
ABSTRACT
Megalolamna is an extinct lamniform shark genus that is known from Miocene marine deposits. In this study, three new specimens of Megalolamna collected from the eastern USA are newly described, and the diagnosis of the genus is emended to include a previously undocumented dental characteristic. The new specimens are represented by one tooth from the upper Oligocene Chandler Bridge Formation in South Carolina and two isolated teeth from the lower Miocene Calvert Formation in Maryland. The specimens from the Calvert Formation represent the northern-most occurrence of Megalolamna. Although the taxonomic identification of the specimen from the Chandler Bridge Formation is somewhat tentative, it is considered to represent geologically the oldest record of Megalolamna, suggesting the likelihood that the enigmatic genus has its origin in the Palaeogene no later than 23.5 Ma.
Introduction
Megalolamna is an extinct lamniform shark genus that is known from Miocene marine deposits and is represented by a single species, M. paradoxodon (Carrillo-Briceño et al. Citation2016, Citation2020; Shimada et al. Citation2017). Although its dental morphology suggests that the taxon belongs to the family Otodontidae, which includes genera such as Cretalamna and Otodus, the species name (paradoxodon) denotes its paradoxical occurrence marked by geologically confined (Aquitanian‒Burdigalian) and sparse but geographically wide distributions with an uncertain phylogenetic origin within the otodontid clade (Shimada et al. Citation2017). Megalolamna paradoxodon has so far been reported from along the Pacific Rim and the western Atlantic Ocean (Carrillo-Briceño et al. Citation2016, Citation2020; Shimada et al. Citation2017), where the earliest stratigraphic record of Megalolamna has been at the Oligocene-Miocene boundary (Carrillo-Briceño et al. Citation2020).
In this study, we report three new specimens of Megalolamna collected from the eastern USA. One comes from the upper Oligocene Chandler Bridge Formation in South Carolina, whereas two specimens come from the lower Miocene Calvert Formation in Maryland (). They are collectively significant because they extend the stratigraphic and geographic ranges of the genus. In addition, they offer new information about the dental morphology of the genus. Therefore, the purpose of this paper is three-fold: (1) to describe and document the new specimens; (2) to emend the generic diagnosis based on a newly recognised dental characteristic; and (3) to highlight their geographic and stratigraphic significance.
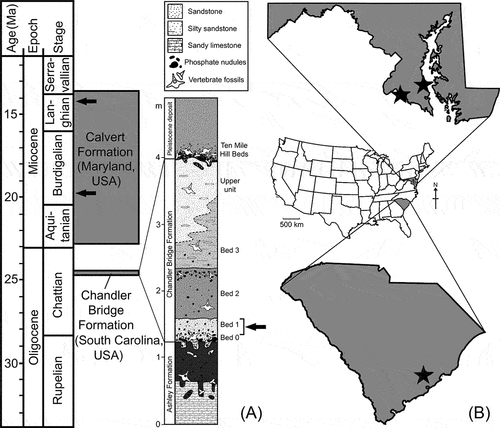
Figure 1. Stratigraphic and geographic positions of specimens of Megalolamna described in this paper. (A) Oligocene to early–middle Miocene geochronology showing stratigraphic position of Calvert Formation of Maryland, USA, and Chandler Bridge Formation of South Carolina, USA, as well as stratigraphic horizons of the specimens (arrow marks; generalized geologic column showing Chandler Bridge Formation is included because the occurrence of Megalolamna marks the oldest geologic record for the genus) (stratigraphic information based on Weems et al. (Citation2016, figure 8), Vogt et al. (Citation2018, figure 1.3), and Fallon and Boessenecker (Citation2020, figure 1C)). (B) map of continental USA (middle) showing Maryland (top) and South Carolina (bottom) along with localities of Megalolamna specimens described in this paper (stars).
Geologic setting
Calvert Formation, Maryland
The early – middle Miocene Calvert Formation () is widely exposed along the western shore of the Chesapeake Bay and along the banks of adjacent tributaries in southern Maryland and Virginia (Harris Citation1893; Shattuck Citation1904; Gernant et al. Citation1971; Gibson Citation1983; Kidwell Citation1984, Citation1989, Citation1997; Ward Citation1992; Ward and Andrews Citation2008; Kidwell et al. Citation2015; Powars et al. Citation2015; Vogt et al. Citation2018; Perez et al. Citation2019). The Calvert Formation is overlain by the middle to late Miocene Choptank and St. Marys formations and underlain by the Eocene Nanjemoy Formation. The three Miocene formations (Calvert, Choptank, and St. Marys) were organised into 24 intraformational units termed ‘zones’ by Shattuck (Citation1904), which were later revised and formalised as ‘beds’ by Ward and Andrews (Citation2008). The Calvert Formation is divided into the Fairhaven Member (Beds 1–3B) and the Plum Point Member (Beds 4–16A). The lithology of the Calvert Formation consists of poorly consolidated siliciclastic sediments and shell beds rich in macroinvertebrate and vertebrate fossils. Chondrichthyan fossils are well documented from the Calvert Formation (e.g. Kent Citation1994, Citation2018), with the peak abundance occurring during the Middle Miocene Climatic Optimum within Beds 10, 12, and 14 (Visaggi and Godfrey Citation2010).
There are two teeth attributed to Megalolamna from the Calvert Formation of Maryland () described in this paper. The first tooth (CMM-V-10306) was found at a locality known as Popes Creek on the Maryland side of the Potomac River. In this locality, the Fairhaven Member of the Calvert Formation lies disconformably above the Eocene Nanjemoy Formation and below an unnamed Pleistocene unit (Ward and Andrews Citation2008, figure 16). CMM-V-10306 was found in situ, within the Popes Creek Sand bed, which is a stratigraphic unit situated between Beds 3A and 3B (Perez et al. Citation2019, figure 1). The second tooth (CMM-V-10270) was found as beach float at Flag Ponds Nature Park along the western shore of the Chesapeake Bay. Flag Ponds is situated in a low elevation gap between the cliffs, but the adjacent cliffs from which the tooth likely originated comprise Beds 16A–21 (Kidwell et al. Citation2015). Although we cannot be certain which bed the tooth came from, the preservation is strongly suggestive of the Calvert Formation rather than the Choptank or St. Marys formations. Bed 16A was deposited during the Middle Miocene Climatic Transition, as the global climate began to cool and eustatic sea level was dropping in association with the expansion of the east Antarctic ice sheet (Vogt and Parrish Citation2012; Pierce et al. Citation2017).
Chandler Bridge Formation, South Carolina
The Chandler Bridge Formation () ranges from 0.3 to 2.5 m and unconformably overlies the Ashley Formation and underlies a series of Pleistocene deposits, where the former is thought to have formed due to Neogene erosion (Katuna et al. Citation1997). The formation is non-calcareous, consisting of massively bedded, fossiliferous phosphatic siltstone and fine sandstone, and it is divided into four subunits, Beds 0–3 (Sanders et al. Citation1982; Katuna et al. Citation1997; Fallon and Boessenecker Citation2020; (). Bed 0 (sensu Boessenecker and Geisler Citation2018) consists of 10‒15 cm, discontinuous, unconsolidated olive-brown sand, silt, and clay, richly containing phosphatic nodules and vertebrate remains. Bed 1 consists of moderately well-sorted, unconsolidated light yellowish-brown siltstone containing very fine-grained quartz sand particles, and it is likewise rich in vertebrate fossils. Bed 2 is a poorly sorted, light tan to brown, unconsolidated silty sandstone with abundant phosphate nodules and vertebrate fossils. Bed 3 is characterised by a compact, light olive grey to dark bluish grey, poorly sorted, silty sandstone sand with phosphate pebbles and quartz discoids (Fallon and Boessenecker Citation2020). The specimen of Megalolamna from the Chandler Bridge Formation described in this paper comes from the basal 1 m of the formation, specifically from an unusual lithology recently identified by Albright et al. (Citation2019) as Bed 1A that is situated somewhere below Bed 2 (). Bed 1A at this locality consists of 2–3 m of greenish-grey, massively bedded, and sparsely fossiliferous sandy silt with occasional phosphatic clasts (RWB, pers. obs.). Although whether Bed 1A is truly a lateral equivalent or different facies of conventional Bed 1 requires further field study, in this vicinity, it directly overlies the Ashley Formation (without beds 0 or 1) and fills in a paleochannel eroded into the Ashley Formation (Albright et al. Citation2019).
The Chandler Bridge Formation is rich in vertebrate fossils, including diverse elasmobranchs (sharks and rays), bony fishes, an estuarine crocodile, sea turtles, sea birds, cetaceans (whales and dolphins), and sea cows (Sanders et al. Citation1982; Cicimurri and Knight Citation2009; Boessenecker and Geisler Citation2018). Although dinoflagellates in the formation have suggested that the Chandler Bridge Formation formed under shelf settings and transitioned towards estuarine and eventually nonmarine deposition through regression (Katuna et al. Citation1997), evidence from marine vertebrate fossils, particularly cartilaginous and osteichthyan fishes, indicates continuous marine deposition under inner to middle shelf environments with temperatures ranging from 20°C to 25°C (Cicimurri and Knight Citation2009; Fierstine and Weems Citation2009). Strontium ratios (87Sr/86Sr) from fossil oyster shells derived from the Chandler Bridge Formation provide an age determination of 24.7 Ma. In concert with a 87Sr/86Sr date of 23.5 Ma from the overlying Edisto Formation, the age of the Chandler Bridge Formation is constrained to late Chattian or latest Oligocene (Weems et al. Citation2016; Boessenecker and Fordyce Citation2017; ().
Systematic palaeontology
Class Chondrichthyes Huxley Citation1880
Subclass Elasmobranchii Bonaparte Citation1838
Cohort Euselachii Hay Citation1902
Subcohort Neoselachii Compagno Citation1977
Order Lamniformes Berg Citation1958
Family Otodontidae Glikman Citation1964
Genus Megalolamna Shimada, Chandler, Lam, Tanaka, and Ward Citation2017
Type species
Megalolamna paradoxodon Shimada, Chandler, Lam, Tanaka, and Ward Citation2017 from Pyramid Hill Sand Member (early Miocene) of Jewett Sand, Kern County, California, USA.
Emended diagnosis
Lamniform differing from all known species of otodontids by the following combination of characters: tooth consisting of a sharply pointed, relatively tall, triangular main cusp, one prominent pair of triangular lateral cusplets, and strongly bilobed root; main cusp erect, slightly inclined distally, or gently curved distally; lingual crown face very convex without ornamentation; labial crown face flat or subtly convex except centre of base with weak depression; height and width of each lateral cusplet nearly equal with tendency to point outward; both mesial and distal cutting edges of main cusp and lateral cusplets smooth and razor-like, and continuous from apex to base; main cusp and lateral cusplets nearly erect to gently curved lingually; concave crown base and distinct, chevron-shaped tooth neck on lingual face characterised as a bourlette covered with thin enameloid layer; prominent tooth neck also on labial face in tall teeth, forming rounded ledge with thin enameloid layer that grades into enameloid of main cusp and lateral cusplets; bilobed root with rounded basal tips and moderately tight basal concavity in between; root overall robust but particularly at lingual protuberance that generally exhibits one or two prominent and a few smaller nutritive foramina; root width slightly wider than total crown width; osteodentine tooth histology.
Megalolamna paradoxodon Shimada, Chandler, Lam, Tanaka, and Ward Citation2017
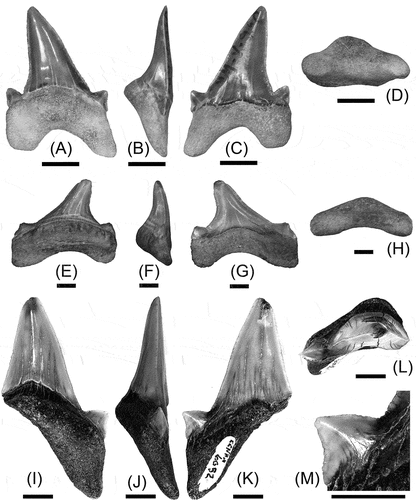
Figure 2. Teeth of Megalolamna described in this paper. (A)–(D), tooth of M. paradoxodon (CMM-V-10306) from Calvert Formation of Maryland, USA, in lingual (A), mesial (B), labial (C), and basal (D) views. (E)–(H), tooth of M. paradoxodon (CMM-V-10270) from Calvert Formation of Maryland, USA, in lingual (A), mesial (B), labial (C), and basal (D) views. (I)–(M), tooth of M. cf. M. paradoxodon (CCNHM 6052) from the Chandler Bridge Formation of South Carolina, USA, in lingual (I), mesial (J), labial (K), and apical (L) views, and close-up view of mesial lateral cusplet (M). All scale bars = 5 mm.
Material
Two isolated teeth (CMM-V-10270 and CMM-V-10306) from the Calvert Formation in Maryland housed in Calvert Marine Museum in Solomons, Maryland, USA.
Locality and stratigraphy
CMM-V-10270 was found as float at Flag Ponds Nature Park in Calvert County along the western shore of the Chesapeake Bay. Flag Ponds is situated in a low elevation gap between the cliffs, but the adjacent cliffs from which the tooth likely originated comprise Ward and Andrews’s (Citation2008) Beds 14–16A which is approximately 14.5–14 Ma in age (Kidwell et al. Citation2015). CMM-V-10306 was found in situ at Popes Creek in Charles County within the Popes Creek Sand of the Fairhaven Member, specifically in a stratigraphic horizon situated between Ward and Andrews’s (Citation2008) Beds 3A and 3B, which is dated approximately 20–19 Ma (Perez et al. Citation2019; ).
Description
CMM-V-10306 () is a complete tooth, measuring approximately 17.5 mm in total tooth height and 13.5 mm in total tooth width. Its crown consists of a relatively tall main cusp, that is gently inclined distally, and a pair of pronounced lateral cusplets, where the total crown height and total crown width measure about 12.5 mm and 12.0 mm, respectively. The mesial and distal cutting edges along the main cusp are slightly convex and slightly concave, respectively. Both cutting edges of the mesial cusplet are slightly concave, and those of the distal cusplet are gently convex. A well-marked tooth neck represented as a bourlette without a thin enameloid coating is present on the lingual side (see below for a possible explanation for the lack of an enameloid coating), whereas the crown base on the labial face is wavy but otherwise relatively straight and slightly overhangs the root. The mesial root lobe is narrower and longer than the distal root lobe, and its U-shaped basal concavity of the root is asymmetrical and moderately deep. The lingual root face is gently rounded with a bulged lingual protuberance that exhibits a few minute nutritive pores, whereas the labial root face is relatively flat.
CMM-V-10270 () is a nearly complete tooth, missing only a small portion of the crown apex. It measures about 24.0 mm in estimated total tooth height and 28.0 mm in total tooth width. Its crown consists of a distally inclined, short main cusp with a pair of low lateral cusplets, where the total crown height and total crown width measure approximately 17.0 mm and 27.5 mm, respectively. The mesial and distal cutting edges of the main cusp are straight and rather strongly concave, respectively, where their basal side continues to become the cutting edges of the lateral cusplets with practically no clear demarcation. A well-marked bourlette with a thin but well-marked enameloid coating is present on the lingual side of the tooth neck, whereas the labial crown base is gently concave. The mesial root lobe is slightly wider than the distal root lobe, and its basal concavity is shallow. The lingual root face is gently rounded with a bulged lingual protuberance that exhibits a few minute nutritive pores, whereas the labial root face is relatively flat.
Remarks
In describing a tooth of ‘Lamniformes gen. et sp. indet’. from the Uitpa Formation in northern Colombia, Carrillo-Briceño et al. (Citation2016) noted that the taxon, which is now recognised as Megalolamna paradoxodon (see Shimada et al. Citation2017, p. 714), occurs along the east coast of North America among some other localities globally and, in particular, stated that the taxon is ‘abundant’ in the Calvert Formation of Maryland (p. 86). However, they did not describe or refer to any specific specimens to substantiate the record. Although our observation is that teeth of M. paradoxodon are exceptionally rare in the Calvert Formation, CMM-V-10270 and CMM-V-10306 described here unequivocally demonstrate the presence of M. paradoxodon in the Calvert Formation of Maryland.
Whereas both CMM-V-10270 and CMM-V-10306 possess characteristics that are overall consistent with M. paradoxodon as described by Shimada et al. (Citation2017), CMM-V-10270 also exhibits a well-marked, enameloid-coated bourlette on the lingual tooth neck that has never been reported previously for the species or genus (see below for further discussion). The main cusp of both specimens shows distal inclination indicating that they both represent lateral teeth (sensu Shimada Citation2002), but the tooth with a more erect main cusp, CMM-V-10306, likely came from a more mesially located position than the other tooth (CMM-V-10270) with a strongly inclined main cusp. In fact, among all the teeth of Megalolamna described to date (Carrillo-Briceño et al. Citation2016, Citation2020; Shimada et al. Citation2017; this study), CMM-V-10270 represents a tooth from the most distal position in the dentition based on its strongly inclined, short main cusp.
Megalolamna cf. M. paradoxodon
Material
An isolated tooth (CCNHM 6052) housed in the College of Charleston’s Mace Brown Museum of Natural History, South Carolina, USA.
Locality and stratigraphy
The tooth was collected from Bed 1A of the Chandler Bridge Formation in the vicinity of the College Park subdivision in Ladson, Berkeley County, South Carolina ().
Description
CCNHM 6052 () preserves a nearly complete, triangular main cusp with a slightly chipped crown apex and parts of the cutting edges, a triangular mesial cusplet, and a mesial root lobe, and is missing the distal cusplet (that was presumably present) and distal root lobe. It is estimated to have originally measured about 36.0 mm in total tooth height and 25.4 mm in total tooth width, where the total crown width including both lateral cusplets is estimated to be about the same as the total tooth width. The crown, including the mesial cusplet, measures about 25.8 mm in estimated height, and the total enameloid height is about 27 mm including a thin enameloid coating on the labial side of the tooth neck that extends basally to the mesial cusplet. Although the main cusp is largely erect, its slightly convex mesial cutting edge and slightly concave distal cutting edge give a slightly distally inclined appearance. The mesial cusplet, which measures 3.7 mm in height and 6.1 mm in width, forms a nearly equilateral triangle, but its slightly convex distal cutting edge and slightly concave mesial cutting edge make the cusplet to point mesially, rather than apicomesially. Much of the cutting edges of the main cusp is smooth with irregular chipping, but the basal-most part of its mesial cutting edge as well as both cutting edges of the mesial cusplet show irregular notches that can be characterised as weak serrations (). The crown base on both lingual and labial faces forms a broad, apically pointed ‘V’. Besides the thin enameloid coating on the labial side of the tooth neck that extends basal to the mesial cusplet (see above), a prominent bourlette marks the lingual side of the tooth neck with a thin enameloid layer also extending below to the mesial cusplet. The preserved mesial root lobe extends basally with a relatively sharp basal point that is slightly directed labially, and the basal concavity of the root is thought to have formed a moderately deep ‘U’. The lingual face of the root is gently rounded and has a prominent lingual protuberance with at least one prominent nutritive pore, whereas the labial face of the root is relatively flat.
Remarks
Overall, CCNHM 6052 shows characteristics consistent with Megalolamna paradoxodon as described by Shimada et al. (Citation2017), but there are two additional notable differences: (1) the presence of a pronounced bourlette with a thin enameloid coating on the lingual tooth neck; and (2) the presence of weak serrations along the cutting edges of the mesial cusplet (see below for further discussion). Particularly because of the presence of serrations, the specimen is here referred to as M. cf. M. paradoxodon (see below for further discussion). The exact tooth position of CCNHM 6052 is uncertain, but the slightly distally inclined main cusp with a rather labiolingually thick crown indicates that it likely represents a mesially located lateral tooth from the lower left jaw.
Discussion
The three specimens described here all possess the original diagnostic characteristics of Megalolamna paradoxodon (Shimada et al. Citation2017), but CMM-V-10270 and CCNHM 6052 also exhibit the presence of an enameloid-covered bourlette representing the lingual tooth neck. Shimada et al. (Citation2017, p. 706) described the morphology of the crown-root transition of Megalolamna as having ‘distinct tooth neck on lingual face; prominent tooth neck also on labial face in tall teeth, forming rounded ledge with a thin enameloid layer that grades into enameloid of main cusp and lateral cusplets.’ However, the presence of a bourlette with a thin enameloid layer on the lingual side of the tooth neck was not explicitly noted. A re-examination of the original materials described by Shimada et al. (Citation2017) reveals that at least three of the five teeth (‘NCSM 30012’, ‘MUSM 3238’, and ‘KPM-NNV735’ therein; their figure 2(k)-(v)) representing specimens from North Carolina (USA), Peru, and Japan are found to possess an enameloid-covered bourlette. The holotype from California (USA) (‘UCMP 112146’) and another referred specimen of M. paradoxodon from North Carolina (‘NCSM 30000’) described by Shimada et al. (Citation2017, figure 2(a)-(j)) do not exhibit an enameloid-covered bourlette, but the junction between their tooth neck and root on the lingual face shows signs of abrasion, suggesting that a thin enameloid layer likely became lost taphonomically. The lack of enameloid layer on the bourlette in CMM-V-10306 () is also interpreted to have been lost due to taphonomic abrasion. In this paper, we thus emended the diagnosis pertaining to the lingual side of the tooth neck in the genus Megalolamna as having a ‘chevron-shaped tooth neck on lingual face characterized as a bourlette covered with thin enameloid layer’ (see above) with the understanding that the thin enameloid coating may be absent for teeth that suffered taphonomic abrasion.
CCNHM 6052 described here was found to possess fine serrations along the base of the mesial cutting edge and mesial lateral cusplet. Shimada et al. (Citation2017, p. 706) described the diagnosis pertaining to the cutting edges of the monospecific genus as ‘both mesial and distal cutting edges of main cusp and lateral cusplets smooth and razor-like, and continuous from apex to base’. Even though we have treated CCNHM 6052 as belonging to Megalolamna (specifically M. cf. M. paradoxodon) here, the specimen cannot technically even be considered as belonging to Megalolamna based on the existing generic diagnosis. However, the fossil record of Megalolamna is exceptionally sparse globally. Therefore, it is difficult to decipher the total morphological variation of teeth in Megalolamna at the present time to determine whether the diagnosis for Megalolamna needs to be emended to accommodate the possible presence of such serrations or a new genus needs to be erected to accommodate CCNHM 6052. Because the serrations are not prominent and confined to a very small portion of CCNHM 6052, we have tentatively considered the presence of such minor serrations to represent a possible variation within Megalomania but have chosen to refrain from emending the generic diagnosis until more specimens of Megalolamna become available for examination.
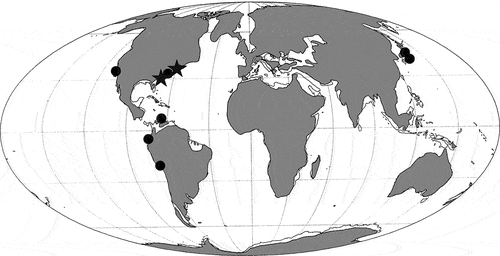
Previously, the fossil record of Megalolamna was represented by specimens from the following seven localities (): the Pungo River Formation of North Carolina, USA; Jewett Sand in southern California, USA; Uitpa Formation in northern Colombia; Dos Bocas Formation of Ecuador; the Chilcatay Formation of Peru; the Oi Formation in Mie Prefecture, Japan; and the O’oshimojo Formation in Nagano Prefecture, Japan (Carrillo-Briceño et al. Citation2016, Citation2020; Shimada et al. Citation2017). The two specimens from the Calvert Formation described in this paper, CMM-V-10270 and CMM-V-10306, are significant because they represent the northern-most occurrence of the taxon. The specimen from the Chandler Bridge Formation, CCNHM 6052, is important because, assuming that it belongs to Megalolamna, it represents geologically the oldest record for the genus. This new stratigraphic record would mean that the origin of this enigmatic genus is rooted in the Palaeogene no later than 23.5 Ma.
Figure 3. Geographic distribution of previous records (circle) and newly described records (star) of Megalolamna (Carrillo-Briceño et al. Citation2016, 2020; Shimada et al. Citation2017; this study) using early Miocene (Aquitanian‒Burdigalian) palaeogeographic map (after Smith et al. Citation1994, p. 27).
Conclusions
Megalolamna paradoxodon was previously documented from the following seven Miocene localities: the Pungo River Formation of North Carolina, USA; Jewett Sand in California, USA; Uitpa Formation in Colombia; Dos Bocas Formation of Ecuador; the Chilcatay Formation of Peru; the Oi Formation in Mie Prefecture, Japan; and the O’oshimojo Formation in Nagano Prefecture, Japan. In this study, we report three new specimens of Megalolamna collected from the eastern USA, consisting of CMM-V-10270 and CMM-V-10306 from the lower Miocene Calvert Formation of Maryland, and CCNHM 6052 from the upper Oligocene Chandler Bridge Formation in South Carolina. CMM-V-10270 represents a tooth from the most distal dental position described to date for Megalolamna. CMM-V-10270 and CCNHM 6052 possess an enameloid-covered bourlette on the lingual side of the tooth neck, which resulted in emending the diagnosis of the monospecific genus Megalolamna. CCNHM 6052 exhibits fine serrations along the base of the mesial cutting edge of the mesial lateral cusplet. The specimen is tentatively identified as M. cf. M. paradoxodon in this paper, where the presence of minor serrations as a possible intrageneric variation may need to be added to the diagnosis of Megalolamna, pending the discovery of more specimens with such serrations. The three new specimens described here are collectively important because they extend the geographic range of the genus as north as Maryland and its stratigraphic range as old as the late Oligocene.
Acknowledgments
We sincerely thank S. Basak (South Carolina resident) who donated CCNHM 6052 for the purpose of this study, J. Geisler for discussions of Chandler Bridge Formation stratigraphy, and S. J. Boessenecker (CCNHM) for access to the CCNHM collections. We also thank W. Ashby (the late Maryland resident) for collecting and donating CMM-V-10306, J. Nance and S. Groff (CMM) for their assistance in curating the specimen, and J. Cooper and two anonymous reviewers for their comments on this manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Albright LI, Sanders AE, Weems RE, Cicimurri DJ, Knight JL. 2019. Cenozoic vertebrate biostratigraphy of South Carolina, U.S.A., and additions to the fauna. Bull Florida Mus Nat Hist. 57:77–236. doi: 10.58782/flmnh.qqgg4577.
- Berg LS. 1958. System der Rezenten und Fossilen Fischartigen und Fische. Berlin: Hochschulbücher für Biologie.
- Boessenecker RW, Fordyce RE. 2017. A new eomysticetid from the Oligocene Kokoamu Greensand of New Zealand and a review of the Eomysticetidae (Mammalia, Cetacea). J Syst Palaeont. 15(6):429‒469. doi: 10.1080/14772019.2016.1191045.
- Boessenecker RW, Geisler JH. 2018. New records of the archaic dolphin Agorophius (Mammalia: Cetacea) from the upper Oligocene Chandler Bridge Formation of South Carolina, USA. PeerJ. 6:e529. doi: 10.7717/peerj.5290.
- Bonaparte CL. 1838. Selachorum tabula analytica. Nuovi Ann Sci Nat Bologna. 1(2):195–214.
- Carrillo-Briceño JD, Argyriou T, Zapata V, Kindlimann R, Jaramillo C. 2016. A new early Miocene (Aquitanian) Elasmobranchii assemblage from the La Guajira Peninsula, Colombia. Ameghiniana. 53(2):77‒99. doi: 10.5710/AMGH.26.10.2015.2931.
- Carrillo-Briceño JD, Villafaña JA, De Gracia C, Flores-Alcívar FF, Kindlimann R, Abella J. 2020. Diversity and paleoenvironmental implications of an elasmobranch assemblage from the Oligocene–Miocene boundary of Ecuador. PeerJ. 8:e9051. doi: 10.7717/peerj.9051.
- Cicimurri DJ, Knight JL. 2009. Late Oligocene sharks and rays from the Chandler Bridge Formation, Dorchester County, South Carolina, USA. Acta Palaeont Polo. 54(4):627–647. doi: 10.4202/app.2008.0077.
- Compagno LJV. 1977. Phyletic relationships of living sharks and rays. Am Zool. 17(2):303–322. doi: 10.1093/icb/17.2.303.
- Fallon BR, Boessenecker RW. 2020. Multispecies leatherback turtle assemblage from the Oligocene Chandler Bridge and Ashley formations of South Carolina, USA. Acta Palaeont Polo. 65:763–776. doi: 10.4202/app.00740.2020.
- Fierstine HL, Weems RE. 2009. Paleontology of the Oligocene Ashley and Chandler Bridge formations of South Carolina, 4: analysis and new records of billfishes (Perciformes: Xiphioidei). Palaeo Ichthyologica. 11:43‒88.
- Gernant RE, Gibson TG, Whitmore FC, editors. 1971. Environmental history of Maryland Miocene Vol. 3. Baltimore (Maryland): Maryland Geol Surv Guidebook.
- Gibson TG. 1983. Key foraminifera from upper Oligocene to lower Pleistocene strata of the central Atlantic Coastal Plain. In: Ray C, editor. Geology and paleontology of the Lee Creek Mine. North Carolina, I; pp. 355–453. Smithsonian Contrib Paleobiol 53. doi: 10.5479/si.00810266.53.1.
- Glikman LS. 1964. [Sharks of Paleogene and their stratigraphic significance]. Moscow: Nauka Press. in Russian.
- Harris GD. 1893. The Tertiary geology of Calvert Cliffs, Maryland. Am J Sci. 45(265):21–31. doi: 10.2475/ajs.s3-45.265.21.
- Hay OP. 1902. Bibliography and catalogue of the fossil vertebrata of North America. Bull US Geol Surv. 179.
- Huxley TH. 1880. On the application of the laws of evolution to the arrangement of the vertebrata, and more particularly of the Mammalia. Proc Zool Soc Lond. 43:649–661.
- Katuna MP, Geisler JH, Colquhoun DJ. 1997. Stratigraphic correlation of Oligocene marginal marine fluvial deposits across the middle and lower coastal plain, South Carolina. Sed Geol. 108(1–4):181‒194. doi: 10.1016/S0037-0738(96)00053-X.
- Kent BW. 1994. Fossil shark teeth of the Chesapeake Bay region. Columbia (Maryland): Egan Rees & Boyer.
- Kent BW. 2018. The cartilaginous fishes (chimaeras, sharks, and rays) of Calvert Cliffs. In: Godfrey S, editor. The geology and vertebrate paleontology of Calvert Cliffs. Vol. 100. Maryland, USA: Smithsonian Contrib Paleobiol; pp. 45–157.
- Kidwell SM. 1984. Outcrop features and origin of basin margin unconformities in the lower Chesapeake Group (Miocene), Atlantic Coastal Plain. In: Schlee J, editor. Interregional unconformities and hydrocarbon accumulation. Vol. 36, Tulsa (Oklahoma): Am Asso Petrol Geol Mem; pp. 37–58.
- Kidwell SM. 1989. Stratigraphic condensation of marine transgressive records: origin of major shell deposits in the Miocene of Maryland. J Geol. 97(1):1–24. doi: 10.1086/629278.
- Kidwell SM. 1997. Anatomy of extremely thin marine sequences land-ward of a passive-margin hinge zone: neogene Calvert Cliffs succession, Maryland, U.S.A. J Sed Res. 67:222–240. doi: 10.1306/D4268563-2B26-11D7-8648000102C1865D.
- Kidwell SM, Powars DS, Edwards LE, Vogt PR. 2015. Miocene stratigraphy and paleoenvironments of the Calvert Cliffs, Maryland. Geol Soc Am Field Guide. 40:231–279.
- Perez VJ, Godfrey SJ, Kent B, Weems R, Nance J. 2019. The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time. J Vert Paleont. 38(6):e1546732. doi: 10.1080/02724634.2018.1546732.
- Pierce EL, van de Flierdt T, Williams T, Hemming SR, Cook CP, Passchier S. 2017. Evidence for a dynamic East Antarctic ice sheet during the mid-Miocene climate transition. Earth Planet Sci Let. 478:1–13. doi: 10.1016/j.epsl.2017.08.011.
- Powars DS, Edwards LE, Kidwell SM, Schindler JS. 2015. Cenozoic stratigraphy and structure of the Chesapeake Bay region: In: Brezinski DK, Halka JP, Ortt RA Jr, editors. Tripping from the fall line: field excursions for the GSA Annual Meeting, Baltimore. Geol Soc Am Field Guide 40. doi: 10.1130/2015.0040(07).
- Sanders AE, Weems RE, Lemon EM Jr. 1982. The Chandler Bridge Formation; a new Oligocene stratigraphic unit in the lower coastal plain of South Carolina. Contributions To Stratigraphy, US Geol Surv Bull. 1529-H:105‒124.
- Shattuck GB. 1904. Geological and paleontological relations, with a review of earlier investigations. In: Clark W, Shattuck G, Dall W, editors. The Miocene deposits of Maryland. Maryland Geol Surv, Baltimore, Maryland; pp. 33–87.
- Shimada K. 2002. Dental homologies in lamniform sharks (Chondrichthyes: Elasmobranchii). J Morphol. 251(1):38–72. doi: 10.1002/jmor.1073.
- Shimada K, Chandler RE, Lam OLT, Tanaka T, Ward DJ. 2017. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Hist Biol. 29(5):704–714. doi: 10.1080/08912963.2016.1236795.
- Smith AG, Smith DG, Funnell BM. 1994. Atlas of Mesozoic and Cenozoic coastlines. Cambridge: Cambridge Univ Press.
- Visaggi CC, Godfrey SJ. 2010. Variation in composition and abundance of Miocene shark teeth from Calvert Cliffs, Maryland. J Vert Paleont. 30(1):26–35. doi: 10.1080/02724630903409063.
- Vogt PR, Eshelman RE, Godfrey SJ. 2018. Calvert Cliffs: eroding mural escarpment, fossil dispensary, and paleoenvironmental archive in space and time. In: Godfrey S, editor. The geology and vertebrate paleontology of Calvert Cliffs. Vol. 100. Smithsonian Contrib Paleobiol; pp. 3–44. doi:10.5479/si.1943-6688.100
- Vogt PR, Parrish M. 2012. Driftwood dropstones in Middle Miocene Climate Optimum shallow marine strata (Calvert Cliffs, Maryland Coastal Plain): erratic pebbles no certain proxy for cold climate. Palaeogeog Palaeoclimat Palaeoecol. 323:100–109. doi: 10.1016/j.palaeo.2012.01.035.
- Ward LW. 1992. Molluscan biostratigraphy of the Miocene, Middle Atlantic Coastal Plain of North America. Vol. 2. Martinsville (Virginia): Virginia Mus Nat Hist Mem.
- Ward LW, Andrews GW. 2008. Stratigraphy of the Calvert, Choptank, and St. Marys formations (Miocene) in the Chesapeake Bay area, Maryland and Virginia. Virginia Mus Nat Hist Mem. 9.
- Weems RE, Albright LB, Bybell LM, Cicimurri DJ, Edwards LE, Harris WB, Lewis WC, Osborne JE, Sanders AE, Self-Trail JM. 2016. Stratigraphic revision of the Cooper Group and the Chandler Bridge and Edisto formations in the Coastal Plain of South Carolina. South Carolina Geol. 49:1‒24.