
ABSTRACT
We report here diverse Late Miocene fish assemblages based on otoliths collected from sedimentary rocks deposited in coastal marine settings in northern Borneo (Seria and Miri formations and Tutong beach). Surface-collected, large-sized otoliths, representing > 96% of the whole assemblage, are dominated by the families Sciaenidae and Ariidae, while Haemulidae, Engraulidae, and Lactaridae can be common at certain sites. Additional taxa were revealed by screen-washing of the sediments from selected sites including taxa from the families of Congridae, Bregmacerotidae, Apogonidae, Gobidae, Sillaginidae, and Cepolidae. Among the sciaenids, we describe six new taxa (Advenasciaena bruneiana, Atrobucca borneensis, Bruneisciaena schwarzhansi, Nibea ambugensis, Nibea stintoni, and Protonibea nolfi), and we also report the first fossil occurrence of some other groups (e.g. Lactarius lactarius, Acanthocepola sp.) from the Indo-Australian Archipelago (IAA). All these discoveries add to our understanding of fish palaeobiodiversity in the IAA where today’s highest marine biodiversity exists. The compositions of the faunas reflect a shallow marine coastal palaeoenvironment with close connection to estuaries where some of the reported fishes might have migrated seasonally or daily with the tide. Turbid, suspension-loaded water might be also presumed, where ariids, sciaenids, and the haemulid Pomadasys are often observed today.
ZooBank LSID for the described genera:
Advenasciaena zoobank.org:act:5D6D3223-462F-4488-88BE-8DDDB107BFB6
Bruneisciaena urn:lsid:zoobank.org:act: 090DAD2F-FB18-43CB-9CA9-5F9FE224CA54
ZooBank LSID for the described species:
Advenasciaena bruneiana urn:lsid:zoobank.org:act: D14930B9-5847-4A1F-9D20-EFC0501F3B3D
Atrobucca borneensis urn:lsid:zoobank.org:act: 36E0F8EC-0266-4BBB-B00A-B43C1B1C7557
Bruneisciaena schwarzhansi urn:lsid:zoobank.org:act: B9B2F637-CC09-44A3-AEE6-6C8EE76FBE34
Nibea ambugensis urn:lsid:zoobank.org:act: 8C780272-A910-404D-89FA-C0BB110B9CEF
Nibea stintoni urn:lsid:zoobank.org:act: 8D750E35-8BD4-481A-822D-7F4DA3816E04
Protonibea nolfi urn:lsid:zoobank.org:act: 81BC0700-50E1-400B-9034-F83DDA6C7710
Introduction
Borneo is part of the Central-Indo-Pacific ecoregion and within that the Indo-Australian Archipelago (IAA), which hosts today’s highest marine fish biodiversity with species richness of over 2000 (e.g. Hoeksema Citation2007; Spalding et al. Citation2007; Cowman Citation2014). The hotspot of biodiversity has existed here since the Miocene as it has shifted from west to east as the closure of the ancient Tethys Ocean advanced during the early Cenozoic (Renema et al. Citation2008). Many invertebrate taxa moved with the hotspot as attested by the fossil record, but in the case of fishes, the ancient origin of some of the groups is supported by molecular phylogeny, which has inferred many new speciation events in the area (e.g. Frédérich et al. Citation2013; Cowman Citation2014; Sorenson et al. Citation2014). In contrast, the Southeast Asian fossil teleost fish record is very sparse, and besides a few reported body fossils (Brouwer and de Citation1923; Beaufort de Citation1926, Citation1931) and sporadic bone remains (e.g. Martin Citation1883; Hennig Citation1911; Koumans Citation1949; Leriche Citation1954), most of our knowledge on the biodiversity of this area comes from otoliths (ear stones) (e.g. Vorstman Citation1927; Nolf and Bajapi Citation1992; Van Hinsbergh and Helwerda Citation2019; Lin et al. Citation2021; Lin and Chien Citation2022). Teleost fish have three pairs of otoliths, the saccular (sagitta), the lagenar (asteriscus), and the utricular (lapillus) otoliths. They are situated in the inner ear at the back of the cranium, and their functions are mainly linked to coordination of movement, sense of balance, and hearing. For most teleosts, the sagittae are the largest otolith, which is made up primarily of aragonite. The lapillus is also built up from aragonite, while asteriscus is primarily composed of vaterite. The size range and shape of the otoliths vary among the taxa, and their morphological patterns, especially that of the sagitta, are an important identifying tool (e.g. Nolf Citation1985, Citation2013). Otoliths are often found in sedimentary rocks and can provide a quick and broad overview on fish biodiversity through time. Several earlier studies described diverse fish faunas from the IAA based on the otoliths (e.g. Frost Citation1925; Vorstman Citation1927; Posthumus Citation1929; Stinton Citation1962). However, the original illustrations of some specimens are difficult to use for precise taxonomy (see Nolf Citation2013). Therefore, to obtain a better understanding on ancient fish diversity, a revision of these faunas is necessary in the future. From northern Borneo, Stinton (Citation1962) studied Neogene otoliths and reported 62 specimens of 18 species pertaining to 10 families. Among these taxa, five were also mentioned from Brunei Darussalam (north Borneo, ). Sixty-six percent of the fauna studied by Stinton represented Sciaenidae (drums and croakers) and Ariidae (sea catfish), with two and four newly described fossil species respectively of these families. Between 2015 and 2021, several field surveys were conducted by the Universiti Brunei Darussalam on the Late Miocene shallow marine deposits in Brunei that resulted in a large number of fish otoliths (~1500). These newly collected specimens are described here, with a thorough comparison with those of Stinton’s collection housed in the Natural History Museum in London and, in some cases, with modern otoliths recovered from Brunei waters.
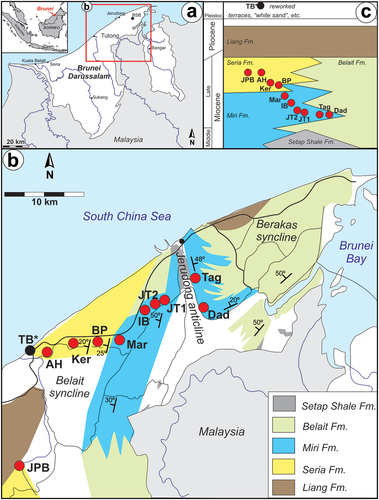
Figure 1. Geographic and geological positions of the localities that yielded the reported otoliths. Note that the depositional age of the outcrops gets younger from east to west. Miri Formation sites: Dad – Dadap, Tag – Tagap, JT1 – Tanjong Nangka, JT2 – Jalan Tutong 2, IB – Ikas Bandung, Mar – Maraburong; Seria Formation sites: PB – Pak Bidang; Ker – Keriam; AH – Ambug Hill, JBP – Jalan Pak Bidang. Reworked specimens were also discovered on Tutong beach (TB*).
Geological setting
Brunei Darussalam is situated in the central part of north Borneo in Southeast Asia (). The country’s geology is dominated by siliciclastic sedimentary rocks (e.g. Liechti et al. Citation1960; Wilford Citation1961; James Citation1984), that were mainly deposited in shallow marine coastal environments with various tide and wave-dominated deltas (Lambiase et al. Citation2003; Lambiase and Cullen Citation2013). The oldest lithostratigraphic unit in the area studied is the Setap Shale Formation, representing a deeper, offshore setting of Early – Middle Miocene age (). These rocks occur in the core of the Jerudong anticline and at the base of the older part of the Belait Formation. This latter unit predominantly represents sandy, shallow-water facies with a strong deltaic influence. The younger part of the Belait Formation is intercalated with the Miri and Seria formations, which were deposited in shallow marine milieu. These units are built up of several small parasequences, often starting with claystone levels rich in shallow marine faunas, which are followed by gradually coarser sediments (siltstone, sandstone) indicating a shallowing environment. The sequences usually end with an erosional surface before the new cycle starts. The age of the Belait and Miri formations in the studied area is Middle and Late Miocene, while the Seria Formation is Late Miocene in age (Sandal Citation1996; Back et al. Citation2005; Kocsis et al. Citation2018, Citation2022). All these sediments are overlain by the Liang Formation, representing a lagoonal and fluvial depositional environment with thick conglomerate beds, lignite horizons, and levels rich in plant remains at some places (e.g. James Citation1984; Wannier et al. Citation2011; Wilf et al. Citation2022). The age of the Liang Formation is latest Miocene to Pliocene. Brunei’s younger geology is characterised by Pleistocene terraces and white sand deposits (James Citation1984; Sandal Citation1996). Erosion of these younger beds together with uplifted older geological units is apparent in the coastal areas. In some places, especially in the Tutong region (), it has resulted in reworked fossils washed up by the sea along the beaches (Kocsis et al. Citation2020). Selected vertebrate fossils, among them an otolith, were dated with Sr-isotope stratigraphy with a result of Late Miocene age (Kocsis et al. Citation2022). The data strongly suggest that most of the vertebrate remains on the beach are originated from the sediments of the Seria Formation. However, the contribution of fossils from older marine beds, such as the Miri Formation, cannot be entirely excluded (Kocsis et al. Citation2020).
Frequent occurrence of fish remains have been recently reported from the clayey beds of the Miri and Seria formations (Kocsis et al. Citation2018; Roslim et al. Citation2020), and in fact, Stinton’s (Citation1962) pioneer work on Borneo otoliths also dealt with material from these sedimentary sequences. Other fish remains, such as shark and ray teeth, were described from the Seria Formation at Ambug Hill (Razak and Kocsis Citation2018; Kocsis et al. Citation2019), while reworked elasmobranch material was mentioned from the coastal region of Tutong (Kocsis et al. Citation2020). Besides the elasmobranch occurrences in the Seria and Miri formations, only a few sporadic elasmobranch teeth are known from some levels of the Belait Formation.
Material and methods
Seven years of regular field surveys to the fossiliferous beds of the Miri and Seria formations yielded a large amount of fossil fish material, among them numerous otoliths (~1500). Here we focus on fossil fish otoliths that were discovered from six Miri and four Seria formation localities. In addition, many specimens were also found reworked at the Tutong beach (, ). The oldest specimens are from the Miri Formation sites of Dadap (Dad) and Tagap (Tag) from the eastern flanks and Tanjong Nangka (JT1) from the western flanks of the Jerudong anticline (). The remaining localities are on the western flank of the anticline, and their age gradually gets younger towards the west: Jalang Tutong 2 (JT2), Ikas Bandung (IB), Maraburong (Mar), Bukit Pangal (BP), Keriam (Ker), and the youngest material is from Ambug Hill (AH) and Jalan Pak Bidang (JPB) outcrops. The overall covered age range is Late Miocene (Tortonian – early Messinian see Kocsis et al. Citation2018, Citation2022).
Table 1. Geographical and geological information on the sampled localities. Note that at the locality of Tanjong Nangka (JT-1), three clay levels yielded fish remains and when it is known for the given specimen, it is referred to in the inventory as JT1-L1, -L2 and -L3, respectively.
The studied otoliths were collected from claystone beds (particle size is dominantly bellow 1/256 mm) from the ten localities (). They were picked from the sediment surface, and then cleaned from the adherent sediment particles in an ultrasonic bath. The reworked Tutong specimens were found on the beach at low tide together with many other vertebrate remains (e.g. shark teeth, turtle bones). The age of these remains is Late Miocene, and most of the material is probably derived from sediments of the Seria Formation (Kocsis et al. Citation2022). Nevertheless, the reworked fauna is handled separately from the in-situ material with an asterisk marking the locality’s abbreviation (e.g. Tutong beach – TB*).
Clay-rich sediment samples (0.2–1 kg) were screen-washed (mesh-sizes of 250 µm, 500 µm, and 1 mm) for micro-fossils (mainly for foraminifera), which occasionally yielded otoliths, especially from AH, Ker, IB, JT1, and Dad. Selected, well-preserved specimens were then photographed in different views either with a Nikon D7000 camera or with a Keyence VHX-7000 digital microscope housed at the Institute of Earth Sciences at the University of Lausanne, Switzerland. A few small specimens were documented with a Scanning Electron Microscope (SEM) JSM-7610F at Universiti Brunei Darussalam. The images are displayed in the photo plates (). The taxonomic work and morphological description of the otoliths are mainly based on Schwarzhans (Citation1993), Ohe (Citation2006), Lin and Chang (Citation2012), and Nolf (Citation2013). For higher taxonomic ranks, Betancur-R et al. (Citation2017) is followed. The majority of the otoliths are housed in the Geological Collection of the Natural History Museum Universiti Brunei Darussalam (GUBD), while the type and many of the figured specimens are stored in the Natural History Museum of London (NHMUK). A few paratypes of newly described species, which also belong to new genera, are also deposited in the Naturalis Biodiversity Center in Leiden, The Netherlands (RGM) and at the Biodiversity Research Museum, Academia Sinica, Taiwan (BRMAS, under the registration code ASIZF). The full species list with corresponding inventory numbers and the figured specimens are listed in Supplementary material 1. Stinton’s collection at the NHMUK was studied for comparison purposes. Those specimens that are referred to in the text with an NHMUK collection number (holotypes, paratypes, etc.) are listed and figured in Supplementary material 2 in the order of appearance in the text. From the following systematic section onwards when the lithostratigraphic origin of the specimens are discussed Formation is abbreviated as Fm.
Figure 2. Congride and Engraulidae. a-c: Ariosoma sp. from AH (a: NHMUK PV P 76628, b-c: GUBD V0172), inner views; d-e: Setipinna cf. retusa from IB (NHMUK PV P 76629), inner and external views; f-g: Setipinna sp. two specimens from Dad (GUBD V0174) inner views; h-m: Coilia planata (NHMUK PV P 76630) from Dad, posterior (f), inner (g), anterior (h), external (i), ventral (j) and dorsal (k) views; n-q: C. planata inner views of specimens from Dad n & o (NHMUK PV P 76631-32), p: IB (GUBD V0175) and q: Ker (GUBD V0176); r-s: C. planata juvenile specimen from JT1-L2, inner and external views (NHMUK PV P 76633); t-u: Coilia cf. mystus from Ker (NHMUK PV P 76634). Scale bars: 0.5 mm.

Figure 3. Ariidae. a-d: Ariidae indet. 1 from AH (GUBD V0125); e-h: Ariidae indet. 2 from JT-1 (GUBD V0141); i-l: Ariidae indet. 3 from AH (GUBD V0130); m-p: Ariidae indet. 4 from JT-1 (GUBD V0143). Orientations: ventral (a-b, e-f, i-j, m-n), dorsal (b, f, j, n), and mesial (d, h, l, p) views. Note that the drawings (a, e, i, m) are emphasizing the differences among the distinguished morpho-groups. Scale bar: 5 mm. Abbreviations: Amp - anteromesial projection, Lb - linea basalis.
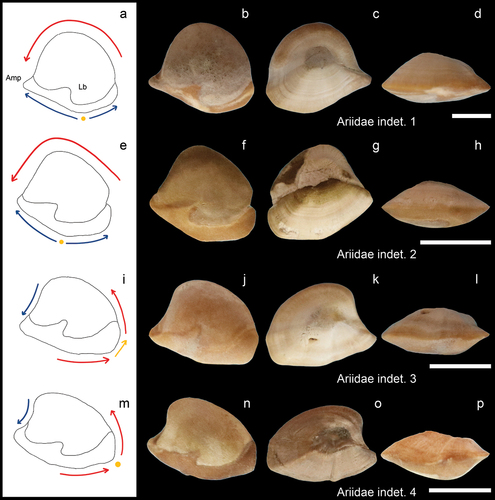
Figure 4. Bregmacerotidae, Apogonidae, and Gobiidae. a-b: Bregmaceros sp. from AH (GUBD V0182) and from IB (NHMUK PV P 76635), inner views; c-h: Apogonidae indet. from AH, c-f (GUBD V0184), inner (c), external (d), ventral (e) and inner dorsal (f) views; g (GUBD V0185) and h (GUBD V0186), inner views; i-n: Gobiidae indet. inner views i-l (GUBD V0187) & n (GUBD V0188) from AH and m from IB (GUBD V0191). Scale bar: 0.1 mm except the big Apogonidae (c-e) which is 0.5mm. The a, g, h, i-l, & n are SEM images.
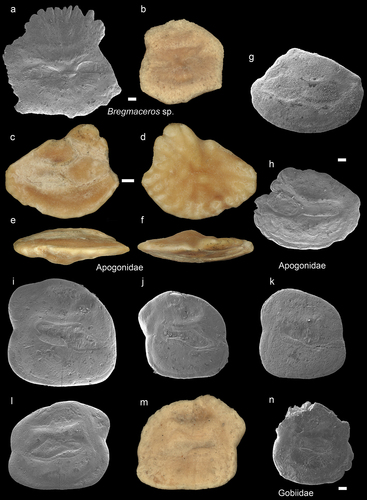
Figure 5. Lactariidae. a-g: Lactarius lactarius fossil specimens from AH. a-d (NHMUK PV P 76636), inner (a), external (b), dorsal (c), and ventral (d) views; e-f inner views of NHMUK PV P 76637 PV P 76638; g – SEM image of a juvenile specimen (GUBD V0194). h-k: modern Lactarius lactarius left and right otoliths BRUJM-20201218-1 BRUJM-20201218-3, TL: 18.5 and 14 cm). Scale bar: 1mm, except 0.1 mm for g.
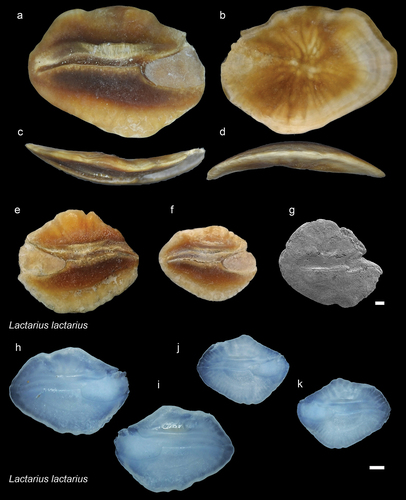
Figure 6. Sciaenidae. a-f: Advenasciaena bruneiana holotype (NHMUK PV P 76639) from JT1 site’s L2 major clay layer: inner (a), external (b), anterior (c), dorsal (d), ventral (e) and posterior (f) views. g-l: A. bruneiana paratype (ASIZF 0100316) from JT1 site’s L2 major clay layer: inner (g), external (h), anterior (i), dorsal (j), ventral (k) and posterior (l) views. m-r: A. bruneiana paratype (RGM.1364031) from Dadap site: anterior (m), inner (n), posterior (o), external (p), ventral (q), dorsal (r) views. Scale bar 1 mm.
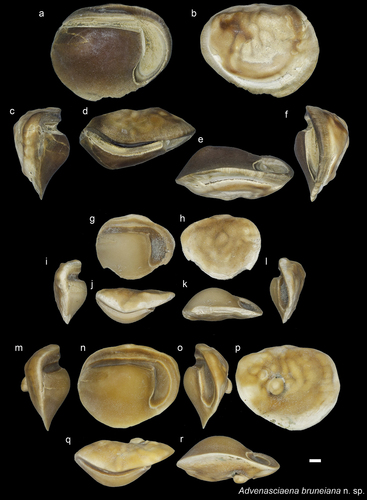
Figure 7. Sciaenidae. a-f: Atrobucca borneensis holotype from AH (NHMUK PV P 76640), posterior (a), inner (b), anterior (c), external (d), dorsal (e) and ventral views (f); g-h: A. borneensis paratype from AH (NHMUK PV P 76641), inner and external views; i-k: A. borneensis from AH inner views of selected size range (NHMUK PV P 766342-44); l-p: Atrobucca aff. borneensis from AH (GUBD V0198), l-n specimen 1: inner (l), dorsal (m) and external (n) views; o-p specimen 2: inner (o) and external (p) views. Scale bar: 1 mm.
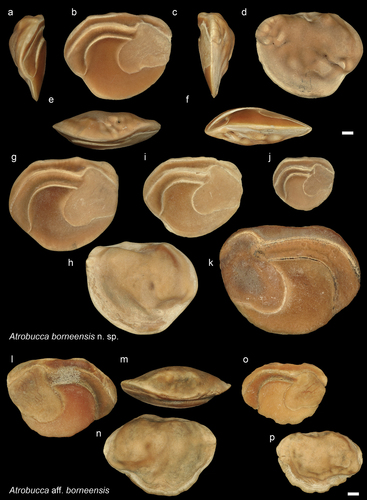
Figure 8. Sciaenidae. a-f: Boesemania obesa from Dad (NHMUK PV P 76645), inner (a), external (b), postrior (c), dorsal (d), anterior (e), ventral (f) views; g: B. obesa from AH inner view (GUBD V0199); h: B. obesa from JT1 inner view (GUBD V0200); i: Boesemania microlepis extant species from Vietnam (GSCN3846). Scale bar: 1mm.
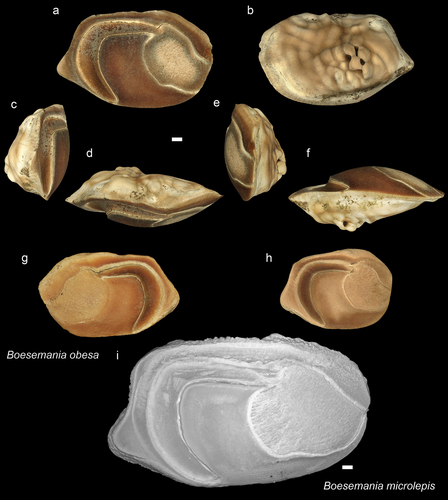
Figure 9. Sciaenidae. a-f: Bruneisciaena schwarzhansi holotype (NHMUK PV P 76646 from Dadap village: posterior (a), inner (b), anterior (c), external (d), dorsal (e), ventral (f) views. g-i: B. schwarzhansi from Dadap paratype (ASIZF 010031): inner (g), dorsal (h), ventral (i) views; j-m: B. schwarzhansi paratype (RGM.1364030) from Bukit Panggal: inner (j), anterior (k), posterior (l), ventral (m) views. n-s: B. schwarzhansi specimens, all with inner view from the type locality of Dadap village (n-p: NHMUK PV P 76647-49 r-s: GUBD V0161). Scale bar 1 mm.
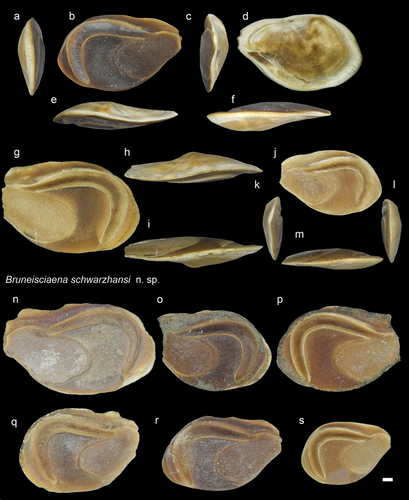
Figure 10. Sciaenidae. a-f: Larimichthys polyactis from AH, inner (a), external (b), posterior (c), dorsal (d), anterior (e), ventral (f) views (NHMUK PV P 76650); g-j: L. polyactis from AH all in inner views (NHMUK PV P 76651-54). Scale bar: 1 mm.
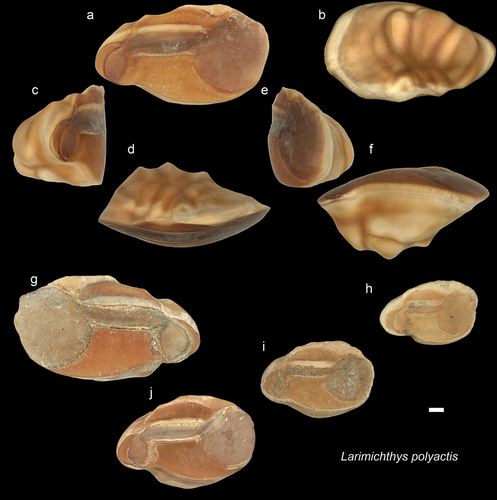
Figure 11. Sciaenidae. a-f: Dendrophysa angulata from AH, posterior (a), inner (b), anterior (c), external (d), dorsal (e) and ventral (f) views (NHMUK PV P 76655); g-n: D. angulata specimens from AH in inner views (g-j: NHMUK PV P 76656-59 k-n: GUBD V0207); o-p: D. angulata from Ker (GUBD V0208 – 2 specimens), inner views. Note the different preservation states. q-r: D. angulata from the Miri Formation all are in inner views q – JT2 (NHMUK PV P 76660), r – JT1-L2 (NHMUK PV P 76661). s-t: Dendrophysa russelii – modern species from Brunei (GSCN4049 TL: 12 cm GSCN4050 TL: 12.9 cm). u-z: Dendrophysa sp. from Ker, posterior (u), inner (v), anterior (w), external (x), dorsal (y) and ventral (z) views; aa-ab: Dendrophysa sp. from Ker and IB, respectively. (u-z aa: NHMUK PV P 76662-63, ab: GUBD V0216). Scale bar: 1 mm.
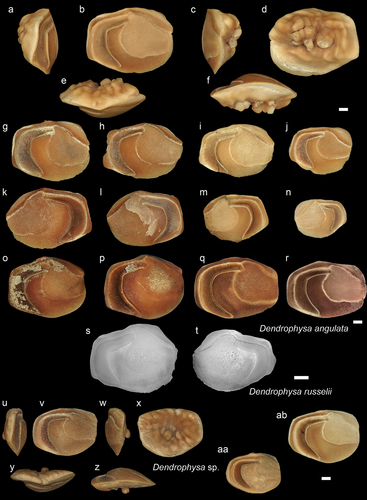
Figure 12. Sciaenidae. a-f: Nibea ambugensis from AH (holotype - NHMUK PV P 76664), posterior (a), inner (b), anterior (c), external (d), dorsal (e) and ventral (f) views; g-q: N. ambugensis specimens from different sizes and localities in inner views. g - from JPB (GUBD V0219), i – from Ker (GUBD V0220), l – Dad (GUBD V0221), h, j, k, m-q from AH (NHMUK PV P 76665-72); r-z: Nibea cf. soldado from AH, specimen-1: anterior (r), inner (s), posterior (t), external (u), dorsal (v) and ventral (w) views (NHMUK PV P 76673), specimen 2: anterior (y) and dorsal (x) views (NHMUK PV P 76674), specimen 3 inner (z) view (NHMUK PV P 76675), aa: Nibea cf. soldado from Ker (GUBD V0224); ab-ac: Nibea soldado modern specimen from Brunei (GSCN4051, TL: 19.7cm and GSCN4053, TL:20.6 cm), inner view. Scale bar: 1mm.
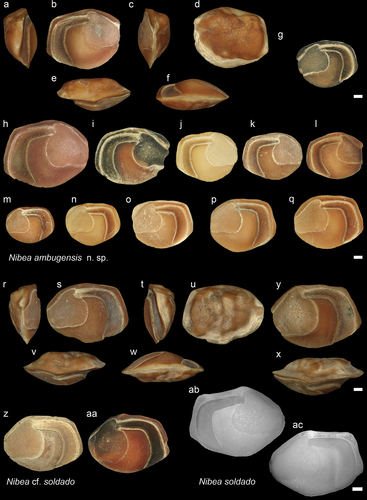
Figure 13. Sciaenidae. a-f: Nibea stintoni holotype from IB (NHMUK PV P 76676), posterior (a), inner (b), anterior (c), external (d), dorsal (e), and ventral (f) views; g-h: N. stintoni from JT1 (GUBD V0227) inner views; i: N. stintoni from AH (GUBD V0228). j-o: Nibea aff. chaoi from Dad, Miri Formation, anterior (j), inner (k), posterior (l), external (M), dorsal (n) and ventral (o) views (NHMUK PV P 76677); p-s: Nibea aff. chaoi from the Miri Formation, inner views, p s from Dad (NHMUK PV P 766778-79), and q & r from JT1 (GUBD V0230). Scale bar: 1 mm.
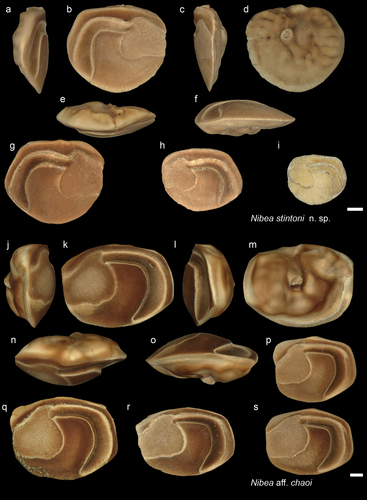
Figure 14. Sciaenidae. a-f: Protonibea nolfi from Tag Miri Formation (holotype - NHMUK PV P 76680), inner (a), external (b), anterior (c), posterior (d), dorsal (e), and ventral (f) views; g-j: P. nolfi two specimens from Dad (NHMUK PV P 76681-82), inner (g i) and dorsal (h j) views; k-m: P. nolfi two paratypes from AH, specimen 1 in inner (k) and dorsal (l) views (NHMUK PV P 76683), specimen 2 in inner view (m) (NHMUK PV P 76684); n: P. nolfi from Ker (GUBD V0233) in inner view. o-q: Protonibea sp. from AH (GUBD V0238); r-t: Protonibea diacanthus modern specimens from Taiwan and Brunei (r: CHLOL4622, TL: 53.5 cm, s: CHLOL5223, TL: 27.6 cm, and t: BRUGM-20220522- TL: 80 cm). Scale bar: 1 mm.
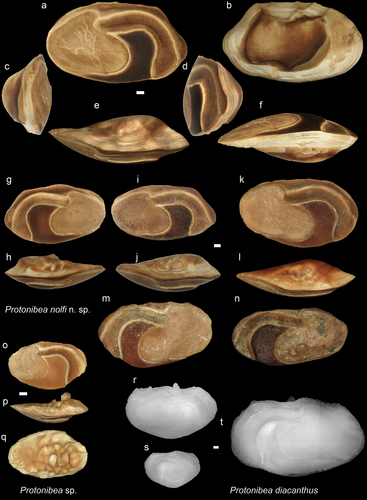
Figure 15. Sciaenidae. a-b: Otolithoides biauritus from AH (NHMUK PV P 76685), inner (a) and dorsal (b) views; c-f: Pterotolithus lateoides from AH, inner (c), external (d), dorsal (e), and ventral (f) views (NHMUK PV P 76686); c-f: P. lateoides from AH, inner (g), dorsal (h), ventral (i), and external (j) views (NHMUK PV P 76687). Scale bar: 1mm.
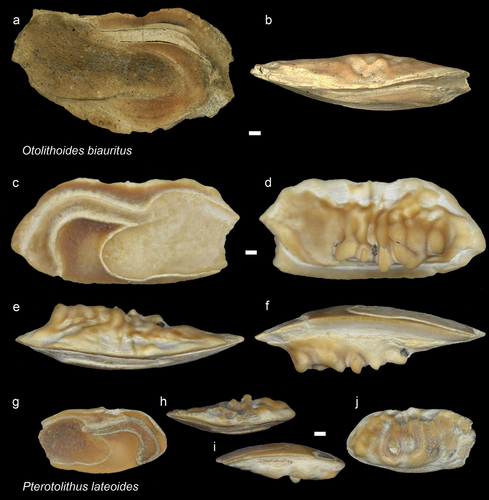
Figure 16. Sillaginidae, Haemulidae, and Cepolidae. a-f: Sillago sp. from AH (NHMUK PV P 76688) posterior (a), inner (b), anterior (c), external (d), dorsal (e), and ventral (f) views; g-h: Sillago sp. from Dad (NHMUK PV P 76689) inner (g) and dorsal (h) views; i-n: Pomadasys sp. from AH anterior (i), inner (j), posterior (k), external (l), dorsal (m), and ventral (n) views; o-r: Pomadasys sp. specimens in different sizes from AH inner views. All the figured Pomadasys are under NHMUK PV P 76690-94; s-u: Acanthocepola sp. from Dad (NHMUK PV P 76695). Scale bars: 1 mm.

The saccular otoliths (from now on referred to as sagitta/sagittae) are usually mesiodistally flattened, therefore inner (mesial) and external (distal) surfaces are defined. Morphometric parameters are the otolith length (OL), height (OH), and thickness (OT), which express anteroposterior, dorsoventral, and mesiodistal extensions, respectively. OL/OH and OL/OT are their respective ratios. In the case of the ariids, the utricular otoliths (from now on referred to as lapillus/lapilli) are the largest and described here. These otoliths are situated dorsoventrally in the skull (e.g. Ohe Citation2006), therefore these otoliths bear enlarged ventral and dorsal surfaces. Morphometric parameters are the otolith length (OL), width (OW), and thickness (OT), which express anteroposterior, mesiodistal, and dorsoventral extensions, respectively.
Systematic Palaeontology
Class Actinopteri Cope, 1871
Infraclass Teleosteii Müller, 1845
Order Anguilliformes Regan, 1909
Family Congridae Kaup, 1856
Genus Ariosoma Swainson, 1838
Ariosoma sp.
Material
3 otoliths from Seria Fm (AH).
Dimensions
OL: 2.79–3.48 mm, OH: 2.64–3.22 mm,1.7 OL/OH: 1.06–1.08.
Description
The specimens are quite worn but still show the characteristic features of a ‘congroid’ sulcus pattern with an anteriorly closed ostium and a vertical anterodorsal ostial channel (Schwarzhans Citation2019). The otoliths show a distinct mid-dorsal expansion and a deeply rounded ventral margin, which together with the S-shaped sulcus, characterises them as representing the genus Ariosoma.
Remarks
From Southeast Asia, eight modern species of Ariosoma are known (Froese and Pauly Citation2022). Fossil Ariosoma otoliths have been reported from Taiwan’s Late Miocene beds (Lin and Chien Citation2022: figs 4a-f), which show some resemblances to the specimens figured in our study, and from the Pleistocene of the Philippines (Van Hinsbergh and Helwerda Citation2019).
Order Clupeiformes Bleeker, Citation1859
Family Engraulidae Gill, 1861
Genus Setipinna Swainson, 1839
Setipinna cf. retusa Stinton, Citation1962
1962 - Setipinna retusa n. sp. Stinton Fig. 19.1.
Material
2 otoliths, 1 from the Miri Fm (IB) and 1 from the Seria Fm (Ker).
Dimensions
OL: 3.32–3.60 mm, OH: 2.66–2.85 mm, OL/OH: 1.25–1.26.
Description
The specimens have a straight, slightly concave dorsal margin, while the ventral margin is strongly convex. The posterodorsal and anterodorsal margins are inclined and straight. The inner surface is convex and bears a deep, straight sulcus. The ostium and cauda have approximately the same length, with a somewhat deeper ostium that opens anteriorly. The sulcus is bordered by strong superior and inferior crista.
Remarks
The species was described by Stinton (Citation1962) based on three specimens from Borneo. The otolith figured here, although quite worn, has all the diagnostic features of the holotype (Stinton Citation1962: fig.19.1 - NHMUK PV P 45561; Nolf Citation2013: plate 30), but the dorsal and the posterodorsal margins tend to be more concave on the type. Therefore, the specimen is described with open nomenclature. In addition, Stinton also reported a myctophid from Dent Peninsula in Sabah (Stinton Citation1962: figs 19.18, NHMUK PV P 45660, see also Supplementary material 2) that very likely represents a Setipinna specimen similar to the one described here, but the anterior end is broken off.
Setipinna sp.
Material
2 broken otoliths from the Miri Fm (Dad).
Dimensions
OH: 2.2–2.5 mm; OL: > 2.8–3.4 mm (note that the specimens are broken).
Description
The anterior part of these specimens is broken, but their preserved sulcus part, particularly the shape of the cauda and the presence of superior and inferior cristae are similar to that of S. cf. retusa (). However, these otoliths are more elongated, and their posterior end is more pointed.
Remarks
From Southeast Asia, four modern Setipinna species are known (Froese and Pauly Citation2022), but their relation to the fossil species needs to be studied in the future.
Genus Coilia Gray, 1830
Coilia planata Stinton, Citation1962
1962 - Coilia planata n. sp. Stinton Fig. 19.2.
2013 - Coilia planata in Nolf Plate 30.
Material
41 sagittae, 19 from the Seria Fm (AH:1, TB*: 10, Ker: 6, BP: 2) and 22 from the Miri Fm (IB: 1, JT1: 3, Dad: 18).
Dimensions
OL: ~3.6–8.46 mm, OH: 3.49–6.33 mm, OL/OH: 1.03–1.34 (note that the smallest specimen is broken).
Description
Subcircular, elongated otoliths with convex inner and external surfaces. The ventral margin is rounded, and the dorsal is rather angular. The otolith becomes narrower anteriorly with a concave or straight anterodorsal margin. The posterior margin is vertical/subrounded. The sulcus is straight, horizontal, with a longer cauda than the ostium (cauda/ostium: 1.26 ± 0.11 n = 5). The latter is open anteriorly. Well-developed crista inferior and superior follows the sulcus, that are respectively paralleled by ventral furrow and dorsal depressions. Two smaller otoliths with broken anterior parts, proportionally display more elevated dorsal otolith margins (), which may be taken as an ontogenetic feature of juvenile specimens or represent a different species.
Remarks
These are the most common anchovy otoliths from Brunei’s Late Miocene beds. Stinton (Citation1962) erected the species based on two specimens. The holotype (NHMUK PV P 45563, Stinton Citation1962: fig.19.2) shows a more circular appearance, but it is rather worn with its anterior end broken. Otherwise, all the characteristics of the specimens described here are similar.
Coilia cf. mystus (Linnaeus, 1758)
t-u
Material
1 left otolith from Seria Fm. (Ker).
Dimensions
OL: 1.14 mm, OH: 0.86 mm, OL/OH: 1.33.
Description
The otolith has a subcircular shape that is elongated anteriorly. The ventral margin is deeply rounded tapering anteriorly, and the posterior margin is straight and slightly curved, such as the dorsal margin. These margins partly or entirely bear large denticles or folds. The anterodorsal margin is straight and tilted, it shows a small notch (excisura) where the ostium opens anteriorly. The sulcus is straight and paralleled by a well-developed crista superior and inferior, similar to C. planata described above.
Remarks
The specimens resemble that of extant C. mystus (see Nolf Citation2013: plate 30), especially in terms of marginal folds, but the modern species has a more rounded dorsal margin. Fishbase reports nine extant Coilia species from the Southeast Asian region (Froese and Pauly Citation2022). The C. mystus is reported from the wider region and is known to enter estuaries (Froese and Pauly Citation2022). It must be mentioned that the otoliths of some engraulid species within the genera Coilia, Setipinna, and Thryssa can have similar features (see BRMAS, http://museum.biodiv.tw/eng).
Order Siluriformes Cuvier, Citation1817
Family Ariidae Bleeker, Citation1858
Sea catfishes are common marine coastal fishes in Southeast Asia today, with many taxa entering estuaries and brackish water, and a few adapted to an exclusive freshwater habitat (Kailola Citation1998; Froese and Pauly Citation2022). In ariids, the utricular otolith (lapillus) is the largest, which is very commonly found in Brunei’s Late Miocene beds. They are easily recognisable, and more than 200 specimens have been collected over the years. Stinton (Citation1962) described four new fossil species from Brunei based on a few specimens (Tachyurus oblongus (n = 12), Netuma radiata (n = 6), Netuma regularis (n = 1), and Arius cavatus (n = 1)). These otoliths are figured only in dorsal view; hence some crucial features, such as the arrangements of the ostium and cauda (after Ohe Citation2006), cannot be observed. Nolf (Citation2013) stated that these otoliths are probably not diagnostic at species level. Indeed, lower taxonomic rank (genus, species) of ariid otoliths can be difficult to assess, mainly due to similarities among the different taxa and the lack of a detailed morphological comparison of otoliths from a large database of modern species. Ohe (Citation2006) investigated and figured otoliths of some modern species from the region, with many also being common in Brunei. Based on Ohe’s paper and studying common ariids collected in Brunei’s waters, the fossil material is sorted into two morpho-groups: Rounded and Elongated. These generally correspond to Ohe (Citation2006)’s Clam- and Maculatus-groups, respectively. Within each group, we further distinguished two sub-groups. For illustration, the otoliths’ dorsal, ventral, and mesial views are shown in . Following the description of the morpho-groups, the otoliths are oriented in ventral view in a way that the anteromesial projection (Amp) and the posterior end of linea basalis (Lb) are aligned along a horizontal line (see ). Altogether 242 lapilli were collected, from which 164 were sorted into four Ariidae groups, two within the rounded and two within the elongated morpho-groups. With future investigation, it is anticipated that a lower level of taxonomy, at least at genus level, could be achieved, and the validity of Stinton’s fossil species could be further evaluated.
Ariidae indet. 1
Material
35 lapilli, 29 from the Seria Fm (AH) and 6 from the Miri Fm (Dad).
Dimensions
OL: 5.1–13.2 mm. OW: 4.3–11.1 mm, OL/OW: 1.20 ± 0.06.
Description
Rounded otoliths with similar length and width. These otoliths have a distal margin that is wide, convex, and symmetrically curved. The anterior and posterior margins are also convex. This makes them have a generally circular appearance except for their linear/concave anteromesial region. They often have the longest width measurement when compared to other morpho-groups.
Remarks
This morphotype closely resembles the common modern species in Brunei’s water, such as Hexanematichthys sagor (Hamilton Citation1822), Netuma bilineata (Valenciennes Citation1840), and N. thalassina (Rüppell Citation1837), but the southern New Guinean and northern Australian taxa of Hemiarius dioctes (Kailola Citation2000), H. stormii (Bleeker Citation1858), and Neoarius graeffei (Kner and Steindachner Citation1867) could be assigned to this group as well (see Ohe Citation2006: fig. 7). One of Stinton’s species, Arius cavatus, can be classified in this morpho-groups since its distal margin appears quite wide and the edge of its cauda is thick (see Stinton Citation1962: fig. 19.6 - NHMUK PV P 44076).
Ariidae indet. 2
Material
28 lapilli otoliths, 19 from the Seria Fm (AH: 18, BP: 1), 9 from the Miri Fm (JT1: 5, Dad: 3).
Dimensions
OL: 5.3–12.6 mm. OW: 4.1–9.8 mm, OL/OW: 1.25 ± 0.07.
Description
Rather diamond-shaped otoliths within the Rounded-group. These otoliths have a symmetrically convex mesial margin. The distal margin is narrower than those classified in Ariidae indet. 1, while the anterior margin is slightly concave towards the mesial end.
Remarks
These otoliths appear like the following modern species in the Southeast Asian region: Plicofollis argyropleuron (Valenciennes Citation1840), P. nella (Valenciennes Citation1840), and P. tonggol (Bleeker Citation1846) (see Ohe Citation2006: fig. 7). Two of Stinton’s species, Netuma radiata and N. regularis, can be classified in this morpho-group as they both appear to have a roughly symmetrical convex mesial margin and narrow distal margin (see Stinton Citation1962: fig. 19.4 – NHMUK PV P 44074–75, and fig 19.5 – NHMUK PV P 45566, respectively).
Ariidae indet. 3
Material
59 lapilli otoliths, 46 from the Seria Fm (AH: 41, Ker: 4, BP: 1), 13 from the Miri Fm (IB: 4, JT1: 7, Dad: 2).
Dimensions
OL: 4.7 –11 mm. OW: 3.4–8.3 mm, OL/OW: 1.34 ± 0.07.
Description
Elongated otoliths with higher length/width ratio range than those that are classified in Ariidae indet. 1 and 2. The distal margin is curved and protrudes more anteriorly that results a more enhanced concave anterior margin. It has three pronounced angular-like points at the posteromesial, anteromesial, and anterodistal margins.
Remarks
This morpho-group is comparable to several modern species in the Southeast Asian region, including Arius arius (Hamilton Citation1822), A. maculatus (Thunberg Citation1792), A. oetik Bleeker Citation1846, and Sciades sona (Hamilton Citation1822) (see Ohe Citation2006: fig. 7), and Batrachocephalus mino (Hamilton Citation1822) and Cryptarius truncatus (Valenciennes Citation1840) collected in Brunei’s waters (Johari et al. in prep).
Ariidae indet. 4
Material
42 lapilli otoliths, 12 from the Seria Fm (Ker: 11, BP: 1), 30 from the Miri Fm (IB: 2, JT2: 1, JT1: 13, Dad: 12, Tag: 2).
Dimensions
OL: 5–9.9 mm. OW: 3.4–7.7 mm, OL/OW: 1.31 ± 0.09.
Description
Elongated otoliths with a rather oval-shape. Their posterior margin can either be sharply or smoothly curved, while the anterior margin is even more concave than that of Ariidae indet. 3.
Remarks
This group is comparable to the local modern species of Nemapteryx nenga (Hamilton Citation1822) (Ohe Citation2006). Stinton’s Tachysurus oblongus can be classified in this morpho-group due its elongated and sharply curved posterior margin, and its strongly concave anterior margin (see Stinton Citation1962: fig. 19.3 – NHMUK PV P 44059).
Order Gadiformes Goodrich, 1909
Family Bregmacerotidae Gill, 1872
Genus Bregmaceros Thompson, 1840
Bregmaceros sp.
Material & Origin
5 sagittae, 3 from the Seria Fm (AH) and 2 from the Miri Fm (IB).
Dimensions
OL: 1.00–1.33 mm, OH: 0.91–1.26 mm, OL/OH: 1.06–1.10.
Description
Thin otoliths that are flat on the interior side, while somewhat convex on the external surface. The genus shows a characteristic sulcus with small, oval, deepened colliculi in ostium and cauda separated by a collum, which is underlain by a small pseudocolliculum. The ventral area has radial ridges from the centre. The better-preserved specimen from Brunei () displays large denticles on the dorsal margin. The posterior margin is concave with a dorsal notch and ventral protrusion. The anterior margin shows an opposite arrangement. The ventral margin is convex, with four angles, two prominent ones at the anterior and posterior margins, one in the middle, and one closer to the posterior end.
Remarks
Codlets are small (<12 cm) pelagic-neritic fishes with six possible modern species occurring in the Southeast Asian region, two that are considered rather typical IAA species, the B. lanceolatus Shen 1960 and B. rarisquamosus Munro Citation1950 (Froese and Pauly Citation2022). Fossil Bregmaceros species were reported from the Pleistocene of the Philippines (Van Hinsbergh and Helwerda Citation2019).
Order Kurtiformes Jordan, 1923
Family Apogonidae Günther, 1859
Apogonidae indet.
Material
5 sagittae from the Seria Fm (AH).
Dimensions
OL: 1.21–4.3 mm, OH: 0.94–3.56 mm, OL/OH: 1.21–1.29.
Description
The inner surface is flat or slightly convex. The ventral margin is rounded with some small folds towards the posterior end. Depending on the preservation state, a predorsal lobe is apparent from which the dorsal margin inclines posteriorly. The sulcus is wide with a straight cauda, and an even wider ostium inclined slightly dorsally and open anteriorly. A dorsal depression is evident above the sulcus.
Remarks
Stinton (Citation1962) described the species Apogon spictatus from the Togopi Fm at the Dent Peninsula, Sabah, Borneo, based on one specimen. In the NHMUK collection, five otoliths are kept together (NHMUK PV P 45670–74), among them the holotype (NHMUK PV P 45670). The drawn holotype in Stinton’s work is a bit misleading (1962: fig 19.10), but the original specimens is shown in the Supplementary material 2). One of the Brunei otoliths () show resemblance to this taxon, and hence may represent the same genus (i.e. Apogon) and probably a closely related species. The other specimens may also represent the genus Apogon. However, cardinal fishes have over 200 species within 34 genera in the wider Indo-West Pacific region (Froese and Pauly Citation2022), hence more detailed study on extant faunas is necessary to further evaluate these specimens. Fossils Apogonidae were also reported from Taiwan (Lin et al. Citation2018; Lin and Chien Citation2022), the Philippines (Van Hinsbergh and Helwerda Citation2019) and from India (Carolin et al. Citation2023).
Order Gobiiformes Günther, 1880
Family Gobiidae Cuvier Citation1816
Gobiidae indet.
Material & Origin
23 sagittae, 17 from the Seria Fm (AH:15, Ker:2) and 6 from the Miri Fm (IB:5, Dad:1).
Dimensions
OL: 0.91–1.24 mm, OH: 0.94–1.24 mm, OH/OL: 0.97–1.00.
Description
The otoliths have a square/rectangle shape with posterodorsal protrusion and well-developed dorsal depression. The area around the sulcus is elevated. The cauda is narrower than the ostium, which inclines and widens ventrally. One specimen (), with its postdorsal protrusion broken off, shows a different anterior margin: it has three small crenulations in the ventral corner, and a small protrusion anterodorsally.
Remarks
Gobiids are one of the most species-rich fish families in the tropical-subtropical region of the Indo-West Pacific, with nearly 600 species (Froese and Pauly Citation2022). Their relatively low abundance in our record could be related to collecting bias, as most of the large otoliths were surface collected and/or to taphonomy and preservation bias for the generally small-sized otoliths, and other microfossils (Roslim et al. Citation2019). Several fossil species have been erected from Southeast Asia (Sumatra (1): Frost (Citation1925); Java (5): Vorstman (Citation1927); Borneo (1): Stinton (Citation1962). However, the validity of these might be questioned in the view of our limited knowledge on the vast amount of extant Gobiidae taxa. Nolf (Citation2013) stated that the fossil species cannot be evaluated based on the illustrations in this early literature. Fossil Gobiidae was recently reported from Taiwan (Lin et al. Citation2018; Lin and Chien Citation2022) and the Philippines (Van Hinsbergh and Helwerda Citation2019). The general morphology of most of our specimens resembles that of Stinton (Citation1962: fig.19.14, Gobius tankilensis Vorstman Citation1927 - NHMUK PV P 45661), Note, it is misspelled as ‘tankilesis’ in Stinton’s work). He also reported three other species from Borneo, but those have not been found in our record. Our gobiid otoliths from Brunei probably represent 2–3 different species.
Order incertae sedis in Carangaria Betancur-R. et al., 2017
Family Lactariidae Boulenger, 1904
Genus Lactarius Valenciennes, Citation1833
Lactarius lactarius (Bloch and Schneider, 1801)
2013 - Lactarius lactarius in Nolf Plate 238.
Material
36 sagittae, 35 from the Seria Fm (AH) and 1 from the Miri Fm. (Dad).
Dimensions
OL: 5.1–12.7 mm, OH: 4.6–9.1 mm, OL/OH: 1.11–1.40.
Description
Thin set, rhomboid-like shaped otoliths with convex inner and concave or flat external surfaces. The dorsal and ventral margins sometimes bear small notches. The dorsal edge is angular, while the ventral one is more curving with anteroventral extension. The ostium is large and opens widely anteriorly/anterodorsally. The cauda is straight with a slight curve at the posterior end, and it is paralleled by prominent superior and inferior cristae. One small specimen (OH: 0.98 mm, OL: 1.14 mm) has been discovered (), which has the general features of these otoliths, however the cauda is entirely straight, and the ostium is incised by an excisura. We have tentatively assigned this specimen to L. lactarius.
Remarks
The fossil otoliths have all the characters of the modern species Lactarius lactarius (), hence they are classified under this taxon, which are commonly called False trevally. This is the only extant species of the genus, and it is commonly found in coastal waters of Southeast Asia (Froese and Pauly Citation2022). On the other hand, there are several fossil species described (Nolf Citation2013: plates 238–239), even from the IAA, such as the middle Eocene species of L. nonfungus (Nolf and Bajpai Citation1992).
Order incertae sedis in Eupercaria Betancur-R. et al., 2017
Family Sciaenidae Cuvier, Citation1829
Genus Advenasciaena gen. nov.
Type species
†Advenasciaena bruneiana sp. nov.
Name
Advena refers to the strange, alien characteristic of these new sciaenid otoliths.
Diagnosis
Thickly built otoliths with an elevated central part in the inner surface. The external surface is irregular but very thick, sometimes with protuberances. The shape of the otolith is oval with a nearly straight dorsal margin with minuscule rounding in the centre, while the posterior, ventral, and anterior margins are smoothly curved. The sulcus is very special. The ostium has an anterior deep groove extending to the margin between the inner and external surfaces and a very shallow, triangular posterior part just under the anterior part of the cauda. It is filled with an ostial colliculum, which runs parallel with the cauda and extends to one-third of its length. The deep groove in the ostium is hardly noticeable in many of the more worn and small specimens. It is best observed in the holotype (). The cauda is also special. It is very deep, with its upper, horizontal part starting nearly at the top of the ostial margin. Its posterior part runs in the ventral direction and gradually widens till the ventral rim. The posterior tip of the cauda is slightly turned anteriorly.
Remarks
The very deep anterior groove and shallow triangular colliculum in the ostium and the coupled very deep, strongly expressed cauda are unique features of this genus. The inner surface of the otoliths of the Stellifer and Bairdiella groups from the Americas (Schwarzhans Citation1993: figs. 382–397) show some superficial resemblance to Advenascianea, mainly because of their deep cauda and inclined ostium. However, the shape and morphology of these otoliths and the position of their ostium are completely different from that of Advenascianea.
Distribution
From late Middle Miocene to Late Miocene layers of the Seria and Miri formations in Brunei Darussalam.
Advenasciaena bruneiana sp. nov.
Name
Referring to Brunei Darussalam, the country of origin.
Holotype: right otolith NHMUK PV P 76639 ().
Type locality
Tanjong Nangka village near Jalan Tutong, Bandar-Muara District, Brunei Darussalam.
Age
Late Miocene, Miri Fm.
Paratypes
ASIZF 0100316 () from Tanjong Nangka village, RGM.1364031 () from Dadap, and NHMUK PV P 456676 from Tutong (Supplementary material 2).
Other material investigated
Miri Fm: Another 5 specimens Dadap outcrop (GUBD V0167). Seria Fm: 2 from Bukit Panggal (GUBDV0168); 1 from Keriam village (GUBDV0169) and 5 from Ambug Hill (GUBDV0170). In addition, 3 reworked specimens from Tutong beach (TB*) (GUBD-V171).
Dimensions
OL/OH: 1.1–1.4 (average 1.2 ± 0.1, n = 13); OL/OT: 1.9–2.6 (average 2.3 ± 0.2, n = 13). The smallest specimens have a length of 6.3–6.5 mm, while the largest specimen is 11.6 mm with a thickness of 5.6 mm.
Description
For detailed characteristics of these otoliths see the genus description.
Diagnosis
The deep anterior groove () and shallow triangular colliculum in the ostium (), and the coupled very deep, strongly expressed cauda () are unique features of this genus and species.
Remarks
In the collection of the Natural History Museum in London the specimen NHMUK PV P 45676 (listed here among the paratypes) was originally catalogued as Pseudosciena angulata, however it clearly represents A. bruneiana (Supplementary material 2).
Distribution
From late Middle Miocene to Late Miocene layers of the Seria and Miri formations in Brunei Darussalam.
Genus Atrobucca Chu, Lo and Wu, 1963
Atrobucca borneensis sp. nov.
Name
Refers to the origin of the otoliths as they are found on Borneo Island.
Holotype
left otolith NHMUK PV P 76640 ().
Type locality
Ambug Hill, Tutong District, Brunei Darussalam.
Age
Late Miocene, Seria Fm.
Paratypes
four specimens NHMUK PV P 76641–44 () from Ambug Hill, Tutong District, Brunei Darussalam.
Other material investigated
224 additional specimens from the Seria Fm (AH) (GUBD V0196) and 4 specimens from TB* (GUBD V0197).
Dimensions
OL: 4.4–11.4 mm, OH: 3–9.1 mm, OT: 1.0–4.8 mm. OL/OH: 1.2–1.3; OH/OT: 1.7–2.4.
Description
Moderately thickset otoliths with a convex, smooth inner surface and an irregular external surface. The otolith is much thicker centro-dorsally and thins ventrally. The dorsal margin is horizontal in the centre, and slightly curves anteriorly, while its posterior quarter is inclined at 30–40 degrees. The ventral margin is deep, smooth anteriorly, and slightly concave posteriorly, where it bears a notch just at the deepest end of the cauda. The smaller specimens lack this notch, and the ventral margin is rather convex posteriorly. The ostium is spoon-shaped with a moderate postostial lobe, bears a small precaudal depression, and narrows to the anterior edge. The anterior part of the cauda is horizontal, and then posteriorly curves at 30–40 degrees and sometimes ends with a buffer-shaped tip.
Diagnosis
These Atrobucca otoliths have a relatively low length-height ratio (1.2–1.3) and a strongly curved cauda compared to other species. The buffer-shaped end of the cauda is less expressed than at other species. It is often not clearly seen, but the slight posterior widenings are visible under the microscope for most of the studied specimens.
Remarks
The otolith of the modern species A. nibe (Jordan and Thompson, Citation1911) is the most similar to A. borneensis (see Schwarzhans Citation1993: figs 356–357). However, the fossil species is shorter and has OL/OH ratio of 1.1 to 1.3 with a strongly curved cauda. The OL/OH ratio for A. nibe is 1.5 (e.g. Schwarzhans Citation1993). The smaller specimens are more affected by diagenesis and have a more worn appearance, but still the key features can be observed.
Distribution
Brunei Darussalam Seria Fm: Ambug Hill (AH) and reworked specimens at the nearby Tutong beach (TB*).
Atrobucca aff. borneensis
Material
2 saggitae from Seria Fm. (AH).
Dimensions
OL: 7.2 & 9.4 mm, OH: 5 & 6.8 mm, OT: 2.1 & 3.1 mm, OL/OH: 1.4; OH/OT: 2.2–2.4.
Description
These otoliths have very similar characters to A. borneensis, however, they appear slightly more elongated and have a higher OL/OH ratio (1.4).
Remarks
The shape of the otoliths also resembles that of the modern species A. nibe and A. brevis (see Schwarzhans Citation1993), but the anterior horizontal part of the cauda is shorter, which is a feature similar to what is seen in A. borneensis. This latter species is abundant in our fossil record, and there is no large variation in their otoliths’ OL/OH ratios. Therefore, either these elongated otoliths represent rare, elongated endmember specimens of A. borneensis, or another species.
Genus Boesemania Trewavas, Citation1977
Boesemania obesa (Stinton Citation1962)
1962 - Otolithoides obesus n. sp. Stinton Fig. 19.12.
1993 - Boesemania obesa in Schwarzhans Figs 234–235.
2013 - Boesemania obesa in Nolf Plate 271.
Material
60 sagittae, 15 from the Seria Fm (AH: 3, Ker: 12); 16 from the Miri Fm (JT1: 14; Dad: 2) and 29 reworked specimens from TB*.
Dimensions
OL: 8–13.6 mm, OH: 5.4–7.9 mm, OL/OH: 1.48–1.72.
Description
Thickset, elongated otoliths with convex inner and external surfaces. The dorsal and ventral margins are rather straight. The anterior one is blunt with an extended predorsal margin. The posterior end is pointed. The ostium is small, inclined downwards by about 40°. The cauda is long, rather narrow close to the ostium, then it widens posteriorly and steeply curved in an U-shape, finally terminating with a pointed end.
Remarks
Boesemania today is a monospecific genus with Boesemania microlepis (Bleeker, Citation1858) living in the freshwater of Southeast Asia (Froese and Pauly Citation2022). The pointed posteroventral margin is characteristic of this genus, and the modern species has a proportionally larger, less inclined ostium (). The holotype of B. obesa is housed in the NHMUK collection (NHMUK PV P 44081, see Stinton Citation1962: fig.19.12, and Supplementary material 2). Another eight specimens of Stinton are in the NHMUK, partially revised by Schwarzhans (Citation1993), and were assigned to this species (e.g. NHMUK PV P 44077, P 44078, P 44082). However, NHMUK PV P 46610 and P 46611 clearly belong to different taxa (see later under Nibea and Protonibea).
Genus Bruneisciaena gen. nov.
Type species
†Bruneisciaena schwarzhansi sp. nov.
Name
Refers to the geographic origin of the new sciaenid genus (Brunei Darussalam).
Diagnosis
Thin-set sciaenid otoliths, with the thinnest part at the anterodorsal margin. The inner surface is slightly convex and smooth, while the external one is irregular. The dorsal margin is horizontal, inclines at the posterior end and forms a curved/pointed posterior margin. The ventral margin is straight or slightly curved in its central and posterior portions, but in the anterior direction it converges dorsally, forming an anterodorsal extension. The ostium is large, extends to the anterior and anteroventral margins, and covers most of the ventral area with a large postostial lobe. The latter can proportionally be larger than the anterior part of the ostium. The ostium bears a small precaudal depression. The cauda is moderately deep. Its anterior part is straight, widens posteriorly, then curves more than 90 degrees, and reaches towards the ventral margin.
Remarks
The thin-built, anterodorsally extended, and hence proportionally higher posteriorly than anteriorly otoliths are different from any genus known so far (see Schwarzhans Citation1993; Nolf Citation2013; Aguilera et al. Citation2016). Some of the Cynoscion, Otolithoides, and Atrobucca species have similar thin-set otoliths and big postostial lobe, but the elongate shape of the cauda and the dorsally oriented collum are clearly different in Bruneisciaena (see Schwarzhans Citation1993, p. 47–48 for C. acoupa, C. microlepidotus, Schwarzhans Citation1993, p. 207 for O. biauritus, and, Schwarzhans Citation1993, p. 201 for A. adusta). On the other hand, the South American freshwater genera Pachypops and Plagioscion (Schwarzhans Citation1993, pp. 28–34) has otoliths with similar long cauda, but again they have an ostium that differs from that of Bruneisciaena.
Distribution
From late Middle Miocene to Late Miocene layers of the Seria and Miri formations in Brunei Darussalam.
Bruneisciaena schwarzhansi sp. nov.
Name
In honour of Werner Schwarzhans (Hamburg, Germany) due to his great works on sciaenid otoliths.
Holotype
left otolith NHMUK PV P 76646 ().
Type locality
Dadap village, Bandar-Muara District, Brunei Darussalam.
Age
Late Miocene, Miri Fm.
Paratypes
ASIZF 0100315 () from Dadap village Brunei Darussalam, Miri Fm, RGM.1364030 () from Bukit Panggal, Brunei Darussalam, Seria Fm, and NHMUK PV P 51234 Tutong road, Seria Fm.
Age
Late Miocene, Seria Fm.
Other material investigated
Miri Fm: 7 other specimens from the type locality (six figured ) (NHMUK PV P 76647–49 - Figs 9n-p; GUBDV0161 – Figur 9q-s), 3 from Tanjong Nangka outcrop (GUBDV0162), and 1 from Ikas Bandung (GUBDV0163). Seria Fm: 1 from Bukit Panggal (GUBDV0160), 2 from Keriam village (GUBDV0164) and 3 from Ambug Hill (GUBDV0165). In addition, 6 reworked specimens from Tutong beach (GUBDV0166).
Dimensions
OL/OH: 1.4–1.8 (average 1.6 ± 0.1, n = 13); OL/OT: 4.6–6.3 (average 5.3 ± 0.5, n = 13).
Description
The taxon is represented by a wide size range in our record, but getting the exact length of the specimens is sometimes difficult because the thinnest anterodorsal part of the otolith is often broken or worn. The smallest specimen recovered has a length of 7.5 mm, while based on the average OL/OH ratios, the largest specimen is estimated to reach over 21 mm. For detailed characteristics of these otoliths, see the genus description.
Diagnosis
The laterally flattened, thin, but anterodorsally extended and narrowed otoliths are different from any other known sciaenid genus. The shape of the ostium, the large postostial lobe, and the curvature of the cauda differentiate this species from other thin-built sciaenid otoliths.
Remarks
Variation seems to occur in the ventral margin of the postostial lobe, which most of the time, curves smoothly and is convex. However, for some specimens, it is rather straight or slightly concave (e.g. ). One specimen was deposited in the NHMUK collection (PV P 51234) and catalogued under the name Pseudosciaena acutirostris n. sp. probably by Stinton. However, this species name has not been published or reported anywhere and hence, it is not available according to the rules of the ICZN. Here we suggested this specimen to be one of the paratypes of Bruneisciaena schwarzhansi.
Distribution
From late Middle Miocene to Late Miocene layers of the Seria and Miri formations in Brunei Darussalam.
Genus Larimichthys Jordan and Starks, 1905
Larimichthys polyactis (Bleeker, 1877)
1993 – Larimichthys polyactis in Schwarzhans Figs. 370–371.
2011 – Larimichthys polyactis in Ohe Figs. 9.
2022 – Larimichthys polyactis in Lin and Chien Figs. 10f-i.
Material
114 sagittae, 109 from the Seria Fm (AH:108, JPB:1), 5 reworked specimens from TB*.
Dimensions
OL: 6.2–11.8 mm, OH: 4.1–6.8 mm, OT: 2.3–5 mm. OL/OH: 1.5–1.8; OH/OT: 1.2–1.7.
Description
Elongated, thick otoliths with a slightly convex and rather smooth internal surface. The external surface bears a strong, well-developed posteroventral umbo. The otoliths’ height is more considerable anteriorly, with both dorsal and ventral margins converging posteriorly. The convergence is smoother on the ventral margin, while sometimes stepwise on the dorsal one. The ostium is fairly circle-shaped with no or very minor precaudal depression. It is anteriorly closed, which is evident only on better preserved specimens. The anterior part of the cauda is straight and posteriorly ends in a round-shaped tip, within it, there is a smaller circular colliculum. A deep dorsal depression parallels the straight part of the cauda, while the ventral area is smooth.
Remarks
The otoliths are considered to represent the modern species L. polycatis due to the resemblance of all the features described above. However, this and the other two extant species, L. crocea (Richardson, Citation1846) and L. pamoides (Munro, Citation1964), are not reported today around Borneo (Froese and Pauly Citation2022). The otoliths of L. crocea bear a posterior tip which distinguishes this species from the others (Schwarzhans Citation1993: figs. 368–369). On the other hand, there is no available otolith specimen to compare with L. pamoides, which occurs in north-western Australia and Papua New Guinea. There is a recently described species, L. terengganui Seah et al. (Citation2015), that is a common form around Malaysia with probable occurrence in northern Borneo (Seah et al. Citation2015). However, based on molecular data, this taxon was suggested to be differentiated as Pseudolarimichthys terengganui (Lo et al. Citation2017). In the general features the otolith of this species (Seah et al. Citation2015: fig. 4) resembles the Bruneian fossil otoliths described here. However, there is no observable distinct deepened colliculum within the rounded end of its cauda. Considering the fossil record, L. polycatis is also known from Japan (Ohe Citation2011) and Taiwan (e.g. Lin and Chien Citation2022).
Genus Dendrophysa Trewavas, Citation1964
Dendrophysa angulata (Stinton Citation1962)
1962 - Pseudosciaena angulata n. sp. Stinton Fig. 19.13.
1993 - Boesemania angulata in Schwarzhans page 128 Figs 236–238.
2013 - Boesemania angulata in Nolf Plate 271.
Material
108 sagittae, 72 from the Seria Fm (AH: 66, Ker: 5, BP: 1), 19 from the Miri Fm (IB: 3, JT2: 1, JT1: 9, Dad: 5, Tag: 1) and 17 reworked specimens from TB*.
Dimensions
OL: 5.1–11.3 mm, OH: 3.6–8.2 mm, OL/OH: 1.38–1.42.
Description
Thick, posteriorly angular otoliths, with much thinner, curved anteroventral edge. The posterodorsal margin is straight or very slightly concave, while the anterodorsal edge is rather high with a well-developed predorsal margin. The ventral margin is curved, especially anteriorly. The external surface bears irregular tuberculation. The internal surface is convex, and most of the area is covered by a large sulcus. The ostium is large with a postostial lobe, then narrows anteriorly, and opens anterodorsally. The cauda is deep and follows the posterior margin and widens slightly then strongly curves back in the anterior direction (i.e. U-shaped).
Remarks
The species was originally described under the genus Pseudosciaena by Stinton (Citation1962) (NHMUK PV P 44084), then later reported under Boesemania together with the other earlier described fossil Bornean sciaenid species B. obesa (Schwarzhans Citation1993; Nolf Citation2013). One of the key features of Boesemania otoliths is the large, pointed posterior tip, which is completely absent from the specimens described here. Schwarzhans (Citation1993) pointed out that the ostium is also somewhat larger than that of Boesemania, but argued that the sulcus has still the characteristic of genus. On the other hand, these otoliths show resemblance to those under the Nibea-group (see Schwarzhans Citation1993). The strongly curved cauda and the extended, thinned anteroventral margin is more similar to the status found in the genus Dendrophysa. There is only one known extant species, D. russelii (Cuvier, Citation1829) () that lives in the coastal regions and estuaries in Southeast Asia (Froese and Pauly Citation2022). The otoliths of D. angulate are often more elongated, especially the specimens from the Miri Fm. (see ), and generally have a longer ostium than the modern species.
It is worth mentioning that in the NHMUK collection, other taxa were catalogued under the name Pseudosciena angulatea. One such specimen is the earlier mentioned Advenasciaena bruneiana (NHMUK PV P 45676), while an otolith from the Dent Peninsula in Sabah (NHMUK PV P 46615) clearly belongs to the extant genus Pennahia (see Supplementary material 2).
Dendrophysa sp.
()
Material
6 sagittae, 2 from the Seria Fm (Ker), 1 from Miri Fm (IB), and 3 reworked from TB*.
Dimensions
OL: 6.1–7.7 mm, OH: 4.6–5.8 mm, OL/OH: 1.33–1.40.
Description
These otoliths are thinner, and less elongated than D. angulata. Their ostium is longer and resembles D. angulata. They have a prominent anterodorsal crest.
Remarks
All these otoliths either represent specimens with a marginal morphology within the species D. angulata, or represent a closely related new species within the Nibea-group.
Genus Nibea Jordan and Thompson, Citation1911
Nibea ambugensis sp. nov.
Name
Derived from the locality of Ambug Hill (Tutong District, Brunei Darussalam) where these otoliths are the most abundant.
Holotype
left otolith NHMUK PV P 76664 ().
Type locality
Ambug Hill (AH), Bandar-Muara District, Brunei Darussalam.
Age
Late Miocene, Seria Fm.
Other material investigated
269 sagittae, 254 from the Seria Fm (JPB: 1 - GUBD V0219, AH: 252- GUBD V0218 & NHMUK PV P 76665–72 (), Ker: 1 - GUBD V0220), 2 from the Miri Fm (JT1, Dad – GUBD V0221-V0222) and 13 reworked specimens from TB* (GUBD V0223).
Dimensions
OL: 4.7–9.3 mm, OH: 3.5–6.7 mm, OL/OH: 1.39–1.43.
Description
Rather square-shaped otoliths with angular posterior and dorsal margins, and a curved ventral margin. They are blunt anteriorly. The otoliths are thick, but they are thin anteroventrally. The external surface is irregular, while the inner surface is convex and rather smooth. The ostium occupies the anterodorsal area with a small postostial lobe that narrows and opens anteriorly. The cauda widens posteriorly, it follows the posterior margin and turns slightly in the anterior direction at its end.
Diagnosis
The general features of these otoliths are similar to those of D. angulata (i.e. Nibea-group see Schwarzhans Citation1993), but their ostium is a bit smaller, less elongated, and the postostial lobe is minor. In addition, the tip of the cauda does not curve as much back anteriorly (i.e. L-shaped rather than U-shaped). These features place these otoliths under the genus Nibea. The shape of the otolith and its OL/OH ratio makes N. ambugensis different from other known Nibea otoliths (e.g. Schwarzhans Citation1993). Nibea chui (Trewavas, Citation1971) is the one that most resembles our otoliths (see Schwarzhans Citation1993: fig. 307), but its posterior margin is relatively vertical and slightly concave, while its ventral area looks smaller when compared to N. ambugensis.
Remarks
These otoliths are among the most common ones in Brunei’s deposits, particularly in the Seria Fm. Currently ten modern Nibea species are known (Froese and Pauly Citation2022). The most similar species, N. chui, occurs today in more temperate waters around Japan, Korea, and China (Froese and Pauly Citation2022). It must be mentioned that the specimen of N. soldado (Lacepède, Citation1802) illustrated by Schwarzhans (Citation1993: fig. 309) is also somewhat alike to our otoliths, but it has smaller and shorter ostium. Moreover, this specimen is less elongated than other figured N. soldado specimens (see , also Wong et al. Citation2016: fig. S1), and therefore it is considered to represent a different species.
Nibea cf. soldado (Lacepède, Citation1802)
Material
24 sagittae, 22 from the Seria Fm (AH: 21, Ker: 1), and 2 reworked specimens from TB*.
Dimensions
OL: 6.6–9.6 mm, OH: 5.0–7.2 mm, OL/OH: 1.41–1.49.
Description
The otoliths are more robust and elongated than the previous species (N. ambugensis). Otherwise, many features are similar.
Remarks
These otoliths clearly belong to the genus Nibea. The collected modern specimens of N. soldado (, and Wong et al. Citation2016: fig. S1) have a similar shape and length -height proportion (1.43 ± 0.04), but as the recovered fossil specimens are quite worn, they are described as Nibea cf. soldado at this stage. Moreover, there are other potential Nibea species in the region whose otoliths have not yet been investigated in detail (e.g. N. coibor (Hamilton, Citation1822)), and therefore the taxonomy of these specimens may be revised in the future.
Nibea stintoni sp. nov.
Name
In memory of Fred C. Stinton who first worked on fossil otoliths from Brunei.
Holotype
left otolith NHMUK PV P 76676 ().
Type locality
Ikas Bandung (IB), Bandar-Muara District, Brunei Darussalam.
Age
Late Miocene, Miri Fm.
Other material investigated
5 sagittae, 2 specimens from the Miri Fm (JT1-L2, GUBD V0227), a juvenile specimen from the Seria Fm. (AH, GUBD V0228), and two possible reworked worn specimens from TB* (GUBD V0229).
Dimension
OL: 4.1–6.2 mm, OH: 3.5–5.6 mm, OL/OH: 1.11–1.17.
Description
Thin, circular-shaped otoliths. The inner surface is convex, while the external is flat/convex with irregular folds. The dorsal margin is straight, slightly rounding in the centre. The posterior end is angular dorsally till the upper half/third of the margin, and then it curves and smoothly joins a deep ventral margin. The type specimen bears small regular ridges at the ventral margin. The anterior margin is regularly upward oriented. The ostium is relatively small, opens anteriorly, and is followed by a predorsal margin. It has a minor postostial lobe. The cauda has a short horizontally oriented portion and then curves downwards. The angle between the horizontal and the downwards oriented part is higher than 90°.
Diagnosis
The laterally compressed, circular-shaped otoliths with deep, centrally oriented ventral margin make N. stintoni different from other Nibea species.
Remarks
A specimen from the NHMUK collection (PV P 46610) belongs to this species. It was catalogued under the name ‘Sciaena’ ovalis n. sp., most probably by Stinton. However, this species name has not been published or reported anywhere, hence it is not valid according to the rules of the ICZN. Schwarzhans (Citation1993) listed this NHMUK specimen under Boesemania obesa. The shapes of the otolith, ostium, and cauda are completely different from B. obesa. The fossil otoliths of Nibea chaoi Lin and Chien (Citation2022) are similar (Lin and Chien Citation2022), but N. stintoni has a more extended ventral area with a more curved ventral margin, and a cauda that bends slightly differently. It must be mentioned that the otolith figured in Schwarzhans (Citation1993: fig. 309) as N. soldado, is also similar to N. stintoni, but the figured specimen is perhaps misidentified and belongs to another species (see previous discussion and images of N. soldado in Wong et al. Citation2016: fig. S1; ). The otolith of N. soldado has a much greater thickness, it is longer than it is high, and has a larger ostium and deeper cauda.
Distribution
From late Middle Miocene to Late Miocene layers of the Seria and Miri formations in Brunei Darussalam.
Nibea aff. chaoi Lin and Chien, Citation2022
Material
11 sagittae, 5 from the Miri Fm (JT1: 2, Dad: 3) and 6 reworked from TB*
Dimensions
OL: 5.9–8.2 mm, OH: 4.8–6.2 mm, OL/OH: 1.23–1.32.
Description
Rectangular shaped otoliths with convex inner and external surfaces. The dorsal margin is straight or slightly rounded towards the centre. The anterior and posterior margins are rather straight and nearly parallel, while the ventral margin is curved. An anterodorsal crest is visible above the ostium. The ostium is small, spoon-shaped, with a minor postostial lobe. The anterior part of the cauda is straight and posteriorly curved in an U-shape.
Remarks
These otoliths resemble that of Nibea chaoi described from Taiwan (Lin and Chien Citation2022), but the Brunei specimens are somewhat more elongated, and their caudal tip curves forward more strongly. This later feature also makes them similar to the genus Dendrophysa (see ), but the anterior part of their cauda is rather horizontal, and their smaller ostium clearly distinguishes them.
Nibea spp.
Material
37 sagittae from the Seria Fm (AH).
Remarks
Worn and broken specimens with Nibea features, but they cannot be assigned under the discussed taxa with certainty due to their preservation state.
Genus Protonibea Trewavas Citation1971
Protonibea nolfi sp. nov.
2022 – Protonibea aff. diacanthus in Lin and Chien, page 14, Figure 12o-p.
Name
In honour of Dirk Nolf for his major contributions to otolith research.
Holotype
right otolith NHMUK PV P 76680 ().
Type locality
Tagap (Tag), Bandar-Muara District, Brunei Darussalam.
Age
Late Miocene, Miri Fm.
Paratypes
Two specimens from Dadap village Brunei Darussalam, Miri Fm NHMUK PV P 76681–82 (), and two specimens from Ambug Hill, Brunei Darussalam, Seria Fm NHMUK PV P 76683–84 (.
Age
Late Miocene.
Other material investigated
42 sagittae, 33 from the Seria Fm (AH: 31 - GUBD V0234, Ker: 2 - GUBD V0233 - ), 2 from the Miri Fm (Dad: 1 - GUBD V0236, Tag: 1 - GUBD V0235), and 7 reworked specimens from TB* (GUBD V0237).
Dimensions
OL: 9.1–17.3 mm, OH: 5.4–10.6 mm, OL/OH: 1.63–1.69.
Description
Elongated, thickset otoliths with convex smooth inner surface and with a big umbo in the external surface, located somewhat to the posterior end. The dorsal margin is horizontal in the centre, while it inclines slightly downwards in its anterior and posterior portions. The ventral margin is very gently curved, with the deepest part near the middle. The posterior margin is angular with a posteroventral protrusion, but in some specimens, the margin can be rather curved (). The sulcus has a large spoon-shaped ostium with a large postostial lobe and minor precaudal depression. The ostium is closed anteriorly, but it reaches the very border of the anterior and anteroventral margins. The anterior part of the cauda is slightly dorsally directed, and its posterior portion strongly curves in a ventral direction. The lower end of the cauda saliently widens and ends with a tip that is directed anteriorly.
Diagnosis
These otoliths are similar to those of the Recent species Protonibea diacanthus (Lacepède Citation1802), but the ostium seems to be more compact and shorter with a more moderate postostial lobe. These features are most remarkable on the stratigraphically older specimens of the Miri Fm (see holotype and ). The recent species often have small folds, ridges on their posterior, ventral, and anterior margins (; see also Lin and Chien Citation2022: fig. 12m-n). These are absent on the fossil specimens probably due to erosion (e.g. abrasion before final deposition). On the other hand, the specimens from the Seria Fm are less angular posteriorly and the size of the postostial lobe is more like that of the modern species (). These differences can partly be explained by different erosional effect, but some features may reflect time-wise changes along the lineage of the species. Nevertheless, here all these fossil Protonibea otoliths recovered from Brunei are classified under the same species, and the observed minor differences are attributed to morphological variability within the species.
Remarks
The genus Protonibea is monospecific, and today, P. diacanthus is common in the coastal areas of the Indo-West Pacific down to a depth of 100 metres (Froese and Pauly Citation2022). It must be mentioned that the NHMUK PV P 46611 specimen from the 1960’s was named as Otolithoides obesus, and later re-evaluated by Schwarzhans (Citation1993, p. 126 under Boesemania obesus), belongs to this species (see Supplementary material 2).
Protonibea sp.
Material
1 left sagitta from the Seria Fm (AH).
Dimensions
OL: 9.8 mm, OH: 6.0 mm, OL/OH: 1.63.
Description
Elongated, oval-shaped otolith with convex, rather curved margins. The inner face is convex, the external one is rather flat, but it bears a large umbo at a thickened posterior end. The ostium is large, reaches the anterior margin, and has a moderate postostial lobe. The cauda is curved by about 90 degrees and reaches the posteroventral margin.
Remarks
The sulcus shape suggests that this specimen belong to the genus Protonibea, but the otolith is thinner and has a smaller ostium with less pronounced postostial lobe than that of the above-described P. nolfi.
Genus Otolithoides Fowler, 1933
Otolithoides biauritus (Cantor, Citation1849)
1993 – Otolithoides biauritus in Schwarzhans page 207, Fig. 365.
Material
1 worn specimen from AH.
Dimensions
OL >16.2 mm; OH >9 mm; OT: 3.3 mm. Note that the exact length and height cannot be given due to the preservation state of the specimen.
Description
The specimen is worn and broken. The otolith is very thin, with a thicker dorsal side. Its inner surface is slightly convex and smooth, while the external surface is more irregular. The dorsal margin is straight in its anterior part and inclines posteriorly, the ventral margin is curved in its central zone. The posterior and anterior ends of the otolith are broken. The ostium is elongated with a large postostial lobe ending very close to the posterior end of the cauda. The anterior part of the cauda is parallel to the dorsal margin and inclines posteriorly.
Remarks
Though several features cannot be observed properly on the specimen, the preserved part of the sulcus is very similar to that of modern O. biauritus otoliths, especially the extended ostium with deep postostial lobe (Schwarzhans Citation1993: fig. 365). This also differs from the other modern species of O. pama (Hamilton, Citation1822), where the ostium has a much deeper ventral margin (Trewavas, Citation1977). Today O. biauritus is common in coastal waters from Pakistan to Borneo (Froese and Pauly Citation2022).
Genus Pterotolithus Fowler, 1933
Pterotolithus lateoides (Bleeker, Citation1849)
1993 - Pterotolithus lateoides in Schwarzhans page 132, Fig. 243.
Material
4 sagittae, 3 from Seria Fm. (AH) and 1 from TB*.
Dimensions
OL: 11.8 – ~19.6 mm, OH: 6.1–8.8 mm, OL/OH: 1.93 – ~2.22. Note, the largest specimen is broken.
Description
Elongated otoliths with a horizontal dorsal margin, and a straight or slightly bent convex ventral margin. The posterior end is angular and pointed; the anterior end is broken on the recovered specimens. The inner surface is rather smooth and slightly convex, while the external side is irregular and has a thicker posterior end that bears notches. The ostium is large, spoon-shaped, and has a clear precaudal depression. Its postostial lobe covers a large part of the ventral area. The anterior part of the cauda starts with a near horizontal portion that runs backwards, then curves down at an almost right angle. It slightly curves back towards the anterior, when it reaches the postventral margin.
Remarks
Today, the species is common around Borneo, Indonesia, and Malaysia (Froese and Pauly Citation2022). The co-occurring other modern species P. maculatus (Cuvier Citation1830) is more widely distributed in the Indo-Australian Archipelago. This species has a more thin-set otolith with less curving cauda than that of P. lateoides (see Schwarzhans Citation1993: Figs 241–242).
Family Sillaginidae Richardson, Citation1846
Genus Sillago Cuvier, Citation1816
Sillago sp.
Material
13 sagittae, 11 from the Seria Fm (AH) and 2 from the Miri Fm (JT1:1, Dad: 1).
Dimensions
OL: 5.1–7.5 mm, OH: 2.9–4.3 mm, OL/OH: 1.74–1.76.
Description
Elongated, otoliths with a wide anteroventral part and a narrow, posterior portion. The inner face is arched, and the external face is concave. The sulcus is long, horizontal, and covers the entire length of the dorsal area. The ostium is greater in height than the cauda, and is slightly anterodorsally directed. The end of the cauda is also curved dorsally.
Remarks
These observed characters are typical for the genus (see Nolf Citation2013: Plate 235 and; Lin and Chang Citation2012, p. 257). The genus Sillago represents demersal fishes with approximately 12 modern species in the wider area of the Indo-Pacific (Froese and Pauly Citation2022). From the Southeast Asian fossil record, it has only been mentioned from Taiwan (Lin et al. Citation2018, Lin et al.Citation2022; Lin and Chien Citation2022).
Order Lutjaniformes Bleeker sensu Betancur-R. et al., 2017
Family Haemulidae Gill, 1885
Genus Pomadasys Lacépède, Citation1802
Pomadasys spp.
Material
150 sagittae, 124 from the Seria Fm (AH: 118, Ker: 6), 21 from the Miri Fm. (IB: 2, JT1: 5, Dad: 13, Tag: 1), and 5 reworked specimens from TB*.
Dimensions
OL: 4.8–14.0 mm, OH: 3.9–10.2 mm, OL/OH: 1.23–1.37.
Description
The otoliths are rather oval-shaped with a large, deep ventral area. The dorsal margin is angular, the posterodorsal area is elevated, and the inclined margin is either smooth or exposes small folds. The anterior margin is rounded, while the posterior one is angular. The otoliths are rather thick with a strongly convex inner surface. The ostium is rectangular, opens anteriorly, and is greater in height and shallower than the cauda. The cauda is horizontal in the anterior portion, but its posterior end is curved ~ 60° ventrally. The cauda’s posterior tip is short and reaches only slightly below the ventral base of the ostium. Above the sulcus, especially above the cauda, there is a relatively deep dorsal depression.
Remarks
Several specimens are fragmentary, but the characteristic of the genus is easily observable. Nine modern species are reported from the wider Indo-West Pacific and Western Pacific regions (Froese and Pauly Citation2022). Today P. argenteus (Forsskål Citation1775), P. auritus (Cuvier Citation1830), P. kaakan (Cuvier Citation1830), and P. maculatus (Bloch Citation1793) are relatively common in Brunei’s coastal waters, and otoliths could be compared with our fossil fauna. Lin et al. (Citation2013) also reported comparative material of Pomadasys with some of these species plus P. quadrilineatus Shen and Lin, Citation1984. The modern otoliths show some inter-specific variations that mainly relate to the extent of the cauda’s curvature and the absence/presence of denticles on the ventral and sometimes on the posterior margins. Such denticles are always absent in our fossil otoliths, which may relate to taphonomic processes. Still, the Bruneian fossil otoliths resemble most closely to that of P. argenteus and P. kaakan. However, the prominent angular posterodorsal margin with a pointed posterior end approximately at the middle height of the otoliths on the fossil specimens cannot be observed on these modern species. The modern species have rather rounded posterior ends (Lin et al. Citation2013).
For the completeness of the record, in the NHMUK, we found the fossil species Pomadasys arcuatus (NHMUK PV P 51238 & PV P 76696) from the Seria Fm of Brunei, a future new species probably marked by Stinton in 1965 according to the labels, but it has not been published. The specimens are worn but show similar features to our Pomadasys specimens. Another specimen in the NHMUK from Miri, Sarawak (NHMUK PV P 45560) is labelled as Lutianus geminans and is also a worn Pomadasys otolith. Other fossil Pomadasys reported from the region originate from Taiwan (e.g. Lin et al. Citation2018, 2022; Lin and Chien Citation2022).
Order Priacanthiformes Bleeker, Citation1871
Family Cepolidae Rafinesque, 1810
Genus Acanthocepola Bleeker, Citation1874
Acanthocepola sp.
Material
1 right otolith from the Seria Fm. (Dad).
Dimensions
OL: 4.4 mm, OH: 2.9 mm, OL/OH: 1.52.
Description
An elongated, oval-shaped otolith with rather pointed posterior and anterior ends. All the margins are smooth. The vertical axis of the convex ventral margin is shifted a bit anteriorly. The inner face is convex, and the external is rather flat. The sulcus is typical for the family, with a wider anteriorly open ostium, situated more ventrally than the cauda, which is shorter and slightly curved.
Remarks
Four extant Acanthocepola species exist in the wider Indo-West Pacific region (Froese and Pauly Citation2022). Some of these have more ragged margins with denticles and a distinct notch at the posterodorsal margins (see species A. indica and A. limbata in http://oto.biodiv.tw/eng/). The absence of these features in the fossil specimen could be related to preservational aspects. The genus Cepola has similar otoliths, but they are more compressed and have less pronounced anteriorly shifted ventral margin (Nolf Citation2013: plates 293–294). Other Cepolidae fossil otoliths from Southeast Asia were reported from the Pleistocene of the Philippines under the genus Owstonia (Van Hinsbergh and Helwerda Citation2019).
Results and discussion
Nearly 1500 fossil fish otoliths were collected from Brunei Neogene deposits that include six sites from the Miri Formation (n = 198), four outcrops from the Seria Formation (n = 1132), and reworked specimens along a 2 km stretch of the beach near Tutong town (n = 163) (, ). The majority of the otoliths come from one single site of the Seria Fm called Ambug Hill (70.8%). It is a medium-sized quarry (<0.5 km2), which is occasionally active with a relatively large area of clay-rich sediment exposure (Kocsis et al. Citation2018). This outcrop is frequently visited for palaeontological courses and for studying molluscs (Harzhauser et al. 2018), foraminifera (Roslim et al. Citation2019), and cartilaginous fish remains (Kocsis et al. Citation2019). Due to the large differences in the recovered number of otoliths among the different sites, the faunas are discussed according to the two lithostratigraphic units (Miri and Seria formations) and the reworked site of the Tutong beach (TB*). Another important issue is the sampling approach. Most of the otoliths were collected from the surface of the sediments, and the majority of the specimens were obtained with this method (>96%). Conversely, several clay-rich samples were screen-washed over the years for foraminifera research and teaching purposes from frequently visited sites (AH, Ker, IB, JT, Dad) that yielded a few small otoliths (<2 mm) (e.g. Roslim et al. Citation2020). It is evident that the sampling is biased towards the hand-picked, large otoliths, and within the micro-remains towards those sites that were more intensively studied. Therefore, these are discussed separately.
Table 2. Fauna list of the investigated outcrops and their combined numbers for the Miri Formation, Seria Formation, and the reworked assemblage of the Tutong beach.
The remains of the families Congridae, Bregmacerotidae, Apogonidae, Gobiidae, and Cepolidae are the sole result of screen-washings, but also most of the Sillaginidae and the smaller Engraulidae (e.g. Setipinna) were found with this method. All these taxa represent only 3.7% of the total fauna (). It is important to mention that the weight of the screen-washed sediments was relatively small (0.2–0.5 kg/layer), and they yielded rarely large otoliths. Even if micro-otoliths have been obtained only from few sites by this method, the preservational state of the small otoliths is often worse than that of the large ones. The investigated sediments in Brunei were deposited in the coastal region, often with influence of estuaries and deltas (Lambiase et al. Citation2003; Lambiase and Cullen Citation2013). The sediments frequently contain the mineral pyrite and sometimes siderite cement, indicating low redox conditions in the early depositional setting (Kocsis et al. Citation2018; Roslim et al. Citation2020). Pyrite-filled voids or chambers of small calcareous shells are also observed (e.g. Roslim et al. Citation2019). When these minerals are exposed to tropical weathering and/or percolating oxygen-rich formational fluids, it would result in low pH conditions that would dramatically lower the preservation potential of the small calcareous remains including the small aragonitic otoliths as well. Conversely, the bigger fossils stayed intact, and are much better preserved.
Figure 17. Abundances graphs with biodiversity information. a: Abundance chart of the large, hand-picked otoliths from the three investigated origins (Seria Formation, Miri Formation, and reworked fauna of Tutong beach – TB*). Pie-chart on the combined fauna show the relative percentages of the recovered fish families. b: Abundance chart of the screen-washed micro-remains (Seria Fm, Miri Fm). Pie-chart on the combined micro-fauna showing relative percentages of the recovered fish families. Note the colour code that corresponds to three faunas on graph (red – Seria Fm, green – Miri Fm, black – reworked fauna TB*). Note that the family Engraulidae appears in both graphs, and the single, common species of Coilia planata represents them among the large otoliths.
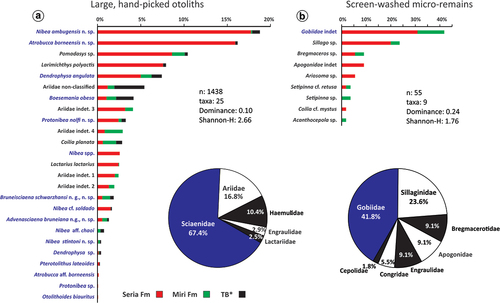
Nevertheless, the screen-washing revealed the presence of many families, even if some are only represented by a few remains. For example, only one Cepolidae otolith was found in the Miri Fm, while the families of Congridae and Apogonidae have so far only been discovered from the Seria Fm. The overall relative abundances of the families are shown in . The Seria Fm sites yielded more micro remains than the Miri Fm (74.5% vs 25.5%), and in both, gobiid otoliths are most prevalent. Today, all these families are common in the Southeast Asian seas, and some with extreme diversity (Apogonidae, Gobiidae, see also ). Therefore, future studies, especially targeting new and freshly opened outcrops, could give further insight about these groups and probably uncover new taxa. It is important to emphasise that future studies must focus on larger bulk sampling with standardised weight. Such an approach could provide a better picture about the fauna’s composition and its diversity.
Table 3. Environment and habitat conditions of the recorded taxa based on modern representatives. Most of the data is derived from Nelson et al. (Citation2016) and Froese and Pauly (Citation2022)
Regarding the large, hand-picked otoliths, in all the three assemblages (Miri, Seria, reworked), the family Sciaenidae (drums and croakers) is the most abundant and diverse group (, ), but in general, sciaenid otoliths also tend to be among the largest. Altogether 17 different sciaenid taxa are reported here, among them are otoliths of two new genera (Advenasciaena, Bruneisciaena). Advenasciaena is especially peculiar with its thick-set otoliths and unique features such as a deep, thin anterior groove and a very shallow, posteriorly extended triangular ostial colliculum (). In addition, one new Atrobucca, two Nibea, and a Protonibea species are presented here. N. stintoni is a rare new species and only a few remains have been discovered so far. Conversely, A. borneensis and Nibea ambugensis are extremely abundant in the Seria Fm. The sciaenids clearly dominate the entire Seria Fm fauna (), which is mainly due to three very common species: Nibea ambugensis (18%), Larimichthys polyactis (8%), and the already mentioned A. borneensis (16%). The latter two were only found at Ambug Hill, and they are also known from a few reworked specimens at Tutong beach. This may suggest Seria Fm origin for the reworked otolith fauna, which is not surprising based on the proximity to the in-situ beds of the Seria Fm (). Furthermore, Sr-isotope ages and taxonomic elements of reworked elasmobranch fauna (e.g. Hemipristis, Otodus megalodon – known so far only from the Seria Fm - Kocsis et al. Citation2019) also support that most of the reworked otoliths are probably derived from the Seria Fm (Kocsis et al. Citation2022).
Some other sciaenids are represented only by a few specimens (e.g. Pterotolithus, Otolithoides), but taxa from the Nibea-group (see groupings in Schwarzhans Citation1993) can be very common (Dendrophysa, Nibea) within all three assemblages. The fossil species Boesemania obesus, described by Stinton (Citation1962), is also recorded from many sites. This taxon is interesting in the sense that it is monospecific today and represented by the species B. microlepis, which is known to inhabit freshwater and brackish environments. Many of the sciaenids occurring in our record live in shallow marine, coastal environments, often in turbid waters, but some also inhabit and/or migrate to brackish environments (e.g. some Nibea, Dendrophysa – Sasaki Citation1998; Froese and Pauly Citation2022, see ).
Other commonly occurring fishes in Brunei’s sediments are the Ariidae (sea catfishes), Haemulidae (grunts – genus Pomadasys), Engraulidae (anchovies – genus Coilia, Setipinna), and Lactariidae (false trevally – genus Lactarius). Their relative proportions within the three assemblages vary somewhat (), but they can be considered an important and integrated part of all otolith assemblages, except the Lactariidae that occurs dominantly in the Seria Fm. The lapilli from the family Ariidae are the only utricular otoliths that are enlarged and found in the Borneo assemblages, and are easily recognised by their shape and lack of typical sagittal features. However, their taxonomy is challenging due to low level variability among the different genera/species. We distinguished four groups here, but it is anticipated that future studies may reveal more detailed information about ariids’ past biodiversity in Borneo. This group is species-rich today in the coastal region of Borneo with fully marine and brackish species (Kailola Citation1998; Froese and Pauly Citation2022). Some taxa visit estuaries either daily with the tide looking for food or seasonally for reproductive reasons (). Based on the data collected, the other families are much less diverse in our record. The Haemulidae are only represented by the genus Pomadasys, which is known to frequent low salinity environments (McKay Citation1998; Froese and Pauly Citation2022) and occurs in the turbid waters of the Sungai Brunei’s estuaries as well. The Engraulidae are dominated by a single taxon, the fossil species Coilia planata. Some modern Coilia species also visit brackish and even freshwater environments (Wongratana et al. Citation1998; Froese and Pauly Citation2022, ), and therefore, similar habitats could probably be expected for the fossil species. There is only one Lactariidae species today, Lactarius lactarius, which was described from our localities as well, representing the first fossil record of the taxon (Nolf Citation2013). By contrast, the otolith fossil records of Lactarius are widely distributed globally since the Paleogene, typically in the shallow marine settings (Stinton Citation1978; Steurbaut and Jonet Citation1982, Nolf and Bajpai Citation1992; Lin and Nolf Citation2022). These fishes are common in shallow marine neritic-pelagic environments but were also observed in brackish milieus (Leis Citation1998; Froese and Pauly Citation2022).
The Seria, Miri, and reworked otolith assemblages appear to indicate a palaeoenvironment of a shallow marine coastal setting, which concurs with sedimentological observation (e.g. Lambiase and Cullen Citation2013; Kocsis et al. Citation2018). Close connection to estuaries is also assumed where many of the recovered fishes might have migrated either for reproduction or for a food source with the tide. Turbid, suspension-loaded water might also be presumed as ariids, sciaenids, and the haemulid Pomadasys are often observed and caught in these conditions today. The overall fauna is dominated by demersal taxa (e.g. Ariidae, Pomadasys, Gobiidae, Sillaginidae, some of the Sciaenidae), while some may indicate rather soft substrate in the area as they generally live in burrows in the sediment (Acanthocepola, Cepolidae – band fish; Arisoma, Congridae – congrid eel, ). The difference between the Seria and the Miri faunas, based on the large-sized otoliths, is best seen by the occurrence of certain sciaenids (Nibea ambugensis, Atrobucca borneensis, Larimichthys polyactis) and Lactarius lactarius in the younger deposit, but they are completely absent or very rare in the beds of the Miri Fm (, ). Modern Atrobucca species (e.g. A. nibe, A. brevis) and the Larimichthys polyactis are marine, benthopelagic fishes, while some extant Nibea species can also inhabit fully marine milieu (e.g. N. chui). The dominant presence of these taxa in the beds of the Seria Fm (AH) could indicate somewhat different environmental conditions with more access to open marine waters in the area. Another interesting fact is that, today Larimichthys polyactis live in temperate water, which means that this species likely had wider distribution in the Miocene and once inhabited tropical coastal regions. Nevertheless, to further trace and assess the habitat and environmental conditions in which these fishes lived in the past, trace element and stable isotope measurements of the otolith could reveal more details in the future.
Regarding the diversity indices (Shannon-H, Dominance), the large hand-picked otoliths reveal a diverse palaeocommunity (). Conversely, the screen-washed derived micro-material show less diversity. The latter is evidently an underestimation due to the lack of large, bulk sediment sampling. In addition, even if our micro-remains are low in number, the gobiid and apogonid each represent probably two-to-three different taxa. Therefore, a more detailed taxonomic study, together with future large-scale sampling could shed more light on the fish biodiversity of the region.
As a comparison to the previous study of Stinton (Citation1962) from Borneo, he reported only 62 otoliths that were sorted into 18 taxa of 10 families. Based on Nolf’s update (2013), all the reported valid nominal species from Brunei have been recovered by our research. However, from the Miri area of Sarawak and the Togopi Fm from the Dent Peninsula in Sabah, Stinton described a few specimens from the families of Carapidae (Carapus extensus (Stinton Citation1962), Holocentridae (Myripristis trigonus), and Lutjanidae (Lutjanus geminans) (Stinton Citation1962: figs 19. 9,11,18 see also Supplementary material 2), that were not yet found in Bruneian deposits. Based on the obtained rarefaction curves, new taxa might well be expected in Brunei, especially from the micro-remains that yielded rather unsaturated curves (). These could certainly add to the overall palaeobiodiversity of the region. Conversely, the rarefaction curves of the large otoliths tend to level out (), and therefore the big picture of the dominant presence of sciaenid and ariids in the large-sized otoliths probably will not change. Nevertheless, detailed studies on the ariids could result in a better assessment on the biodiversity of this group in the future.
Figure 18. Rarefaction curves. Note that the curves of the large otoliths level out, while the micro-remains one is undersaturated. The latter indicates that with collecting more otoliths, additional taxa can be expected.
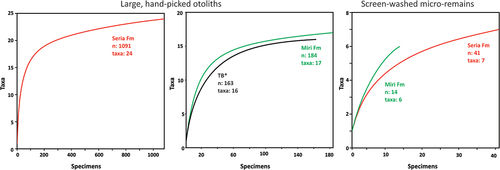
Conclusion
Our study reports diverse coastal marine fish faunas based on otoliths from the Neogene beds of Brunei Darussalam in Borneo. The recovered 1493 otoliths belong to 34 taxa of 11 families, and among them six new species and two new genera (Advenasciaena bruneiana gen. nov. sp. nov., Atrobucca borneensis sp. nov., Bruneisciaena schwarzhansi gen. nov. sp. nov., Nibea ambugensis sp. nov., Nibea stintoni sp. nov., and Protonibea nolfi sp. nov.) are reported. The overall fauna is dominated by Sciaenidae, followed by Ariidae, Haemulidae, Engraulidae, and Lactariidae. Most of these specimens were collected from the surface of the outcrops, however, other families were revealed by occasional screen-washing (Gobiidae, Sillaginidae, Bregmacerotidae, Apogonidae, Congridae, and Cepolidae). Many of these groups are represented only by a few specimens and collectively the micro-remains compose 3.7% of the entire fauna. Future bulk sampling of the sediments could shed further light on the abundance and diversity of the micro-otolith assemblages. Based on the large-sized otoliths, the fish fauna indicates a palaeoenvironment with a coastal marine setting with nearby estuaries and deltas, probably with turbid water and, from time to time with lower salinity conditions. The younger deposits of the Seria Fm (AH) may reflect more open marine connections but still with a shallow setting with close proximities of river inputs. When comparing our findings with modern biodiversity and previous studies of the fossil record from the region, it is evident that our results increase our understanding of fish palaeobiodiversity in the Indo-Australian Archipelago, where today’s highest marine biodiversity exists. The described faunas and new taxa further highlight that the region’s fossil record is understudied, and new palaeontological findings are expected in the future.
Supplemental Material
Download PDF (9.2 MB)Supplemental Material
Download PDF (100.9 KB)Acknowledgments
The research was supported by two university research grants of Universiti Brunei Darussalam: UBD/PNC2/2/RG/1(325) and UBD/RSCH/1.4/FICBF(b)/2019/023. Antonino Briguglio’s suggestions for the naming of the new taxa and their proper use, and Amajida Roslim help with some of the illustrations and specimen donations are greatly appreciated. We thank to the Department of Museums, Brunei and the Institute for Biodiversity and Environmental Research (IBER) at UBD for their aid in the donation process of key specimens to other, internationally recognised collections. Travel funding to Brunei was provided by the Ministry of Science and Technology, Taiwan (Grant No. 108-2116-M-029-001-MY2, 109-2116-M-001-022-) to Chien-Hsiang Lin. The manuscript has benefited from the constructive comments of Werner W. Schwarzhans, Gary Stringer, and Dirk Nolf. We are grateful for their thorough and detailed inputs.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2023.2271489
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Aguilera OA, Schwarzhans W, Béarex P. 2016. Otoliths of the Sciaenidae from the Neogene of tropical America. PalaeoIchthyologica. 14:7–90.
- Back S, Tioe HJ, Thang TX, Morley CK. 2005. Stratigraphic development of synkinematic deposits in a large growth-fault system, onshore Brunei Darussalam. J Geol Soc. 162(2):243–257. doi: 10.1144/0016-764903-006.
- Beaufort de LF. 1926. On a collection of marine fishes from the Miocene of South Celebes. Jaarboek van het Mijnwezen in Nederlandsch Oost–Indië. 1925:117–148.
- Beaufort de LF 1931. Pisces, Reptilia and Aves. Leidsche Geologische Mededelingen, 5: 461–470.
- Betancur-R R, Wiley EO, Arratia G, Acero A, Bailly N, Miya M, Lecointre G, Orti G. 2017. Phylogenetic classification of bony fishes. BMC Evol Biol. 17(162):1–40. doi: 10.1186/s12862-017-0958-3.
- Bleeker P. 1846. Siluroideorum bataviensium conspectus diagnosticus. [Inside title: Overzigt der Siluroieden, welke te Batavia voorkomen]. Verhandelingen van het Bataviaasch Genootschap van Kunsten en Wetenschappen. 21:1–60.
- Bleeker P. 1849. Bijdrage tot de kennis der Sciaenoïden van den Soenda-Molukschen Archipel, met beschrijving van 7 nieuwe soorten. Verhandelingen van het Bataviaasch Genootschap van Kunsten en Wetenschappen. 23(5):1–31.
- Bloch ME. 1793. Naturgeschichte der ausländischen Fische. Berlin. 7:144.
- Brouwer HA, Beaufort de LF. 1923. On tertiary marine deposits with fossil fishes from south celebes. Proceedings, Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, 26: 159–166.
- Cantor TE. 1849. Catalogue of malayan fishes. J Asiatic Soc Bengal. 18:983–1443.
- Carolin N, Bajpai S, Maurya SA, Schwarzhans W. 2023. New perspectives on late Tethyan Neogene biodiversity development of fishes based on Miocene (~ 17 ma) otoliths from southwestern India. PalZ. 97(1):43–80. doi: 10.1007/s12542-022-00623-9.
- Cowman PF. 2014. Historical factors that have shaped the evolution of tropical reef fishes: a review of phylogenies, biogeography, and remaining questions. Front Genet. 5(394):1–15. doi: 10.3389/fgene.2014.00394.
- Cuvier G. 1829. Le Règne Animal, distribué d'après son organisation, pour servir de base à l'histoire naturelle des animaux et d'introduction à l'anatomie comparée. Edition. 2:406.
- Cuvier G. 1830. Histoire naturelle des poissons. Tome cinquième. Livre cinquième. Des Sciénoïdes. F. G. Levrault, Paris. p. 499.
- Forsskål P. 1775. Descriptiones animalium avium, amphibiorum, piscium, insectorum, vermium; quae in itinere orientali observavit Petrus Forskål. Post mortem auctoris edidit Carsten Niebuhr, Hauniae. p. 164.
- Frédérich B, Sorenson L, Santini F, Slater GJ, Alfaro ME. 2013. Iterative ecological radiation and convergence during the evolutionary history of damselfishes (pomacentridae). Am Nat. 181(1):94–113. doi: 10.1086/668599.
- Froese R, Pauly D (Eds) 2022. FishBase. World Wide Web electronic publication. www.fishbase.org, version (02/2022).
- Frost GA. 1925. Description of fish otoliths from the tertiary formations of Atcheen, northern Sumatra. Wetenschappelijke mededeelingen Dienst van den Mijnbouw in Nederlandsch-Oost Indie . 2:1–28.
- Hamilton F. 1822. An account of the fishes found in the river Ganges and its branches. Edinburgh & London. p. 405.
- Hennig E. 1911. Die Fischreste. In: Selenka L Blanckenhorn M, editors. Die Pithecanthropus-Schichten auf Java. Geologische und palaontologische Ergebnisse der Trinil-Expedition (1907 und 1908). Leipzig: Verlag Von Wilhelm Engelmann; pp. 54–60.
- Hoeksema BW. 2007. Delineation of the Indo-Malayan centre of maximum marine biodiversity: the coral triangle. In: Renema W, editor. Biogeography, Time and place: distributions, barriers, and islands. Dordrecht, The Netherlands: Springer; pp. 117–178.
- James DMD. 1984. The geology and Hydrocarbon Resources of Negara Brunei Darussalam. Brunei Darussalam: Bandar Seri Begawan, Muzium Brunei. p. 165.
- Jordan DS, Thompson WF. 1911. A review of the sciaenoid fishes of Japan. Proceedings of the United States National Museum. 39: 241–261.
- Kailola PJ. 1998. Order siluriformes. Ariidae (=Tachysuridae). Sea catfishes (fork-tailed catfishes). In: Carpenter K, and Niem V, editors. FAO species identification guide for fishery purposes. The living marine resources of the western central pacific. Volume 3 batoid fishes, chimaeras and bony fishes part 1 (elopidae to linophrynidae). Vol. 2001. Rome: FAO; pp. 1827–1879.
- Kailola PJ. 2000. Six new species of fork-tailed catfishes (pisces, Teleostei, ariidae) from Australia and new Guinea. The beagle, records of the museums and art galleries of the northern territory. 16:127–144.
- Kner R, Steindachner F. 1867. Neue Fische aus dem Museum derNeue Fische aus dem Museum der. Mathematisch-Naturwissenschaftliche Classe. 54:356–395.
- Kocsis L, Briguglio A, Roslim A, Razak H, Corić S, Frijia G. 2018. Stratigraphy and age estimate of Neogene shallow marine fossiliferous deposits in Brunei Darussalam (Ambug Hill, Tutong district). J Asian Earth Sci. 158:200–209. doi: 10.1016/j.jseaes.2018.02.023.
- Kocsis L, Razak H, Briguglio A, Szabó M. 2019. First report on a diverse Neogene fossil cartilaginous fish fauna from Borneo (Ambug Hill, Brunei Darussalam). J Syst Paleontol. 17(10):791–819. doi: 10.1080/14772019.2018.1468830.
- Kocsis L, Botfalvai G, Qamarina Q, Razak H, Király E, Lugli F, Wings F, Lambertz M, Raven H, Briguglio A, et al. 2020. Geochemical analyses suggest stratigraphic origin and late Miocene age of reworked vertebrate remains from Penanjong beach in Brunei Darussalam (Borneo). Hist Biol. 33(11):2627–2638. doi: 10.1080/08912963.2020.1819999.
- Kocsis L, Briguglio A, Cipriani A, Frijia G, Vennemann T, Baumgartner C, Roslim A. 2022. Strontium isotope stratigraphy of late Cenozoic fossiliferous marine deposits in North Borneo (Brunei, and Sarawak, Malaysia). J Asian Earth Sci. 231:105213. doi: 10.1016/j.jseaes.2022.105213.
- Koumans FP. 1949. On some fossil fish remains from Java. Zoologische mededeelingen. 30(5):77–82.
- Lacepède BGE. 1802. Histoire naturelle des poissons. 4:1–728.
- Lambiase JJ, Cullen BA. 2013. Sediment supply systems of the champion “delta” of NW Borneo: implications for deepwater reservoir sandstones. J Asian Earth Sci. 76:356–371. doi: 10.1016/j.jseaes.2012.12.004.
- Lambiase JJ, Razak Damit A, Simmons MD, Abdoerrias R, Hussin A. 2003. A depositional model and the stratigraphic development of modern and ancient tide-dominated deltas in NW Borneo. In: Posamentier H, Hasan Sidi F, Darman H, and Nummedal D, editors. Deltas of the Asia Pacific Region: modern and ancient, Claremore, OK, U.S.A: Society of Sedimentary Geology Special Publication; Vol. 76, pp. 109–124.
- Leis JM. 1998. Lactariidae. False trevally (milk trevally). In: Carpenter K, and Niem V, editors. FAO species identification guide for fishery purposes. The living marine resources of the western central pacific. Volume 4 Bony fishes part 2 (mugilidae to carangidae). Vol. 2001. Rome: FAO; p. 2649.
- Leriche M. 1954. Les faunes ichthyologiques marines du Néogène des Indes Orientales. Mémoires de la Société Paléontologique Suisse. 70:1–21.
- Liechti P, Roe FN, Haile NS, Kirk HJC. 1960. The geology of Sarawak, Brunei and the western part of North Borneo. Bulletin 3, volumes 1&2. Geological survey department British territories in Borneo. Kuching, Sarawak: Government Printing Office. p. 360.
- Lin CH, Chang CW. 2012. Otolith atlas of Taiwan fishes. Pingtung: National Museum of Marine Biology and Aquarium. p. 415.
- Lin CH, Chien CW. 2022. Late Miocene otoliths from northern Taiwan: insights into the rarely known Neogene coastal fish community of the subtropical northwest Pacific. Hist Biol. 34(2):361–382. doi: 10.1080/08912963.2021.1916012.
- Lin CH, Chien CW, Lee SW, Chang CW. 2021. Fish fossils of Taiwan: a review and prospection. Hist Biol. 33(9):1362–1372. doi: 10.1080/08912963.2019.1698563.
- Lin CH, Li KT, Chang CW. 2013. Identification of Pomadasys species (pisces, Haemulidae) from an archaeological midden site in Nankuanli east (Taiwan), based on otolith morphology. Raff Bullet Zool. 61(1):293–302.
- Lin CH, Nolf D. 2022. Middle and late Eocene fish otoliths from the eastern and southern USA. Eur J Taxon. 814:1–122. doi: 10.5852/ejt.2022.814.1745.
- Lin CH, Wang LC, Wang CH, Chang CW. 2018. Common early Pleistocene fish otoliths from Niubu in Chia-Yi County, southwestern Taiwan. J National Taiwan Mus. 71:47–68. Chinese.
- Lin CH, Wang YC, Ribas-Deulofeu L, Chang CW, Li KT. 2022. Changes in marine resource consumption over the past 5000 years in southwestern Taiwan revealed by fish otoliths. J Archaeol Sci Rep. 42:103400. doi: 10.1016/j.jasrep.2022.103400.
- Lo P, Liu S, Mohd nor SA, Chen W. 2017. Molecular exploration of hidden diversity in the Indo-west Pacific sciaenid clade. PloS One. 12(4):e0176623. doi: 10.1371/journal.pone.0176623.
- Martin K. 1883. Palaeontologische Ergebnisse von Tiefbohrungon auf Java, nebst allgemeineren Studien über das Tertiar von Java, Timor und einiger anderer Inseln. Samml Geol Reichs-Museum Leiden ser I Bd. III:380.
- McKay EJ. 1998. Haemulidae (=Pomadasyidae). Grunts (also sweetlips, rubberlips, hotlips, and velvetchins). In: Carpenter K, and Niem V, editors. FAO species identification guide for fishery purposes. The living marine resources of the western central pacific. Volume 5. Bony fishes part 3 (menidae to pomacentridae). Vol. 2001. Rome: FAO; pp. 2961–2989.
- Munro ISR. 1964. Additions to the fish fauna of New Guinea. Papua New Guinea Agricultural Journal. 16(4):141–186.
- Nelson SJ, Grande CT, Wilson HVM. 2016. Fishes of the world. 5th ed. Hoboken, New Jersey: Wiley & Sons; p. 707.
- Nolf D. 1985. Otolithi piscium. In: Schultze H, editor. Handbook of paleoichthyology. Vol. 10, Stuttgart: Gustav Fischer Verlag; p. 146.
- Nolf D. 2013. The diversity of fish otoliths, past and present. Brussels: Royal Belgian Institute of Natural Sciences. p. 581.
- Nolf D, Bajapi S. 1992. Marine middle Eocene fish otoliths from India and Java. Bulletin de l’Institut royal des Sciences naturelles de belgique. Science de la Terre. 62:175–221.
- Ohe F. 2006. Skulls and otoliths of eleven sea catfishes (family Ariidae) from Malaysia and one species related to them from the east China sea. Natural Environ Sci Res. 19:11–28.
- Ohe F. 2011. Otolith figures of sciaenid fish inhibiting the waters around Japan, with the comments for them from the geohistorical background. Kaseki No Tomo. 56:32–39.
- Posthumus O. 1929. Vischotolieten van N.O, Borneo. Wetenschappelijke mededeelingen Dienst van den Mijnbouw in Nederlandsch-Oost Indie. 9:87–108.
- Razak H, Kocsis L. 2018. Late Miocene otodus (megaselachus) megalodon from Brunei Darussalam: body length estimation and habitat reconstruction. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen Stuttgart. 288(3):299–306. doi: 10.1127/njgpa/2018/0743.
- Renema W, Bellwood DR, Braga JC, Bromfield K, Hall R, Johnson KG, Lunt P, Meyer CP, McMonagle LB, Morley RJ, et al. 2008. Hopping hotspots: global shifts in marine biodiversity. Sci. 321(5889):654–657. doi: 10.1126/science.1155674.
- Richardson J. 1846. Report on the ichthyology of the seas of China and Japan. Report of the British Association for the Advancement of Science 15th Meeting. 187–320.
- Roslim A, Briguglio A, Kocsis L, Corić S, Gebhardt H. 2019. Large rotaliid foraminifera as biostratigraphic and palaeoenvironmental indicators in northwest Borneo: an example from a late Miocene section in Brunei Darussalam. J Asian Earth Sci. 170:20–28. doi: 10.1016/j.jseaes.2018.10.019.
- Roslim A, Briguglio A, Kocsis L, Abd. Rahman F, Bahrein IF, Goeting S, Razak H. 2020. Sedimentology and stratigraphy of Late Miocene outcrops (Miri and Seria formations) along Jalan Tutong in Brunei Darussalam. Bull Geolog Soc Malaysia. 70(1):39–56. doi: 10.7186/bgsm70202004.
- Rüppell WPES. 1837. Neue Wirbelthiere zu der Fauna von Abyssinien gehörig. Fische des Rothen Meeres. Siegmund Schmerber, Frankfurt am Main. 53–80.
- Sandal ST. 1996. The Geology and Hydrocarbon Resources of Negara Brunei Darussalam (1996 revision). Brunei Shell Petroleum Co. and Muzium Negara, Syabas Bandar Seri Begawan, Brunei Darussalam, p. 243.
- Sasaki K. 1998. Sciaenidae. Croakers (drums). In: KE C, and Niem V, editors. FAO species identification guide for fishery purposes. The living marine resources of the western central pacific. Volume 5. Bony fishes part 3 (menidae to pomacentridae). Vol. 2001. Rome: FAO; pp. 3117–3174.
- Schwarzhans W. 1993. A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). Piscium catalogus: part otolithi piscium. Vol. 1. München: Verlag Dr. Friedrich Pfeil; p. 245.
- Schwarzhans W. 2019. A comparative morphological study of recent otoliths of the congridae, muraenesocidae, nettastomatidae and colocongridae (Anguilliformes). Memorie della Società Italiana di Scienze Naturali e del Museo di Storia Naturale di Milano. 46:327–354.
- Seah YG, Hanafi N, Ghaffar Mazlan A, Chao NL. 2015. A new species of Larimichthys from Terengganu, east coast of Peninsular Malaysia (Perciformes: Sciaenidae). Zootaxa. 3956(2):271–280. doi: 10.11646/zootaxa.3956.2.7.
- Shen S-C, Lin W-W. 1984. Some new records of fishes from Taiwan with descriptions of three new species. Taiwan Museum Special Publications Series. 4:1–25.
- Sorenson L, Santini F, Alfaro ME. 2014. The effect of habitat on modern shark diversification. J Evol Biol. 27(8):1536–1548. doi: 10.1111/jeb.12405.
- Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson M, Halpern BS, Jorge MA, Lombana A, Lourie SA, et al. 2007. Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. Bioscience. 57(7):573–583. doi: 10.1641/B570707.
- Steurbaut E, Jonet S. 1982. Revision des otolithes de téléostéens du Miocène Portugais. Bulletin de la Société Belge de Géologie. 90(3):191–229.
- Stinton FC. 1962. Teleostean otoliths from the upper tertiary strata of Sarawak, Brunei and North Borneo. British Borneo Geological Survey, Annual Report. 1962:75–92.
- Stinton FC. 1978. Fish otoliths from the English Eocene (part 3). Palaeontographical Society Monographs. 555(555):127–189. doi: 10.1080/25761900.2022.12131741.
- Thunberg CP. 1792. Tvånne Japanske fiskar. Kongliga Vetenskaps-Academiens Handlingar. Stockholm. 13:29–32.
- Trewavas E. 1971. The syntypes of the sciaenid corvina albida Cuvier and the status of Dendrophysa hooghliensis Sinha and Rao and Nibea coiborThe syntypes of the sciaenid corvina albida Cuvier and the status of Dendrophysa hooghliensis Sinha and Rao and Nibea coibor. J Fish Biol. 3(4):453–461.
- Trewavas E. 1977. The sciaenid fishes (croakers or drums) of the lndo-West-Pacific. Trans Zoolog Soc London. 33:253–541.
- Valenciennes A. 1840. Histoire naturelle des poissons. Tome quatorzième. Suite du livre seizième. Labroïdes. Livre dix-septième. Des Malacoptérygiens. Pitois-Levrault, Paris in Cuvier & Valenciennes. 464.
- Van Hinsbergh VWM, Helwerda RA. 2019. Fish otoliths from the Cabarruyan Piacenzian-Gelasian fauna found in the Philippines. Zootaxa. 4563(3):401–443. doi: 10.11646/zootaxa.4563.3.1.
- Vorstman AG. 1927. Tertiaire vischotolieten van Java. Wetenschappelijke mededeelingen Dienst van den Mijnbouw in Nederlandsch-OostIndie. 5:1–24.
- Wannier M, Lesslar P, Lee C, Raven H, Sorkhabi R, Ibrahim A. 2011. Geological excursions around Miri, Sarawak. Miri, Sarawak, Malaysia: Ecomedia. p. 279.
- Wilford GE. 1961. The geology and mineral resources of Brunei and adjacent parts of Sarawak with description of Seria and Miri Oilfields. In: Memoir geological survey department. Vol. 10. Brunei: British Territories in Borneo; p. 319.
- Wilf P, Zou X, Donovan MP, Kocsis L, Briguglio A, Shaw D, Slik F, Lambiase JJ. 2022. First fossil-leaf floras from Brunei Darussalam show dipterocarp dominance in Borneo by the pliocene. PeerJ. 10:e12949. doi: 10.7717/peerj.12949.
- Wong JY, Chu C, Chong VC, Dhillon SK, Loh KH. 2016. Automated otolith image classification with multiple views: an evaluation on Sciaenidae. J Fish Biol. 89(2):1324–1344. doi: 10.1111/jfb.13039.
- Wongratana T, Munroe TA, Nizinski MS. 1998. Order Clupeiformes. Engraulidae. Anchovies. In: Carpenter K, and Niem V, editors. FAO species identification guide for fishery purposes. The living marine resources of the western central pacific. Volume 3 batoid fishes, chimaeras and bony fishes part 1 (elopidae to linophrynidae). Vol. 2001. Rome: FAO; pp. 1698–1753.