology
From the name Lingwu County, which is the administrative region with which the new locality is affiliated.
Holotype
IVPP V 25399, a mandible, with nearly all dentition but missing the vertical ramus.
Type locality and horizon
Qingshuiying locality of Ningdong town, Lingwu County, Ningxia Province, China. Fossil from the yellow sandstone, with horizontal or trough cross-bedding. Early Oligocene, Wulantatalian (Chinese Land Mammal Age), corresponding to Suevian (European Land Mammal Ages).
Diagnosis
The first lower cheek tooth dp1 is present, with a single root that is stronger than the root of p2. At the moderate wear stage, in the lower molars, the protolophid runs more lingually than the hypolophid did, and its labial wall is a cute angle; the posterior valley is almost U-shaped in occlusal view. The labial cingulid is present and continuous in the lower premolars but is interrupted at the hypoconid in the lower molars. The lingual cingulid is present but interrupted at the metaconid of all lower cheek teeth. The symphysis extends to the level of p3. Compared to other species, the diastema length between dp1 and the canine of the new species is the shortest (24 mm, ratio to the length of lower cheek row 0.114), and the length of the symphysis posterior to the first cheek tooth is the longest one. In size, the new species is the largest one of Eggysodon.
Description
The vertical ramus of the mandible is lost. The symphysis is narrow and anteriorly upturning, and its angle with the horizontal plane is 31°; its posterior end is at the level of the anterior border of p3. The distance between the canine and dp1 is short, approximately 24 mm. The distance from dp1 to the posterior end of the symphysis is approximately 39 mm. The ventral border of the mandible is horizontal. In dorsal view, the axis of the cheek tooth row is not consistent with the mandibular corpus.
There are two alveoli of incisors on each side, namely i1 and i2, and the alveoli of i1 are larger than those of i2. The preserved root of i1 is long and extends forward and upward, at an angle of 33° with horizontal plane. The third incisor i3 is completely reduced (). The canine is conical-shaped, but the lingual wall is more convex than the labial wall. It is greatly upturned, at an angle of 82° with horizontal plane (). The root is rounded and robust, with an anteroposterior diameter of 22 mm. The crown is slightly flattened laterally, and the height is 15 mm. The right canine shows that the enamel in the lingual wall has been worn off, leaving two deep and wide depressions at both the anterior and posterior edges. The occlusal surface is continuous to the root but constricted to the area adjacent to the enamel.
Figure 2. Shadowgraphs of mandibles by micro-CT, blue line marks outline of canine. A Proeggysodon qiui, IVPP V 18099, from the Erden Obo locality; B, Eggysodon lingwuensis sp. nov., IVPP V 25399, from the Qingshuiying locality. Not to scale.
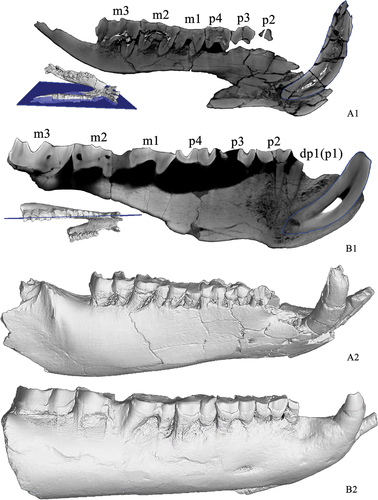
Figure 1. Mandible of Eggysodon lingwuensis sp. nov. from the Qingshuiying locality, Ningxia, China. A, occlusal view; B, right view; C, left view.
The cheek teeth have been heavily worn. The right dp1 (or p1) is preserved, with a columnar root, and simple crown, which has been crushed. The labial cingulid is present. The enamels on the occlusal surface of p2-p4 have been worn off, so that dentins of different lophids are fused with each other on the occlusal surface. The outline of p2 is triangular. The labial wall of the protolophid has a clear tendency to extend inward. The labial cingulid is strong and complete, but the lingual cingulid is not complete and is absent at the lingual wall of the metaconid. The ectoflexid is shallow and wide.
The secondary structures on the occlusal surface of p3 and p4 are also fused. The labial walls of the protolophid and the hypolophid are parallel to each other. The trigonid valley and the talonid valley are triangular in outline. The ectoflexid is shallow and narrow. The lingual cinguluid is reduced at the lingual wall of the metaconid. The labial cingulid is strong and complete.
The enamel of the occlusal surface of m1 has been fully worn off, and the dentine has been fused. The labial walls of the trigonid and talonid are rounded. The ectoflexid is shallow and wide. The lingual cinguluid is interrupted at the lingual wall of the metaconid. The labial cingulid is also interrupted at the labial wall of both the protolophid and the hypolophid.
The trigonid of m2 is crushed. The talonid valley has a U-shaped outline in occlusal view. The labial wall of the talonid is rounded. The lingual cingulid is absent at the lingual wall of the metaconid, but present at the entrance of the talonid valley. The labial cingulid is reduced and absent at the labial wall of the hypolophid.
The paralophid of m3 is short. The labial wall of the protolophid is not parallel to that of the hypolophid, and the former bends more lingually. The metalophid is parallel to the entolophid. The trigonid valley is triangular, but the talonid valley is U-shaped. The lingual cingulid is reduced and only present at the entrance of the talonid valley. The labial cingulid is strong and complete.
Comparison
Roman (Citation1910, Citation1911) suggested a new genus Eggysodon and assigned the European species ‘Ronzotherium’ osborni as the type species Eggysodon osborni. Stehlin (Citation1930) and Brunet (Citation1979) added more materials from France to this genus. Breuning (Citation1924) suggested a new subfamily Eggysodontinae and referred it to the family Hyracodontidae. Qiu and Wang (Citation2007) considered Eggysodontinae an independent family Eggysodontidae. Under the cladistic analysis, however, the content of this family becomes complex and uncertain (Wang et al. Citation2013, Citation2016; Bai et al. Citation2020). In Europe, Eggysodon is the only undebatable genus of this family.
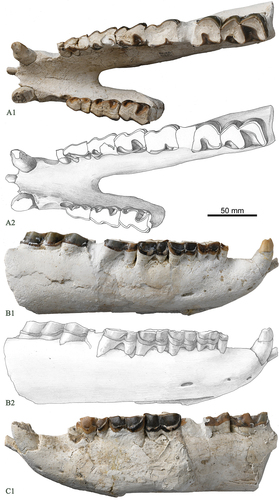
In East Asia, the known eggysodontids are mainly reported from the Eocene strata of Yunnan and Inner Mongolia, exemplified by Ilianodon, Proeggysodon and Guangnanodon (Chow and Xu Citation1961; Bai and Wang Citation2012; Wang et al. Citation2013). Ilianodon was excavated from the Late Eocene strata of the Lunan Basin, Yunnan, with ample materials (Chow and Xu Citation1961). This genus has features close to those of the new mandible: the symphysis is very short, ending at the level of p2, the diastema is very short, and the alveola is interpreted as an enlarged i2. However, in Ilianodon, the first lower cheek tooth dp1 is reduced, and the lower cheek teeth of are also distinct from the new teeth in that they have a set of special features: the lower premolars are not molariform, and they are much longer than wide; the metalophid in p3 and p4 is extends strongly posterolingually, and their entolophid is very weak; the paralophid in m1 and m2 is short, low and slender, but in m3 it becomes longer and extends nearly parallel to the metalophid, albeit still low and slender.
Guangnanodon is from the Late Eocene strata of Guangnan, Yunnan, close to the locality of Ilianodon (Wang et al. Citation2013). In this genus, the lower premolars are not molariform, less advanced than that of new mandible; in p4, the protolophid and the hypolophid are nearly parallel with each other, and its protolophid is close to the paralophid in length rather than longer than the latter in the new mandible; in the lower molars, the protolophid is oblique more lingually than that of the new material, and the paralophid is also longer albeit is low and slender; the lingual cingulid is reduced in all lower cheek teeth but is present with remains in the new material.
Proeggysodon from the Late Eocene strata of Erden Obo, China, is similar to the new material in having the two incisors, and the lower canine is large. However, the entoconid and the entolophid of Proeggysodon are very weak in the premolars, but the paralophid is long and forms a U-shaped trigonid in the molars (Bai and Wang Citation2012).
Borissiak (Citation1915) described some skull, mandible and postcranial bones from an Oligocene locality around Chelkar-Tenis in Kazakhstan, and assigned them as a new local species: ‘Epiaceratherium’ turgaicum. When discussing hyracodontids, Wood (Citation1932) combined Eggysodon and ‘Epiaceratherium’ turgaicum and named a new genus, namely, Allacerops, consisting of specimens from Europe and Asia. Because the presence of the incisor is unknown, this referral caused a long-term dispute over the taxonomical identity of Allacerops and Eggysodon (Beliajeva Citation1954; Brunet Citation1979; Dashzeveg Citation1991). Reshetov et al. (Citation1993) described a mandible from the Tort-Mola locality of Kazakhstan and noted that Allacerops differs from Eggysodon in the fully developed three incisors and premolars. In new mandible, two incisors are present, and the third one has been reduced. In addition, the lower molars of Allacerops have a V-shaped occlusal outline of talonid, different from the wide and U-shaped outline of the new mandible (Reshetov et al. Citation1993). We hence could not refer to the new mandible to Allacerops.
To date, four species have been recorded in Eggysodon, E. osborni, E. gaudryi, E. pomeli, and E. reichenaui. When establishing the genus Eggysodon, Roman (Citation1911) referred to three species, namely E. osborni, E. gaudryi and E. reichenaui, and named a new species, E. pomeli. Following Roman, ample materials were described (Stehlin Citation1930; Uhlig (Citation1999). E. pomeli from the Late Oligocene (MP29–30) has the specimen of only the upper cheek tooth and is not mentioned in the following comparison. E. reichenaui from the Early Oligocene has the very limited materials (Roman Citation1911; Uhlig Citation1999). Its lower cheek teeth are exemplified by the lower molars, similar to the new specimens in having a rectangular outline of whole-tooth outline, acute angle of the trigonid and right angle of the talonid, V-shaped trigonid valley and U-shaped talonid valley. The difference is mainly in the trigonid: the metalophid of the new lower molars is obliquely extending, not parallel with the paralophid, but these two lophs are nearly parallel and the metalophid is nearly transverse in E. reichenaui. In addition, E. reichenaui has lost the lingual cinguluid of the lower molars in the Early Oligocene specimens, whereas the lingual cingulid is present in the new specimens.
The materials of E. osborni are mainly from the Oligocene localities of France and Germany (Roman Citation1911). The lower cheek teeth share several similarities with the new specimen: except for p2, the trigonid is U-shaped and the talonid is L-shaped; the paralophid is absent in p2 but well developed in other cheek teeth; p3 and p4 are molariform; the labial cingulid is strong and continuous in the lower premolars but is slightly reduced and not continuous in the lower molars; the lingual cingulid is strong but interrupted at the metaconid in the lower premolars, and it is also interrupted in the lower molars but becomes weak. The mandible shown by Uhlig (Citation1999, text fig. 90, plate 3) suggested that it probably has a longer diastema of approximately 39 mm, but if dp1 is present, the diastema should be approximately 30 mm. Nevertheless, given its small-sized mandible (Uhlig Citation1999, table 122, p2-m3 length of 154 mm), the diastema of E. osborni is proportionally much longer than that of the new material, but the total length of the symphysis is much shorter (). Another difference lies in the metalophid, which extends slightly posterolingually in the lower molars and is not parallel with the entolophid; however, in the new material, the paralophid is parallel with the entolophid.
Eggysodon gaudryi is from the Oligocene localities of France, with ample materials (Roman Citation1911; Stehlin Citation1930; Uhlig Citation1999; Antoine et al. Citation2011). The mandibulae of E. gaudryi have a short symphysis, which ends at the anterior part of p2, and the diastema from the canine to p2 is 34 mm, which is approximately two times longer than the posterior part of the symphysis, namely, the distance from p2 to the posterior end of the symphysis. In the new mandible, however, the posterior part is nearly twice as long as the diastema length. Furthermore, the lower molar of E. gaudryi has a continuous labial cinguluid, which is reduced in new materials. It seems that, together with the smaller size of E. gaudryi (with an m1-m3 length of 96 mm), the new mandible is more advanced. However, among all mandibles of E. gaudryi, the first lower cheek tooth dp1 is completely reduced.
The above comparisons indicate that the new mandible belongs to the genus Eggysodon but differs from all known species of this genus (except for E. pomeli, which has no mandible fossil). We hence recognise this mandible as an endemic species, in the name of the place of production, Eggysodon lingwuensis sp. nov.
Discussion
Radinsky (Citation1966) united Allaceropinae and Eggysodontinae to Hyracodontidae, and Qiu and Wang (Citation2007) elevated Eggysodontinae to the family level Eggysodontidae. Here, we tentatively follow the view of the latter that considered Eggysodontidae as an independent family. When establishing the genus Eggysodon, Roman (Citation1911) highlighted the close distance between the lower canine and the first lower cheek tooth. Breuning (Citation1924) added the enforcement of the canine as diagnosis of this group. In this study, we presented the largest canine (length of crown and root, 16 mm and 22 mm) of this group. According to micro-CT results, the longer symphysis is caused by the enlarged and elongated lower canine: its root extends to the edge of dp1/p2 (). In Proeggysodon, both the symphysis and the root of the enlarged canine posteriorly extend to the same level, namely, the anterior edge of p2. In contrast, Ardynia has a canine smaller than the third incisor, and its symphysis ends before the first lower cheek tooth (Radinsky, Citation1967; Bai et al., Citation2018). Notably, Hyracodon nebraskensis has a canine close to the incisor in size (), but the posterior end of symphysis is at the level of the anterior part of p2, similar to that of Proeggysodon (Scott et al. Citation1941). Not only because the enlarged canine, backward retraction of the symphysis is also correlated with a shortened rostral area, especially the length of the diastema. Furthermore, the symphysis of the new mandible is not only proportionally short but also posteriorly elongated, representing the most advanced stage within Eggysodontidae according to the evolutionary tendency discussed by Roman (Citation1911) and Qiu and Wang (Citation1999).
Table 1. Measures of teeth of Eggysodon lingwuensis sp. nov. and other rhinoceroses (length/width/height) (mm).
In terms of the shortening of diastema, E. lingwuensis sp. nov. represents the most advanced species of this genus, and its canine is also among the largest canine (). In contrast, Eggysodon osborni has a relatively smaller canine and a shorter symphysis, and its diastema is longer than that of the other two species. As for E. gaudryi, it shows a different evolutionary direction: the first lower cheek tooth dp1 is completely reduced; however, this species is not more advanced than E. lingwuensis sp. nov., because of the proportionally longer symphysis and diastema. In contrast, E. lingwuensis sp. nov. is close to Proeggysodon qiui in size of canine and length of diastema, but symphysis of the former is much longer than the latter. These indicate that the short diastema has evolved since the Late Eocene, and the reduction of dp1 and the elongation of the symphysis have latterly occurred. Therefore, we suggest that the evolutionary stage of both two species E. gaudryi and E. lingwuensis sp. nov. are comparable, and E. osborni is the most primitive species. Fossil records show that E. osborni is from the earliest Oligocene, and E. gaudryi is from the early Late Oligocene to early Late Oligocene.
Table 2. Measures of mandible and teeth of Eggysodon lingwuensis sp. nov. and other rhinoceroses (mm).
The earliest publication of fossil from the Qingshuiying locality is by Yang and Zhou (Citation1956), followed by Hu (Citation1959). There are no other reports from this locality. As for now, five genera are found from the Qingshuiying locality. According to the newest suggestion of the Palaeogene integrative stratigraphy and timescale of China, Cyclomylus and Eumeryx are restricted to the Early Oligocene Ulantatalian Stage, Schizotherium is from the Late Eocene Baiyinian Stage. Paraceratherium is from the Oligocene (Deng et al., Citation2021). These records indicate an uncertain age, the Late Eocene or the Early Oligocene. Eggysodon is from the Early Oligocene to Late Oligocene, and the evolutionary stage of new species Eggysodon lingwuensis sp. nov. is also restricted to this age. This allows us to determine with certainty that the age of sediments that produced new materials is in the Early Oligocene.
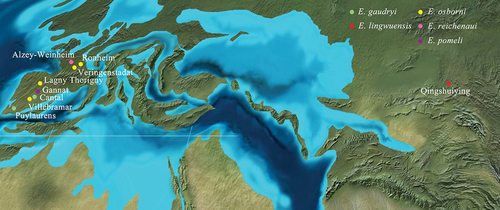
During the Palaeogene, mammalian communications between Asia and Europe were scarce. Nevertheless, several genera are shared between two continents, such as Artiodactyla Entelodon, Carnivora Hyaenodon, and Rodentia Eucricetodon (Brunet Citation1979; Dawson Citation2003; Wang et al. Citation2019). The oldest records of genera within Rhinocerotidae shared by Europe and Asia are Ronzotherium from the Late Eocene strata in Mongolia and Epiaceratherium from the Late Eocene strata in Vietnam (Dashzeveg Citation1991; Wang et al. Citation2013). Another case is the giant rhinos, which has ample fossils reported in China, Balkan Peninsula, Turkey, and Pakistan (Deng et al. Citation2021). On the other hand, although the primitive eggysodontid Proeggysodon has been reported from the Late Eocene of Inner Mongolia (Bai and Wang Citation2012; Bai et al. Citation2020), Eggysodon from the Oligocene is thus far absent in other areas of Asia until this paper. The new species of Eggysodon from the Northwest of China is the first reliable record of this genus in Asia (). These records further confirmed that East Asia and Europe are not separated through the Eocene to the Oligocene. The shortest route of migration is through East Europe and Central Asia, but North America is probably an alternative pathway.
Figure 3. Localities of Eggysodon around Europe and Asia, and five species are marked by different colours. The locality data are from Stehlin (Citation1930); Brunet (Citation1979); Uhlig (Citation1999); Antoine et al. (Citation2011).
Acknowledgments
We thank Wang Shiqi, Bai Bin, Gao Wei, Zang Hailong, Liu Yonghong, Zhang Xuelin, Chen Jin, and Si Hongwei (Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China), for their help to this work. We are grateful to Xu Yong and Chen Yu for preparing the drawing.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Antoine PO, Métais G, Orliac MJ, Peigné S, Rafaÿ S, Solé F, Vianey-Liaud M. 2011. A new late Early Oligocene vertebrate fauna from Moissac, South-West France. C R Palevol. 10(4):239–250. doi: 10.1016/j.crpv.2011.01.004.
- Bai B, Meng J, Zhang C, Gong YX, Wang YQ. 2020. The origin of Rhinocerotoidea and phylogeny of Ceratomorpha (Mammalia, Perissodactyla). Commun Biol. 3(1):509. doi: 10.1038/s42003-020-01205-8.
- Bai B, Wang YQ. 2012. Proeggysodon gen. nov., a primitive Eocene eggysodontine (Mammalia, Perissodactyla) from Erden Obo, Siziwangqi, Nei Mongol, China. Vertebrata of PalAsiatica. 50(3):204–218.
- Bai B, Wang YQ, Zhang ZQ. 2018. The late Eocene hyracodontid perissodactyl Ardynia from Saint Jacques, Inner Mongolia, China and its implications for the potential Eocene–Oligocene boundary. Palaeoworld. 27:247–257.
- Beliajeva EI. 1954. New material of Tertiary Rhinocerotids (in Russian). Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR. 47:24–54.
- Borissiak AA. 1915. Mammifères fossiles de Sébastopol II, Trudy Geolegicheskago Komiteta Novaja Seria. 137:1–45.
- Breuning S. 1924. Beiträge zur Stammesgeschichte der Rhinocerotidae. Verhandlungen der Zoologischen-Botanischen Gesellschaft in Wien. 73:5–46.
- Brunet M. 1979. Les grands mammifères chefs de file de l’immigration Oligocène et le problème de la limite Éocène-Oligocène en Europe. Paris: Foundation Singer-Polignac Mémoires du Muséum national d’Histoire naturelle; p. 281.
- Chow MC, Xu YX. 1961. New primitive true rhinoceroses from the Eocene of Iliang, Yunnan. Vert PalAs. 5:291–304.
- Cope ED. 1879. On the extinct American rhinoceroses and their allies. The American Naturalist. 13:771a–771j.
- Dashzeveg D. 1991. Hyracodontids and rhinocerotids (Mammalia, Perissodactyla, Rhinocerotoidea) from the Paleogene of Mongolia. Palaeovertebrata. 21(1–2):1–84.
- Dawson MR. 2003. Paleogene rodents of Eurasia. Deinsea. 10(1):97–126.
- Deng T, Lu X, Wang S, Flynn LJ, Sun D, He W, Chen S. 2021. An Oligocene giant rhino provides insights into Paraceratherium evolution. Commun Biol. 4(1):639.
- Deninger K. 1903. Ronzotherium Reichenaui aus dem Oligocän Ton Weinheim bei Alzey. Zeitschrift der Deutschen Geologischen Gesellschaft. 55:93–97.
- Guérin C. 1980. Les rhinocéros (Mammalia, Perissodactyla) du Miocène terminal au Pléistocène supérieur en Europe occidentale: comparaison avec les espèces actuelles. Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon. 79:1–1182.
- Heissig K. 1989. Rhinocerotidae. In: Prothero D, and Schoch R, editors. The Evolution of Perissodactyls. New York: Oxford University Press; pp. 399–417.
- Hu CK. 1959. Chalicotherids from the Tertiary in North China. Vert PalAs. 1:33–40.
- Owen R. 1848. On the Archetype and Homologies of the Vertebrate Skeleton. London: John van Voorst.
- Qiu ZX, Wang BY. 1999. Allacerops (Rhinocerotoidea, Perissodactyla), its discovery in China and its systematic position. Vertebrata of PalAsiatica. 37(1):48–61.
- Qiu ZX, Wang BY. 2007. Paracerathere fossils of China. Palaeontologia Sinica. 29:1–396.
- Radinsky LB. 1966. The families of the Rhinocerotoidea (Mammalia, Perissodactyla). Journal of Morphology. 47(4):631–639. doi: 10.2307/1377893.
- Radinsky LB. 1967. A review of the Rhinocerotoid family Hyracodontidae (Perissodactyla). Bulletin of the American Museum of Natural History. 136(1):1–45.
- Rames MB. 1886. Note sur l’âge des argiles du Cantal et sur les débris fossiles qu'elles ont fournis. Bull Soc géol Fr. 3:357–360.
- Reshetov V, Spassov N, Baishashov B. 1993. Tenisia gen. nov.: taxonomic revaluation of the Asian Oligocene rhinocerotoid Eggysodon turgaicum (Borissiak, 1915) (Mammalia, Perissodactyla, Hyracodontidae). Geobios. 26(6):715–722. doi: 10.1016/S0016-6995(93)80054-U.
- Roman MF. 1910. Sur les Rhinocerides de l'Oligocene d'Europe et leur filiation. Compte Rendu des Seances de l'Academie des Sciences de Paris. 150:1558–1560.
- Roman MF. 1911. Les rhinocéridés de l’oligocène d’Europe. Archives Muséum d’Histoire Naturelle de Lyon. 11(1):1–110. doi: 10.3406/mhnly.1912.966.
- Schlosser M. 1902. Beiträge zur Kenntnis der Säugethierreste aus den süddeutschen Bohnerzen. Geologische und Palaöntologische Abhandlungen, Jena, NF. 5:117–258.
- Scott WB, Jepsen GL, Wood AE. 1941. The mammalian fauna of the White River Oligocene, Part Ⅴ, Perissodactyla. Transactions of the American Philosophical Society. 28(5):747–980. doi: 10.2307/1005518.
- Stehlin HG. 1930. Bemerkungen zur Vordergebissformel der Rhinocerotiden. Eclogae Geologicae Helvetiae. 23(2):644–648.
- Uhlig U. 1999. Die Rhinocerotoidea (Mammalia) aus der unteroligozänen Spaltenfüllung Möhren 13 bei Treuchtlingen in Bayern. Bayerische Akademie der Wissenschaften Mathematisch-Naturwissenschaftliche Klasse, Abhandlungen NF. 170: 1–254.
- Wang HB, Bai B, Gao F, Ch HW, WY Q. 2013. New eggysodontid (Mammalia, Perissodactyla) material from the Paleogene of the Guangnan Basin, Yunnan Province, China. Vertebrata PalAsiatica. 51(4):305–320.
- Wang H, Bai B, Meng J, Wang Y. 2016. Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. Scientific Reports. 6(1):39607. doi: 10.1038/srep39607.
- Wang YQ, Li Q, Bai B, Jin X, Mao FY, M J. 2019. Paleogene integrative stratigraphy and timescale of China. Science China Earth Sciences. 62(1):287–309. doi: 10.1007/s11430-018-9305-y.
- Wood IH. 1932. Status of Epiaceratherium (Rhinocerotidae). J Mammal. 13(2):169–171.
- Yang CJ, Zhou MZ. 1956. Mammalian fossils from the Oligocene in Lingwu, Gansu. Acta Palaeontologica Sinica 4:3–16.