
ABSTRACT
In this short communication, we describe the first reported case of an in-situ observation and subsequent sampling of a fossil Otodus megalodon tooth from a deep-sea locality in the Pacific Ocean (14.11332°N, 167.39357°W; 3090 m depth). The additional documentation of its location prior to collection allows for tentative conclusions on surrounding factors leading to its current state, such as sedimentation rate and manganese encrusting.
Introduction
Fossil teeth of the extinct shark species Otodus megalodon, which have been documented from numerous sites of Miocene and Pliocene sediments, indicate an almost global distribution (Cappetta Citation2012; Boessenecker et al. Citation2019).
As the last chronospecies of the Paleocene-Pliocene megatooth shark lineage, it is one of the largest known top marine predators and reached an estimated length of up to 20 m (Shimada Citation2021; Perez et al. Citation2021; Shimada et al. Citation2023). The reasons for its extinction at the transition of the early to the late Pliocene (~3.6 MA) remain enigmatic (Boessenecker et al. Citation2019). Population fragmentation caused by oceanographic changes along with competition for food with the great white shark (Carcharodon carcharias) may have driven the extinction (Boessenecker et al. Citation2019; McCormack et al. Citation2022).
However, the fossilised teeth are commonly found in marine sediments, with the majority of findings occurring in sediments that are easily accessible from land, while sediments in deep waters remain largely unsampled. The Challenger expedition (1872–1876) was the first to report fossil shark teeth in general from deep-sea sediment samples (Murray Citation1889, Citation1891). Since then, numerous expeditions have documented fossil shark teeth (Agassiz and Murray Citation1902; Eastman Citation1903, Citation1906; Belyaev and Glikman Citation1970; Richer de Forges Citation1986; Séret Citation1987), including the megatooth shark O. megalodon. All Pacific records of fossil shark teeth and O. megalodon are summarised in Pollerspöck and Straube (Citation2023a, Citation2023b). In this short communication, we add a fossil finding of O. megalodon from a deep-sea site in the Central Pacific Ocean. This finding is of special interest as the tooth was discovered during a remotely operated vehicle (ROV) dive and filmed in situ prior to being collected.
Geological settings
The sample was collected on an isolated seamount located in a geologically complex region between the Mid-Pacific Mountains and the Line Islands Ridge in the Central Pacific Ocean, which is characterised by numerous isolated seamounts interspersed among several linear submarine volcanic chains with varied orientations. The seamount is located on ocean basement created about 118 Ma (Early Cretaceous) (Müller et al. Citation2008); however, the age of the seamount volcanism is unknown. Other seamounts in the region have eruption dates ranging from 55 Ma (Eocene) to 82 Ma (Late Cretaceous) (Clouard and Bonneville Citation2005). This geologic complexity has been attributed to multiple plume interaction models (Crough and Jarrard Citation1981; Duncan and Clague Citation1985; Pockalny et al. Citation2021) or sub-lithospheric melt generated by tensional cracks (Natland and Winterer Citation2005).
Material and methods
The sample was collected during a recent expedition aboard E/V Nautilus that explored the deep-sea geology and biology around Johnston Atoll (NA141; June 19-14 July 2022). The tooth was collected on a dive using the ROV Hercules at a depth of 3090 m on an unnamed seamount located ~350 km southeast of Johnston Atoll (Dive H1926, 14.11332°N, 167.39357°W, 23 June 2022), within the Pacific Remote Islands Marine National Monument. The ROV dive during which the sample was collected surveyed a narrow ridge that runs in a southwest to northeast direction for about 5 km with slopes on the ridge crest averaging about 20 degrees.
The site where the sample was collected was a relatively flat area on the ridge crest covered with nodules of various sizes. Between these were areas of fine grained sediment. In an area with smaller nodules, the ROV sampled the seafloor sediments using a shovel. The sample (NA141-006-01) consisted of smaller nodules, fine sandy sediment and the tooth. Subsequent viewing of the high-resolution video collected by the ROV revealed that a cutting edge of the tooth was protruding from the sediment (), and the tooth was embedded upright in the sand. Prior to collecting the sample, the expedition log recorded pillow basalts welded together by ferromanganese crust that were covered by deep-sea corals. Upon approach, the flat part of the steep ridge where the tooth sample was eventually sampled was covered by nodules and rubble; the benthic fauna in this area consisted of occasional stalked crinoids, unstalked crinoids, cutthroat eels, and sea cucumbers.
Figure 1. Screenshot from the documentation video of the (ROV) Hercules at a depth of 3090 m on an unnamed seamount located ~350 km southeast of Johnston Atoll (dive H1926, 14.11332°N, 167.39357°W, 23 June 2022) showing the site with the Otodus megalodon tooth.

Once brought back to shore, the tooth was discovered during sample accessioning at the Marine Geological Samples Laboratory at the University of Rhode Island. Curators removed portions of the enclosing ferromanganese crust to expose the tooth surface, and photographed the specimen using a Nikon D500 SLR camera. Further detailed images were taken using a digital binocular camera (Leica IC80 HD, Software Leica Application Suite). Up to 13 images were taken per view, which were then combined into one image using the focus stacking software CombineZP (Hadley Citation2010). GIMP2 (https://www.gimp.org/) was used to crop the images and generate the reconstruction ().
Figure 2. Otodus megalodon tooth NA141-006-01-MT. A, labial view; B, profile view; C, lingual view; D, basal view. Scale bar 2 cm.

Figure 3. Close up images of Otodus megalodon tooth NA141-006-01-MT. A, tooth enamel; B, broken apex with cavity; C, view of the basal manganese crust; D, lateral serration.

The fossil tooth described herein is deposited at the University of Rhode Island Marine Geological Samples Laboratory in Narragansett, Rhode Island under the sample number NA141-006-01-MT (IGSN: http://doi/org/10.58052/URI00027Z).
Institutional Abbreviations
URI, University of Rhode Island; Narragansett, Rhode Island, U.S.A.
Results
Family †Otodontidae Glikman, Citation1964
Genus †Otodus Agassiz, Citation1838
Type species
†Otodus auriculatus (Blainville, Citation1818)
†Otodus (Megaselachus) megalodon Agassiz, Citation1835
()
Referred material
NA141-006-01-MT, upper tooth, Pacific Ocean (Lat, Lon: 14.1133275, −167.393574), Date/Time 23 June 2022 T 11:53:18, Temperature 1.59°C, Depth: 3089 m, Nautilus Dive Report: H1926.
Description
The dimensions of the tooth are as follows: height: 68.0 mm (calculated: 93 mm), width: 63.3 mm (calculated: 71 mm), crown height (CH): calculated: 64.0 mm, length cutting edge (labial view, right side): 66,0 mm, left side: 58,7 mm, weight: 37 gr (). Only the tooth crown is preserved, which has a triangular shape. The crown is broad, the cutting edges are clearly serrated on both sides (). The lingual side is convex, while the labial side is comparatively flat and shows a slight bulge in the middle. The tip of the tooth is broken and only the enameloid part fossilised leaving, as far as recognisable, the inner part of the capsule hollow (). All damaged areas, including the broken tip, the enameloid folds () as well as the tooth basis, are covered by a blackish manganese crust. The outer structure of this crust is granular ().
The lower edge of the crown is clearly visible when inspecting the tooth from its lingual side; the enameloid forms an arch (). Only the enameloid is preserved, the root is lacking. The cutting edges are sharply serrated. A side-facing view reveals a slight curvature of the upper crown labially.
The dental morphological characteristics of the tooth crown described here agree with the diagnosis given in Cappetta (Citation2012) for O. megalodon and we therefore assign the fossil to this species.
Discussion
It is noteworthy that the in situ documentation shows that the tooth was only partially embedded in sediment. Further, the well-preserved serration of the cutting edges suggests that the tooth did not undergo large transportation or drifting from which abrasion of the cutting edges would be expected. Exposure of the cutting edge () indicates that the sampling location did not accumulate overlying sediment, despite a minimum age of the fossil tooth of 3.5 million years of age (Boessenecker et al. Citation2019). ROV dives of the E/V Nautilus program often target steep topographical features like the ridge on which the tooth was collected, because stronger currents prevent sediment accumulation; the result is the exposure of rocks and higher abundances of suspension and filter feeders, which are a typical focus of the program.
The in situ observation shows that the tooth was partially encrusted with manganese. Accumulation of manganese around fossil nuclei has been described by Church (Citation1876). In such instances, the majority of fossil shark teeth are incomplete and represented by the enameloid only (Belyaev and Glikman Citation1970). The tooth-husks may indeed represent an ideal basis for manganese growth. The tooth described herein was sampled prior to a complete manganese encapsulation. A recent publication (Rouse and Goffredi Citation2023) reports on two annelid species, Osedax packardorum and O. talkovici, boring into Alopias vulpinus teeth at a depth of 1000 m in the Pacific, with feeding on the dentin pulp leaving the tooth crowns unharmed. Whether Osedax annelids fed on O. megalodon teeth is speculative, although the notably large teeth of megatooth sharks would certainly pose a great food source (Rouse and Goffredi Citation2023).
Pimiento et al. (Citation2016) analysed the palaeogeographic distribution of O. megalodon based on its fossil record. Results of this study suggest O. megalodon to be coastal based on the fact that the fossils originate from coastal sediments (). Records from offshore sediments were not discussed in this study. After an intensive review of the literature of historical deep-sea expeditions (Eastman Citation1903, Citation1906; Belyaev and Glikman Citation1970; Séret Citation1987), numerous further deep-sea finds surfaced in addition to the fossil described herein. All deep-sea records of O. megalodon are documented from depths of 350 to 5570 metres. A common characteristic of all deep-sea sites is their great distance to any dry land (>900 km). A possible explanation of the finding localities’ distribution could be transoceanic migration of O. megalodon (Cooper et al. Citation2022). Large-scale migratory behaviour is known from several extant lamniform sharks (e.g. Coffey et al. Citation2017; Doherty et al. Citation2019), including the Great White Shark Carcharodon carcharias (Bonfil et al. Citation2005)
Figure 4. Fossil occurrences of Otodus megalodon and Pacific records of cetacean ear bones. Blue dots: O. megalodon records from Pimiento et al. (Citation2016); yellow stars: records from Eastman (Citation1903, Citation1906), Belyaev and Glikman (Citation1970), and Séret (Citation1987); red star: this study; brown whale symbol: cetacean ear bones from Eastman (Citation1906); green whale symbols: common fossil findings of cetacean ear bones and O. megalodon from Eastman (Citation1906).
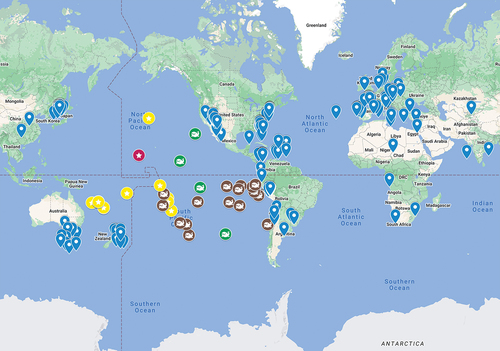
Reasons for migrations can be associated with mating behaviour, nursery areas and following prey (Weng et al. Citation2008; Fitzpatrick et al. Citation2012; Jorgensen et al. Citation2012; Domeier and Nasby-Lucas Citation2013). As O. megalodon is known to have been feeding on various whale species (Collareta et al. Citation2017; Godfrey et al. Citation2018), prey-induced migration cannot be excluded as this also aligns with an abundant fossil record of Cetacean ear bones (e.g. genera Hyperoodon (Ziphiidae) or Kogia (Kogiidae) as well as Delphinidae) at oceanic deep-sea sites in the South Pacific (Eastman Citation1906).
Conclusion
This short communication describes the first reported case of an in situ observation and subsequent sampling of a fossil O. megalodon tooth from a deep-sea locality in the Pacific Ocean. The additional documentation of its location prior to collection allows for tentative conclusions on surrounding factors leading to its current state, such as sedimentation rate and manganese encrusting. The first in situ documentation of a megatooth shark fossil from the deep sea highlights the importance of using advanced deep-diving technologies to survey the largest and least explored parts of our ocean.
Author contributions
JP took detailed images, performed all the analysis. NS and JP wrote the manuscript with contributions from all authors. DC, KK, and RR curate the specimen at the MGSL. All authors approved the submitted version.
Acknowledgments
This research used data collected by E/V Nautilus expedition NA141, which was supported by NOAA Ocean Exploration through the Ocean Exploration Cooperative Institute, and executed under permit 12543-22002 authorized by the U.S. Fish and Wildlife Service. We thank the captain and crew of E/V Nautilus, the Nautilus Corps of Exploration, the Ocean Exploration Trust, and all that supported the expedition from shore. National Science Foundation grant OCE-2116199 provides support for the curation and distribution of geological samples at the Marine Geological Samples Laboratory, University of Rhode Island, where the tooth, among other marine geological specimens, is openly accessible for scientific research and education. DAE would like to thank the Save Our Seas Foundation Grant 594. We would like to thank Charlie Underwood, Jack Cooper and another anonymous reviewer for their constructive comments on the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Agassiz L. 1835. Recherches sur les poissons fossiles. 5th livraison (June 1835). Petitpierre et Prince (text) and H. Nicolet (plates). Neuchâtel, Imprimerie de Petitpierre.
- Agassiz L. 1838. Recherches sur les poissons fossiles. 11th livraison (November 1838). Petitpierre et Prince (text) and H. Nicolet (plates). Neuchâtel, Imprimerie de Petitpierre.
- Agassiz A, Murray J. 1902. Prelimiпary report and list or stations with remarks on the deep-sea deposits. Memoirs of The Museum of Comparative Zoology at Harvard College. 26(1):1–114.
- Belyaev GM, Glikman LS. 1970. The teeth of sharks on the floor of the Pacific Ocean. Trudy Instituta Okaenologii, Academiia Nauk SSSR. 88:236–276.
- Blainville deHMD. 1818. Sur les ichthyolites ou les poissons fossiles. Nouveau Dictionnaire d’Histoire Naturelle. 27:310–391.
- Boessenecker RW, Ehret DJ, Long DJ, Churchill M, Martin E, Boessenecker SJ. 2019. The early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific. PeerJ. 7:Article e6088. doi: 10.7717/peerj.6088.
- Bonfil R, Meyer MA, Scholl MC, Johnson RL, O’Brian S, Oosthuizen WH, Swanson S, Kotze D, Patterson M. 2005. Transoceanic migration, long-distance return migration and local movement patterns in the great white shark. Sci. 301(5745):100–103. doi: 10.1126/science.1114898.
- Cappetta H. 2012. Handbook of Paleoichthyology, Vol. 3E: Chondrichthyes Mesozoic and Cenozoic Elasmobranchii: Teeth. Munich: Pfeil Verlag.
- Church AH. 1876. Manganese in the sea. Mineralogical Magazine and Journal of the Mineralogical Society. 1(2):50–53. doi: 10.1180/minmag.1876.001.2.06.
- Clouard V, Bonneville A. 2005. Ages of seamounts, islands, and plateaus on the Pacific plate. Special Papers-Geological Society of America. 388:71–90. doi: 10.1130/0-8137-2388-4.71.
- Coffey DM, Carlisle AB, Hazen EL, Block BA. 2017. Oceanographic drivers of the vertical distribution of a highly migratory, endothermic shark. Sci Rep. 7(1):Article 10434. doi: 10.1038/s41598-017-11059-6.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M, Bianucci G. 2017. Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeogr Palaeoclimatol Palaeoecol. 469:84–91. doi: 10.1016/j.palaeo.2017.01.001.
- Cooper JA, Hutchinson JR, Bernvi DC, Cliff G, Wilson RP, Dicken ML, Menzel J, Wroe S, Pirlo J, Pimiento C. 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Sci Adv. 8(33):Article eabm9424. doi: 10.1126/sciadv.abm9424.
- Crough ST, Jarrard RD. 1981. The Marquesas-Line Swell. J Geophys Res. 86(B12):11763–11771. doi: 10.1029/JB086iB12p11763.
- Doherty PD, Baxter JM, Godley BJ, Graham RT, Hall G, Hall J, Hawkes LA, Henderson SM, Johnson L, Speedie C, et al. 2019. Seasonal changes in basking shark vertical space use in the north-east Atlantic. Mar Biol. 166(10):Article 129. doi: 10.1007/s00227-019-3565-6.
- Domeier ML, Nasby-Lucas N. 2013. Two-year migration of adult female white sharks (Carcharodon carcharias) reveals widely separated nursery areas and conservation concerns. Anim Biotelemetry. 1(1):Article 2. doi: 10.1186/2050-3385-1-2.
- Duncan RA, Clague DA. 1985. Pacific plate motion recorded by linear volcanic chains. In: Nairn AEM, Stehli FG, Uyeda S, editors. The Ocean Basins and Margins: volume 7A the Pacific Ocean. Boston, MA: Springer US; pp. 89–121.
- Eastman CR. 1903. Sharks’ teeth and Cetacean bones from the Red Clay of the tropical Pacific. Memoirs of the Museum of Comparative Zoology at Harvard College. 26(4):178–189.
- Eastman CR. 1906. Sharks’ teeth and Cetacean bones from the Red Clay of the tropical Pacific. Bulletin of the Museum of Comparative Zoology at Harvard College. 50(4):75–98.
- Fitzpatrick R, Thums M, Bell I, Meekan MG, Stevens JD, Barnett A, Hays GC. 2012. A comparison of the seasonal movements of tiger sharks and green turtles provides insight into their predator-prey relationship. PLoS ONE. 7(12):Article e51927. doi: 10.1371/journal.pone.0051927.
- Glikman LS. 1964. (Sharks of Paleogene and their stratigraphic significance). «in russian». Moscou: Nauka Press.
- Godfrey SJ, Ellwood M, Groff S, Verdin MS. 2018. Carcharocles-bitten odontocete caudal vertebrae from the Coastal Eastern United States. Acta Palaeontol Pol. 63(3):463–468. doi: 10.4202/app.00495.2018.
- Hadley A. (2010). CombineZp, Image Stacking Software. [last access 2016 Apr 1]. https://hadleyweb.pwp.blueyonder.co.uk.
- Jorgensen SJ, Arnoldi NS, Estess EE, Chapple TK, Rückert M, Anderson SD, Block BA, Goldstien SJ. 2012. Eating or meeting? Cluster analysis reveals intricacies of white shark (Carcharodon carcharias) migration and offshore behavior. PLoS ONE. 7(10):Article e47819. doi: 10.1371/journal.pone.0047819.
- McCormack J, Griffiths ML, Kim SL, Shimada K, Karnes M, Maisch H, Pederzani S, Bourgon N, Jaouen K, Becker MA, et al. 2022. Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nat Commun. 13(1):Article 2980. doi:10.1038/s41467-022-30528-9.
- Müller RD, Sdrolias M, Gaina C, Roest WR. 2008. Age, spreading rates, and spreading asymmetry of the world’s ocean crust. Geochemistry, Geophysics, Geosystems. 9(4):Article Q04006. doi: 10.1029/2007GC001743.
- Murray JA. 1889. On marine deposits in the Indian, Southern, and Antarctic oceans. Scott Geogr Mag. 5(8):405–436. doi: 10.1080/00369228908732390.
- Murray JA. 1891. Report on Deep-Sea Deposits Based on the Specimens Collected during the Voyage of HMS Challenger in the Years 1872 to 1876. Deep-Sea Deposits. London: Majesty’s Goverment.
- Natland JH, Winterer EL. 2005. Fissure control on volcanic action in the Pacific. Special Papers-Geological Society of America. 388:687–710. doi: 10.1130/0-8137-2388-4.687.
- Perez VJ, Leder RM, Badaut T. 2021. Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions. Palaeontol Electron. 24(1):Article a09. doi: 10.26879/1140.
- Pimiento C, MacFadden BJ, Clements CF, Varela S, Jaramillo C, Velez-Juarbe J, Silliman BR. 2016. Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. J Biogeogr. 43(8):1645–1655. doi: 10.1111/jbi.12754.
- Pockalny R, Barth G, Eakins B, Kelley KA, Wertman C. 2021. Multiple melt source origin of the Line Islands (Pacific Ocean). Geology. 49(11):1358–1362. doi: 10.1130/G49306.1.
- Pollerspöck J, Straube N. (2023a). Pacific Records of Fossil Shark Teeth. [Data set]. https://www.google.com/maps/d/edit?mid=1DzptDIUdLc1uUQsz9hRK3Klw4ffsJMo&usp=sharing.
- Pollerspöck J, Straube N. (2023b). Records of Otodus Megalodon and Cetacean Ear Bones. [Data set]. https://www.google.com/maps/d/edit?mid=1mvcAiY5A9EcOEqIkA2XXXEgIVGJW9eg&usp=sharing.
- Richer de Forges B. 1986. La campagne MUSORSTOM 4 en Nouvelle-Caléonie, mission du N.O. “Vauban”, septembre-octobre 1985. Rapports Scientifiques et Techniques. 38:1–31.
- Rouse GW, Goffredi SK. 2023. Osedax (Siboglinidae: Annelida) utilizes shark teeth for nutrition. J Mar Biol Assoc Uk. 103:e35. doi: 10.1017/S0025315423000243.
- Séret B. 1987. Découverte d’une faune à Procarcharodon megalodon (Agassiz, 1835) en Nouvelle-Calédonie (Pisces, Chondrichthyes, Lamnidae). Cybium. 11(4):389–394.
- Shimada K. 2021. The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. Hist Biol. 33(7):904–911. doi: 10.1080/08912963.2019.1666840.
- Shimada K, Maisch HM, Perez VJ, Becker MA, Griffiths ML. 2023. Revisiting body size trends and nursery areas of the Neogene megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), reveals Bergmann’s rule possibly enhanced its gigantism in cooler waters. Hist Biol. 35(2):208–217. doi: 10.1080/08912963.2022.2032024.
- Weng KC, Foley DG, Ganong JE, Perle C, Shillinger GL, Block BA. 2008. Migration of an upper trophic level predator, the salmon shark Lamna ditropis, between distant ecoregions. Mar Ecol Prog Ser. 372:253–264. doi: 10.3354/meps07706.