
ABSTRACT
Supposed dinosaur remains were collected between 1859 and 1906 in the Lower Cretaceous Recôncavo Basin (Northeast Brazil). Since these materials remained undescribed, and most were considered lost. Recently, some of these historical specimens were rediscovered in the Natural History Museum of London, providing an opportunity to revisit them after 160 years. The specimens come from five different sites, corresponding to the Massacará (Berriasian-Barremian) and Ilhas (Valanginian-Barremian) groups. Identified bones comprise mainly isolated vertebral centra from ornithopods, sauropods, and theropods. Appendicular remains include a theropod pedal phalanx, humerus, and distal half of a left femur with elasmarian affinities. Despite their fragmentary nature, these specimens represent the earliest dinosaur bones discovered in South America, enhancing our understanding of the Cretaceous dinosaur faunas in Northeast Brazil. The dinosaur assemblage in the Recôncavo Basin resembles coeval units in Northeast Brazil, such as the Rio do Peixe Basin, where ornithopods coexist with sauropods and theropods. This study confirms the presence of ornithischian dinosaurs in Brazil based on osteological evidence, expanding their biogeographic and temporal range before the continental rifting between South America and Africa. Additionally, these findings reinforce the fossiliferous potential of Cretaceous deposits in Bahia State, which have been underexplored since their initial discoveries.
Introduction
The Cretaceous strata in Brazil cover approximately 19% of its territory (Schobbenhaus and Neves Citation2003; Da Silva et al. Citation2003) and have yielded a noteworthy ichnological and osteological dinosaur fossil record (Campos and Kellner Citation1991; Kellner and Campos Citation1999, Citation2000; Bittencourt and Langer Citation2011). The main dinosaur-bearing localities come from Lower Cretaceous deposits, especially in the Northeast region of the country (; see also Supplementary Data). The dinosaur components are mainly represented by rebbachisaurids (Lindoso et al. Citation2019) and titanosaurian sauropods (Zaher et al. Citation2011; Ghilardi et al. Citation2016; Carvalho et al. Citation2017), associated with abelisauroid (Zaher et al. Citation2020; De Souza et al. Citation2021), spinosaurid (Kellner and Campos Citation1996; Sues et al. Citation2002, França et al. Citation2022; Lacerda et al. Citation2023), carcharodontosaurid (Medeiros et al. Citation2014, Citation2019; Pereira et al. Citation2020), megaraptoran (Aranciaga-Rolando et al. Citation2018), and coelurosaurian theropods (Kellner Citation1999; Naish et al. Citation2004; Sayão et al. Citation2020), including avialans (Carvalho et al. Citation2015, Citation2021).
Figure 1. Cretaceous deposits from Brazil and their dinosaur assemblages, emphasising occurrences from the Lower Cretaceous units. 1–3, Botucatu Formation (Paraná Basin); 4–5, Goio Erê and Rio Paraná formations (Caiuá Group); 6–8, Abaeté, Quiricó, and Três Barras formations (Sanfranciscana Basin); 9–10, Maceió and Feliz Deserto formations (Sergipe-Alagoas Basin); 11, Antenor Navarro, Sousa, and Rio Piranhas formations (Rio do Peixe basins); 12, Quixoá (Icó) and Malhada Vermelha formations (Iguatu basins); 13, Rio da Batateira, Crato, and Romualdo formations (Araripe Basin); 14, Açu Formation (Potiguar Basin); 15, Itapecuru Formation (Parnaíba Basin); 16, Alcântara Formation (São Luís-Grajaú Basin). The asterisk exhibits the occurrences from the Recôncavo Basin presented by this work. Detailed information on each occurrence is provided in the Supporting Information (File S1). Modified from ‘Carta Geológica do Brasil ao Milionésimo’ (Souza et al. Citation2004).
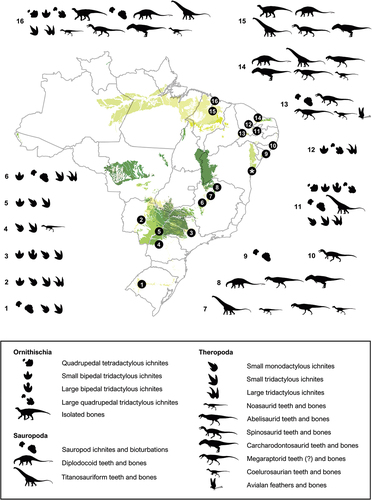
Regarding the ornithischian record, the occurrences are scarce and most of them doubtful. They include putative iguanodontian centra coming from the Recôncavo Basin (Mawson and Woodward Citation1907), an ornithischian ischium (MN 7021-V) – later reinterpreted as a spinosaurid rib (Leonardi and Borgomanero Citation1981; Machado and Kellner Citation2007)—, and a partial preserved caudal vertebra (UFMA 1.10.240) from the Alcântara Formation (Medeiros and Schultz Citation2002; Medeiros et al. Citation2007), which was briefly described and assigned to cf. Ouranosaurus by Novas (Citation2009). In contrast, the ichnological record concerning this group is abundant and widespread amongst the Brazilian Lower Cretaceous deposits, mainly in the small half-grabens of the Rio do Peixe and Iguatu basins. This record includes ichnites of putative thyreophorans, small-to-medium-sized bipedal and graviportal ornithopod trackmakers (Leonardi Citation1994; Leonardi and Carvalho Citation2002, 2021).
Although most of these dinosaur-bearing units have been extensively explored over the last decades, the dinosaur components of the Recôncavo Basin units remain poorly understood. Most of the data regarding the Recôncavo Basin dinosaurs come from historical publications older than a century ago (e.g. Allport Citation1860; Marsh Citation1869; Woodward Citation1888; Mawson and Woodward Citation1907). In addition, most of these materials are considered lost by subsequent works (e.g. Kellner and Campos Citation2000). Although some re-evaluations of other archosaurs from the Recôncavo Basin have been performed in recent years (e.g. Rodrigues and Kellner Citation2010; Souza et al. Citation2015, Citation2019), the putative dinosaur material remained undescribed in detail, with few mentions of these specimens in the literature (e.g. Bittencourt and Langer Citation2011).
Notwithstanding, some of these historical materials were recently located in the palaeontological collection of the Natural History Museum of London (NHM) and are presented herein. The specimens described below represent the first dinosaur bones collected in South America (e.g. Allport Citation1860) and revealed a diversified dinosaur fauna in the Recôncavo Basin. This historical dinosaur bones enhance our understanding about the faunal succession in Gondwana during the earliest Cretaceous. Furthermore, this work aims to encourage Brazilian palaeontologists to conduct further fieldwork in the basins of Bahia State, as they represent one of the earliest Cretaceous faunas in South America and indicates potential new important discoveries (e.g. Souza and Campos Citation2018).
Historical background and geological settings
The Recôncavo Basin is a NW – SE-oriented half-graben located in Bahia State, covering approximately 12.000 km2 and reaching about 10 km of maximum thickness (Prates and Fernandez Citation2015). The basement of this basin is composed of Neoproterozoic rocks from the São Francisco Craton, characterised by a network of high-dip synthetic normal faults divided into three structural blocks: Low, High, and Platform (Destro et al. Citation2003). The depositional history of the Recôncavo Basin can be separated into five phases: a syneclysis stage in the Permian, a pre-rift stage during the Tithonian to Early Berriasian (Dom João Local Age), a syn-rift stage starting in the Early Berriasian and extending to the Early Aptian (Rio da Serra, Aratu, Buracica, and Jiquiá local ages), and a post-rift stage ranging from the Late Aptian to Early Albian (Alagoas Local Age), later covered by Neogene (Miocene and Pliocene) and Quaternary deposits (Viana et al. Citation1971). The syn-rift sequence of the Recôncavo Basin is commonly referred in the literature as the ‘Bahia Supergroup’, which comprises the Santo Amaro Group (Itaparica, Água Grande, Candeias and Maracangalha formations), the Ilhas Group (Marfim, Pojuca and Taquipe formations), the Massacará Group (Salvador and São Sebastião formations), and the Marizal Formation (Caixeta et al. Citation1994).
According to Da Silva et al. (Citation2007), the beginning of the syn-rift phase resulted in the formation of several tectonic lakes with differential subsidence, represented by the Candeias and Maracangalha formations. The Tauá and Gomo members (Candeias Formation) are deposited under lacustrine conditions, characterised by clays rich in organic matter. They are associated with gravitational flows and fluvial system tracts of the Pitanga and Caruaçu members (Maracangalha Formation), which are related to the reactivation of transcurrent faults. By contrast, the Ilhas Group encompasses a prograding deltaic paleoenvironment, based on the succession of sandstones and shales (e.g. Marfim Formation), as well as mudstones and limestones (e.g. Pojuca Formation). The Catu Member (Marfim Formation) indicates several changes in the development of lacustrine systems. The high relief formed by the Salvador Fault on the eastern margin gradually intruded flagstones and conglomeratic sandstone facies of the Sesmaria Member (Salvador Formation) into these tectonic lakes. The establishment of the prograding deltaic system of the Pojuca Formation indicates episodic marine incursions (Santiago Member) that supplied these tectonic lakes. A subsidence decrease, which probably occurred during the Early Aptian (Arai Citation2014; but see also Assine et al. Citation2016; Parméra et al. Citation2019), progressively silted up these lacustrine systems with fluvial sediments from the São Sebastião Formation. In the western part of the basin, erosive processes on the Ilhas Group developed a canyon system that is filled by the alluvial fans deposits of the Taquipe Formation. The post-rift phase, represented by the Marizal Formation, was deposited through the formation of alluvial and delta fans.
The deposits of Recôncavo Basin hold a relatively diversified fossil record, including microfossils (e.g. foraminifera, ostracods, and ‘conchostraceans’), molluscs (e.g. bivalves and gastropods), and plant remains, such as palynomorphs, amber, and silicified conifer logs (Milhomem et al. Citation2003). Vertebrate remains are represented by cephalic fins of hybodontiform sharks (Acrodus nitidus Woodward Citation1888), several occurrences of actinopterygians (e.g. Ellimmichthys longicostatus (Cope Citation1886); ‘Lepidotes’ mawsoni Woodward Citation1888) and sarcopterygians (e.g. Mawsonia gigas Mawson and Woodward Citation1907), remains of chelonians and putative plesiosaurians (Marsh Citation1869; Woodward Citation1891), some crocodilian teeth and bones (e.g. ‘Thoracosaurus bahiaensis’, Sarcosuchus hartti; Souza et al. Citation2015; Souza Citation2019), pterosaurian teeth (Woodward Citation1891; Rodrigues and Kellner Citation2010), and supposed dinosaur specimens (Allport Citation1860; Hartt Citation1870; Mawson and Woodward Citation1907; Figueiredo and Gallo Citation2021).
The outcrops from which the historical material was collected are recognised by some studies as belonging to the ‘Bahia Supergroup’, but without an assignment to a specific horizon (e.g. Rodrigues and Kellner Citation2010; Bittencourt and Langer Citation2011). Hartt (Citation1870) and Hartt and Agassiz (Citation1875) regarded these deposits as potentially equivalent to the ‘Neocomian’ strata of Europe. Later, Derby (Citation1878) assigned the entire coastal basin from Bahia into the Lower Cretaceous. More recently, Souza et al. (Citation2015) suggested that these historical localities probably belong to the Late Valanginian to Early Barremian Ilhas Group (Caixeta et al. Citation1994). However, given the high urbanisation of the Salvador area and the absence of a refined stratigraphic control during the collection of these materials, Souza et al. (Citation2019) suggested some cautiousness to assign such rocks to any more specific unit and age.
Nevertheless, we bring here some novelties based on the descriptions of NHM collection labels, in which explain the specific lithofacies that each specimen recovered and allowing a more refined horizon assignment. According to the collection labels, the specimens reviewed here were recovered from five different localities through field seasons from 1859 to 1906 (). The outcrops were first described by Allport (Citation1860) and later explored by Mawson, in which compiled them into a detailed map in Mawson and Woodward (Citation1907). We evaluated these sites and compared the data of collection labels within stratigraphic studies of the Recôncavo Basin (e.g. Da Silva et al. Citation2007), plotting them at the ‘Carta Geológica do Brasil ao Milionésimo’ (Souza et al. Citation2004). The revised map () shows that the prospected localities match with the outcrops of the Ilhas Group and the Salvador Formation, correlating with the specific lithofacies in which the specimens were found ().
Figure 2. Historical fossil-bearing localities of the Lower Cretaceous Recôncavo Basin in Northeast Brazil, modified from Allport (Citation1860) and Mawson and Woodward (Citation1907). Localities which have yielded dinosaur remains are described in Table 1. Geographic range of the geological units was taken from ‘Carta Geológica do Brasil ao Milionésimo’ (Souza et al. Citation2004). Scale bar = 25 km.
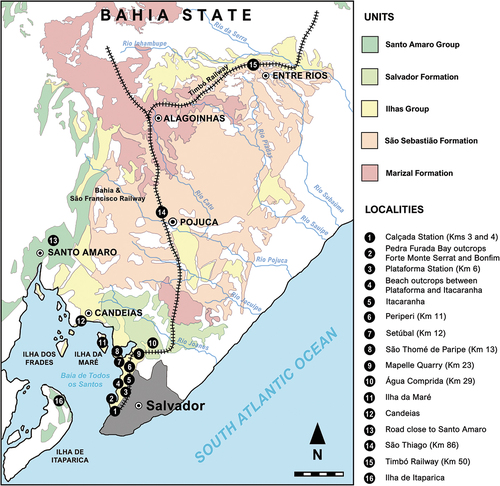
Figure 3. Stratigraphic chart of the Lower Cretaceous Massacará and Ilhas groups (Recôncavo Basin), highlighting their vertebrate assemblages (coloured boxes). 1, hybodontiforms (Acrodus nitidus); 2, semionotids (‘Lepidotes’ spp.); 3, mawsoniids (Mawsonia gigas); 4, undetermined testudines; 5, pholidosaurids (Sarcosuchus hartti); 6, putative gavialoids (‘Thoracosaurus bahiaensis’); 7, pterosaurs (anhanguerids); 8, elasmarians (Tietasaura derbyiana gen. et sp. nov.); 9, iguanodontians; 10, diplodocoids; 11, spinosaurids; 12, carcharodontosaurians; 13, undetermined small theropods (tyrannosauroids?); 14, titanosaurs (lithostrotians); 15, amiids (Calamopleurus mawsoni); 16, cladocyclids (Cladocyclus mawsoni); 17, aspidorhynchids (‘Belonostomus’ carinatus); 18, undetermined clupeomorphs (Scrombroclupeoides scutata); 19, ellimmichthyiforms (Ellimmichthys longicostatum, Ellimmichthys spinosus, Scutatuspinosus itapagipensis). References regarding the vertebrate occurrences were taken through bibliographic survey (see text). Geological chart modified from Da Silva et al. (Citation2007). Silhouettes from several sources.
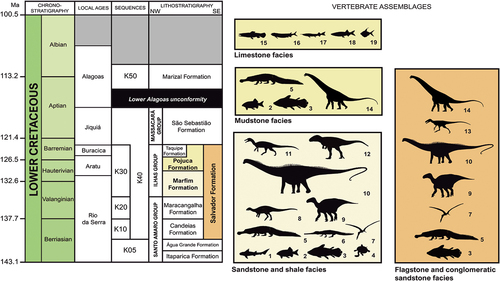
Table 1. Collection details of the specimens described in this work.
Institutional abbreviations
CPPLIP, Centro de Pesquisas Paleontológicas Llewellyn Ivor Price, Peirópolis, Brazil; GSI, Geological Survey of India, Kolkata, India; IRSNB, MACN, Museo Argentino de Ciencias Naturales ‘Bernadino Rivadavia’, Buenos Aires, Argentina; MCF-PVPH, Museo ‘Carmen Funes’ - Paleontología de Vertebrados Plaza Huincul, Plaza Huincul, Argentina; Museo de La Plata, La Plata, Argentina; MN, Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; MPM, Museo Regional Provincial ‘Padre Jesús Molina’, Río Gallegos, Argentina; MUCPv, Museo de la Universidad Nacional del Comahue, Neuquén, Argentina; NHM, Natural History Museum, London, United Kingdom; SGB, Serviço Geológico Brasileiro, Rio de Janeiro, Brazil; SHN, Sociedade de História Natural, Torres Vedras, Portugal; UFMA, Universidade Federal do Maranhão, São Luís, Brazil; UFRJ, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; UNPSJB, Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina. CPPLIP, Centro de Pesquisas Paleontológicas Llewellyn Ivor Price, Peirópolis, Brazil; GSI, Geological Survey of India, Kolkata, India; IRSNB, MACN, Museo Argentino de Ciencias Naturales ‘Bernadino Rivadavia’, Buenos Aires, Argentina; MCF-PVPH, Museo ‘Carmen Funes’ - Paleontología de Vertebrados Plaza Huincul, Plaza Huincul, Argentina; MLP, Museo de La Plata, La Plata, Argentina; MN, Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; MPM, Museo Regional Provincial ‘Padre Jesús Molina’, Río Gallegos, Argentina; MUCPv, Museo de la Universidad Nacional del Comahue, Neuquén, Argentina; NHM, Natural History Museum, London, United Kingdom; SGB, Serviço Geológico Brasileiro, Rio de Janeiro, Brazil; SHN, Sociedade de História Natural, Torres Vedras, Portugal; UFMA, Universidade Federal do Maranhão, São Luís, Brazil; UFRJ, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; UNPSJB, Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina.
Systematic Palaeontology
Dinosauria Owen Citation1842
Ornithischia Seeley Citation1888
Genasauria Sereno Citation1986
Neornithischia Cooper Citation1985
Cerapoda Sereno Citation1986
Ornithopoda Marsh Citation1881
Elasmaria Calvo et al. Citation2007
Tietasaura gen. nov.
urn:lsid:zoobank.org:act:814725BB-A522-465F-975D-1755A11C3956
Etymology
The generic epithet is a combination of Tieta (nickname for Antonieta in Portuguese) and -saura (σαύρα), the genitive form of -saurus and meaning lizard in ancient Greek. The name Tieta honours the main character from the homonymous novel ‘Tieta do Agreste’ by the famous author Jorge Amado, who was born in Bahia and lived in Salvador City. The name Antonieta further means ‘priceless’, alluding to the value of Tietasaura derbyiana sp. nov. as the first nominal ornithischian species from Brazil.
Type species
Tietasaura derbyiana sp. nov.
Diagnosis
As for type and only known species, by monotypy.
Tietasaura derbyiana sp. nov.
(; Table S3)
Figure 4. Holotypic distal half of the left femur of Tietasaura derbyiana gen. et. sp. nov. (NHM-PV R.3424) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at Plataforma Station (Locality 3). A, anterior; B, medial; C, posterior; D, lateral; E, distal views. Anatomical abbreviations: ali, anterior linea muscularis; cd, condylid process; cr, crest; fc, fibular condyle; gr, groove; ieg, intracondylar extensor groove; iff, intracondylar flexor fossa; lfs, lateral fossa; rg, rugosity; sc, sulcus; sf, spiral fracture; scr, supracondylar ridge; st, striae; tc, tibial condyle. Scale bar = 100 mm.

urn:lsid:zoobank.org:act:52683BA8-C4C5-4A81-90 DB-3BC6431946D2
Etymology
The specific epithet is an eponym honouring Orville A. Derby (1851–1915), founder and the first director from Brazilian Mineralogical and Geological Commission (Serviço Geológico e Mineralógico do Brasil, nowadays SGB), being also the former director of the National Museum of Rio de Janeiro (MN) and one of the pioneers of palaeontology in the Recôncavo Basin. Despite all tragedies in his life and the blatant lack of governmental support, Derby valiantly fought for the scientific progress of the Brazilian geosciences.
Holotype
NHM-PV R.3424, represented by a distal half of a small left femur.
Diagnosis
Small sized elasmarian ornithopod exhibiting an unique combination of character states on the femur (putative autapomorphies marked with an asterisk): presence of a marked anterior linea muscularis followed by several longitudinal striae converging distally towards the intercondylar extensor groove; broad but shallow intercondylar extensor groove; stout supracondylar ridges that extends medially; fibular supracondylar ridge sinuous and bearing a lateral fossa*; hemispherical distal femoral condyles in the posterior view, being the tibial condyle twice as large as fibular condyle; distinct prominent crest in the median margin of the tibial condyle*; fibular condyle with straight lateral margin and continuous in the distal view, lacking an indentation formed by a condyloid (rectangular) process; presence of an offset condylid, medial to the fibular condyle; broad and deep intercondylar flexor fossa, subtriangular in shape and much extending into the diaphysis.
Type locality and horizon
The holotype of Tietasaura derbyiana was recovered at a beach near the Plataforma Station (Locality 3), Salvador City, Bahia State. The shale facies outcropping in this locality are associated with the Valanginian – Hauterivian Marfim Formation (Ilhas Group, Recôncavo Basin).
Description and comparisons
Tietasaura derbyiana comprise a moderately preserved distal end of a small left femur (231.09 mm in preserved length; see Table S3), with less than a half of its total proximodistal length. This specimen was designated as a Hyposaurus sp. in the collection labels of the NHM, a dyrosaurid crocodyliform genus present in the Upper Cretaceous to Early Paleocene strata of North America, North Africa, and Brazil (e.g. Veloso et al. Citation2023). Here, however, we reinterpret it as belonging to an elasmarian ornithopod based on several characteristics (see below). When compared with crocodyliform femora, remarkable anatomical distinctions are observable in Tietasaura derbyiana. In the anterior view, for example, the femur NHM-PV R.3424 exhibits a slightly convex surface towards the distal portion, with the extensor groove more developed on the distal surface than the anterior one. By contrast, in Coelognathosuchia taxa (Goniopholidae+Tethysuchia sensu Martin et al. Citation2014), such as Burkesuchus mallingrandensis (see Novas et al. Citation2021, fig. 5) and Anteophthalmosuchus hooleyi (Martin et al. Citation2016, fig. 16), Hyposaurus rogersii (Pellegrini et al. Citation2021, fig. 1), and an unnamed dyrosaurid from Senegal (Martin et al. Citation2019, fig. 6), the extensor groove is broad and extends proximally in the diaphysis. In the posterior view, Tietasaura derbyiana has a remarkably deep and broad flexor fossa, whereas in coelognathosuchians this region is more restricted, developing as a vertical groove. Additionally, the fibular condyle in coelognathosuchians is much more developed distally than the tibial one, contrasting with the inverse pattern observed in Tietasaura derbyiana. Finally, in the distal view, the femoral epiphysis of Tietasaura derbyiana is wider mediolaterally, while in crocodyliform femora it develops anteroposteriorly.
Although the bone exhibits signs of pre-burial transportation, its surface is well preserved, and the periosteum preserves several muscular striae and rugosities. Moreover, the diaphysis in Tietasaura derbyiana shows a spiral fracture () typical of fresh bone breakage (e.g. Galloway Citation1999). In the median portion of the diaphyseal surface, the periosteum is well-preserved and particularly smooth. This feature could be interpreted as an indicator of osteological maturity, contrasting with the long-grained or dimpled patterns seen in immature specimens of pterosaurs (Bennett Citation1993), ceratopsians (Tumarkin‐Deratzian Citation2009), and birds (Tumarkin-Deratzian et al. Citation2006). Comparing with other dinosauromorphs, the bone cortex thickness in NHM-PV R.3424 further indicates that the individual reached its asymptotic mass (Sander and Tückmantel Citation2003; Griffin et al. Citation2019). However, due to the unavailability of paleohistological or CT-scan data for this individual, a precise ontogenetic assessment for Tietasaura derbyiana cannot be assumed. Therefore, such inferences are considered tentative and should be approached with caution (see Griffin et al. Citation2021 for an overview).
The diaphysis, as preserved, is swelled towards the distal end surface in anterior and posterior views (). In the lateral and medial views, the preserved shaft exhibits an accentuated posterior curvature, indicating a sigmoid profile for the femur (). This feature is widely distributed, being present in some early-diverging ornithischians, such as Eocursor parvus Butler et al. Citation2007 and Pisanosaurus mertii Casamiquela Citation1967 (see Butler Citation2010; Agnolín and Rozadilla Citation2018), thescelosaurids (Gilmore Citation1915; Scheetz Citation1999; Boyd et al. Citation2009; Brown et al. Citation2011, Citation2013), and some elasmarians, such as Anabisetia saldiviai Coria and Calvo Citation2002.
The anterior surface in Tietasaura derbyiana is almost planar, except for a shallow ‘V-shaped’ intercondylar extensor groove that does not reach the fibular and tibial condyles in the distal view (). This condition differs from the condition observed in most theropods, in which this region presents a deep and elongated groove (Carrano et al. Citation2012). This condition resembles the morphology found in thescelosaurids and ornithopods (e.g. Krumenacker Citation2017; Madzia et al. Citation2018). In addition, the anterior surface exhibits remarkable anterior linea intermuscularis and several striae, which could be associated with M. femorotibialis. Both condyles are visible in the anterior view, being the tibial more bevelled and extending further distally than the fibular one, but not reaching the diaphyseal portion of the distal end ().
In the posterior view (), the tibial condyle is much larger than the fibular one, but both are broad and hemispherical in shape. In lateral and medial views (), a pair of prominent supracondylar ridges are present, enclosing a fully opened, deep posterior intercondylar flexor fossa. This fossa is vertically developed and tapers towards the diaphysis acquiring a ‘V-shaped’ aspect. In the fibular condyle, the supracondylar ridge acquires a sinuous profile in the posterior view (). In contrast, the supracondylar ridge is confluent and merges with the condyle in the lateral view (). The tibial condyle is slightly oblique, posteromedially inclined, with a round lateral aspect. In distal view (), the fibular condyle has a straight and continuous lateral margin, lacking an indentation that detaches a condyloid process parasagittal to the femoral shaft (–G). In contrast, medially to the fibular condyle, a small accessory condylid is present, as in Notohypsilophodon comodorensis Martínez Citation1998 ().
Figure 5. Schematic draw comparing selected elasmarian femora in (1) posterior and (2) distal views. A, Tietasaura derbyiana gen. et. sp. nov. (NHM-PV R.3424); B, Notohypsilophodon comodorensis (UNPSJB-Pv 942); C, Gasparinisaura cincosaltensis (MUCPv-208); D, Fulgurotherium australe (NHM-PV R.3719); E, Trinisaura santamartensis (MLP 08-III-1-1); F, Morrosaurus antarcticus (MACN-Pv 197); G, Isasicursor santacrucensis (MPM 21,533); H, Anabisetia saldiviai (MCF-PVPH 74). Red arrows exhibit the taxa that exhibits a distinct condylid processes between the condyles, while the blue arrows indicate the development and shifting of the condyloid (‘rectangular’) processes. Dotted lines highlight different patterns on general shape (convex or almost rectilinear) and condyle development of femoral surfaces in the posterior (1) and distal (2) views. Anatomical abbreviations: id, indentation; ieg, intracondylar extensor groove. Without scale for comparative purposes.
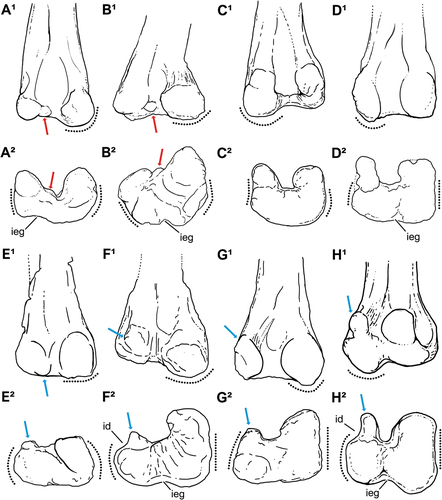
In lateral view, the strongly sigmoidal shaft of Tietasaura derbyiana differs from the condition seen in Hypsilophodon foxii Huxley Citation1870, some elasmarians (e.g. Gasparinisaura cincosaltensis Coria and Salgado Citation1996a, Talenkauen santacrucensis Novas et al. Citation2004, iguanodontians, and theropods, such as noasaurids and maniraptorans, in which the femora display relatively straighter profiles (e.g. Hulke Citation1882; Madzia et al. Citation2018; Rozadilla et al. Citation2019). However, it is remarkable that the femur shaft is slightly sigmoid in perinate specimens of Iguanodon galvensis Verdú et al. Citation2015 but acquires a fully straight profile through its ontogenetic development.
Compared with theropods, the new species lacks any sign of an anteromedial crest (Carrano and Sampson Citation2008). The fibular supracondylar crest confluent with the condyle also contrasts with what is observed in theropods, in which this crest is posteriorly projected and detached from this condyle (e.g. Carrano Citation2007; Gianechini and Apesteguia Citation2011; Egli et al. Citation2016; Rauhut and Carrano Citation2016; Novas et al. Citation2018). In paravians, excepting dromaeosaurids, the articular regions are separated by a deep groove, forming a trochlea-like articular surface in distal view (Baumel and Witmer Citation1993; Chiappe Citation1996; Norell and Makovicky Citation1997; Forster et al. Citation1998).
The mostly planar anterior surface of Tietasaura derbyiana further differs from the condition observed in most theropods, in which this region presents a deep and elongated anterior intercondylar groove (Carrano et al. Citation2012). In this respect, Tietasaura derbyiana is similar to the morphology found in thescelosaurids and ornithopods (e.g. Krumenacker Citation2017).
Tietasaura derbyiana differs from iguanodontians in lacking a medially inflated medial condyle that partially covers the posterior intercondylar groove (e.g. Madzia et al. Citation2018). It also differs from most ornithopods in lack of mediolaterally compressed distal condyles in the posterior view. As in elasmarians, the condyles in Tietasaura derbyiana are often subcircular, as seen in Kangnasaurus coetzeei Haughton Citation1915 (Cooper Citation1985), Gasparinisaura cincosaltensis, Anabisetia saldiviai, Trinisaura santamartaensis Coria et al. Citation2013, Notophysilophodon comodoriensis, and Morrosaurus antarcticus Rozadilla et al. Citation2016.
Tietasaura derbyiana is further distinguished from most neornithischians by the absence of an indentation on the posterolateral edge of the femur in distal view, between the main corpus of the fibular condyle and the condyloid (or rectangular) process, as seen in iguanodontians (e.g. Galton Citation1977; Forster Citation1990; Bertozzo et al. Citation2017; Maidment et al. Citation2022), Burianosaurus augustai Madzia et al. Citation2018, some elasmarians such as Kangnasaurus coetzeei, Anabisetia saldiviai, Trinisaura santamartaensis, and Morrosaurus antarcticus, thescelosaurids (e.g. Barta and Norell Citation2021; Sues et al. Citation2023), and, although quite gentle, basal marginocephalians (e.g. Butler and Zhao Citation2009; Han et al. Citation2018). Tietasaura derbyiana lacks this indentation (thus lacking a distinct rectangular/condyloid process) and thus exhibits a straight medial edge of the fibular condyle in distal view. In this respect, Tietasaura derbyiana is like some elasmarians, such as Fulgurotherium australe Huene von Citation1932 (e.g. Molnar and Galton Citation1986), Gasparinisaura cincosaltensis, Notohypsilophodon comodoriensis, Isasicursor santacrucensis Novas et al. Citation2019, and the Victorian femur type I (Rich and Rich Citation1989). Within elasmarians, Tietasaura derbyiana resembles Notohypsilophodon comodorensis in the conspicuously sharp posterior supracondylar ridges (Ibiricu et al. Citation2010), as well as the presence of a condylid between both condyles. The fibular condyle of Tietasaura derbyiana is also strikingly like that of Gasparinisaura cincosaltensis, roughly in the shape of a rectangular triangle with a slightly rounded medial edge in the distal view.
Ornithopoda Marsh Citation1881
Iguanodontia Baur Citation1891
Ankylopollexia Sereno Citation1986
Styracosterna Sereno Citation1986
Gen. et sp. indet.
Specimen A
Referred material
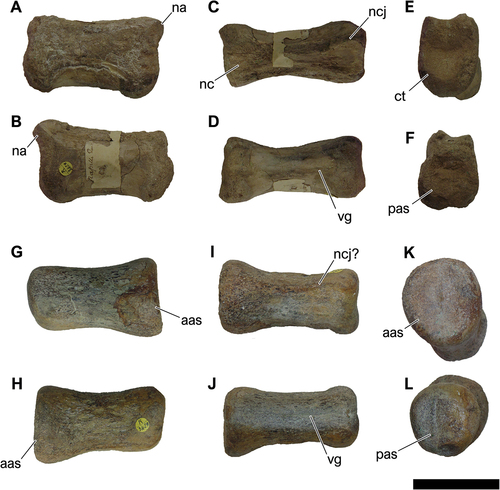
NHM-PV R.3425, represented by an isolated middle dorsal centrum (; Table S2).
Figure 6. NHM-PV R.3425, isolated middle dorsal centrum with iguanodontian affinities (Specimen A) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at Plataforma Station (Locality 3). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: avh, anteroventral heel; ct, cotyle; nvf, neurovascular foramina; mco, median concavity; ncj, neurocentral joint; pas, posterior articulation surface; pvg, posteroventral groove. Scale bar = 100 mm.

Locality and horizon
In contrast to the other materials studied here, the collection label of NHM-PV R.3425 only contains the description ‘Cretaceous. Bahia, Brazil’. in the provenance data, hindering major details about the exact locality that the specimen came. Nonetheless, according to the site descriptions provided by Mawson and Woodward (Citation1907), we can safely ascribe this specimen to the Ilhas Group (Valanginian – Barremian). The specimen NHM-PV R.3425 might have been recovered from some outcrop at the beach near the Plataforma Station (Locality 3; see ), based on taphonomical features of the specimen, such as the colour and the abrasion degree. The year in which this specimen was collected (1906) also supports this assumption, wherein only three other similar materials were reported from this locality.
Description and comparisons
NHM-PV R.3425 is referred to a middle dorsal centrum given the absence of articular facets of chevrons. The centrum is dorsoventrally taller than anteroposteriorly long, as in some middle dorsal centra of Iguanodon bernissartensis Boulenger Citation1881, Mantellisaurus atherfieldensis (Hooley Citation1925), Hippodraco scutodens, Iguanacolossus fortis McDonald et al. Citation2010, and Morelladon beltrani Gasulla et al. Citation2015 (see Norman Citation1980, Citation1986; McDonald et al. Citation2012). Both anterodorsal and anteroventral corners of the anterior articular surface surpass the posteroventral one (). In addition, the anteroventral expansion creates a rounded anteroventral heel that is slightly deflected anteriorly. The centrum exhibits an opisthoplatyan intercentral articulation, bearing a gently concave anterior surface and an almost flat posterior surface that exhibits ellipsoid outlines (). Signals of lateral striations circumscribing the articular surfaces are absent but could correspond to a taphonomic artefact since this facet is well abraded. In the lateral view, the posterior articular surface displays subparallel margins, which slightly diverges dorsally. The lateral surfaces of NHM-PV R.3425 exhibit a series of small neurovascular foramina, mainly on the right surface. A marked neurocentral joint is observable on the dorsal surface, indicating that NHM-PV R.3425 could correspond to an immature individual (). The ventral margin is gently concave in lateral view (). A marked posteroventral groove is present ventrally, lacking crests or ridges ().
The centrum is slightly constricted medially and has an ‘hourglass shape’ in both dorsal and ventral views (), which is the typical morphology observed in ornithischian dorsal centra (e.g. Norman et al. Citation2004a). NHM-PV R.3425 differs from thyreophorans (e.g. Raven et al. Citation2020; Breeden et al. Citation2021) and marginocephalians (e.g. Butler and Zhao Citation2009) in exhibiting taller than wide articular centra, with a posteroventral margin deeper than the anteroventral one in lateral view. These features ally NHM-PV R.3425 to some iguanodontian ornithopods, such as Iguanodon bernissartensis Boulenger Citation1881, Mantellisaurus atherfieldensis (Hooley Citation1925), Hippodraco scutodens McDonald et al. Citation2010, and Iguanacolossus fortis McDonald et al. Citation2010. Another typical ornithopod feature that can be seen in NHM-PV R.3425 corresponds to a series of small neurovascular foramina on the lateral surface of the centra, like what is seen in elasmarians (e.g. Calvo et al. Citation2007; Rozadilla et al. Citation2019), Sektensaurus sanjuanboscoi Ibiricu et al. Citation2019, and iguanodontians (e.g. Wang et al. Citation2010), including hadrosaurids (e.g. Cruzado-Caballero Citation2017).
The lack of the median ventral keel in NHM-PV R.3425 differ it from the middle dorsal centra of several ornithischians such as Hypsilophodon foxii (see Hulke Citation1882), Psittacosaurus mongoliensis (Osborn Citation1923) (see Sereno and Chao Citation1988; Averianov et al. Citation2006), thescelosaurids (including ‘jeholosaurids’, with exception of Orodromeus), such as Changchunsaurus parvus Zan et al. Citation2005, Jeholosaurus shangyuanensis Xu et al. Citation2000, Yueosaurus tiantaiensis Zheng et al. Citation2012 (Scheetz Citation1999; Butler et al. Citation2011; Han et al. Citation2012). This further differs from some elasmarians, such as Mahuidacursor lipanglef Cruzado-Caballero et al. Citation2019 and Macrogryphosaurus gondwanicus (see Rozadilla et al. Citation2019), even though this feature is also absent in other elasmarians, such as Trinisaura santamartaensis and Notohypsilophodon comodoriensis (Coria et al. Citation2013; Ibiricu et al. Citation2014).
The absence of a median ventral keel is also observed in the unnamed ornithopod from the Bajo de la Carpa Formation (MAU-Pv-LE-617; Jiménez Gomis et al. Citation2018), non-hadrosauriform dryomorphans (e.g. Tenontosaurus tilletti Ostrom Citation1970), dryosaurids, and ankylopollexians, such as Camptosaurus dispar (Marsh Citation1879), Cumnoria prestwichii (Hulke Citation1880), Hippodraco scutodens, Iguanacolossus fortis, and Dakotadon lakotaensis Weishampel and Björk Citation1989 (e.g. Gilmore Citation1909; Galton Citation1981; Forster Citation1990; McDonald et al. Citation2010). Rhabdodontomorphs, such as Zalmoxes robustus (Nopcsa Citation1900) and Mochlodon suessi (Bunzel Citation1871) (see Godefroit et al. Citation2009; Ősi et al. Citation2012), exhibit a reduction of the ventral keel, showcasing an evolutionary tendency to diminish in non-hadrosauriform iguanodontians. Hadrosauriformes demonstrate, conversely, a secondary development of a ventral keel, as evidenced in Iguanodon bernissartensis, Mantellisaurus atherfieldensis, Morelladon beltrani Gasulla et al. Citation2015, Ouranosaurus nigeriensis Taquet Citation1972, and various hadrosaurids (e.g. Hooley Citation1925; Gasca et al. Citation2015; Verdú et al. Citation2015). In this regard, NHM-PV R.3425 shares more similarities with non-hadrosauriform dryomorphans.
Notably, NHM-PV R.3425 shares with the middle dorsal centra of some basal ankylopollexians asymmetrical ventral margins in the lateral view, with a shallower anteroventral heel compared to the deeper posteroventral one. This configuration is observed in the middle dorsal centra of the non-hadrosauriform ankylopollexians Cumnoria prestwichii (Maidment et al. Citation2022), Camptosaurus dispar (Gilmore Citation1909), Uteodon aphanoecetes (Carpenter and Wilson Citation2008), and Hippodraco scutodens (McDonald et al. Citation2010). This trait is also present in Sektensaurus sanjuanboscoi, initially interpreted as an indeterminate ornithopod (either elasmarian or iguanodontian; see Ibiricu et al. Citation2019) and recognised here as a non-hadrosauriform styracosternan (see further below).
Additionally, this feature is absent in thyreophorans (e.g. Norman et al. Citation2004b; Raven et al. Citation2020; Breeden et al. Citation2021), elasmarians (e.g. Novas et al. Citation2019; Cruzado-Caballero et al. Citation2019; Rozadilla et al. Citation2019, 2020), the indeterminate ornithopod MAU-Pv-LE-617 (Jiménez Gomis et al. Citation2018), dryosaurids (Galton Citation1981; Barrett Citation2016), Tenontosaurus tilleti Ostrom Citation1970, rhabdodontids (Godefroit et al. Citation2009; Ősi et al. Citation2012), Hypsilophodon foxii (Hulke Citation1882), and more advanced styracosternans, such as Iguanacolossus fortis, Lanzhousaurus magnidens You et al. Citation2005, Barilium dawsoni (Lydekker Citation1888; Norman Citation2011), Hypselospinus fittoni (Lydekker Citation1889; Norman Citation2010), Morelladon beltrani, Iguanodon bernissartenis (Norman Citation1980), Mantellisaurus atherfieldensis Hooley Citation1925 (Norman Citation1986), Ouranosaurus nigeriensis (Bertozzo et al. Citation2017), Equijubus normani You et al. Citation2003, and Probactrosaurus gobiensis (Rozhdestvensky Citation1966). Hence, NHM-PV R.3425 exhibits similarities with some basal ankylopollexians like Camptosaurus dispar, Cumnoria prestwichii, Uteodon aphanoecetes, Hippodraco scutodens, and Sektensaurus sanjuanboscoi (Gilmore Citation1909; Carpenter and Wilson Citation2008; McDonald et al. Citation2010; Ibiricu et al. Citation2019; Maidment et al. Citation2022).
NHM-PV R.3425 further differs from the dorsal centra of most ornithischians given that it bears taller than wide, ellipsoid articular outlines. In this respect, it differs from the wider than taller, or as wide as high, articular surfaces found in the dorsal centra of thyreophorans (e.g. Norman et al. Citation2004b; Raven et al. Citation2020; Breeden et al. Citation2021), marginocephalians (e.g. Butler and Zhao Citation2009), rhabdodontomorphs (Godefroit et al. Citation2009; Ősi et al. Citation2012), Tenontosaurus tilleti (Forster Citation1990), and dryosaurids (Galton Citation1981; Barrett Citation2016). The same is true for some elasmarians, such as Talenkauen santacrucensis (see Rozadilla et al. Citation2019), Mahuidacursor lipanglef (see Cruzado-Caballero et al. Citation2019), and Macrogryphosaurus gondwanicus (see Rozadilla et al. Citation2020), even though other elasmarians show similarly higher than wide dorsal centra, such as Isasicursor santacrucensis (which still differs from NHM-PV R.3425 in exhibiting a ventral groove; see Novas et al. Citation2019). Like Isasicursor santacrucensis, the dorsal centrum of the indeterminate ornithopod MAU-Pv-LE-617 also resembles NHM-PV R.3425 in being higher than wide, and distinct in bearing a ventral groove (Jiménez Gomis et al. Citation2018). This morphology also differs from what is seen in the basal ankylopollexians Camptosaurus dispar Marsh Citation1879 and Uteodon aphanoecetes (Carpenter and Wilson Citation2008) but matches with an array of some non-hadrosauroid ankylopollexians middle dorsal centra. This feature is absent in hadrosauriforms, such as Ouranosaurus nigeriensis (see Bertozzo et al. Citation2017) and Eolambia caroljonesa Kirkland Citation1998 (McDonald et al. Citation2012). NHM-PV R.3425 further resembles Hippodraco scutodens in the flushed dorsal edge of the posterior articular surface relative to the anterior articular surface.
In summary, NHM-PV R.3425 resembles the middle dorsal vertebrae of non-hadrosauriform ankylopollexians due to the following combination of characters: centrum being subequal in length and height; higher than wide, ellipsoid articular outlines; concave ventral margin in lateral view, with the posterior ventral projection deeper than the posterior one; and ventral surface lacking a pronounced keel or ridge. Based on this combination of features, we regard that NHM-PV R.3425 most closely resembles Cumnoria prestwichii, Hippodraco scutodens, and Sektensaurus sanjuanboscoi. In particular, NHM-PV R.3425 is most similar to the styracosternan Hippodraco scutodens due to the articular surfaces not being parallel in lateral view.
Specimen B
Referred material
NHM-PV R.2459, a nearly preserved middle to posterior dorsal centrum (; Table S2).
Figure 7. NHM-PV R.2459, isolated middle to posterior dorsal centrum with iguanodontian affinities (Specimen B) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at Forte Monte Serrat (Locality 2). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: avh, anteroventral heel; ct, cotyle; nvf, neurovascular foramina; na, neural arch; nc, neural canal; ncj, neurocentral joint; pas, posterior articulation surface; st, striae. Scale bar = 100 mm.

Locality and horizon
The specimen NHM-PV R.2459 was recovered from sandy shale facies at the Forte Montserrat (Locality 2), which is located close to the beaches of Pedra Furada Bay, Salvador City, Bahia State. The rock layers in this locality are associated with the Ilhas Group, based on the faciological characterisation provided by Allport (Citation1860), and correspond to the level of the Valanginian – Hauterivian Marfim Formation.
Description and comparisons
NHM-PV R.2459 is an isolated dorsal centrum and, as in several bones studied here, presents a certain degree of weathering. The centrum is taller than wider, being taller than NHM-PV R.3425, and has an opisthoplatyan intercentral articulation. The centrum exhibits well-defined longitudinal striations circumscribing the articular surfaces (), as well as presents numerous lateral neurovascular foramina as in NHM-PV R.3425. The anterior articular surface has a subcircular to oval outline (). In addition, the anterior articular face, as preserved, is slightly transversely wider than the posterior one (), defining a wedge-shape in lateral and ventral views () – a common feature among ornithopods (e.g. Norman et al. Citation2004a). The ventral margin is strongly concave in lateral view, showing an offset between the anterior and posterior faces, with the anteroventral heel projecting further ventrally than the posteroventral one, as in NHM-PV R.3425. The lateral and ventral surfaces of the centrum bear several small neurovascular foramina. The absence of a neural arch hinders refined comparisons regarding the position of this element along the dorsal series.
The specimen NHM-PV R.2459 has already been referred as a dorsal centrum of the tetanuran Megalosaurus by Owen (in Allport Citation1860) and after attributed as a probable material of the pholidosaurid Sarcosuchus hartti by Buffetaut and Taquet (Citation1977). However, NHM-PV R.2459 differs substantially from theropods due to the lack of external and internal pneumatic traits, such as pneumatopores, as usual in presacral elements for Theropoda (e.g. Benson et al. Citation2012). This specimen also differs from pholidosaurids, as well as several other crocodyliforms, given their taller centra, opisthoplatyan intercentral articulations, and the lack of hypapophysis (Souza Citation2019).
As with NHM-PV R.3425 (see above), this specimen notably resembles the morphology of non-hadrosauriform ankylopollexians, such as Cumnoria prestwichii, Hippodraco scutodens, and Sektensaurus sanjuanboscoi, due to the lack of either a ventral keel or groove, strong concave ventral margin in lateral view (with the anterior ventral projection deeper than the anterior one; ellipsoid to suboval articular surfaces, and the centrum being subequal in length and height. Like NHM-PV R.3425 and Hippodraco scutodens, NHM-PV R.2459 also exhibits articular surfaces that are not parallel in lateral view.
Specimen C
Referred material
NHM-PV R.3426, represented by a fragmentary middle caudal centrum; NHM-PV R.3428(a) and NHM-PV R.3428(b), composed of two posterior caudal centra, which might belong to the same individual. (; Table S2).
Figure 8. NHM-PV R.3426, middle caudal centrum with iguanodontian affinities (Specimen C) from the Berriasian–Barremian Salvador Formation (Massacará Group) at Mapelle Quarry (Locality 9). A, right lateral; B, left lateral; C, anterior; D, posterior; E, ventral views. Anatomical abbreviations: avh, anteroventral heel; ct, cotyle; pas, posterior articulation surface; pvh, posteroventral heel. Scale bar = 100 mm.

Figure 9. NHM-PV R.3428(a) and NHM-PV R.3428(b), associated middle to posterior caudal centra with iguanodontian affinities (specimen C) from the Berriasian–Barremian Salvador Formation (Massacará Group) at Mapelle Quarry (Locality 9). A-G, right lateral; B-H, left lateral; C-I, dorsal; D-J, ventral; E-K, anterior; F-L, posterior views. Anatomical abbreviations: aas, anterior articulation surface; ct, cotyle; na, neural arch; nc, neural canal; ncj, neurocentral joint; pas, posterior articulation surface; vg, ventral groove. Scale bar = 100 mm.
Locality and horizon
The three elements were recovered at the Mapelle Quarry (Locality 8). In this locality outcrops flagstones and conglomeratic sandstones of the Salvador Formation (Massacará Group), which is partially synchronous with the Ilhas Group, ranging from the Berriasian to Barremian time-interval.
Description and comparisons
NHM-PV R.3426 is an isolated partial opisthoplatyan centrum with unpreserved neural arch (). The centrum is approximately rectangular, with only a faint middle constriction, but does not produce an accentuated wedge shape (). The ventral margin, although damaged, seems almost flat (). It is identified here as a middle caudal centrum due to its moderate elongation, only slightly longer than tall, as seen in non-hadrosauriform styracosternan middle-to-posterior caudal centra (e.g. Norman Citation1980; McDonald et al. Citation2012). It is interesting to note that the anterior articular surface is quadrangular, like the single known anterior caudal centrum of the holotype of Iguanacolossus fortis (see McDonald et al. Citation2010). Such configuration cannot be found in the other well-known iguanodontian caudal elements, such as those of Valdosaurus canaliculatus Galton Citation1975 (Barrett Citation2016), Tenontosaurus tilletti (Forster Citation1990), Cumnoria prestwichii (Maidment et al. Citation2022), Hypselospinus fittoni (Norman Citation2011), and Ouranosaurus nigeriensis (Bertozzo et al. Citation2017). This can also not be found in other known elasmarian caudal centra (Novas et al. Citation2019; Rozadilla et al. Citation2020), nor in other indeterminate South American ornithopods (Rubilar-Rogers et al. Citation2013; Cruzado-Caballero Citation2017).
The elements NHM-PV R.3428(a) and R.3428(b) are anteroposteriorly elongated and slightly dorsoventrally taller than transversally wide. The element ‘a’ () is better preserved than the element ‘b’ (), which is very abraded, and both are apneumatic. In lateral view, the ventral margins of the centra are slightly concave (). Ventrally, the element ‘a’ also present a shallow longitudinal groove (). The intercentral articulation varies from a slightly platycoelous () to amphyplatyan ().
The specimen NHM-PV R.3426 exhibit a general quadrangular centrum shape, differing from the middle to posterior caudal centra of most ornithopods which, although the high of varying shapes, usually exhibit articular surfaces approximately subequal in width and height, as seen in elasmarians (e.g. Coria and Salgado Citation1996a; Novas et al. Citation2019; Rozadilla et al. Citation2019), rhabdodontomorphs (e.g. Weishampel et al. Citation2003; Ősi et al. Citation2012), Tenontosaurus (e.g. Forster Citation1990), dryosaurids (e.g. Barrett Citation2016), and iguanodontians such as Cumnoria prestwichii (Maidment et al. Citation2022), Camptosaurus dispar, Uteodon aphanoecetes (Carpenter and Wilson Citation2008), and Sektensaurus sanjuanboscoi (Ibiricu et al. Citation2014). By exhibiting taller than wide and quadrangular to subcircular articular surfaces, the Salvador Formation caudal centra are similar to some styracosternans such as Iguanacolossus fortis (McDonald et al. Citation2010), Iguanodon bernissartensis (Norman Citation1980), Mantellisaurus atherfieldensis (Norman Citation1986), Ouranosaurus nigeriensis (at least the middle elements; Bertozzo et al. Citation2017), and the El Castellar iguanodont (García-Cobeña et al. Citation2023). In this respect, it also resembles the indeterminate ornithopod from Bajo de la Carpa Formation (Cruzado-Caballero Citation2017).
However, the Salvador Formation distal caudal centra lack the hexagonal shape seen in the articular surfaces found in many ankylopollexians, such as Cumnoria prestwichii, Hypselospinus fittoni, Iguanodon bernissartensis, Mantellisaurus atherfieldensis, and Ouranosaurus nigeriensis. It also differs from the middle and distal caudal centra of elasmarians, such as Gasparinisaura cincosaltensis, Talenkauen santacrucensis, and Isasicursor santacrucensis, in which the articular surfaces are circular and wider than higher (Coria and Salgado, Citation1996a; Rozadilla et al. Citation2019; Novas et al. Citation2019). Instead, the articular surfaces of the distal caudal centra of the Salvador Formation form are closer to the higher than wide centra with subcircular shape seen in the basal styracosternan Iguanacolossus fortis, the El Castellar iguanodontian (García-Cobeña et al. Citation2023), and the Bajo de la Carpa Formation ornithopod (Cruzado-Caballero Citation2017).
Summarising, the middle caudal NHM-PV R.3426 differs from most ornithopods, except for Iguanacolossus fortis, in exhibiting a quadrangular articular surface (McDonald et al. Citation2010). The distal centra in NHM-PV R.3428 exhibits a combination of higher than wide articular surfaces, an opisthoplatyan condition, mild concave ventral margin in lateral view, presence of a lateral horizontal ridge, and presence of a slightly ventral groove. This synapomorphic combination of characters matches with the morphology seen in the posterior caudal centra of other styracosternans, such as Iguanacolossus fortis and the El Castellar iguanodontian, and resembles the Bajo de la Carpa Formation ornithopod as well (Cruzado-Caballero and Powell Citation2017). This is consistent with the presence of non-hadrosauriform styracosternans in the Recôncavo Basin assemblage as already indicated by the dorsal centra NHM-PV R.3425 and NHM-PV R.2459.
Specimen D
Referred material
NHM-PV R.3429, a middle caudal vertebra (; Table S2).
Figure 10. NHM-PV R.3429, isolated middle caudal vertebra with iguanodontian affinities (Specimen D) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at Plataforma Station (Locality 3). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: alf, anterolateral fossa; avh, anteroventral heel; ct, cotyle; ns, neural spine; pas, posterior articulation surface; prz, prezygapophysis. Scale bar = 100 mm.

Locality and horizons
The NHM-PV R.3429 specimen was found at the beach near Plataforma Station (Locality 3). The shale facies outcropping in this locality are associated with the Valanginian – Hauterivian Marfim Formation (Ilhas Group, Recôncavo Basin).
Description and comparisons
NHM-PV R.3429 is an incomplete vertebra, comprised by most of the centrum and part of the pedicel of the neural arch (). It is interpreted as a caudal due to the lack of a transverse process, and as a middle caudal due to its moderate elongation (about as wide as tall) and the lack of articular facets for chevrons. The intercentral articulation is platycoelous with subcircular, higher than wide articular surfaces (). The vertebra is apneumatic, and the neural arch covers entire the dorsal extension of the centrum (). The ventral surface is smooth, lacking keels or grooves (). In these respects, this element closely resembles for the middle caudal vertebrae of several ankylopollexians, such as Cumnoria prestwichii, Hypselospinus fittoni, Iguanodon bernissartensis, and Ouranosaurus nigeriensis. It differs from early-diverging iguanodontians, such as rhabdodontomorphs (Ősi et al. Citation2012) and dryosaurids (e.g. Galton Citation1981), which tend to exhibit mid caudal articular surfaces about as wide as high. It also differs from theropods and sauropods for being taller than wide, while in those saurischians the reverse is more common (Weishampel et al. Citation2003).
NHM-PV R.3429 differs from crocodylomorphs, such as notosuchians and other mesoeucrocodylians, due to the ventral offset presented and the absence of hypapophyses (e.g. Leardi et al. Citation2015). It is notorious that, in general pattern, this specimen closely matches with the other Marfim Formation iguanodontian material (NHM-PV R.2459 and R.3425), except for the lack of an asymmetrical ventral margin of the centrum in lateral view (). Given the morphology of this specimen is consistent with ankylopollexians and closely matches the general morphology, we regard that these specimens possibly represent the same taxon.
Saurischia Seeley Citation1888
Sauropoda Marsh Citation1878
Neosauropoda Bonaparte Citation1986
Diplodocoidea Marsh Citation1884
Diplodocidae Marsh Citation1884
Gen. et sp. indet.
Specimen A
Referred material
NHM-PV R.2981(b), a posterior end of a distal middle or proximal posterior caudal centrum, with a minor portion of the neural arch (; Table S2).
Figure 11. NHM-PV R.2981(b), fragment of a distal middle or proximal posterior caudal centrum with diplodocid affinities (Specimen A) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at beach between Plataforma and Itacaranha (Locality 4). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: bf, blind fossa; na, neural arch; pdc, posterodorsal concavity; pas, posterior articulation surface; pdc, posterodorsal concavity; vh, ventral hollow. Scale bar = 100 mm.

Locality and horizon
NHM-PV R.2981(b) comes from beach outcrops between Plataforma Station and Itacaranha (Locality 4), Pedra Furada Bay, and was recovered in the fossiliferous shale intercalated with sandstone lenses associated, here associated with the Valanginian – Hauterivian Marfim Formation.
Description and comparisons
NHM-PV R.2981(b) is represented by an incomplete centrum lacking most of its anterior portion. However, based on the posterior surface suggests that it might be platycoelous (). Part of the periosteum suffered some abrasion produced by sedimentary particles against the bone and possibly due to some degree of reworking of the carcass before the final burial (e.g. Shipman Citation1981). The specimen NHM-PV R.2981(b) exhibits strongly concave lateral surfaces (), as in the rebbachisaurids Amanzosaurus maranhensis Carvalho et al. Citation2003, Limaysaurus tessonei Calvo and Salgado Citation1995, and Itapeuasaurus cajapioensis Lindoso et al. Citation2019. In the anterior view, the centrum is strongly compressed medially and forms an 8-shaped outline, most by the development of deep blind fossae (). Nonetheless, any sign of internal pneumaticity on bone tissue is observable. In addition, there is no signal of an anteroposteriorly oriented ridge in NHM-PV R.2981(b), differing from the observed in UNPSJB-PV 1004/3, Dicraeosaurus hansemanni Janensch Citation1914 and Suuwassea emiliae Harris and Dodson Citation2004 (Harris Citation2006; Ibiricu et al. Citation2012). In the dorsal view (), a shallow but relatively broad posterodorsal concavity is present, which is not seen in other diplodocoids (e.g. Tschopp et al. Citation2015). Ventrally, NHM-PV R.2981(b) bears a markedly well-developed hollow that extends longitudinally (), as in NHM-PV R.3427 (see below), UNPSJB-PV 1004/3, and Tornieria africana Fraas 1908 (Remes Citation2006).
In the posterior view (), the centrum is dorsoventrally elongated like other diplodocids (see ), such as Apatosaurus ajax Marsh Citation1877, Brontosaurus parvus Peterson and Gilmore Citation1902, and Galeamopus hayi (Holland Citation1924), but less than middle to posterior caudal vertebrae of the rebbachisaurid Demandasaurus darwini Fernández-Baldor et al. Citation2011. This condition contrasts with the dorsoventrally compressed caudal centra of Amazonsaurus maranhensis and other rebbachisaurids, which are more similar to those observed in flagellicaudatans. The posterior articular face in NHM-PV R.2981(b) exhibits a roughly circular outline with flat ventral margin (), like Barosaurus lentus Marsh Citation1890 and Suuwassea emiliae, differing from the trapezoidal outlines seen in Amazonsaurus maranhensis, the unnamed rebbachisaurids from Cajual island (UFMA 1.10.015, UFMA 1.10.168, UFMA 1.10.188, and UFMA 1.10.806; see Medeiros and Schultz Citation2004), and Limaysaurus tessonei Calvo and Salgado Citation1995 (Salgado et al. Citation2004). In addition, NHM-PV R.2981(b) displays prominent dorsolateral expansions, but less developed than in the titanosaurian Mnyamawamtuka moyowamkia Gorscak et al. Citation2019.
Table 2. Caudal centrum ratios (width-to-height, length-to-height) for selected sauropod taxa (Modified from Tschopp et al. Citation2015: table S39).
Specimen B
Referred material
NHM-PV R.3427, represented by a posterior caudal centrum (; Table S2).
Figure 12. NHM-PV R.3427, isolated posterior caudal centrum with diplodocid affinities (Specimen B) from the Berriasian–Barremian Salvador Formation (Massacará Group) at Mapelle Quarry (Locality 9). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: aas, anterior articular surface; bf, blind fossa; na, neural arch; ncj, neurocentral joint; pdc, posterodorsal concavity; pas, posterior articulation surface; pdc, posterodorsal concavity; rm, rock matrix; vlr, ventrolateral ridge; vh, ventral hollow. Scale bar = 50 mm.

Locality and horizon
NHM-PV R.3427 was found at the Mapelle Quarry (Locality 9). In this locality outcrops flagstones and conglomerate sandstones of the Salvador Formation (Massacará Group), that ranges from the Berriasian to Barremian time-interval.
Description and comparisons
NHM-PV R.3427 is a moderately preserved posterior caudal centrum, showing a platycoelous type intercentral articulation and strongly dorsoventral compression and elongation (), such as the posterior caudal vertebra of Diplodocus carnegii Hatcher Citation1901 and Barosaurus lentus (). A minor part of an anteriorly displaced neural arch is also observable (), and both anterior and posterior articulations have an ellipsoid outline (), like Barosaurus lentus and Tornieria africana.
The lateral surfaces of the centrum are flat. In addition, restricted blind fossae are present, which are asymmetrically developed (). Dorsally, the neural canal appears wide, occupying almost entirely the dorsal surface of the centrum (). As in NHM-PV R.2981(b), the dorsal surface also exhibits a broad posterodorsal concavity. However, the concavity in NHM-PV R.3427 is relatively shallower than in the NHM-PV R.2981(b), due to its more posterior position in the caudal series than in the latter specimen. In the ventral view (), two well-marked ventrolateral ridges are present and extend until the distal end of the centrum. The ventrolateral ridges in NHM-PV R.3427 are sharper than the present in NHM-PV R.2981(b) () and increase in width towards the lateral edges of the centrum, being more pronounced anteriorly. The development of these ridges delimits a deep and elongated longitudinal ventral hollow, as in Tornieria africana (Remes Citation2006), being straight in shape through most of its extension. Lastly, NHM-PV R.3427 differs from NHM-PV R.2981(b) due to the absence of prominent dorsolateral expansions in the posterior portion of the centrum. This absence of dorsolateral expansions is also observed in Barosaurus lentus, especially in the middle to posterior caudal vertebrae (e.g. Lull Citation1919). However, this corresponds to another feature that could represent a change along the individual caudal series.
Neosauropoda Bonaparte Citation1986
Macronaria Wilson and Sereno Citation1998
Titanosauriformes Salgado et al. Citation1997
Somphospondyli Wilson and Sereno Citation1998
Titanosauria Bonaparte and Coria Citation1993
Lithostrotia Upchurch et al. Citation2004
Gen. et sp. indet.
Referred material
NHM-PV R.2129(a), represented by a complete anterior to mid-caudal centrum (; Table S2).
Figure 13. NHM-PV R.2129(a), isolated middle caudal centrum with lithostrotian affinities from the Hauterivian–Barremian Pojuca Formation at Itacaranha (Locality 5). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: cd, condyle; ce, condylar edge; cf, chevron facet; ct, cotyle; cvj, costovertebral joint; lco, lateral concavity; nc, neural canal; ncj, neurocentral joint; vh, ventral hollow; vlr, ventrolateral ridge. Scale bar = 100 mm.

Locality and horizon
The specimen NHM-PV R.2129(a) was found at the beach outcrops near Itacaranha (Locality 5), associated with other vertebrate elements. These specimens come from the gravelly shale facies on yellowish mudstones that could pertain to the Hauterivian – Barremian Pojuca Formation.
Description and comparisons
NHM-PV R.2129(a) is a well-preserved and relatively small vertebral centrum, entirely lacking the neural arch. Based on NHM collection labels, NHM-PV R.2129(a) has been referred to the dyrosaurid Hyposaurus, given the procoelous nature of neosuchian vertebrae (e.g. Romer Citation1956). However, most Hyposaurinae taxa display platycoelous to amphyplatyan centra (e.g. Schwarz-Wings et al. Citation2009), contrasting with strong procoelous morphology seen in NHM-PV R.2129(a). The neural arch restricted on the anterior portion of the centrum () denotes an unambiguous titanosauriform synapomorphy (e.g. Salgado et al. Citation1997; Wilson Citation2002; D’Emic Citation2012; Mannion et al. Citation2013). In contrast, the neural arch in neosuchians occupies entirely the dorsal surface of the centrum (e.g. Salisbury and Frey Citation2001).
The presence of a well-marked serrated neurocentral and costovertebral joints (), and the diminutive size of the material as well (ca. 10 cm in length), leads us to believe that the specimen represents an immature individual (Griffin et al. Citation2021). The presence of marked scars of the M. caudofemoralis or M. ischiocaudalis on both dorsolateral edges of the centrum () suggest that NHM-PV R.2129(a) might be a distal anterior or a proximal middle caudal element (C10?), especially when compare with more complete caudal series known (e.g. Kellner et al. Citation2005; Lacovara et al. Citation2014). Additionally, the anterior margin of the neurocentral suture is undifferentiated from the dorsal edge of the cotyle, strongly linked to the advanced titanosaur clade Aeolosaurini (Franco-Rosas et al. Citation2004; Santucci and Arruda-Campos Citation2011; Bandeira et al. Citation2019).
The centrum has a developed procoelous type of intercentral articulation, in which the condyle apex is flushed above the longitudinal axis of the centrum, as in advanced titanosaurians (Powell Citation2003; Poropat et al. Citation2021). This condition differs from that observed in mamenchisaurids, turiasaurians, dicraeosaurids, and diplodocids, in which the procoely is limited to the anteriormost elements and its convexity apex is there below the longitudinal axis of the centrum (Mannion et al. Citation2019). The presence of procoelous middle caudal vertebrae is observed on several Upper Cretaceous titanosaur taxa, such as Baurutitan britoi and Dreadnoughtus schrani. In contrast, titanosaurs from the Lower Cretaceous exhibit a wide array of intercentral articulation types, varying from platycoelous, amphyplatyan, and mildly procoelous middle caudal vertebrae, as in Malawisaurus dixeyi Haughton Citation1928 (Gomani Citation2005) and Mnyamawamtuka moyowamkia (Gorscak et al. Citation2019). NHM-PV R.2129(a) exhibits strongly procoelous middle caudal centra as in the contemporaneous (Valanginian – Hauterivian) taxa Tengrisaurus starkovi Averianov and Skutschas Citation2017 (see also Averianov et al. Citation2021, Citation2023), Volgatitan simbirskiensis Averianov and Efimov Citation2018, as well as ‘Iuticosaurus valdensis’ (NHM-PV R. 146 and R.151) and the unnamed titanosaur NHM-PV R.5333 from Wessex Formation (Le Loeuff Citation1993; Upchurch et al. Citation2011).
In the anterior view (), the cotyle has a subcircular outline that slightly tapers ventrally. The ventral margin in this region is almost straight, a common feature of anterior to middle caudal vertebrae due to the attachment area decreasing for the M. caudofemoralis longus along the tail (Salgado and García, Citation2002). Laterally, the centrum of NHM-PV R.2129(a) is markedly concave, as in Maxakalisaurus topai Kellner et al. Citation2006 and aeolosaurines. The cotyle margin is slightly inclined anteriorly, but not as strongly as in Aeolosaurus rionegrinus Powell Citation1987, Gondwanatitan faustoi Kellner and Azevedo Citation1999, and Uberabatitan ribeiroi Salgado and Carvalho Citation2008 (Bandeira et al. Citation2019; Silva et al. Citation2019). The ventral face of the centrum bears prominent ventrolateral ridges (), likewise many titanosaurian taxa, such as Andesaurus delgadoi Calvo and Bonaparte Citation1991, Hamititan xinjiangensis Wang et al. Citation2021, and Saltasaurus loricatus Bonaparte and Powell Citation1980 (Powell Citation1992). These ridges border a shallow longitudinal hollow, as in the holotypic caudal vertebra of ‘Titanosaurus indicus’ (GSI-2191.2194; Mohabey et al. Citation2013), Isisaurus colberti Jain and Bandyopadhyay Citation1997 (Wilson and Upchurch Citation2003) and saltasaurines. However, NHM-PV R.2129(a) differs from the latter due this fossa is not well excavated, as in Saltasaurus loricatus and Rocasaurus muniozi Salgado and Azpilicueta Citation2000 (Sanz et al. Citation1999; Powell Citation2003; Zurriaguz and Cerda Citation2017). Similarly, to GSI-2191.2194, the NHM-PV R.2129(a) specimen has two chevron facets, present on both anterior and posterior portions of the centrum, but less developed than in the Indian taxon (see Wilson and Upchurch Citation2003: ). In addition, NHM-PV R.2129(a) lacks a marked mediolaterally compression, which gives a houglass-like shape to the centrum, as present in GSI-2191.2194. The articular surfaces for chevrons in the anterior face of NHM-PV R.2129(a) are oriented anteroventrally, while on the posterior one, the facets are ventrally oriented. Although not preserved in this specimen, this pattern indicates that chevrons of the NHM-PV R.2129(a) specimen had double articular facets, as in aeolosaurines (Santucci and Arruda-Campos Citation2011). Posteriorly, the developed condyle is featureless, lacking characteristic notches or sulcus ().
Saurischia Seeley Citation1888
Theropoda Marsh Citation1881
Averostra Paul Citation2002
Gen. et sp. indet.
Referred material
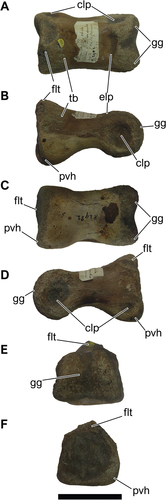
NHM-PV R.2982, a right pedal phalanx IV-1 (; Table S3).
Figure 14. NHM-PV R.2982, right pedal phalanx IV-1 of an indeterminate averostran from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at beach between Plataforma and Itacaranha (Locality 4). A, dorsal; B, right lateral; C, ventral; D, left lateral; E, anterior; F, posterior views. Anatomical abbreviations: clp, colateral pits; elp, extensor ligament pit; flt, flexor ligament tubercle; gg, ginglymus; pvh, posteroventral heel; tb, tubercle. Scale bar = 50 mm.
Locality and horizon
The specimen have been found in the beach outcrops between Plataforma Station and Itacaranha (Locality 4), Pedra Furada Bay, and recovered from fossiliferous shale intercalated with sandstone lenses. These facies are associated with the Valanginian – Hauterivian Marfim Formation.
Description and comparisons
The pedal phalanx NHM-PV R.2982 is relatively well preserved and presents some erosion on its surface, along the articular face of the proximal region. The specimen is dorsoventrally compressed in its midpoint (), acquiring a subtriangular shape on the distal portion (). It is elongated, robust, and rectangular in both dorsal and ventral views (). The phalanx is asymmetrical, maintaining a misalignment of its margins (). The distal articulation is ginglymoid (), while the proximal one is rounded and has a deep and wide collateral ligament pit. Robust and dorsoventrally compressed pedal phalanx are remarkable among abelisauroids and basal tetanurans (Coria et al. Citation2002; Novas et al. Citation2004; Carrano Citation2007; Brusatte et al. Citation2009; White et al. Citation2013). It contrasts with the slender, dorsoventrally taller, and elongated phalanx of noasaurids, some basal coelurosaurians, ornithomimosaurians, and maniraptorans (Novas Citation1997; Choiniere et al. Citation2010, Citation2012, Citation2014; Egli et al. Citation2016; Langer et al. Citation2019). Laterally, NHM-PV R.2982 has two heels proximally: a proximomedial (), which is more expanded and rounded distally; and a proximolateral () that is slightly taller and triangular. In lateral view, the specimen has a rectangular outline rather than a sharp triangular one (), as observed in Majungasaurus crenatissimus (Depéret Citation1896), Sinraptor dongi Currie and Zhao Citation1994, Allosaurus fragilis (Marsh Citation1877), and the carcharodontosaurid SHN.039 (Malafaia et al. Citation2019).
The flexor tubercle, in the lateral view, is aligned with the bottom of the proximal articulation. The dorsal surface of the distal articulation is marked by a shallow extensor groove, as in most theropods (). On the medial surface, an ellipsoid collateral pit marks the proximal portion, close to the articulation and above the posteroventral heel. The lateral collateral pit is slightly wider than the medial one. The ventral surface is almost flat and has a rectangular aspect with a small concavity in the proximal direction. Medially, the wider collateral pit and tall and laterally inclined ginglymus indicate that the NHM-PV R.2982 is a right pedal phalanx. The concave proximal articulation lacking a medial keel, the deepness of the extensor groove and the presence of a posteroventral heel indicates that the specimen is a phalanx IV-1 (e.g. Currie and Zhao Citation1993; Carrano Citation2007; Malafaia et al. Citation2019). The lack of comparative specimens with completely preserved pedes, as well as diagnostic features among postcranial skeleton of several theropod lineages, only allows an assignment to a large-bodied averostran theropod. NHM-PV R.2982 was recovered with other theropod remains (see below), suggesting that the specimen may belong to the same individual of NHM-PV R. 2980 or NHM-PV R.2981(a).
Averostra Paul Citation2002
Tetanurae Gauthier Citation1986
Orionides Carrano et al. Citation2012
Megalosauroidea Fitzinger Citation1843
aff. Spinosauridae Stromer Citation1915
Gen. et sp. indet.
Referred material
NHM-PV R.2980, one complete and well-preserved anterior caudal centrum (; Table S2). The theropod phalanx NHM-PV R.2982 was recovered from the same locality. However, the lack of additional data regarding the collection of the specimens hampers a secure association between both specimens as pertaining to the same individual.
Figure 15. NHM-PV R.2980, anterior caudal centrum with putative spinosaurid affinities from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at beach between Plataforma and Itacaranha (Locality 4). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: as, anterior articular surface; bf, blind fossa; cvj, costovertebral joint; nc, neural canal; ncj, neurocentral joint; pas, posterior articulation surface; pvh, posteroventral heel; vh, ventral hollow; vlr, ventrolateral ridge. Scale bar = 100 mm.

Locality and horizon
The specimen was found in Locality 4, the beach outcrops between Plataforma Station and Itacaranha, Pedra Furada Bay, recovered from fossiliferous shale intercalated with sandstone lenses. These facies are associated with the Valanginian – Hauterivian Marfim Formation.
Description and comparisons
The vertebral centrum NHM-PV R.2980 is rectangular in both lateral, dorsal, and ventral views (), being subequal in elongation and height. The intercentral articulation is amphicoelous (), as in megalosauroids and abelisaurids, such as Pycnonemosaurus nevesi Kellner and Campos Citation2002 (Benson Citation2010; Mateus et al. Citation2011; Delcourt Citation2017; Malafaia et al. Citation2017, 2020). This feature contrasts with other ceratosaurians and avetheropods, in which the caudal centrum is constricted towards the midline, conferring a spool-like shape (e.g. Bonaparte Citation1991; Brochu Citation2003; Benson Citation2008; Persons and Currie Citation2011). NHM-PV R.2980 presents the posterior articulation dorsoventrally taller than the anterior articulation area (). NHM-PV R.2980 has a greater ventral extension in the lateral view (), presenting a lateral offset. It also differs from the unnamed spinosaurid from the Araripe Basin (MN 4743-V; Bittencourt and Kellner Citation2004) and Ichthyovenator laosensis Allain et al. Citation2012, which presents a parallel dorsoventral extension of articular areas. The dorsal region bears a well-marked neurocentral and costovertebral joints, with several crenulations, suggesting that the centrum and neural arch were unfused at the time of death as in other archosaurians (Brochu Citation1996) and other specimens described here.
In dorsal view (), the midpoint of the centrum is constricted, forming an ‘X’ aspect due to the unfused region of the neural arch pedicel and transverse processes. The lateral surfaces have broad and shallow pleurocentral depressions (), which is common among theropods. A small blind fossa is present on the anteroventral portion on the left surface but absent on the right one. The ventral surface of the centrum has a longitudinal hollow, bounded by slightly sharp ventrolateral ridges (). Close to the posterior articulation, the ventral surface bears a pair of chevron facets, which characterises this vertebra as a caudal element.
The shallow and wide ventral hollow bounded by ridges is a remarkable feature among Megalosauroidea, in particular spinosaurids (Benson Citation2010; Mateus et al. Citation2011; Rauhut et al. Citation2016; Malafaia et al. Citation2019). Among Abelisauroidea, some taxa present the ventral surface with a deep but narrow groove running along the midline, such as Masiakasaurus knopfleri Sampson et al. Citation2001 (Carrano et al. Citation2002), Eoabelisaurus mefi Pol and Rauhut Citation2012, Aucasaurus garridoi Coria et al. Citation2002, Carnotaurus sastrei Bonaparte Citation1985, Ilokelesia aguadagrandensis Coria and Salgado Citation2000 (sensu Delcourt Citation2017), and Viavenator exxoni Filippi et al. Citation2016. Compared to MN 4743-V, the specimen NHM-PV R.2980 differs in the presence of a shallower hollow and fainter paired keels more restricted to the posterior portion of the centrum. In ceratosaurians and tetanurans, the ventral surface bears no structure such as narrow grooves nor a deep ventral hollow (Méndez Citation2014). The shape of the centrum resembles the Barremian species from Europe than to the Aptian – Albian specimen from Brazil MN 4743-V, such as Vallibonavenatrix cani from Spain (Malafaia et al. Citation2019), Baryonyx walkeri Charig and Milner Citation1986, and Iberospinus natarioi Mateus and Estraviz-López Citation2022, recovered, respectively, in England and Portugal (Mateus et al. Citation2011). However, given the absence of a preserved neural arch, we cannot assign it to Baryonychinae or Spinosaurinae.
Orionides Carrano et al. Citation2012
Avetheropoda Paul Citation1988
Allosauroidea Marsh Citation1878
aff. Carcharodontosauria Benson, Carrano and Brusatte, Citation2010
Gen. et sp. Indet.
Specimen A
Referred material
NHM-PV R.2981(a), posterior half of the dorsal centrum (; Table S2). The theropod phalanx NHM-PV R.2982 was recovered from the same locality. However, the lack of additional data regarding the collection of the specimens hampers a secure association between both specimens as pertaining to the same individual.
Figure 16. NHM-PV R.2981(a), isolated partial dorsal centrum with putative carcharodontosaurian affinities (Specimen A) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at beach between Plataforma and Itacaranha (Locality 4). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: aas, anterior articular surface; aae, articulation expasion; bf, blind fossa; nc, neural canal; ncj, neurocentral joint; ipt, internal pneumatic tissue. Scale bar = 100 mm.

Locality and horizon
The specimen NHM-PV R.2981(a) comes from Locality 4, the beach between Plataforma and Itacaranha, Pedra Furada Bay, and was recovered from sandy shale facies intercalated with sandstones, corresponding to the Valanginian-Hauterivian Marfim Formation.
Description and comparisons
NHM-PV R.2981(a) is an incomplete centrum, presenting a ventral expansion of the posterior articulation which is prominent even in lateral view and lacking pneumatic foramina (). The eroded posteriormost region has some rounded internal chambers with thick septa, but lacking sign of ramifications (). Although its thickness, the pattern of chambers organisation is quite like the polycamerate vertebrae in avetheropods (Benson et al. Citation2012). The lateral surface exhibits a shallow pleurocentral depression, presenting an eroded foramen, which is not preserved on the right side ().
Dorsally, the centrum exhibits a pronounced constriction (), as the posterior dorsal vertebrae of Allosaurus fragilis and the carcharodontosaurids Concavenator corcovatus Ortega et al. Citation2010, Lajasvenator ascheriae Coria et al. Citation2020 and Siamraptor suwati Chokchaloemwong et al. Citation2019 (see also Apesteguía et al. Citation2016; Cuesta et al. Citation2019). Ventrally, NHM-PV R.2981(a) lacks chevron articular surfaces, as well as grooves or ventral keels (), as typical in the caudal series of theropods. The posterior face is eroded, but the anterior intercentral articular surface seems to be flat (). Those features, together with the absence of the parapophysis, indicate that NHM-PV R.2981(a) is probably a posterior dorsal vertebra. The specimen lacks diagnostic features and, albeit the hourglass shape also occurs in some basal coelurosaurs, it is mainly present among allosauroids, especially carcharodontosaurians (Carrano et al. Citation2012).
Specimen B
Referred material
NHM-PV R.2129(b), a partial mid-caudal centrum lacking most of its dorsal portion (; Table S2).
Figure 17. NHM-PV R.2129(b), isolated partial middle caudal centrum with putative carcharodontosaurian affinities (Specimen B) from the Valanginian–Hauterivian Marfim Formation (Ilhas Group) at Itacaranha (Locality 5). A, right lateral; B, left lateral; C, dorsal; D, ventral; E, anterior; F, posterior views. Anatomical abbreviations: aas, anterior articular surface; aae, articulation expasion; mc, medial constriction; pae, posterior articular edge. Scale bar = 100 mm.
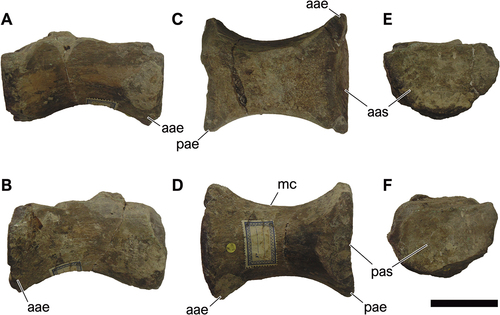
Locality and horizon
The specimen NHM-PV R.2129(b) was found near Locality 5, the Itacaranha site, recovered from sandy shale facies intercalated with sandstones corresponding to the Valanginian-Hauterivian Marfim Formation.
Description and comparisons
NHM-PV R.2129(b) is a slightly amphicoelous element, interpreted here as a probable mid-caudal vertebra. The centrum is anteroposteriorly elongated (), being strongly constricted in its median portion. This constriction confers a hourglass-shape in dorsal and ventral views, marking an accentuated ventral concavity (). The centrum is anteroposteriorly short compared to the carcharodontosaurids from the Potiguar Basin (UFRJ 523-R and 524-R; Pereira et al. Citation2020), which could be a variation along the axial sequence. NHM-PV R.2129(b) shows slight platycoelous intercentral articulations and, although the centrum is crushed, it is more developed transversely than dorsoventrally (). Hourglass-shaped centrum is synapomorphic to allosauroids (Carrano et al. Citation2012; Bandeira et al. Citation2021) and some basal coelurosaurs, especially megaraptorans (Brochu Citation2003; Carpenter et al. Citation2005; Sereno et al. Citation2008; Choiniere et al. Citation2010; Samathi et al. Citation2019). The shape of the centrum is remarkable in both groups, however, NHM-PV R.2129(b) lacks diagnostic features to refer either to Carcharodontosauridae or Megaraptora. However, the centrum lacks the pneumatization associated typical of the latter, suggesting a carcharodontosaurian affinity.
Additional dinosaur remains from Bahia State
Some additional specimens from the Recôncavo Basin have been briefly reported in the literature as undetermined reptiles (e.g. Gallo et al. Citation2023). Based on the collection labels, these remains were collected at Locality 9 (Mapelle quarry) in the Salvador Formation (; ). The specimens, however, were not described nor identified as dinosaur remains until now. Unfortunately, this additional material can no longer be firsthand evaluated, as they were housed in the National Museum at the time of the fire that occurred in September 2018. Hence, the status of such specimens is presumably lost, as they have not yet been located by the rescue team (Luciana Carvalho, pers. comm. 2022). There are a few photographs of these specimens (), although not all anatomical views are covered (Gallo Citation1993; see Supplementary Data). It is important to provide some observations of these materials given that they are an indicative of a hidden abundance of dinosaur material in the Recôncavo Basin, as well as suggests that Bahia State dinosaur diversity could be greater than previously expected.
Figure 18. Additional dinosaur specimens from Bahia state (modified from Gallo Citation1993; Gallo et al. Citation2023). A, partial neosauropod tooth crowns (1–3); B, probable theropod humerus with tyrannosauroid affinities in anterior view. Anatomical abbreviations: ap, apical portion; ec, epicondyle; le, laterodistal expansion; ls, labial surface; sm, sinuous margin. Scale bar = 10 mm.

Among the specimens, some sauropod teeth are figured (). All identified teeth lack a collection number and bear a general narrow morphology. The most complete crown () appears to be D-shaped in cross-section, with its apicobasal axis gently curved lingually, and exhibit a high SI value (>4.16; Table S4). The second partial crown () appears to be a straight pencil-like tooth, with cylindrical shape in cross-section, and displays a moderate SI value (2.88). Narrow crowns are present in neosauropod dinosaurs, first appearing in the Late Jurassic diplodocoids and later in the late-diverging titanosaurs by homoplasy (e.g. Sereno and Wilson Citation2005). The morphology observed in the tooth crowns from Bahia State coincides with that observed in diplodocoids and titanosaurs as well (see Chure et al. Citation2010), in which all tooth crowns bear moderate-to-high SI values. Given the presence of both clades in the Recôncavo Basin through axial material (see above), and the impossibility of a firsthand observation of these materials (thus hampering the observation of wear facets patterns), we cannot assume whether they belong to a diplodocoid or a titanosaur and regard these specimens as indeterminate neosauropods.
Regarding appendicular elements, only MN 4451/1-V was identified, and it is represented by the shaft of a long bone missing the proximal and distal regions due to breakage (). On the distal end, this bone is relatively flat, wider mediolaterally than anteroposteriorly. The shaft becomes progressively subcircular proximally. It exhibits a slight mid-length constriction in anterior view. In anterior view, the lateral side of the distal region exhibits a sigmoidal contour, with a lateral convexity immediately followed by a distal concavity. Due to this combination of features, the general anatomy of this bone closely matches to that of the humeral shaft of the tyrannosauroid Eotyrannus lengi Hutt et al. Citation2001 (see Naish and Cau Citation2022: fig. 19B). It seems that the proximal breakage occurred roughly at the level where the deltopectoral crest would be present (Naish and Cau Citation2022: fig. 19). The cross-section pattern (with a subcircular and constricted mid-shaft with flattened distal region) matches the humeri morphology of other theropods as well, such as noasaurids (e.g. Langer et al. Citation2019, De Souza et al. Citation2021). However, the sigmoidal laterodistal contour of the shaft in anterior view is like the one seen in Eotyrannus lengi. This feature is absent in noasaurids (e.g. Carrano et al. Citation2002), abelisaurids (e.g. Carrano Citation2007), and carcharodontosaurids (e.g. Canale et al. Citation2015; Cuesta et al. Citation2019). We thus regard that this specimen could possibly represents the right humerus of a tyrannosauroid theropod, due to its similarities to Eotyrannus lengi. However, some caution is needed regarding this interpretation given the fragmentary nature of the specimen. The presence of tyrannosauroids in the Berriasian – Barremian of South America would not be surprising, given the presence of the probable tyrannosauroid Santanaraptor placidus Kellner Citation1999 in the Aptian Romualdo Formation of Northeast Brazil (Delcourt and Grillo Citation2018).
Phylogenetic relationships of the Bahia State dinosaurs
Considering the implications associated with the identification of Tietasaura derbyiana and the Bahia iguanodontians as the first unambiguous ornithischian osteological remains from Brazil, we performed two analyses to examine their phylogenetic relationships. However, it is crucial to note that, given the fragmentary nature of the specimens, the obtained results do not alone elucidate the less inclusive phylogenetic connections of these specimens with other taxa. Rather, such analyses are useful for identifying shared morphological characters with distinct clades or taxa and contributes to understanding of the systematic affinities of these taxa alongside the anatomical comparisons carried. Furthermore, do they serve as evidence contributing to the recognising of palaeodiversity within the analysed region.
For the purpose of these analyses, the Bahia iguanodontian sample was divided into two operational taxonomic units (OTUs): the Marfim Formation iguanodontian (represented by the dorsal elements NHM-PV R.2459 and R.3425, which were not found associated, but are redundant regarding phylogenetic scoring) and the Salvador Formation iguanodontian (represented by the caudal elements NHM-PV R.3426, R.3428(a), and R.3428(b), which are likely associated). Additionally, we also carried a phylogenetic analysis to investigate the positioning of the lithostrotian specimen NHM-PV R.2129(a). Although the theropod (NHM-PV R.2129b R.2980, R.2981a, and R.2982) and diplodocoid (NHM-PV R.2981b, R.3427) specimens described in this work encompass diagnostic materials, the limited codifiable characters (less than 3) prevented their inclusion in the known matrices for these groups. Thus, we only provide the comparable characters of these specimens in the Supplementary Data.
Datasets and analytical protocol
To investigate the phylogenetic relationships of the ornithischian specimens, all three OTUs (Tietasaura derbyiana, the Marfim Formation iguanodont, and the Salvador Formation iguanodont) were included in an updated version of the neornithischian phylogenetic matrix of Rozadilla et al. (Citation2019), which is derived from Boyd (Citation2015). For this analysis, five unstable OTUs were pruned a posteriori (Yueosaurus tiantaiensis, Elrhazosaurus nigeriensis, Callovosaurus leedsi, Qantassaurus intrepidus, and Atlascopcosaurus loadsi). The two iguanodontian OTUs were further included in an updated version of the iguanodontian phylogenetic matrix of Verdú et al. (Citation2015), derived from McDonald et al. (Citation2012). For this analysis, four unstable OTUs were treated as pruned taxa (Muttaburrasaurus langdoni, Barilium dawsoni, Dakotadon lakotaensis, and Fukuisaurus tetoriensis). For the Lithostrotia specimen, we use the dataset of Navarro et al. (Citation2022), a modified version of Carballido et al. (Citation2020). For this analysis, six unstable OTUs were pruned a priori (Zapalasaurus bonapartei, Bellusaurus sui, Brachiosaurus altithorax, Abydosaurus mcintoshi, Chubutisaurus insignis, and Kaijutitan maui). All data matrices and modifications incorporated are described in the Supplementary Data.
We apply an equally weighted parsimony approach in all analyses performed here using the software TNT 1.5 (Goloboff and Catalano Citation2016). In both cases, to first identify the minimum length trees (MLTs), we performed tree searches using the ‘New Technology’ algorithms: Sectorial Search, Ratchet (25 substitutions made, or 99% swapping completed, 6 up-weighting probabilities, 6 down-weighting probabilities, with 10 total iterations), and Tree Fusing. Driven Search included 15 initial add sequences, random seed equal 0, 50 times of minimum length, without collapsing trees after search. Subsequently, the stored trees were subjected to a Traditional Search using TBR algorithm (by command ‘bb’) to find all most parcimonious trees (MPTs).
For the construction of strict consensus trees (SCT), zero-length branches will be collapsed in the MPTs (collapsing rule 3), if they lack support under any of the most parsimonious reconstructions. The obtained SCTs were time-scaled in the R software v4.3.1 by using the ‘ape’, ‘paleotree’, ‘phytools’, and ‘strap’ packages, following the method Minimum-Branch-Length described by Laurin (Citation2004). Node supports for the MPTs will be calculated using Bremer procedures (Decay Index; Bremer Citation1994), calculated using the TNT script ‘bremsup.run’ that combines heuristic searches of suboptimal trees allied to tree searches under negative constraints. The complete SCTs and Bremer supports were also presented in the Supplementary Data.
Results
Ornithischian material
Phylogenetic analysis 1
The first analysis (modified from Rozadilla et al. Citation2019) resulted in 14,760 minimum-length trees (911 steps), with ensemble consistency index (CI) of 0.655 and retention index (RI) of 0.360 (). In the strict consensus tree, Tietasaura derbyiana is recovered as an elasmarian ornithopod, sister-group of the Argentinian Notohypsilophodon comodoriensis based on character 261 (0→1), strong and sharp posterior supracondylar ridges of the femur). These two taxa fell in a polytomy with Gasparinisaura cincosaltensis and Fulgurotherium australe Huene 1932, as supported by character 260 (1→0), indentation of fibular condyle forming distinct condylid absent. Character 259 (0→1), distal femoral condyles rounded in posterior view, is recovered as an elasmarian synapomorphy (homoplastic with Thescelosauridae).
Figure 19. Time-scaled strict consensus trees depicting the phylogenetic relationships of the Recôncavo Basin ornithischians. A, all taxa included in an updated version of the neornithischian phylogenetic matrix of Rozadilla et al. (Citation2019), with Tietasaura derbyiana Gen. et sp. nov. found as an elasmarian, while the material of the Marfim and Salvador formations were recuperated as basal styracosternans. B, phylogenetic affinities of the Marfim and Salvador formations specimens in the phylogenetic matrix of Verdú et al. (Citation2015), which is focused on internal relationships of Iguanodontia. Bremer supports of both analyses are summarised in the supplementary data. Silhouettes from several sources.
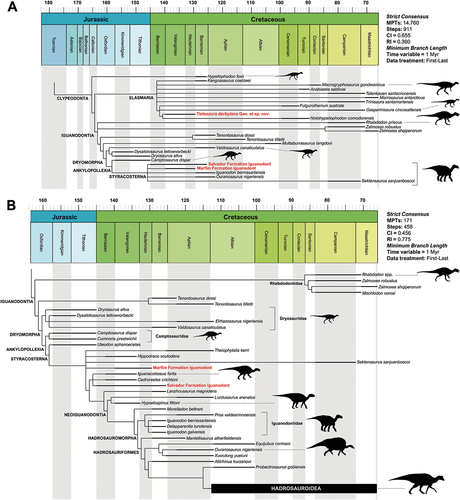
Both Marfim and Salvador formations iguanodontians are nested in a polytomic node that corresponds to Styracosterna (). This node is supported by character 262 (0→1), middle dorsal centra articular surfaces taller than wide, as a synapomorphy; and character 265 (0→1), posterior caudal centra articular surfaces taller than wide, as an ambiguous synapomorphy. Such trait is absent in Sektensaurus sanjuaboscoi and unknown in the Marfim Formation iguanodontian, but present in the Salvador Formation iguanodontian, Iguanodon bernissartensis, and Ouranosaurus nigerensis, thus supporting the Marfim Formation iguanodontian as a styracosternan. Character 263 (1→0), middle dorsal centra lacking a ventral keel, is recovered as a synapomorphy of the clade comprising Tenontosaurus spp. + Dryomorpha, supporting the Marfim Formation iguanodontian as a member of such clade (reversed in Ouranosaurus nigerensis). Character 264 (0→1), an asymmetrical ventral margin of middle dorsal centra (with a posterior ventral projection deeper than the anterior one), is recovered as a synapomorphy of Ankylopollexia (reversed in Iguanodon bernissartensis and Ouranosaurus nigerensis). This character places the Marfim Formation iguanodontian as an ankylopollexian.
Phylogenetic analysis 2
The second analysis (modified from Verdú et al. Citation2015) resulted in 171 trees, with lengths of 456 steps, with a CI of 0.456 and RI of 0.775 (). In this analysis, the Marfim Formation iguanodontian was recovered as a relatively basal styracosternan, in a node that excludes Uteodon aphanoecetes Carpenter and Wilson Citation2008 as the basalmost styracosternan (Node A, ). This node includes a polytomy involving Hippodraco scutodens, Sektensaurus sanjuanboscoi, and Theiophytalia kerri Brill and Carpenter Citation2006, as well as a branch that includes all remaining styracosternans. This latter branch includes a small clade which englobes the Salvador Formation iguanodontian, Iguanacolossus fortis, and Cedrorestes crichtoni Gilpin et al. Citation2007 (Node B, ).
Character 151 (1→0), middle dorsals lacking a ventral keel, is recovered as a synapomorphy of Dryomorpha + Tenontosaurus (later reversed in hadrosauriforms), as in the first analysis performed here. This character supports the placement of the Marfim Fm. iguanodont as a non-hadrosauriform member of such group. Character 149 (0→1), an asymmetrical ventral margin of the middle dorsal centra (with a posterior ventral projection deeper than the anterior one), is recovered here as a synapomorphy present at the base of Ankylopollexia (later reversed in node B). This character places the Marfim Formation iguanodont as an ankylopollexian. Character 150 (0→1), middle dorsal articular surfaces higher than wide, is recovered as a synapomorphy of Node A (being a homoplasy with Cumnoria prestwichii) within a sub-clade of styracosternans, later reversed within derived hadrosauroids (). This character supports the Marfim Formation iguanodontian as a styracosternan. In some trees, character 152 (0→1), non-parallel articular surfaces of the middle dorsal centra in lateral view, is recovered as an ambiguous synapomorphy joining the Marfim Formation iguanodontian and Hippodraco scutodens.
Character 153 (1→2), hexagonal-shaped posterior caudal articular surfaces, is recovered as a synapomorphy of a clade that contains Hypselospinus fittoni and Hadrosauriformes (homoplastically with Cumnoria prestwichii), excluding the Salvador Formation iguanodontian from such clade. Characters 153 (0→1), wedge-shaped posterior caudal articular surfaces, and 154 (0→1), taller than wide posterior caudal articular surfaces, are recovered as synapomorphies joining the Salvador Formation iguanodontian and Iguanacolossus fortis, within a sub-clade of non-hadrosauriform styracosternans.
Given the fact that there are no overlapping elements between the Marfim Formation and the Salvador Formation iguanodontians, and that these specimens are too fragmentary, we suggest that these results are best seen with caution. Nonetheless, it can be said that, based on the current available material and analysis, there exists evidence for the presence of at least two distinct iguanodontians in the Recôncavo Basin.
Titanosaurian material
For the specimen NHM PV R. 2129a, we found 327 MPTs of 1799 steps (CI = 0.317/RI = 0.704), showing a well-resolved topology (). The overall topology in consensus tree is quite similar with the previous iterations of this matrix, in particular Navarro et al. (Citation2022). The established most inclusive clades (Titanosauria, Lithostrotia, Eutitanosauria, Colossosauria, and Saltasauroidea) are recovered. As in previous analysis, branch support values for most macronarian nodes (Titanosauria included) received low to moderate support values (see Supplementary Data).
Our analysis recovered NHM PV R. 2129a as a rinconsaurian lithostrotian, in a harsh polytomy within Uberabatitan ribeiroi and Brasilotitan nemophagus Machado et al. Citation2013, nested in a more inclusive clade where Aeolosaurini is included. This relationship indicates NHM PV R. 2129a as one of the most ancient rinconsaurian known to date, being supported by four synapomorphies: the presence of the ventrolateral ridges on the anterior caudal vertebrae (Char. 233: 0→1); a ventral hollow in the anterior and middle caudal vertebrae (Char. 250: 0→1); anterior caudal centra with the dorsal margin of cotyle slightly inclined anteroventrally (Char. 253: 0→1); and the presence of forked chevrons with anterior and posterior projections (Char. 266: 0→1). Regarding the last feature, it could be scored due to the articular facets of chevrons in the Pojuca specimen are present in both anterior and posterior ventral surfaces. This feature is also observed in ‘Titanosaurus indicus’, indicating of the chevrons in these taxa have two articular surfaces.
Albeit the ventral hollow and the ventrolateral ridges in the centrum are widely distributed among Titanosauria, these characters are often considered as synapomorphies of Lithostrotia (Averianov and Lopatin Citation2020; Wang et al. Citation2021). The Pojuca specimen unveils a potential for discovering new titanosaurian taxa within Recôncavo Basin. Finally, the analysis further reaffirms that Rinconsauria as an exclusively South American clade.
Discussion
Systematic affinities of the dinosaurs from Bahia State
The first mention of possible dinosaur specimens in the Recôncavo Basin was made by Richard Owen in Allport (Citation1860), which consists of an incomplete dorsal vertebra (NHM-PV R.2459) collected in 1859 at ‘Fort Montserrate’ (sic). This specimen has been also referred to as Sarcosuchus hartti (Buffetaut and Taquet Citation1977; Souza et al. Citation2019), and further assigned as an indeterminate crocodyliform (e.g. Kellner and Campos Citation2000). Likewise, the specimens NHM-PV R.2129(a) and R.3424 were related to the crocodyliform Hyposaurus ‒ based on the collection labels of the NHM. However, these specimens received little attention in the literature, being never described in detail. Dinosaur occurrences from pre-Barremian deposits are rarer worldwide and considered a product of a global scarcity of Berriasian – Hauterivian-aged continental deposits (Tennant et al. Citation2017). Given the relative paucity of dinosaur occurrences in this interval, the specimens described here represent not only one of the most diverse dinosaur faunas from this time-interval, but also an important historical finding.
The specimen NHM-PV R.2459 (as well as the specimens NHM-PV R.3425, R.3426, R.3428, and R. 3429, hereafter collectively referred to as the Bahia specimens) can be set apart from crocodyliforms due to their taller than wider centra and the lack of hypapophyses (Souza Citation2019). These specimens can also not be viewed as sauropods since both have taller than wide intercentral articulation facets and the neural arches extends onto entire the dorsal surface of the centrum, a morphology not observed on neosauropods (McIntosh Citation1990). The Bahia specimens also differ from any theropod from the Cretaceous due to the lack of either fossae, pneumatic foramina or pleurocoel on the lateral surface of the centrum, which can be found in both ceratosaurs and avetheropods (e.g. Carrano et al. Citation2012). Furthermore, theropod centra usually are ventrally keeled and lack a distinctively deeper posteroventral margin, with exception of Viavenator exxoni and Unenlagia comahuensis which instead presents a ventral groove (Gianechini and Apesteguia Citation2011; Filippi et al. Citation2016). In contrast, the Bahia specimens closely resemble with the morphology seen in ornithischians, which typically exhibit hourglass-shaped centra and lack any sign of pneumatic traits (e.g. pneumatopores, pleurocoels, fossae, or a pneumatic internal bone tissue; Norman et al. Citation2004a; Wilson et al. Citation2011).
Concerning ornithischians, the Bahia specimens differ from several major clades, lacking a median ventral keel on the dorsal centra. In basal thyreophorans, the dorsal centra are also spool-shaped and lack ventral keels; but differ from the Bahia ornithischian material in bearing circular intercentral articular surfaces (Norman et al. Citation2004b). In neornithischians, however, the presence of a ventral keel in the dorsal centra is widespread, found in forms such as Hypsilophodon foxii (Hulke Citation1882), Psittacosaurus mongoliensis (Sereno and Chao Citation1988; Averianov et al. Citation2006), the thescelosaurids Changchunsaurus parvus (Zan et al. Citation2005; Butler et al. Citation2011), Jeholosaurus shangyuanensis (Xu et al. Citation2000), Yueosaurus tiantaiensis (Zheng et al. Citation2012), but absent in Orodromeus makelai (Scheetz Citation1999). In other hand, elasmarians, such as Notohypsilophodon comodorensis (Ibiricu et al. Citation2014), Trinisaura santamartaensis (Coria et al. Citation2013), Sektensaurus sanjuanboscoi (Ibiricu et al. Citation2019), and Mahuidacursor lipanglef (Cruzado-Caballero et al. Citation2019) presents ventral median keels.
The secondary loss of the ventral keel in posterior dorsal centra can be viewed as a basal iguanodontian feature: it is reduced in rhabdodontomorphs such as Zalmoxes robustus and Mochlodon suessi (Godefroit et al. Citation2009; Ősi et al. Citation2012) and completely absent in dryomorphans, such as Tenontosaurus tilletti, dryosaurids, Camptosaurus dispar, Hippodraco scutodens, Iguanacolossus forti, Dakotadon lakotaensis and Barilium dawsoni (Lydekker Citation1888; Gilmore Citation1909; Galton Citation1981; Weishampel and Björk Citation1989; Forster Citation1990; McDonald et al. Citation2010). The ventral keel develops secondarily in the clade Cumnoria prestwichii + Uteodon aphanocoetes (see Hulke Citation1880; Carpenter and Wilson Citation2008) as well as in the Hadrosauriformes, as seen in Iguanodon bernissartensis, Mantellisaurus atherfieldensis, Morelladon beltrani, ‘Delapparentia turolensis’, Ouranosaurus nigeriensis, and hadrosaurids (Hooley Citation1925; Ibiricu et al. Citation2014; Gasca et al. Citation2015; Gasulla et al. Citation2015; Verdú et al. Citation2015; Bertozzo et al. Citation2017). Therefore, the ornithischian specimens from the Ilhas Group conform to non-hadrosauriform, and non-rhabodontomorph iguanodontians (i.e. basal ankylopollexians).
Another feature typically seen in ornithopods that can be seen in specimens NHM-PV R.2459 and R.3425 is a series of small neurovascular foramina on the lateral and ventral surfaces of the centra, like what is seen in elasmarians (e.g. Calvo et al. Citation2007; Ibiricu et al. Citation2019) and iguanodontians (e.g. Wang et al. Citation2010). In addition, longitudinal striations circumscribing the articular surface, on the lateral and ventral surfaces of the centrum, are also typically seen in ornithopods, as in the elasmarian Macrogryphosaurus gondwanicus and the iguanodontians Zalmoxes shqiperorum, Tenontosaurus tilleti, Camptosaurus dispar, Sektensaurus sanjuanboscoi, Iguanodon bernissartensis, Morelladon beltrani, Ouranosaurus nigeriensis and hadrosaurids (Galton and Powell Citation1980; Norman Citation1980; Weishampel et al. Citation2003; Cruzado-Caballero and Powell Citation2017). This feature is seen in NHM-PV R.2459, though not in R.3425, possibly due to erosion.
Within ornithopods, the Bahia ornithischian material stands apart from the posterior dorsal centra of most elasmarians in several aspects. It is characterised by being higher than wider, displaying a posteroventral margin deeper than the anteroventral one, and featuring smooth ventral surfaces, contrasting with the conditions observed in Talenkauen santacrucensis (see Rozadilla et al. Citation2019), Mahuidacursor lipanglef (see Cruzado-Caballero et al. Citation2019), and Macrogryphosaurus gondwanicus (see Rozadilla et al. Citation2019), which clearly exhibit ventral keels. Bahia material further differs of Isasicursor santacrucensis where the posterior dorsal centra are higher than wide and ventrally grooved (see Novas et al. Citation2019).
Within the Iguanodontia, the posterior dorsal centra are as wide as high in the Rhabdodontomorpha (Godefroit et al. Citation2009; Ősi et al. Citation2012), thus not matching the Bahia material. In Tenontosaurus tilleti, anterior dorsal centra exhibit comparatively straight ventral margins, being equally wide as tall (except for dorsal 1 and 2) and exhibit ventral keels (Forster Citation1990), thus not matching the Bahia material. The posterior dorsal centra of Tenontosaurus tilleti lack ventral keels and exhibit concave ventral margins; still, they differ from the Bahia material in being about twice as high as long (instead of as high as long), having subcircular/weakly heart-shaped articular surfaces, and in lacking a posteroventral margin deeper than the anteroventral margin. In the Dryosauridae, the presence of a ventral keel is also limited to the anterior dorsal centra, with middle and posterior dorsal centra lacking it (Galton Citation1981; Barrett Citation2016). Still, they differ from the Bahia material in that the articular surfaces are round (slightly heart-shaped) and as wide as tall in all dorsal centra, as can be seen in Valdosaurus canaliculatus, Dysalotosaurus lettowvorbecki, and Dryosaurus altus.
Within the Ankylopollexia, the Hadrosauriformes can be characterised by exhibiting ventral keels throughout the whole dorsal series, as can be seen in Iguanodon bernissartensis, Mantellisaurus atherfieldensis, Ouranosaurus nigeriensis, Morelladon beltrani and hadrosaurids (Norman Citation1980; Gasulla et al. Citation2015; Bertozzo et al. Citation2017; Rozadilla et al. Citation2022), including the Argentinian hadrosaurids Huallasaurus australis, Lapampasaurus chollinoi and Bonapartesaurus rionegrensis (Prieto-Marquez and Salinas Citation2010; Coria et al. Citation2012; Cruzado-Caballero and Powell Citation2017). The Bahia material further differs from the Argentinian hadrosaurids Huallasaurus australis, Bonapartesaurus rionegrensis and Lapampasaurus chollinoi in that the posterior dorsal centra of these forms are opisthocoelous, ventrally keeled, and with heart-shaped articular surfaces.
Therefore, the Bahia material can be excluded from the Rhabdodontomorpha, Dryosauridae, Tenontosaurus and Hadrosauriformes – thus leaving the basal ankylopollexian grade. Within non-hadrosauriform Ankylopollexia, as mentioned above, ventral keels are absent in the posterior dorsals, as can be seen in Camptosaurus dispar, Hippodraco scutodens, Iguanacolossus fortis, Dakotadon lakotaensis and Barilium dawsoni; to the exception of only Cumnoria prestwichii and Uteodon aphanocoetes. In Camptosaurus dispar, posterior dorsal centra are slightly opisthocoelous, about as tall as wide and as tall as long, with articular surfaces round/slightly heart-shaped, posteroventral margin deeper than the anterior one, and parallel articular surfaces; thus, differing from the Bahia material.
In Hippodraco scutodens, the posterior dorsal centra are slightly platycoelous, taller than wide, with oval articular surfaces, with the posteroventral margin conspicuously deeper than the anterior one, and the anterior articular surface inclined relative to the other – matching the Bahia material. In Iguanacolossus fortis, the relative height, inclination, and relief of the articular surfaces of the posterior dorsal centra cannot be assessed due to damage (anteroposterior crushing), but the centra exhibit the posteroventral margin deeper than the anterior one, they are as wide as tall, and the articular surfaces are round/slightly heart shaped. In Barilium dawsoni, the articular surfaces are rounded, with everted margins, as tall as wide, taller than long, and the posteroventral margin is not deeper than the anteroventral (Norman Citation2010). Furthermore, the Bahia material also shares with basal ankylopollexians an interesting feature: the posteroventral margin of the centrum, in lateral view, is deeper than the anteroventral margin. This same configuration can be seen in Camptosaurus dispar, Uteodon aphanoecetes, Cumnoria prestwichii, Hippodraco scutodens, Iguanacolossus fortis, Barilium dawsoni, Hypselospinus fittoni and Morelladon beltrani. It is absent, however, in elasmarians, dryosaurids, Tenontosaurus tilleti, rhabdodontids, Hypsilophodon foxii, and more advanced ankylopollexians such as Mantellisaurus altherfieldensis, Ouranosaurus nigeriensis, Equijubus normani, Probactrosaurus gobienesis, and hadrosaurids. Therefore, the Bahia material matches the morphology of basal ankylopollexians. Amongst all known iguanodontian posterior dorsals, the Bahia iguanodont material most closely resembles Hippodraco scutodens. They share the oval articular surfaces, higher than wider articular surfaces (Table S2), and the posterior inclination of the anterior articular surface relative to the posterior articular surface (see ).
The dinosaur fauna from Bahia State and its paleobiogeographic implications
Ornithischia
The fossil record of Ornithischia from Gondwana is relatively scarce compared to Laurasian occurrences, particularly in South America (see Cruzado-Caballero et al. Citation2018). Sauropods dominated as primary large herbivores in this region throughout the Mesozoic interval, suggesting that ornithischians likely played a secondary role in these dinosaur communities (e.g. Bonaparte and Kirlan-Jawaroska Citation1987; but see; Ezcurra and Agnolín Citation2012). The uneven presence of ornithischians in South America may be influenced by various factors, including differential collection efforts, taphonomical biases, or even reflect a true provincialism driven by climatic constraints, generating a conspicuous latitudinal diversity gradient (see Upchurch et al. Citation2002; Ibiricu et al. Citation2021; Chiarenza et al. Citation2022). The recognition of new ornithischian taxa in South America, however, has experienced its more prolific period since its first discoveries (e.g. Brett-Surman Citation1979; Bonaparte et al. Citation1984). Most of the ornithischian diversity from South America comes from Upper Cretaceous deposits of Argentina (), with few occurrences from Jurassic deposits (e.g. Pol et al. Citation2011; Barrett et al. Citation2014; Salgado et al. Citation2017; Rauhut et al. Citation2020) or from other localities (e.g. Huene von Citation1934; Soto et al. Citation2012; Soto-Acuña et al. Citation2021; Alarcón-Muñoz et al. Citation2023).
Table 3. Main occurrences of ornithischian bones from South America.
Aside from osteological remains, ornithischian ichnites have also been recovered from several localities in South America, mainly in Uruguay, Bolivia, and Brazil (Leonardi Citation1994). Regarding the Brazilian record, it is the most expressive in terms of diversity and number of occurrences, including trackways and some isolated footprints of quadrupedal ornithischians (mostly related to thyreophorans) and several morphotypes related to ornithopods (see and Supplementary data). Notably, the fauna of Bahia State exhibits more similarities with the ichnocoenosis from the coeval Rio do Peixe and Iguatu basins (Arai Citation2006).
In the Rio do Peixe Basin two ichnogenera have been attributed to iguanodontian trackmakers: Caririchnium magnificus and Sousaichnium pricei (Leonardi Citation1979, Citation1989). It is inferred that these tracks were produced by large-sized graviportal iguanodontians, conforming to the basal styracosternans identified here from the Bahia State. In addition, some small tracks of bipedal ornithopods from the Rio do Peixe Basin (Staurichnium diogenis) have been tentatively attributed to juvenile ‘iguanodontids’ or even possibly dryosaurids (Leonardi and Carvalho Citation2002, Citation2021). These referrals of probable trackmakers were based on the purported presence of such clades in Argentina, such as Gasparinisaura cincosaltensis (Coria and Salgado Citation1996a), Anabisetia saldiviai (Coria and Calvo Citation2002), and Talenkauen santacrucensis (Novas et al. Citation2004). However, all these taxa are currently regarded as components of an endemic Gondwanan clade, Elasmaria (e.g. Calvo et al. Citation2007; Madzia et al. Citation2018). Thus, we consider that these ornithopod tracks from the Rio de Peixe Basin most likely were produced by basal styracosternans and elasmarians, rather than dryosaurids.
Until this contribution, elasmarians were known from slightly younger strata than those from Bahia State, such as the Barremian to Cenomanian deposits of Australia (Herne et al. Citation2019), the Cenomanian to Maastrichtian of Argentina (Ibiricu et al. Citation2019), and the Campanian – Maastrichtian of Chile and Antarctica (Coria et al. Citation2013; Rubilar-Rogers et al. Citation2013; Rozadilla et al. Citation2016). By contrast, the osteological remains of styracosternans in South America are limited to hadrosaurid records, most from Argentina (). Hadrosaurids are globally restricted to the Late Cretaceous, with Argentinian forms dating from the Campanian-Maastrichtian. A longstanding view is that the presence of hadrosaurids in South America probably represents a dispersion event coming from North America at some point in the Late Cretaceous (Prieto-Marquez and Salinas Citation2010; Coria et al. Citation2012; Cruzado-Caballero and Powell Citation2017; Alarcón-Muñoz et al. Citation2023). Nonetheless, it is unclear whether basal styracosternans were still present in South America when hadrosaurids colonised the continent, although it is tempting to speculate that hadrosaurids could have replaced the earlier basal lineages. The presence of elasmarians and early-diverging styracosternans in the Lower Cretaceous of Northeast Brazil, however, does not support the temporal coexistence of all these distinct lineages in South America.
The newly identified fauna from Bahia fills a critical gap in ornithischian evolution and extends the presence of styracosternans in South America into the Lower Cretaceous, increasing the diversity of ornithischians in this region. The identification of the Bahia ornithischian specimens as elasmarians and early-diverging styracosternans confirm its presence in Brazil through osteological remains, which is consistent with the long-known record of ornithopod tracks in the Rio do Peixe Basin.
Sauropoda
Diplodocoidea
The clade Diplodocidae experienced its maximum diversity during the Late Jurassic, in particular North America and Europe (Tschopp et al. Citation2015), with few taxa known from the Gondwnana, such as Tornieria africana from the Kimmeridgian – Tithonian of Tanzania (Remes Citation2006) and an unnamed diplodocinae from Argentina (Rauhut et al. Citation2015). In exception of Leinkupal laticauda, from the Berriasian – Valanginian Bajada Colorada Formation, any other Gondwanan and post-Tithonian diplodocine remains is still dubious (Whitlock Citation2011; Gallina et al. Citation2014). Concerning the NHM-PV R.2981(b) and R.3427 specimens, not only represent the first confirmed diplodocines from Brazil but also encompass one of the rare occurrences of Cretaceous diplodocines. This establishes the Bahia diplodocines a late-surviving lineage, and further highlights their presence as some of the northernmost occurrence of the clade in South America.
The similarity between NHM-PV R.3427 within some Jurassic taxa, especially Tornieria africana, fills a gap concerning the ‘Jurassic-type’ neosauropods and their survival at the Jurassic – Cretaceous boundary, as observed in some earliest Cretaceous deposits, such as the Kirkwood Formation. The fauna found in the Recôncavo Basin is very similar to this unit, sharing the presence of iguanodontians (Forster et al. Citation2023), theropods (De Klerk et al. Citation2000; Forster et al. Citation2009), as well as diplodocine and titanosauriform sauropods (McPhee et al. Citation2016). Our work includes the first evidence that Diplodocidae coexisted with advanced lithostrotian titanosaurs in the Lower Cretaceous, given the presence of NHM-PV R.2129(a). This reinforces that the transition of the Jurassic to Cretaceous neosauropod turnover was a gradual replacement, as observed in previous works (e.g. McPhee et al. Citation2016). The identification of at least two distinct neosauropod lineages in the Recôncavo Basin reinforces that the fossil record obfuscates attempts at estimating comparative faunal diversities throughout time if even the incomplete material is ignored. The estimates concerning dinosaurian diversity are strongly biased in nominal species as the only unit of comparison, due to the earliest Cretaceous record is still largely composed of fragmentary specimens. This is compatible with contemporaneous formations such as the Wessex Formation, which appears to have superficially low diversity concerning neosauropods specimens.
Titanosauria
Neosauropods at the Lower Cretaceous are quite rare and comparing with the latest Jurassic represented by the richness of species from the Morrison and Tendaguru formations, its being suggested a sharp decline in neosauropod diversity at the outset of the Cretaceous (Mannion et al. Citation2011; Cashmore et al. Citation2020). Within the Lower Cretaceous, the presence of titanosaurians is even more limited, especially in pre-Aptian deposits (Gorscak and O’Connor Citation2016). Notably, some previously assigned titanosaurs from this period, such as Triunfosaurus leonardii and Ligabuesaurus leanzai, have been recently reclassified as non-titanosaur somphospondylians (Poropat et al. Citation2017; Bellardini et al. Citation2023). Currently, the oldest recorded titanosaur occurrence is represented by Ninjatitan zapatai from the Berriasian – Valanginian Bajada Colorada Formation (Gallina et al. Citation2021), recovered as a probable Colossosauria. However, this taxon only preserves the anteriormost caudal vertebra, displaying a slightly procoelous condition, which hampers major comparisons with the NHM-PV R.2129(a).
As NHM-PV R.2129(a), several other pre-Aptian titanosaur taxa also exhibit well-developed procoely, particularly in the middle and posterior elements. They include Tengrisaurus starkovi from the Murtoi Formation (Valanginian), Volgatitan simbirskiensis from an unnamed unit (Hauterivian), both in Russia, and NHMUK 5333 from the Wessex Formation (Barremian) on the Isle of Wight, United Kingdom. Additional taxa that display procoelous caudal vertebrae, include the early lithostrotian titanosaurian Mnyamawamtuka moyowamkia from the Aptian Mtuka Member of the Galula Formation, which has anterior caudal vertebrae with ‘mild procoely’. In contrast, the basal lithostrotian Malawisaurus dixeyi presents strongly procoelous anterior caudal vertebrae followed by platycoelous and amphyplatyan middle and posterior caudal vertebrae.
The acquisition of strongly procoelous middle caudal vertebrae appears to be a non-uniform osteological feature throughout titanosaurian evolution. This characteristic might be biomechanically advantageous as it provides greater stabilisation against shear loads (Fronimos et al. Citation2016). Interestingly, the irradiative evolution of Titanosauria appears to coincide with the faunal turnover event occurred in the Cenomanian – Turonian (Coria and Salgado Citation2005), aligning with the acquisition of the mechanically advantageous procoelous vertebrae and their rotational stability. Changes in the landscape and habitat distribution could have influenced the diversification and dispersal of these dinosaurs, and the acquisition of the procoelous vertebrae, may have allowed certain lineages of titanosaurs to explore new palaeoecological niches. For example, the rotational stability provided by the presence of the procoelous vertebrae may have important functional implications, impacting the ability of titanosaurs to move efficiently and other behavioural aspects, such as foraging strategies, locomotion, or reproduction.
Gondwanan taxa in the post-Cenomanian, such as Andesaurus delgadoi (Mannion and Calvo Citation2011) and Mendozasaurus neguyelap (González-Riga et al. Citation2018), exhibit strongly procoelous anterior caudal vertebrae, transitioning from slightly procoelous to amphyplatyan in the middle and posterior caudal vertebrae. In contrast, older Laurasian taxa like Volgatitan simbirskiensis display well-developed procoelous anterior and middle caudal vertebrae. This implies that the presence of strongly procoelous anterior vertebrae has a much older origin within the titanosaur evolutionary history – an unexpected pattern considering the more primitive somphospondylan taxa, where the platycoelous and amphyplatyan condition is widespread. An example of this can be seen in somphospondylan Malarguesaurus florenciae from Upper Cretaceous (Turonian-Coniacian) of Argentina, which exhibits platycoelous anterior to mildly procoelous middle caudal vertebrae (González-Riga et al. Citation2009). The presence of NHM PV R. 2129a not only supports the occurrence of procoelous anterior to middle caudal vertebrae, but also indicates an intricate scenario in the diversification of lithostrotian titanosaurs during the Early Cretaceous.
Theropoda
Spinosauridae
Spinosauridae is classically divided into Baryonychinae and Spinosaurinae (e.g. Sereno et al. Citation1998; Bertin Citation2010; Malafaia et al. Citation2019), where the former comprises mostly Laurasian and North African forms, while the latter was widely distributed (Barrett et al. Citation2011; Hone and Holtz Citation2017). Most of the spinosaurid fossil record come from Aptian – Albian deposits (e.g. Bittencourt and Kellner Citation2004; Kellner et al. Citation2011; Allain et al. Citation2012; Sales et al. Citation2017). Dubious occurrences are reported for older stages of the Lower Cretaceous, as some teeth from Tanzania (including Ostafrikasaurus crassiserratus; Buffetaut Citation2013) and Niger. These findings indicate a putative Jurassic occurrence of this clade (Buffetaut Citation2008; Serrano-Martínez et al. Citation2015). Santonian remains are known from some teeth from China (Hone et al. Citation2010) and an isolated ungual from North America (Allain et al. Citation2012). The evolutionary placement of those taxa is critical to support a Pangean distribution of spinosaurids before the Tethys opening.
Recent studies suggest that the divergence between spinosaurines and baryonychines occurred at least before Barremian (Sales et al. Citation2017). The main divergence regards the Iberian and North African forms and its evolutionary significance (Buffetaut and Ouaja Citation2002; Mateus et al. Citation2011). Sereno et al. (Citation1998) proposed a split between baryonychines and spinosaurines at the end of the Jurassic using a vicariant and dispersion model. Some studies have suggested an evolvement of spinosaurines in North Africa from Albian baryonychine forms, which migrated from Europe to North Africa during the Cretaceous (Machado and Kellner Citation2005).
The rise of spinosaurines in Gondwana has been explained as a faunal turnover with baryonychines in Aptian-Albian North Africa but recent studies suggest a niche partitioning between these theropod forms, at least in Kem Kem beds (Arden et al. Citation2019). Recent fossil findings in both the Iberian Peninsula and North Africa suggests a European origin of Baryonychinae, as well as the similarity between African and European forms, corroborates the migration through Gondwana during Barremian-Aptian (Ruiz-Omeñaca et al. Citation2005; Sánchez-Hernández et al. Citation2007; Canudo et al. Citation2013).
The spinosaurids in South America are recovered mostly from the Northeast of Brazil. Some teeth from the Feliz Deserto Formation (Sergipe-Alagoas Basin, Berriasian – Valanginian) represent the oldest spinosaurines known to date (Lacerda et al. Citation2023). Those findings reinforce a baryonychine/spinosaurine splitting before the Barremian. Prolific spinosaurine remains come from the Aptian record of the Araripe Basin (e.g. Kellner and Campos Citation1996; Martill et al. Citation1996; Sues et al. Citation2002; Bittencourt and Kellner Citation2004; Machado and Kellner Citation2007; Aureliano et al. Citation2018), from the Albian-Cenomanian São Luis-Grajaú Basin (Medeiros and Schultz Citation2002; Medeiros Citation2006; Kellner et al. Citation2011; França et al. Citation2022), and recently by a Spinosauridae tooth from the Albian-Cenomanian of the Potiguar Basin (Pereira et al. Citation2020). A dubious tooth was from the Cenomanian-Turonian of Argentina (Canudo et al. Citation2004), which potentially comprises the southernmost record of a spinosaurid in South America. The materials NHM-PV R.2980 (and possibly R.2982) represent the oldest osteological record in Brazil and the southernmost in the country. Although detailed comparisons are scarce, the centrum of NHM-PV R.2980 is remarkably distinct from the Brazilian specimen MN 4743-V (Bittencourt and Kellner Citation2004). Regarding its robust shape, the ventral surface with a paired ventral keel, a shallow and wide groove, NHM-PV R.2980 resembles the Barremian taxa, Baryonyx walkeri (Charig and Milner Citation1986) and the Spanish spinosaurine Vallibonavenatrix cani (Malafaia et al. Citation2019). The specimens described here corroborate a complex relation within spinosaurid forms of Iberian-North Africa regions. Also, NHM-PV R.2980 represents a wide distribution of spinosaurids in the Lower Cretaceous Brazilian coast, as well as a wide time range of the clade.
Carcharodontosauridae
Carcharodontosaurids are notable as they represent large-bodied theropods with a widespread distribution. The group achieved a wide distribution during the ‘Middle’ Cretaceous (Aptian-Cenomanian), from which most of the fossil record of this group comes and is considered extinct before the Turonian interval. The carcharodontosaurian record spans North America (Stovall and Langston Citation1950), Asia (Brusatte et al. Citation2009; Chokchaloemwong et al. Citation2019), Europe (Ortega et al. Citation2010), Africa (Sereno et al. Citation1996; Sereno and Brusatte Citation2008; Cau et al. Citation2013; Evers et al. Citation2015), and South America (Coria and Salgado Citation1995; Novas et al. Citation2005; Coria and Currie Citation2006; Canale et al. Citation2015; Motta et al. Citation2016; Coria et al. Citation2020). Oldest carcharodontosaurian occurrences are dated back to the Jurassic and include isolated teeth from China (Han et al. Citation2011) and Germany (Gerke et al. Citation2016), Veterupristisaurus milneri from Tanzania (Rauhut Citation2011; Carrano et al. Citation2012), Lusovenator santosi from Portugal (Malafaia et al. Citation2020), and an unnamed taxon from the Sergi Formation of Brazil (Bandeira et al. Citation2021), which suggests that its diversification is related to the Pangean fragmentation (Sereno and Brusatte Citation2008).
However, Pre-Aptian records of carcharodontosaurians are scarce globally. One exception, Lajasvenator ascheriae from the Valanginian of Argentina (Coria et al. Citation2020), indicates that carcharodontosaurids are present in southern regions of South America in pre-Aptian stages. Other carcharodontosaurian occurrences remain dubious. The reassessment of a theropod tooth from Romania revealed that the group could also be present in the Valanginian of Laurasia, but such assignment could be an artefact of the analysis given the incompleteness of such specimen (Csiki-Sava et al. Citation2016). Briefly speaking, the Tithonian to Aptian interval has scarce occurrences concerning reliable carcharodontosaurian records.
Carcharodontosaurid remains from Brazil are usually represented by isolated teeth from the Albian-Cenomanian deposits of the São Luís-Grajaú and Potiguar basins (e.g. Medeiros and Schultz Citation2002), as well as few fragmentary occurrences of axial bones (Medeiros et al. Citation2014, Citation2019; Pereira et al. Citation2020). Although NHM-PV R.2981(a) and R.2129(b) lack diagnostic features, the general shape of the partial centra of those specimens could refer them to Carcharodontosauridae, broadening both the temporal and spatial range of this clade. Despite their fragmentary nature, these specimens fill a critical gap in a poorly sampled period of the carcharodontosaurian evolution.
Conclusions
In this contribution, we revisited the putative dinosaur remains collected between 1859 and 1906 in the Lower Cretaceous coastal deposits (Massacará and Ilhas groups) of the Recôncavo Basin, Bahia State. The evaluation of this historical material provided a new glimpse into the dinosaur faunas present in Northeast Brazil, enhancing the understanding about dinosaur diversity of northern South America during the earliest Cretaceous (.
Figure 20. Time-scaled strict consensus tree depicting the phylogenetic relationships of NHM-PV R.2129(a) within Titanosauria, included in an updated version of the matrix of Navarro et al. (Citation2022). Bremer supports are summarised in the supplementary data. Silhouettes from several sources.
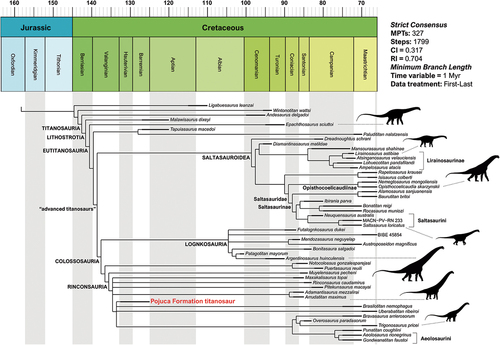
Figure 21. Paleontographical reconstruction of the hypothetical paleoenvironment of the Recôncavo Basin during Lower Cretaceous: A, faunal components of the Marfim Formation (Valanginian–Hauterivian); B, faunal components of the Pojuca Formation (Hauterivian–Barremian). The Salvador Formation is partially synchronous with both units, sharing coeval components. Artwork by Matheus Gadelha.

Albeit most of the recognised specimens comprise fragmentary material, they offer several diagnostic characters that allow the identification at least of two lineages of theropods, sauropods, and ornithischians. Among them, includes the first nominal species of an elasmarian from Brazil, being the most ancient record of this group, as well as remains of probable two iguanodontian taxa. Alongside the ichnological data, these findings enhance our understanding about paleobiogeographical aspects of this group prior to the Gondwanan rifting. Among the two identified sauropod clades, includes the first diplodocid material from Brazil, being the northmost occurrence of Gondwana, as well as the earliest known rinconsaurian titanosaur to date. Regarding theropod dinosaurs, our contribution identified the probable presence of spinosaurids and carcharodontosaurians, which, as in other chronocorrelated basins around the world, comprise the apex predators of this paleoecosystem.
The dinosaurian components of Recôncavo Basin resemble those recorded in contemporaneous units of Northeast Brazil, such as the Sergipe-Alagoas, Rio do Peixe, and Iguatu basins, sharing the faunal association of ornithischians alongside sauropod and theropod dinosaurs. In a broad perspective, the dinosaur fauna from Recôncavo Basin also exhibits striking similarities with African dinosaur faunas from the Jurassic – Cretaceous boundary.
The specimens described in this study hold historical significance as they represent the first dinosaur bones collected from South America. They highlight the fossiliferous potential of the Cretaceous deposits in Bahia State, which have been relatively unexplored since their initial discoveries.
Supplemental Material
Download Zip (1.1 MB)Acknowledgments
We are deeply grateful to the curators Sandra D. Chapman and Lorna Steel (NHM), for allowing access to the specimens under their care, and for their support during the examination of the materials described here. We express our gratitude to Matheus Fernandes Gadelha for providing the reconstruction of the hypothetical paleoenvironment of the Massacará and Ilhas groups and their dinosaurian components. The paper was improved by the thoughtful comments from Penélope Cruzado Caballero and an anonymous reviewer. We also want to thank the Willi Hennig Society and R Core Team for sponsoring the TNT and R software’s.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplemental data
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2024.2318406
Additional information
Funding
References
- Agnolín FL, Rozadilla S. 2018. Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina. J Syst Palaeontol. 16(10):853–879. doi: 10.1080/14772019.2017.1352623.
- Alarcón-Muñoz J, Vargas AO, Püschel HP, Soto-Acuña S, Manríquez L, Leppe M, Kaluza J, Milla V, Gutstein CS, Palma-Liberona J, et al. 2023. Relict duck-billed dinosaurs survived into the last age of the dinosaurs in subantarctic Chile. Sci Adv. 9(24):eadg2456. doi:10.1126/sciadv.adg2456.
- Allain R, Xaisanavong T, Richir P, Khentavong B. 2012. The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos. Naturwissenschaften. 99(5):369–377. doi: 10.1007/s00114-012-0911-7.
- Allport S. 1860. On the discovery of some fossil remains near Bahia, in South America (with notes on the fossil by P. Edgerton, J. Morris and T. Rupert Jones). Quart J Geol Soc London. 16(3):263–266. doi: 10.1144/GSL.JGS.1860.016.01-02.36.
- Ameghino F. 1899. Nota preliminar sobre el Loncosaurus argentinus, un representante de la familia Megalosauridae de la República Argentina. An Soc Cient Argentina. 49:61–62.
- Apesteguía S, Smith ND, Juárez Valieri R, Makovicky PJ, Farke AA. 2016. An unusual new theropod with a didactyl manus from the Upper Cretaceous of Patagonia, Argentina. PloS One. 11(7):e0157793. doi: 10.1371/journal.pone.0157793.
- Arai M. 2006. Revisão estratigráfica do Cretáceo Inferior das bacias interiores do nordeste do Brasil. Geociências. 25(1):7–15.
- Arai M. 2014. Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: a paleontological perspective. Braz J Geol. 44(2):339–350. doi: 10.5327/Z2317-4889201400020012.
- Aranciaga-Rolando AM, Egli FB, Sales MAF, Martinelli AG, Canale JI, Ezcurra MD. 2018. A supposed Gondwanan oviraptorosaur from the Albian of Brazil represents the oldest South American megaraptoran. Cret Res. 84:107–119. doi: 10.1016/j.cretres.2017.10.019.
- Arden TMS, Klein CG, Zouhri S, Longrich NR. 2019. Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurids. Cret Res. 93:275–284. doi: 10.1016/j.cretres.2018.06.013.
- Assine ML, Quaglio F, Warren LV, Simões MG. 2016. Comments on paper by M. Arai “Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: a paleontological perspective”. Braz J Geol. 46(1):3–7. doi: 10.1590/2317-4889201620150046A.
- Aureliano T, Ghilardi AM, Buck PV, Fabbri M, Samathi A, Delcourt R, Fernandes MA, Sander M. 2018. Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil. Cret Res. 90:283–295. doi: 10.1016/j.cretres.2018.04.024.
- Averianov A, Efimov V. 2018. The oldest titanosaurian sauropod of the Northern Hemisphere. Bio Comm. 63(3):145–162. doi: 10.21638/spbu03.2018.301.
- Averianov AO, Lopatin AV. 2020. An unusual new sauropod dinosaur from the Late Cretaceous of Mongolia. J Syst Palaeontol. 18(12):1009–1032. doi: 10.1080/14772019.2020.1716402.
- Averianov AO, Sizov AV, Grigoriev DV, Pestchevitskaya EB, Vitenko DD, Skutschas PP. 2023. New data on dinosaurs from the Lower Cretaceous Murtoi Formation of Transbaikalia, Russia. Cret Res. 138:105287. doi: 10.1016/j.cretres.2022.105287.
- Averianov AO, Sizov AV, Skutschas PP. 2021. Gondwanan affinities of Tengrisaurus, Early Cretaceous titanosaur from Transbaikalia, Russia (Dinosauria, Sauropoda). Cret Res. 122:104731. doi: 10.1016/j.cretres.2020.104731.
- Averianov AO, Skutschas PP. 2017. A new lithostrotian titanosaur (Dinosauria, Sauropoda) from the Early Cretaceous of Transbaikalia, Russia. Biol Comm. 62(1):6–18. doi: 10.21638/11701/spbu03.2017.102.
- Averianov AO, Voronkevich AV, Leshchinskiy SV, Fayngertz AV. 2006. A ceratopsian dinosaur Psittacosaurus sibiricus from the Early Cretaceous of West Siberia, Russia and its phylogenetic relationships. J Syst Palaeontol. 4(4):359–395. doi: 10.1017/S1477201906001933.
- Baez A, Marsicano C. 2001. A heterodontosaur ornithischian dinosaur from the Upper Triassic of Patagonia. Ameghiniana. 38:271–279.
- Bandeira KLN, Brum AS, Pêgas RV, Souza LG, PVLGC P, Pinheiro AEP. 2021. The first Jurassic theropod from the Sergi Formation, Jatobá Basin, Brazil. An Acad Bras Ciênc. 93(2):e20201557. doi: 10.1590/0001-3765202120201557.
- Bandeira KLN, Machado EB, Campos D, Kellner AWA. 2019. New titanosaur (Sauropoda, Dinosauria) records from the Morro do Cambambe Unit (Upper Cretaceous), Mato Grosso state, Brazil. Cret Res. 103:104155. doi: 10.1016/j.cretres.2019.06.001.
- Barrett PM. 2016. A new specimen of Valdosaurus canaliculatus (Ornithopoda: Dryosauridae) from the Lower Cretaceous of the Isle of Wight, England. Mem Mus Vic. 74:24–48. doi: 10.24199/j.mmv.2016.74.04.
- Barrett PM, Benson RBJ, Rich TH, Vickers-Rich P. 2011. First spinosaurid dinosaur from Australia and the cosmopolitanism of Cretaceous dinosaur faunas. Biol Lett. 7(6):933–936. doi: 10.1098/rsbl.2011.0466.
- Barrett PM, Butler RJ, Mundil R, Scheyer TM, Irmis RB, Sánchez-Villagra MR. 2014. A palaeoequatorial ornithischian and new constraints on early dinosaur diversification. Proc R Soc B. 281(1791):20141147. doi: 10.1098/rspb.2014.1147.
- Barta DE, Norell MA. 2021. The osteology of Haya griva (Dinosauria: Ornithischia) from the Late Cretaceous of Mongolia. Bull Am Mus Nat Hist. 445(1):1–112. doi: 10.1206/0003-0090.445.1.1.
- Baumel JJ, Witmer LM. 1993. Osteology. In: Baumel, JJ editor. Handbook of avian anatomy: nomina anatomica avium. 2nd ed. Cambridge, Massachusetts: Nuttal Ornithological Club; p. 45–132.
- Baur G. 1891. Remarks on the reptiles generally called Dinosauria. Am Nat. 25(293):434–454. doi: 10.1086/275329.
- Bellardini F, Coria RA, Pino DA, Windholz GJ, Baiano MA, Martinelli AG. 2023. Osteology and phylogenetic relationships of Ligabuesaurus leanzai (Dinosauria: Sauropoda) from the Early Cretaceous of the Neuquén Basin, Patagonia, Argentina. Zoo J Linn Soc. 196(4):1333–1393. doi: 10.1093/zoolinnean/zlac003.
- Bennett SC. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology. 19(1):92–106. doi: 10.1017/S0094837300012331.
- Benson RBJ. 2008. New information on Stokesosaurus, a tyrannosauroid (Dinosauria: Theropoda) from North America and the United Kingdom. J Vert Palaeontol. 28(3):732–750. doi: 10.1671/0272-4634(2008)28[732:NIOSAT]2.0.CO;2.
- Benson RBJ. 2010. A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods. Zoo J Linn Soc. 158(4):882–935. doi: 10.1111/j.1096-3642.2009.00569.x.
- Benson RBJ, Butler RJ, Carrano MT, O’Connor PM. 2012. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the ‘reptile’–bird transition. Biol Rev. 87(1):168–193. doi: 10.1111/j.1469-185X.2011.00190.x.
- Benson RBJ, Carrano MT, Brusatte SL. 2010. A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic. Naturwissenschaften. 97(1):71–78. doi: 10.1007/s00114-009-0614-x.
- Bertin T. 2010. A catalogue of material and review of the Spinosauridae. PalArch’s J Vert Paleontol. 7(4):1–39.
- Bertozzo F, Dalla Vecchia FM, Fabbri M. 2017. The Venice specimen of Ouranosaurus nigeriensis (Dinosauria, Ornithopoda). PeerJ. 5:e3403. doi: 10.7717/peerj.3403.
- Bittencourt JS, Kellner AWA. 2004. On a sequence of a sacrocaudal theropod dinosaur vertebrae from Lower Cretaceous Santana Formation, Northeastern Brazil. Arq Mus Nac. 62:309–320.
- Bittencourt JS, Langer MC. 2011. Mesozoic dinosaurs from Brazil and their biogeographic implications. An Acad Bras Ciênc. 83(1):23–60. doi: 10.1590/S0001-37652011000100003.
- Bonaparte JF. 1985. A horned Cretaceous carnosaur from Patagonia. Nat Geogr Res. 1(1):149–151.
- Bonaparte JF. 1986. Les Dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés) du Jurassique moyen de Cerro Condor (Chubut, Argentine). Ann Palaeontol. 72:325–386.
- Bonaparte JF. 1991. The Gondwanan theropod families Abelisauridae and Noasauridae. Hist Biol. 5(1):1–25. doi: 10.1080/10292389109380385.
- Bonaparte JF, Coria RA. 1993. Un nuevo y gigantesco sauropodo titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la Provincia del-Neuquen, Argentina. Ameghiniana. 30(3):271–282.
- Bonaparte JF, Franchi MR, Powell JES, Sepúlveda E G. 1984. La Formación Los Alamitos (Campaniano–Maastrichtiano) del sudeste de Río Negro, con descripción de Kritosaurus australis n. sp. (Hadrosauridae). Significado paleogeográfico de los vertebrados. Rev Asoc Geol Argentina. 39(3&4):284–299.
- Bonaparte JF, Kirlan-Jawaroska Z. 1987. Late Cretaceous dinosaurs and mammal faunas of Laurasia and Gondwana. In: Currie, PJ, Koster, EH editors. Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Alberta, Canada: Royal Tyrrell Museum of Palaeontology; p. 24–29.
- Bonaparte JF, Powell JE. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Mém Soc Géol France. 139:19–28.
- Boulenger GA. 1881. Sur l’arc pelvien chez les dinosauriens de Bernissart [On the pelvic arch in the dinosaurs of Bernissart]. Bull mém Acad r méd Belg. 1(5):1–11.
- Boyd CA. 2015. The systematic relationships and biogeographic history of ornithischian dinosaurs. PeerJ. 3:e1523. doi: 10.7717/peerj.1523.
- Boyd CA, Brown CM, Scheetz RD, Clarke JA. 2009. Taxonomic revison of the basal neornithischian taxa Thescelosaurus and Bugenasaura. J Vert Palaeontol. 29(3):758–770. doi: 10.1671/039.029.0328.
- Breeden BT, Raven TJ, Butler RJ, Rowe TB, Maidment SCR. 2021. The anatomy and palaeobiology of the early armoured dinosaur Scutellosaurus lawleri (Ornithischia: Thyreophora) from the Kayenta Formation (Lower Jurassic) of Arizona. R Soc Open Sci. 8(7):201676. doi: 10.1098/rsos.201676.
- Bremer K. 1994. Branch Support and Tree Stability. Cladistics. 10(3):295–304. doi: 10.1111/j.1096-0031.1994.tb00179.x.
- Brett-Surman MK. 1979. Phylogeny and paleobiogeography of hadrosaurian dinosaurs. Nature. 277(5697):560–562. doi: 10.1038/277560a0.
- Brill K, Carpenter K. 2006. A description of a new ornithopod from the Lytle Member of the Purgatoire Formation (Lower Cretaceous) and a reassessment of the skull of Camptosaurus. In: Carpenter, K editor. Horns and Beaks: ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press; p. 49–67.
- Brochu CA. 1996. Crocodylian skeletal maturity. J Vert Palaeontol. 16(1):1–14.
- Brochu CA. 2003. Osteology of Tyrannosaurus rex: Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. J Vert Palaeontol. 22(sup4):1–138. doi: 10.1080/02724634.2003.10010947.
- Brown CM, Boyd CA, Russel AP. 2011. A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. Zoo J Linn Soc. 163(4):1157–1198. doi: 10.1111/j.1096-3642.2011.00735.x.
- Brown CM, Evans DC, Ryan MJ, Russell AP. 2013. New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta. J Vert Palaeontol. 33(3):495–520. doi: 10.1080/02724634.2013.746229.
- Brusatte SL, Benson RBJ, Chure DJ, Xu X, Sullivan C, Hone DWE. 2009. The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids. Naturwissenschaften. 96(9):1051–1058. doi: 10.1007/s00114-009-0565-2.
- Buffetaut E. 2008. Spinosaurid teeth from the Late Jurassic of Tendaguru, Tanzania, with remarks on the evolutionary and biogeographical history of the Spinosauridae. In: Mazin J, Pouech P, Hantzpergue P, Lacombe V, editors. Mid-Mesozoic life and environments. Lyon: Documents des Laboratoires de Géologia Lyon; p. 26–28.
- Buffetaut E. 2013. An early spinosaurid dinosaur from the Late Jurassic of Tendaguru (Tanzania) and the evolution of the spinosaurid dentition. Oryctos. 10:1–8.
- Buffetaut E, Ouaja M. 2002. A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae. Bull Soc Géol France. 173(5):415–421. doi: 10.2113/173.5.415.
- Buffetaut E, Taquet P. 1977. The giant crocodilian Sarcosuchus in the early Cretaceous of Brazil and Niger. Palaeontol. 20(1):203–208.
- Bunzel E. 1871. Die Reptilfauna der Gosauformation in der Neuen Welt bei Weiner-Neustadt. Abh K-kön geol Reichsanst. 5:1–18.
- Butler RJ. 2010. The anatomy of the basal ornithischian dinosaur Eocursor parvus from the lower Elliot Formation (Late Triassic) of South Africa. Zoo J Linn Soc. 160(4):648–684. doi: 10.1111/j.1096-3642.2009.00631.x.
- Butler RJ, Liyong J, Jun C, Godefroit P. 2011. The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian-Cenomanian) of Jilin Province, north-eastern China. Palaeontology. 54(3):667–683. doi: 10.1111/j.1475-4983.2011.01046.x.
- Butler RJ, Smith RMH, Norman DB. 2007. A primitive ornithischian dinosaur from the Late Triassic of South Africa, and the early evolution and diversification of Ornithischia. Proc R Soc B. 274(1621):2041–2046. doi: 10.1098/rspb.2007.0367.
- Butler RJ, Zhao Q. 2009. The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China. Cret Res. 30(1):63–77. doi: 10.1016/j.cretres.2008.03.002.
- Caixeta JM, Bueno GV, Magnavita LV, Feijó FJ. 1994. Bacias do Recôncavo, Tucano e Jatobá. Bol Geoc Petrobras. 8(1):163–172.
- Calvo JO, Bonaparte JF. 1991. Andesaurus delgadoi gen. et sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquén, Argentina. Ameghiniana. 28(3–4):303–310.
- Calvo JO, Porfiri JD, Novas FE. 2007. Discovery of a new ornithopod dinosaur from the Portezuelo formation (Upper Cretaceous), Neuquén, Patagonia, Argentina. Arq Mus Nac Rio de Janeiro. 65(4):471–483.
- Calvo JO, Salgado L. 1995. Rebbachisaurus tessonei sp. nov. A new sauropod from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia. 11:13–33.
- Campos DDA, Kellner AWA. 1991. Dinosaurs of the Santana Formation with comments on other Brazilian occurrences. In: Masey J, editor. Santana Fossils, an illustrated atlas. Neptune, New Jersey: T.F.H. Pubns Inc; p. 372–375.
- Canale JI, Novas FE, Pol D. 2015. Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) from the Lower Cretaceous of Patagonia, Argentina. Hist Biol. 27(1):1–32. doi: 10.1080/08912963.2013.861830.
- Canudo JI, Gasulla JM, Gómez-Fernández D, Ortega F, Sanz JL, Yagüe P. 2013. Primera evidencia de dientes aislados atribuidos a Spinosauridae (Theropoda) en el Aptiano inferior (Cretácico Inferior) de Europa: Formación Arcillas de Morella (España). Ameghiniana. 45(4):649–662.
- Canudo JI, Salgado L, Barco JL, Bolatti R, Ruiz-Omeñaca JI. 2004. Dientes de dinosaurios terópodos y saurópodos de la Formación Cerro Lisandro (Cenomaniense superior-Turoniense inferior, Cretácico superior) en Río Negro (Argentina). Geo Temas. 6(5):31–34.
- Carballido JL, Scheil M, Knötschke N, Sander PM. 2020. The appendicular skeleton of the dwarf macronarian sauropod Europasaurus holgeri from the Late Jurassic of Germany and a re-evaluation of its systematic affinities. J Syst Palaeontol. 18(9):739–781. doi: 10.1080/14772019.2019.1683770.
- Carpenter K, Miles C, Cloward K. 2005. New small theropod from the Upper Jurassic Morrison Formation of Wyoming. In: Carpenter K, editor. The Carnivorous Dinosaurs. Bloomington and Indianapolis: Indiana University Press; p. 23–48.
- Carpenter K, Wilson Y. 2008. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of Its forelimb. Ann Carnegie Mus. 76(4):227–263. doi: 10.2992/0097-4463(2008)76[227:ANSOCO]2.0.CO;2.
- Carrano MT. 2007. The appendicular skeleton of Majungasaurus crenatissimus (Theropoda; Abelisauridae) from the Late Cretaceous of Madagascar. J Vert Palaeontol. 27(sup2):163–179. doi: 10.1671/0272-4634(2007)27[163:TASOMC]2.0.CO;2.
- Carrano MT, Benson RBJ, Sampson SD. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). J Syst Palaeontol. 10(2):211–300. doi: 10.1080/14772019.2011.630927.
- Carrano MT, Sampson SD. 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). J Syst Palaeontol. 6(2):183–236. doi: 10.1017/S1477201907002246.
- Carrano MT, Sampson SD, Forster CA. 2002. The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. J Vert Palaeontol. 22(3):510–534. doi: 10.1671/0272-4634(2002)022[0510:TOOMKA]2.0.CO;2.
- Carvalho IDS, Agnolín FL, Rozadilla S, Novas FE, Andrade JAFG, Xavier-Neto J. 2021. A new ornithuromorph bird from the Lower Cretaceous of South America. J Verteb Paleontol. 41(4):4. doi: 10.1080/02724634.2021.1988623.
- Carvalho IDS, Avilla LS, Salgado L. 2003. Amazonsaurus maranhensis gen. et sp. nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) of Brazil. Cret Res. 24(6):697–713. doi: 10.1016/j.cretres.2003.07.005.
- Carvalho IDS, Novas FE, Agnolín FL, Isasi MP, Freitas FI, Andrade JA. 2015. A new genus and species of enantiornithine bird from the Early Cretaceous of Brazil. Braz J Geol. 45(2):161–171. doi: 10.1590/23174889201500020001.
- Carvalho IDS, Salgado L, Lindoso RM, Araújo-Júnior HID, Nogueira FCC, Soares JA. 2017. A new basal titanosaur (Dinosauria, Sauropoda) from the Lower Cretaceous of Brazil. J South Am Earth Sci. 75:74–84. doi: 10.1016/j.jsames.2017.01.010.
- Casal GA, Martínez RD, Luna M. 2016. Ordenamiento y caracterización faunística del Cretacico Superior del Grupo Chubut, Cuenca del Golfo San Jorge, Argentina. Rev Bras Paleontol. 19(1):53–70.
- Casamiquela RM. 1967. Un nuevo dinosaurio ornitisquio triasico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto, Argentina. Ameghiniana. 5(2):47–64.
- Cashmore DD, Mannion PD, Upchurch P, Butler RJ. 2020. Ten more years of discovery: revisiting the quality of the sauropodomorph dinosaur fossil record. Palaeontol. 63(6):951–978. doi: 10.1111/pala.12496.
- Cau A, Dalla Vecchia FM, Fabbri M. 2013. A thick-skulled theropod (Dinosauria, Saurischia) from the Upper Cretaceous of Morocco with implications for carcharodontosaurid cranial evolution. Cret Res. 40:251–260. doi: 10.1016/j.cretres.2012.09.002.
- Charig AJ, Milner AC. 1986. Baryonyx, a remarkable new theropod dinosaur. Nature. 324(6095):359–361. doi: 10.1038/324359a0.
- Chiappe LM. 1996. Late Cretaceous birds of southern South America: anatomy and systematics of Enantiornithes and Patagopteryx deferrariisi. Münchner Geowissenschaftliche Abhandlungen. 30:203–244.
- Chiarenza AA, Mannion PD, Farnsworth A, Carrano MT, Varela S. 2022. Climatic constraints on the biogeographic history of Mesozoic dinosaurs. Curr Bio. 32(3):570–585. doi: 10.1016/j.cub.2021.11.061.
- Choiniere JN, Clark JM, Forster CA, Norell MA, Eberth DA, Erickson GM, Chu H, Xu X. 2014. A juvenile specimen of a new coelurosaur (Dinosauria: Theropoda) from the Middle–Late Jurassic Shishugou Formation of Xinjiang, People’s Republic of China. J Syst Palaeontol. 12(2):177–215. doi: 10.1080/14772019.2013.781067.
- Choiniere JN, Clark JM, Forster CA, Xu X. 2010. A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People’s Republic of China. J Vert Palaeontol. 30(6):1773–1796. doi: 10.1080/02724634.2010.520779.
- Choiniere JN, Forster CA, de Klerk WJ. 2012. New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa. J Afr Earth Sci. 71–72:1–17. doi: 10.1016/j.jafrearsci.2012.05.005.
- Chokchaloemwong D, Hattori S, Cuesta E, Jintasakul P, Shibata M, Azuma Y, Liu J. 2019. A new carcharodontosaurian theropod (Dinosauria: Saurischia) from the Lower Cretaceous of Thailand. PloS One. 14(10):e0222489. doi: 10.1371/journal.pone.0222489.
- Chure D, Britt BB, Whitlock JA, Wilson JA. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften. 97(4):379–391. doi: 10.1007/s00114-010-0650-6.
- Cooper MR. 1985. A revision of the ornithischian dinosaur Kangnasaurus coetzeei Haughton, with a classification of the Ornithischia. Ann S Afr Mus. 95(8):281–317.
- Cope ED. 1886. A contribution to the vertebrate paleontology of Brazil. Proc Am Philos Soc. 23(121):1–21.
- Coria RA. 2014. South American hadrosaurs: considerations on their diversity. In: Eberth D, Evans D, editors. Hadrosaurs. Bloomington: Indiana University Press; p. 332–339.
- Coria RA. 2016. An overview of the ornithischian dinosaurs from Argentina. Cont Del Mus Argent Cienc Nat. 6:109–117.
- Coria RA, Calvo JO. 2002. A new iguanodontian ornithopod from Neuquén Basin, Patagonia, Argentina. J Vert Palaeontol. 22(3):503–509. doi: 10.1671/0272-4634(2002)022[0503:ANIOFN]2.0.CO;2.
- Coria RA, Chiappe LM, Dingus L. 2002. A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia. J Vert Palaeontol. 22(2):460–465. doi: 10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2.
- Coria RA, Currie PJ. 2006. A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. Geodiversitas. 28(1):71–118.
- Coria RA, Currie PJ, Ortega F, Baiano MA. 2020. An Early Cretaceous, medium-sized carcharodontosaurid theropod (Dinosauria, Saurischia) from the Mulichinco Formation (upper Valanginian), Neuquén Province, Patagonia, Argentina. Cret Res. 111:104319. doi: 10.1016/j.cretres.2019.104319.
- Coria RA, González-Riga BJ, Casadío S. 2012. Un nuevo hadrosáurido (Dinosauria, Ornithopoda) de la Formación Allen, provincia de la Pampa, Argentina. Ameghiniana. 49(4):552–572. doi: 10.5710/AMGH.9.4.2012.487.
- Coria RA, Moly JJ, Reguero M, Santillana S, Marenssi S. 2013. A new ornithopod (Dinosauria; Ornithischia) from Antarctica. Cret Res. 41:186–193. doi: 10.1016/j.cretres.2012.12.004.
- Coria RA, Salgado L. 1995. A new giant carnivorous dinosaur from the Cretaceous of Patagonia. Nature. 377(6546):224–226. doi: 10.1038/377224a0.
- Coria RA, Salgado L. 1996a. A basal iguanodontian (Ornithischia: Ornithopoda) from the Late Cretaceous of South America. J Vert Palaeontol. 16(3):445–457. doi: 10.1080/02724634.1996.10011333.
- Coria RA, Salgado L. 1996b. “Loncosaurus argentinus” Ameghino, 1899 (Ornithischia, Ornithopoda): a revised description with comments on its phylogenetic relationships. Ameghiniana. 33:373–376.
- Coria RA, Salgado L. 2000. A basal Abelisauria Novas, 1992 (Theropoda-Ceratosauria) from the Cretaceous of Patagonia, Argentina. GAIA. 15:89–102.
- Coria RA, Salgado L. 2005. Mid-Cretaceous turnover of saurischian dinosaur communities: evidence from the Neuquén Basin. Geol Soc London Spec Pub. 252(1):317–327. doi: 10.1144/GSL.SP.2005.252.01.16.
- Cruzado-Caballero P. 2017. New hadrosaurid remains from the Late Cretaceous of Río Negro Province (Argentina, Late Cretaceous). J Iberian Geol. 43(2):307–318. doi: 10.1007/s41513-017-0023-5.
- Cruzado-Caballero P, Coria RA. 2016. Revisiting the hadrosaurid (Dinosauria: Ornithopoda) diversity of the Allen Formation: a re-evaluation of Willinakaqe salitralensis from salitral Moreno, Río Negro province, Argentina. Ameghiniana. 53(2):231–237. doi: 10.5710/AMGH.25.09.2015.2943.
- Cruzado-Caballero P, Filippi LS, Méndez AH, Garrido AC, Díaz-Martínez I. 2018. First ornithopod remains from the Bajo de la Carpa Formation (Santonian, Upper Cretaceous), northern Patagonia, Argentina. Cret Res. 83:182–193. doi: 10.1016/j.cretres.2017.07.022.
- Cruzado-Caballero P, Gasca JM, Filippi LS, Cerda IA, Garrido AC. 2019. A new ornithopod dinosaur from the Santonian of Northern Patagonia (Rincón de Los Sauces, Argentina). Cret Res. 98:211–229. doi: 10.1016/j.cretres.2019.02.014.
- Cruzado-Caballero P, Powell J. 2017. Bonapartesaurus rionegrensis, a new hadrosaurine dinosaur from South America: implications for phylogenetic and biogeographic relations with North America. J Vert Palaeontol. 37(2):e1289381. doi: 10.1080/02724634.2017.1289381.
- Csiki-Sava Z, Brusatte SL, Ş V. 2016. “Megalosaurus cf. superbus” from southeastern Romania: The oldest known Cretaceous carcharodontosaurid (Dinosauria: Theropoda) and its implications for earliest Cretaceous Europe-Gondwana connections. Cret Res. 60:221–238. doi: 10.1016/j.cretres.2015.12.004.
- Cuesta E, Ortega F, Sanz JL. 2019. Axial osteology of Concavenator corcovatus (Theropoda; Carcharodontosauria) from the Lower Cretaceous of Spain. Cret Res. 95:106–120. doi: 10.1016/j.cretres.2018.10.026.
- Currie PJ, Zhao X-J. 1993. A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Can J Earth Sci. 30(10):2037–2081. doi: 10.1139/e93-179.
- Currie PJ, Zhao X-J. 1994. A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian J Earth Sci. 30(10–11):2037–2081. doi: 10.1139/e93-179.
- Da Silva OB, Caixeta JM, Milhomem PS, Kosin MD. 2007. Bacia do Recôncavo. Bol Geoci Petrobras. 15(2):423–431.
- Da Silva AJP, Lopes RDC, Vasconcelos AM, Bahia RBC. 2003. Bacias sedimentares Paleozóicas e Meso-Cenozóicas interiores. In: Bizzi L, Schobbenhaus C, Vidotti R, Gonçalves J, editors. Geologia, Tectônica e Recursos Minerais do Brasil: texto, mapas & SIG. Brasília: CPRM - Serviço Geológico do Brasil; p. 55–85.
- De Klerk WJ, Forster CA, Sampson SD, Chinsamy A, Ross CF. 2000. A new coelurosaurian dinosaur from the Early Cretaceous of South Africa. J Vert Palaeontol. 20(2):324–332. doi: 10.1671/0272-4634(2000)020[0324:ANCDFT]2.0.CO;2.
- Delcourt R. 2017. Revised morphology of Pycnonemosaurus nevesi Kellner & Campos, 2002 (Theropoda: Abelisauridae) and its phylogenetic relationships. Zootaxa. 4276(1):1–45. doi: 10.11646/zootaxa.4276.1.1.
- Delcourt R, Grillo ON. 2018. Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy. Palaeogeogr Palaeoclimatol Palaeoecol. 511:379–387. doi: 10.1016/j.palaeo.2018.09.003.
- D’Emic MD. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoo J Linn Soc. 166(3):624–671. doi: 10.1111/j.1096-3642.2012.00853.x.
- Depéret C. 1896. Note sur les dinosauriens sauropodes & théropodes du Crétacé supérieur de Madagascar. Bullde Soc Géol France. 3(24):176–194.
- Derby OA. 1878. A Bacia Cretáceo da Bahia. Arq Do Mus Nac. 3:1–135.
- De Souza GA, Soares MB, Weinschütz LC, Wilner E, Lopes RT, De Araújo OMO, Kellner AWA. 2021. The first edentulous ceratosaur from South America. Sci Rep. 11(1):22281. doi: 10.1038/s41598-021-01312-4.
- Destro N, Szatmari P, Alkmim FF, Magnavita LP. 2003. Release faults, associated structures, and their control on petroleum trends in the Recôncavo rift, northeast Brazil. AAPG Bull. 87(7):1123–1144. doi: 10.1306/02200300156.
- De Valais S, Apesteguía S, Udrizar Sauthier D. 2003. Nuevas evidencias de dinosaurios de la Formación Puerto Yeruá (CretácicoProvincia de Entre Ríos, Argentina). Ameghiniana. 40(3):502–507.
- Egli FB, Agnolín FL, Novas F. 2016. A new specimen of Velocisaurus unicus (Theropoda, Abelisauroidea) from the Paso Córdoba locality (Santonian), Río Negro, Argentina. J Vert Palaeontol. 36(4):e1119156. doi: 10.1080/02724634.2016.1119156.
- Evers SW, Rauhut OWM, Milner AC, McFeeters B, Allain R. 2015. A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco.Farke A, editor. PeerJ. 3:e1323. doi: 10.7717/peerj.1323.
- Ezcurra MD, Agnolín FL. 2012. A New Global Palaeobiogeographical Model for the Late Mesozoic and Early Tertiary. Syst Bio. 61(4):553–566. doi: 10.1093/sysbio/syr115.
- Fernández-Baldor FT, Canudo JI, Huerta P, Montero D, Suberbiola XP, Salgado L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontol Pol. 56(3):535–552. doi: 10.4202/app.2010.0003.
- Figueiredo FJD, Gallo V. 2021. Revision of Ellimmichthys longicostatus (Clupeomorpha: Ellimmichthyiformes) from the Lower Cretaceous of Brazil with comments on the taxonomy of related species. J South Am Earth Sci. 105:103006. doi: 10.1016/j.jsames.2020.103006.
- Filippi LS, Méndez AH, Juárez-Valieri RD, Garrido AC. 2016. A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids. Cret Res. 61:209–219. doi: 10.1016/j.cretres.2015.12.018.
- Fitzinger LJ. 1843. Systema Reptilium. Wien: Braumüller et Seidel. doi:10.5962/bhl.title.4694.
- Forster CA. 1990. The postcranial skeleton of the ornithopod dinosaur Tenontosaurus tilletti. J Vert Paleontol. 10(3):273–294. doi: 10.1080/02724634.1990.10011815.
- Forster CA, De Klerk WJ, Poole KE, Chinsamy-Turan A, Roberts EM, Ross CF. 2023. Iyuku raathi, a new iguanodontian dinosaur from the Early Cretaceous Kirkwood Formation, South Africa. Anat Rec. 306(7):1762–1803. doi: 10.1002/ar.25038.
- Forster CA, Farke AA, McCartney JA, De Klerk WJ, Ross CF. 2009. A “basal” tetanuran from the Lower Cretaceous Kirkwood Formation of South Africa. J Vert Palaeontol. 29(1):283–285. doi: 10.1671/039.029.0101.
- Forster CA, Sampson SD, Chiappe LM, Krause DW. 1998. The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar. Sci. 279(5358):1915–1919. doi: 10.1126/science.279.5358.1915.
- França TC, Brilhante NS, Nobre YDOM, Medeiros MAA, Lindoso RM, Rodrigues FC. 2022. The first record of a spinosaurid pedal ungual from Brazil (Boca do Forno Ravine, Itapecuru Formation, Parnaíba Basin). Hist Biol. 34(9):1817–1826. doi: 10.1080/08912963.2021.1981890.
- Franco-Rosas AC, Salgado L, Rosas CF, Carvalho IDS. 2004. Nuevos materiales de titanosaurios (Sauropoda) en el Cretácico superior de Mato Grosso, Brasil. Rev Bras Paleont. 7(3):329–336. doi: 10.4072/rbp.2004.3.04.
- Fronimos JA, Wilson JA, Baumiller TK. 2016. Polarity of concavo-convex intervertebral joints in the necks and tails of sauropod dinosaurs. Paleobiol. 42(4):624–642. doi: 10.1017/pab.2016.16.
- Gallina PA, Apesteguía SN, Haluza A, Canale JI, Farke AA. 2014. A Diplodocid Sauropod Survivor from the Early Cretaceous of South America. PloS One. 9(5):e97128. doi: 10.1371/journal.pone.0097128.
- Gallina PA, Canale JI, Carballido JL. 2021. The earliest known titanosaur sauropod dinosaur. Ameghiniana. 58(1):35–51. doi: 10.5710/AMGH.20.08.2020.3376.
- Gallo V. 1993. A “Coleção Cope” do Museu Nacional/UFRJ: Vertebrados fósseis da Bahia, Sergipe e Pernambuco (Brasil), e sua correlação estratigráfica [ Master thesis]. Instituto de Geociências, Universidade Federal do Rio de Janeiro; p. 173.
- Gallo V, Absolon BA, Figueiredo FJ. 2023. A “Coleção Cope” e os fósseis na Estrada de Ferro da Bahia-São Francisco. Filos Hist Biol. 18(1):73–95. doi: 10.11606/issn.2178-6224v18i1p73-95.
- Galloway A. 1999. Principles for interpretation of blunt force trauma: the biomechanics of fracture production. In: Galloway A, editor. Broken Bones: anthropological analysis of blunt force trauma. Springfield: Charles C. Thomas; p. 35–62.
- Galton PM. 1977. The ornithopod dinosaur Dryosaurus and a Laurasia–Gondwanaland connection in the Upper Jurassic. Nat. 268(5617):230–232. doi: 10.1038/268230a0.
- Galton PM. 1981. Dryosaurus, a hypsilophodontid dinosaur from the Upper Jurassic of North America and Africa postcranial skeleton. Palaontol Z. 55(3–4):271–312. doi: 10.1007/BF02988144.
- Galton PM, Powell H. 1980. The ornithischian dinosaur Camptosaurus prestwichii from the Upper Jurassic of England. Palaeontol. 23(2):411–443.
- García-Cobeña J, Cobos A, Verdú FJ. 2023. Ornithopod tracks and bones: paleoecology and an unusual evidence of quadrupedal locomotion in the Lower Cretaceous of eastern Iberia (Teruel, Spain). Cret Res. 144:105473. doi: 10.1016/j.cretres.2023.105473.
- Gasca J, Moreno-Azanza M, Ruiz-Omeñaca J, Canudo J. 2015. New material and phylogenetic position of the basal iguanodont dinosaur Delapparentia turolensis from the Barremian (Early Cretaceous) of Spain. J Iber Geol. 41(1):57–70. doi: 10.5209/rev_JIGE.2015.v41.n1.48655.
- Gasulla JM, Escaso F, Narváez I, Ortega F, Sanz JL, Claessens L. 2015. A new sail-backed styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PloS One. 10(12):e0144167. doi: 10.1371/journal.pone.0144167.
- Gauthier J. 1986. Saurischian monophyly and the origin of birds. Mem California Acad Sci. 8:1–55.
- Gerke O, Wings O, Lötters S. 2016. Multivariate and cladistic analyses of isolated teeth reveal sympatry of theropod dinosaurs in the Late Jurassic of Northern Germany. PloS One. 11(7):e0158334. doi: 10.1371/journal.pone.0158334.
- Ghilardi AM, Aureliano T, Duque RRC, Fernandes MA, Barreto AMF, Chinsamy A. 2016. A new titanosaur from the Lower Cretaceous of Brazil. Cret Res. 67:16–24. doi: 10.1016/j.cretres.2016.07.001.
- Gianechini FA, Apesteguia S. 2011. Unenlagiinae revisited: dromaeosaurid theropods from South América. An Acad Bras Ciênc. 83(1):163–195. doi: 10.1590/S0001-37652011000100009.
- Gilmore CW. 1909. Osteology of the Jurassic reptile Camptosaurus: with a revision of the species of the genus, and description of two new species. Proc US Nat Mus. 36(1666):197–332. doi: 10.5479/si.00963801.36-1666.197.
- Gilmore CW. 1915. Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance formation of Wyoming. Proc US Nat Mus. 49(2127):591–616. doi: 10.5479/si.00963801.49-2127.591.
- Gilpin D, DiCroce T, Carpenter K. 2007. A possible new basal hadrosaur from the Lower Cretaceous Cedar Mountain Formation of eastern Utah. In K Carpenter and and Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press; p. 79–89.
- Godefroit P, Codrea V, Weishampel DB. 2009. Osteology of Zalmoxes shqiperorum (Dinosauria, Ornithopoda), based on new specimens from the Upper Cretaceous of Nălaţ-Vad (Romania). Geodiversitas. 31(3):525–553. doi: 10.5252/g2009n3a3.
- Goloboff PA, Catalano SA. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics. 32(3):221–238. doi: 10.1111/cla.12160.
- Gomani EM. 2005. Sauropod dinosaurs from the early Cretaceous of Malawi, Africa. Palaeontol Elec. 8(1):1–37.
- González-Riga BJ, Mannion PD, Poropat SF, Ortiz-David LD, Coria JP. 2018. Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships. Zoo J Linn Soc. 184(1):136–181. doi: 10.1093/zoolinnean/zlx103.
- González-Riga BJ, Previtera E, Pirrone CA. 2009. Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Cret Res. 30(1):135–148. doi: 10.1016/j.cretres.2008.06.006.
- Gorscak E, O’Connor P. 2016. Time-calibrated models support congruency between Cretaceous continental rifting and titanosaurian evolutionary history. Biol Lett. 12(4):20151047. doi: 10.1098/rsbl.2015.1047.
- Gorscak E, O’Connor PM, Joger U. 2019. A new African titanosaurian sauropod dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, Southwestern Tanzania. PloS One. 14(2):e0211412. doi: 10.1371/journal.pone.0211412.
- Griffin CT, Bano LS, Turner AH, Smith ND, Irmis RB, Nesbitt SJ. 2019. Integrating gross morphology and bone histology to assess skeletal maturity in early dinosauromorphs: new insights from Dromomeron (Archosauria: Dinosauromorpha). PeerJ. 7:e6331. doi: 10.7717/peerj.6331.
- Griffin CT, Stocker MR, Colleary C, Stefanic CM, Lessner EJ, Riegler M, Formoso K, Koeller K, Nesbitt SJ. 2021. Assessing ontogenetic maturity in extinct saurian reptiles. Biol Rev. 96(1):470–525. doi: 10.1111/brv.12666.
- Han F-L, Barrett PM, Butler RJ, Xu X. 2012. Postcranial anatomy of Jeholosaurus shangyuanensis (Dinosauria, Ornithischia) from the Lower Cretaceous Yixian Formation of China. J Vert Palaeontol. 32(6):1370–1395. doi: 10.1080/02724634.2012.694385.
- Han F, Clark JM, Xu X, Sullivan C, Choiniere J, Hone DWE. 2011. Theropod teeth from the Middle-Upper Jurassic Shishugou Formation of northwest Xinjiang, China. J Vert Palaeontol. 31(1):111–126. doi: 10.1080/02724634.2011.546291.
- Han F, Forster CA, Xu X, Clark JM. 2018. Postcranial anatomy of Yinlong downsi (Dinosauria: Ceratopsia) from the Upper Jurassic Shishugou Formation of China and the phylogeny of basal ornithischians. J Syst Palaeontol. 16(14):1159–1187. doi: 10.1080/14772019.2017.1369185.
- Harris JD. 2006. The axial skeleton of the dinosaur Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontol. 49(5):1091–1121. doi: 10.1111/j.1475-4983.2006.00577.x.
- Harris JD, Dodson P. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontol Pol. 49:197–210.
- Hartt CF. 1870. On the geology of Brazil. J Am Geogr Stat Soc. 2(2):55. doi: 10.2307/196251.
- Hartt CF, Agassiz L. 1875. Geology and physical geography of Brazil: scientific results of a journey in Brazil by Louis Agassiz and his traveling companions. Huntington, New York: Krieger; p. 620.
- Hatcher JB. 1901. Diplodocus (Marsh): its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Mem Carnegie Mus. 1(1):1–63. doi: 10.5962/p.234818.
- Haughton SH. 1915. On some dinosaur remains from Bushmankand. Trans R Soc S Afr. 5(1):259–264. doi: 10.1080/00359191509519723.
- Haughton SH. 1928. On some reptilian remains from the Dinosaur Beds of Nyasaland. Trans R Soc South Afr. 16(1):67–75. doi: 10.1080/00359192809519658.
- Herne M, Nair J, Evans A, Tait A. 2019. New small-bodied ornithopods (Dinosauria, Neornithischia) from the Early Cretaceous Wonthaggi Formation (Strzelecki Group) of the Australian-Antarctic rift system, with revision of Qantassaurus intrepidus Rich and Vickers-Rich, 1999. J Paleontol. 93(3):543–584. doi: 10.1017/jpa.2018.95.
- Holland WJ. 1924. The skull of Diplodocus. Mem Carnegie Mus. 9(3):379–403. doi: 10.5962/p.234841.
- Hone DWE, Holtz TR Jr. 2017. A Century of Spinosaurs - A Review and Revision of the Spinosauridae with Comments on Their Ecology. Acta Geol Sin - English Edition. 91(3):1120–1132. doi: 10.1111/1755-6724.13328.
- Hone DWE, Xu X, Wang D-Y. 2010. A probable baryonychine (Theropoda: Spinosauridae) tooth from the Upper Cretaceous of Henan Province, China. Vert PalAs. 48(1):19–26.
- Hooley RW. 1925. On the skeleton of Iguanodon atherfieldensis sp. nov., from the Wealden Shales of Atherfield (Isle of Wight). Quart J Geol Soc London. 81(2):1–61. doi: 10.1144/GSL.JGS.1925.081.01-04.02.
- Huene von F. 1932. Die fossile Reptilordnung Saurischia: ihre Entwicklung und Geschichte. [The fossil reptile order Saurischia, their development and history]. Monographien zur Geologie und Palaeontologie s1. 4(1–2):1–361.
- Huene von F. 1934. Nuevos dientes de saurios del Cretáceo del Uruguay. Bol Inst Geol Uruguay. 21:13–20.
- Hulke JW. 1880. Iguanodon prestwichii, a new species from the Kimmeridge Clay, distinguished from I. mantelli of the Wealden Formation in the S.E. of England and Isle of Wight by Differences in the Shape of the Vertebral Centra, by fewer than five Sacral Vertebrœ, by the simpler Character of its Tooth-serrature, &·c., founded on numerous fossil remains lately discovered at Cumnor, near Oxford. Quart J Geol Soc London. 36(1–4):433–456. doi: 10.1144/GSL.JGS.1880.036.01-04.36.
- Hulke JW. 1882. XXIV. An attempt at a complete osteology of Hypsilophodon foxii; a British Wealden dinosaur. Phil Trans R Soc. 173:1035–1062. doi: 10.1098/rstl.1882.0025.
- Hutt S, Naish D, Martill DM, Barker MJ, Newbery P. 2001. A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cret Res. 22(2):227–242. doi: 10.1006/cres.2001.0252.
- Huxley TH. 1870. On Hypsilophodon Foxii, a new Dinosaurian from the Wealden of the Isle of Wight. Quart J Geol Soc London. 26(1–2):3–12. doi: 10.1144/GSL.JGS.1870.026.01-02.07.
- Ibiricu LM, Casal GA, Alvarez BN, De Sosa Tomas A, Lamanna MC, Cruzado-Caballero P. 2021. New hadrosaurid (Dinosauria: Ornithopoda) fossils from the uppermost Cretaceous of central Patagonia and the influence of paleoenvironment on South American hadrosaur distribution. J South Am Earth Sci. 110:103369. doi: 10.1016/j.jsames.2021.103369.
- Ibiricu LM, Casal GA, Lamanna MC, Martínez RD, Harris JD, Lacovara KJ. 2012. The southernmost records of Rebbachisauridae (Sauropoda: Diplodocoidea), from early Late Cretaceous deposits in central Patagonia. Cret Res. 34:220–232. doi: 10.1016/j.cretres.2011.11.003.
- Ibiricu LM, Casal GA, Martínez RD, Luna M, Canale JI, Álvarez BN, González Riga B. 2019. A new ornithopod dinosaur (Dinosauria: Ornithischia) from the Late Cretaceous of central Patagonia. Cret Res. 98:276–291. doi: 10.1016/j.cretres.2019.02.001.
- Ibiricu LM, Martínez RD, Lamanna MC, Casal GA, Luna M, Harris JD, Lacovara KJ. 2010. A medium-sized ornithopod (Dinosauria: Ornithischia) from the Upper Cretaceous Bajo Barreal Formation of Lago Colhué Huapi, Southern Chubut Province, Argentina. Ann Carnegie Mus. 79(1):39–50. doi: 10.2992/007.079.0103.
- Ibiricu LM, Martínez RD, Lamanna MC, Casal GA, Luna M, Harris J, Lacovara KJ. 2010. A Medium-Sized Ornithopod (Dinosauria: Ornithischia) from the Upper Cretaceous Bajo Barreal Formation of Lago Colhué Huapi, Southern Chubut Province, Argentina. Ann Carnegie Mus. 79(1):39–50. doi: 10.2992/007.079.0103.
- Ibiricu LM, Martínez RD, Luna A, Casal GA. 2014. A reappraisal of Notohypshilophodon comodorensis (Ornithischia: Ornithopoda) from the Late Cretaceous of Patagonia, Argentina. Zootaxa. 3786(4):401. doi: 10.11646/zootaxa.3786.4.1.
- Jain SL, Bandyopadhyay S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of central India. J Vert Paleontol. 17(1):114–136. doi: 10.1080/02724634.1997.10010958.
- Janensch W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten [Overview of the vertebrate fauna of the Tendaguru beds]. Archiv für Biontol. 3:81–110.
- Jiménez Gomis C, Cruzado Caballero P, Gasca JM, Filippi LS. 2018. Nuevos fósiles de dinosaurios ornitópodos en el Santoniense (Cretácico Superior) de la Formación Bajo de la Carpa del norte de Patagonia (Neuquén, Argentina). Geogaceta. 64:83–86.
- Juárez-Valieri R, Haro J, Fiorelli L, Calvo J. 2010. A new hadrosauroid (Dinosauria: Ornithopoda) from the Allen Formation (Late Cretaceous) of Patagonia, Argentina. Rev Mus Argent Cienc Nat N S. 12(2):217–231. doi: 10.22179/REVMACN.12.242.
- Kellner AWA. 1999. Short note on a new dinosaur (Theropoda, Coelurosauria) from the Santana Formation (Romualdo Member, Albian), northeastern Brazil. Bol Mus Nac Rio de Janeiro N S. 49:1–8.
- Kellner AWA, Azevedo SAK. 1999. A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. In: Tomida Y, Rich T, Vickers-Rich P, editors. Proceedings of the Second Gondwanan Dinosaur Symposium. National Science Museum Monographs, Tokyo; 15: p. 111–142.
- Kellner AWA, Azevedo SAK, Machado EB, Carvalho LBD, Henriques DDR. 2011. A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil. An Acad Bras Ciênc. 83(1):99–108. doi: 10.1590/S0001-37652011000100006.
- Kellner AWA, Campos DDA. 1996. First Early Cretaceous theropod dinosaur from Brazil with comments on Spinosauridae. N Jb Geol Paläont Abh. 199(2):151–166. doi: 10.1127/njgpa/199/1996/151.
- Kellner AWA, Campos DDA. 1999. Vertebrate paleontology in Brazil – a review. Episode. 22(3):238–251. doi: 10.18814/epiiugs/1999/v22i3/012.
- Kellner AWA, Campos DDA. 2000. Brief review of dinosaur studies and perspectives in Brazil. An Acad Bras Ciênc. 72(4):509–538. doi: 10.1590/S0001-37652000000400005.
- Kellner AWA, Campos DDA. 2002. On a theropod dinosaur (Abelisauria) from the continental Cretaceous of Brazil. Arq Mus Nac Rio de Janeiro. 60(3):163–170.
- Kellner AWA, Campos DDA, Azevedo SAK, Trotta MNF, Henriques DDR, Craik MMT, Silva HP. 2006. On a new titanosaur sauropod from the Bauru Group, Late Cretaceous of Brazil. Bol Mus Nac Rio de Janeiro N S Geol. 74:1–31.
- Kellner AWA, Campos DA, Trotta MN. 2005. Description of a titanosaurid caudal series from the Bauru Group, Late Cretaceous of Brazil. Arq Mus Nac. 63(3):529–564.
- Kirkland JI. 1998. A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: cretaceous) of eastern Utah - the oldest known hadrosaurid (lambeosaurine?). Bull N M Mus Nat Hist Sci. 14:283–295.
- Krumenacker L. 2017. Osteology, phylogeny, taphonomy, and ontogenetic histology of Oryctodromeus cubicularis, from the Middle Cretaceous (Albian-Cenomanian) of Montana and Idaho [ Doctoral thesis]. Montana, USA: Montana State University; p. 267.
- Lacerda MBS, De Andrade MB, Sales MAF, Aragão PRL, Vieira FS, Bittencourt JS, Liparini A. 2023. The vertebrate fossil record from the Feliz Deserto Formation (Lower Cretaceous), Sergipe, NE Brazil: paleoecological, taphonomic, and paleobiogeographic implications. Cret Res. 147:105463. doi: 10.1016/j.cretres.2022.105463.
- Lacovara K, Lamanna M, Ibiricu L, Poole JC, Schroeter ER, Ullmann PV, Voegele KK, Boles ZM, Carter AM, Fowler EK, et al. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from Southern Patagonia, Argentina. Sci Rep. 4(1):6196. doi:10.1038/srep06196.
- Langer MC, Martins NDO, Manzig PC, Ferreira GD, Marsola JDA, Fortes E, Lima R, Santana LCF, Vidal LDS, Lorençato RDS, et al. 2019. A new desert-dwelling dinosaur (Theropoda, Noasaurinae) from the Cretaceous of south Brazil. Sci Rep. 9(1):9379. doi: 10.1038/s41598-019-45306-9.
- Laurin M. 2004. The evolution of body size, Cope’s Rule and the origin of amniotes. Syst Bio. 53:594–622. doi: 10.1080/10635150490445706.
- Leardi JM, Pol D, Novas FE, Riglos MS. 2015. The postcranial anatomy of Yacarerani boliviensis and the phylogenetic significance of the notosuchian postcranial skeleton. J Vert Paleontol. 35(6):e995187. doi: 10.1080/02724634.2014.995187.
- Le Loeuff J. 1993. European titanosaurs. Rev Paléobiol. 7:105–117.
- Leonardi G. 1979. Nota preliminar sobre seis pistas de dinossauros Ornithischia da Bacia do Rio do Peixe (Cretáceo Inferior) em Sousa, Paraíba, Brasil. An Acad Bras Ciênc. 51(3):501–516.
- Leonardi G. 1989. Inventory and statistics of the South American dinosaurian ichnofauna and its paleobiological interpretation. In: Gillette D, Lockley M, editors. Dinosaur Tracks and Traces. Cambridge: Cambridge University Press; p. 165–178.
- Leonardi G. 1994. Annotated atlas of South America tetrapod footprints: devonian to Holocene: with an appendix on Mexico and Central America. Brasilía: Serviço Geológico do Brasil (CPRM); p. 248.
- Leonardi G, Borgomanero G. 1981. Sobre uma possÍvel ocorrência de ornithischia na formação Santana, Chapada do Araripe (Ceara). Rev Bras Geoc. 11(1):1–4. doi: 10.25249/0375-7536.198114.
- Leonardi G, Carvalho IDS. 2002. Jazigo icnofossilífero do Ouro-Araraquara (SP). In: Schobbenhaus C, Campos D, Queiroz E, Winge M, Berbert-Born M, editors. Sítios Geológicos e Paleontológicos do Brasil. Brasilía: Serviço Geológico do Brasil (CPRM) - Comissão Brasileira de Sítios Geológicos e Paleobiológicos - (SIGEP). p. 39–48.
- Leonardi G, Carvalho IDS. 2021. Dinosaur Tracks from Brazil - A Lost World of Gondwana. Bloomington: Indiana University Press; p. 462.
- Lindoso RM, Medeiros MAA, Carvalho IDS, Pereira AA, Mendes ID, Iori FV, Sousa EP, Arcanjo SHS, Silva TCM. 2019. A new rebbachisaurid (Sauropoda: Diplodocoidea) from the middle Cretaceous of northern Brazil. Cret Res. 104:104191. doi: 10.1016/j.cretres.2019.104191.
- Lull RS. 1919. The sauropod dinosaur Barosaurus Marsh. Mem Connecticut Acad Art Sci. VI:1–42.
- Luna M, Casal GA, Martínez RD, Lamanna MC, Ibiricu LM, Ivany E. 2003. La presencia de un Ornitopoda en el Miembro Superior de La Formación Bajo Barreal, Cretácico Superior (Campaniano-Maastrichtiano?) del Sur de Chubut. Ameghiniana. 40:61R.
- Lydekker R. 1888. Note on a new Wealden iguanodont and other dinosaurs. Quart J Geol Soc London. 44(1–4):46–61. doi: 10.1144/GSL.JGS.1888.044.01-04.08.
- Lydekker R. 1889. Notes on new and other dinosaurian remains. Geol Mag. 6(8):352–356. doi: 10.1017/S0016756800176587.
- Machado EB, Avilla LS, Nava WR, Campos DDA, Kellner AWA. 2013. A new titanosaur sauropod from the Late Cretaceous of Brazil. Zootaxa. 3701(3):301–321. doi: 10.11646/zootaxa.3701.3.1.
- Machado EB, Kellner AWA. 2005. Notas sobre Spinosauridae (Theropoda, Dinosauria). Anu Inst Geoc. 28(1):158–173. doi: 10.11137/2005_1_158-173.
- Machado EB, Kellner AWA. 2007. On a supposed ornithischian dinosaur from the Santana Formation, Araripe Basin, Brazil. In: Carvalho I, Cassab R, Schwanke C, Carvalho M, Fernandes A, Rodrigues M, De Carvalho M, Arai M, Oliveira M, editors. Palentologia: Cenários da Vida.Rio de Janeiro: Editora Interciência. p. 291–299.
- Madzia D, Boyd CA, Mazuch M. 2018. A basal ornithopod dinosaur from the Cenomanian of the Czech Republic. J Syst Palaeontol. 16(11):967–979. doi: 10.1080/14772019.2017.1371258.
- Maidment SCR, Chapelle KEJ, Bonsor JA, Button D, Barrett PM. 2022. Osteology and relationship of Cumnoria prestwichii (Ornithischia: Ornithopoda) from the Late Jurassic of Oxfordshire, UK. Palaeontogr Soc Monogr. 176(664):1–55. doi: 10.1080/02693445.2022.2162669.
- Malafaia E, Gasulla JM, Escaso F, Narváez I, Sanz JL, Ortega F. 2019. A new spinosaurid theropod (Dinosauria: Megalosauroidea) from the upper Barremian of Vallibona, Spain: Implications for spinosaurid diversity in the Early Cretaceous of the Iberian Peninsula. Cret Res. 106:104221. doi: 10.1016/j.cretres.2019.104221.
- Malafaia E, Mocho P, Escaso F, Dantas P, Ortega F. 2019. Carcharodontosaurian remains (Dinosauria, Theropoda) from the Upper Jurassic of Portugal. J Paleontol. 93(1):157–172. doi: 10.1017/jpa.2018.47.
- Malafaia E, Mocho P, Escaso F, Ortega F. 2017. New data on the anatomy of Torvosaurus and other remains of megalosauroid (Dinosauria, Theropoda) from the Upper Jurassic of Portugal. J Iber Geol. 43(1):33–59. doi: 10.1007/s41513-017-0003-9.
- Malafaia E, Mocho P, Escaso F, Ortega F. 2020. A new carcharodontosaurian theropod from the Lusitanian Basin: evidence of allosauroid sympatry in the European Late Jurassic. J Vert Paleontol. 1(1):e1768106. doi: 10.1080/02724634.2020.1768106.
- Mannion PD, Calvo JO. 2011. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics. Zoo J Linn Soc. 163:155–181. doi: 10.1111/j.1096-3642.2011.00699.x.
- Mannion PD, Upchurch P, Barnes RN, Mateus O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoo J Linn Soc. 168(1):98–206. doi: 10.1111/zoj.12029.
- Mannion PD, Upchurch P, Carrano MT, Barrett PM. 2011. Testing the effect of the rock record on diversity: a multidisciplinary approach to elucidating the generic richness of sauropodomorph dinosaurs through time. Biol Rev Camb Philos Soc. 86(1):157–181. doi: 10.1111/j.1469-185X.2010.00139.x.
- Mannion PD, Upchurch P, Schwarz D, Wings O. 2019. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zoo J Linn Soc. 2019:1–126.
- Marsh OC. 1869. Notice of some new reptilian remains from the Cretaceous of Brazil. Am J Sci. 2-47(141):390–392. doi: 10.2475/ajs.s2-47.141.390.
- Marsh OC. 1877. Notice of some new dinosaurian reptiles from the Jurassic Formation. Am J Sci. 3(14):514–516. doi: 10.2475/ajs.s3-14.84.514.
- Marsh OC. 1878. Principal characters of American Jurassic dinosaurs, part I. Am J Sci. 3(95):411–416. doi: 10.2475/ajs.s3-16.95.411.
- Marsh OC. 1879. Notice of new Jurassic reptiles. Am J Sci. 3(108):501–505. doi: 10.2475/ajs.s3-18.108.501.
- Marsh OC. 1881. Principal characters of American Jurassic dinosaurs, part V. Am J Sci. 3(125):417–423. doi: 10.2475/ajs.s3-21.125.417.
- Marsh OC. 1884. Principal characters of American Jurassic dinosaurs, part VII. On the Diplodocidae, a new family of the Sauropoda. Am J Sci. 3(27):160–168. doi: 10.2475/ajs.s3-27.160.329.
- Marsh OC. 1890. Description of new dinosaurian reptiles. Am J Sci. 3-39(229):81–86. doi: 10.2475/ajs.s3-39.229.81.
- Martill DM, Cruickshank ARI, Frey E, Small PG, Clarke M. 1996. A new crested maniraptoran dinosaur from the Santana Formation (Lower Cretaceous) of Brazil. J Geol Soc London. 153(1):5–8. doi: 10.1144/gsjgs.153.1.0005.
- Martin JE, Delfino M, Smith T. 2016. Osteology and affinities of Dollo’s goniopholidid (Mesoeucrocodylia) from the Early Cretaceous of Bernissart. Belgium. J Vertebr Paleontol. 36(6):e1222534. doi: 10.1080/02724634.2016.1222534.
- Martínez RD. 1998. Notohypsilophodon comodorensis gen. et. sp. nov., un Hypsilophodontidae (Ornithischia: Ornithopoda) del Cretácico Superior de Chubut, Patagonia central, Argentina. Acta Geol Leopold. 21(46/47):119–135.
- Martin JE, Lauprasert K, Buffetaut E, Liard R, Suteethorn V, Angielczyk K. 2014. A large pholidosaurid in the Phu Kradung Formation of north-eastern Thailand. Palaeontology. 57(4):757–769. doi: 10.1111/pala.12086.
- Martin JE, Sarr R, Hautier L. 2019. A dyrosaurid from the Paleocene of Senegal. J Paleontol. 93(2):343–358. doi: 10.1017/jpa.2018.77.
- Mateus O, Araújo R, Natário C, Castanhinha R. 2011. A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus. Zootaxa. 2827(1):54. doi: 10.11646/zootaxa.2827.1.3.
- Mateus O, Estraviz-López D. 2022. A new theropod dinosaur from the Early Cretaceous (Barremian) of Cabo Espichel, Portugal: implications for spinosaurid evolution. PloS One. 17(2):e0262614. doi: 10.1371/journal.pone.0262614.
- Mawson J, Woodward AS. 1907. On the Cretaceous Formation of Bahia (Brazil), and on vertebrate fossils collected therein. Quart J Geol Soc London. 63(1–4):128–. doi: 10.1144/GSL.JGS.1907.063.01-04.11.
- McDonald AT, Espilez E, Mampel L, Kirkland JI, Alcala L. 2012. An unusual new basal iguanodont (Dinosauria: Ornithopoda) from the Lower Cretaceous of Teruel, Spain. Zootaxa. 3595(2012):61–76. doi: 10.11646/zootaxa.3595.1.3.
- McDonald AT, Kirkland JI, DeBlieux DD, Madsen SK, Cavin J, Milner ARC, Panzarin L, Farke AA. 2010. New basal iguanodonts from the Cedar Mountain Formation of Utah and the evolution of thumb-spiked dinosaurs. PloS One. 5(11):e14075. doi: 10.1371/journal.pone.0014075.
- McIntosh JS. 1990. Sauropoda. In: Weishampel D, Dodson P, Ósmolska H, editors. The Dinosauria. Berkeley: University California Press; p. 345–401.
- McPhee BW, Mannion PD, de Klerk WJ, Choiniere JN. 2016. High diversity in the sauropod dinosaur fauna of the Lower Cretaceous Kirkwood Formation of South Africa: implications for the Jurassic–Cretaceous transition. Cret Res. 59:228–248. doi: 10.1016/j.cretres.2015.11.006.
- Medeiros M. 2006. Large theropod teeth from the Eocenomanian of northeastern Brazil and the occurrence of Spinosauridae. Rev Bras Paleontol. 9(3):333–338. doi: 10.4072/rbp.2006.3.08.
- Medeiros MAA, Arcanjo SHS, Lindoso RM, Carvalho IS, Pereira AA, Sousa EP, Mendes ID, Mendes DCS. 2019. Cenomanian vertebrates of the São Luís Basin. In: Teixeira S, Chaves C, editors. Contribuições à geologia da Amazônia. Belém, Brazil: Sociedade Brasileira de Geologia (Núcleo Norte); p. 179–187.
- Medeiros MAA, Freire PC, Pereira AA, Santos RAB, Lindoso RM, Coêlho AFA, Passos EB, Sousa E. 2007. Another African dinosaur recorded in the Eocenomanian of Brazil and a revision on the paleofauna of the Laje do Coringa site. In: Carvalho I, Cassab R, Schwanke C, Carvalho M, Fernandes A, Rodrigues M, De Carvalho M, Arai M, Oliveira M, editors. Palentologia: Cenários da Vida (1) Rio de Janeiro: Editora Interciência. p. 413–423.
- Medeiros MAA, Lindoso RM, Mendes ID, Carvalho IDS. 2014. The Cretaceous (Cenomanian) continental record of the Laje do Coringa flagstone (Alcântara Formation), northeastern South America. J South Am Earth Sci. 53:50–58. doi: 10.1016/j.jsames.2014.04.002.
- Medeiros MAA, Schultz CL. 2002. A fauna dinossauriana da “Laje do Coringa”, Cretáceo Médio do Nordeste do Brasil. Arq Mus Nac. 60(3):155–162.
- Medeiros MAA, Schultz CL. 2004. Rayososaurus (Sauropoda, Diplodocoidea) no meso-Cretáceo do Norte-Nordeste Brasileiro. Rev Bras Paleontol. 7(2):255–279. doi: 10.4072/rbp.2004.2.22.
- Méndez AH. 2014. The caudal vertebral series in abelisaurid dinosaurs. Act Palaeontol Pol. 59(1):99–107. doi: 10.4202/app.2011.0129.
- Milhomem PS, Milani EJ, Oliveira FM, Carvalho MSS, Souza-Lima W. 2003. Bacia do Recôncavo Bacias Sedimentares Brasileiras. Fundação Paleontológica Phoenix. 5(51):1–3.
- Mohabey DM, Sen S, Wilson JA. 2013. India’s first dinosaur, rediscovered. Curr Sci. 104(1):34–37.
- Molnar RE, Galton PM. 1986. Hypsilophodontid dinosaurs from Lightning Ridge, New South Wales, Australia. Geobios. 19(2):231–243. doi: 10.1016/S0016-6995(86)80046-8.
- Motta MJ, Aranciaga Rolando AM, Rozadilla S, Agnolín FE, Chimento NR, Brissón Egli F, Novas FE. 2016. New theropod fauna from the Upper Cretaceous (Huincul formation) of northwestern Patagonia, Argentina. New Mexico Mus Nat Hist Sci Bull. 71:231–253.
- Naish D, Cau A. 2022. The osteology and affinities of Eotyrannus lengi, a tyrannosauroid theropod from the Wealden Supergroup of southern England. PeerJ. 10:e12727. doi: 10.7717/peerj.12727.
- Naish D, Martill DM, Frey E. 2004. Ecology, systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil. Hist Biol. 16(2–4):57–70. doi: 10.1080/08912960410001674200.
- Navarro BA, Ghilardi AM, Aureliano T, Díez Díaz V, Bandeira KLN, Cattaruzzi AGS, Iori FV, Martine AM, Carvalho AB, Anelli LE, et al. 2022. A new nanoid titanosaur (Dinosauria: Sauropoda) from the Upper Cretaceous of Brazil. Ameghiniana. 59(5):317–354. doi: 10.5710/AMGH.25.08.2022.3477.
- Nopcsa BF. 1900. Dinosaurierreste aus Siebenbürgen (Schädel von Limnosaurus transsylvanicus nov. gen. et spec.) [Dinosaur remains from Transylvania (skull of Limnosaurus transsylvanicus nov. gen. et spec.)]. Denkschriften der Kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Classe. 68:555–591.
- Norell M, Makovicky PJ. 1997. Important features of the dromaeosaur skeleton: information from a new specimen. Am Mus Novit. 3215:1–45.
- Norman DB. 1980. On the ornithischian dinosaur Iguanodon bernissartensis from the Lower Cretaceous of Bernissart (Belgium). Bull Inst R Sci Nat Belg Sci Terre. 178:103.
- Norman DB. 1986. On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda). Bull Inst R Sci Nat Belg Sci Terre. 56:281–372.
- Norman DB. 2010. A taxonomy of iguanodontians (Dinosauria: Ornithopoda) from the lower Wealden Group (Cretaceous: Valanginian) of southern England. Zootaxa. 2489(1):47–66. doi: 10.11646/zootaxa.2489.1.3.
- Norman DB. 2011. On the osteology of the lower Wealden (Valanginian) ornithopod Barilium dawsoni (Iguanodontia: Styracosterna). Spec Pap Palaeontol. 86:165–194.
- Norman DB, Witmer LM, Weishampel DB. 2004a. Basal Ornithischia. In: Weishampel D, Dodson P, Osmólska H, editors. The Dinosauria. 2nd ed. Berkeley: University of California Press; p. 325–334.
- Norman DB, Witmer LM, Weishampel DB. 2004b. Basal Thyreophora. In: Weishampel D, Dodson P, Osmólska H, editors. The Dinosauria. 2nd ed. Berkeley: University of California Press; p. 335–342.
- Novas FE. 1997. Anatomy of Patagonykus puertai (Theropoda, Avialae, Alvarezsauridae), from the Late Cretaceous of Patagonia. J Vert Palaeontol. 17(1):137–166. doi: 10.1080/02724634.1997.10010959.
- Novas FE. 2009. The age of dinosaurs in South America. Bloomington: Indiana University Press; p. 367–368.
- Novas FE, Agnolin F, Bandyopadhyay S. 2004. Cretaceous theropods from India: a review of specimens described by Huene and Matley (1933). Rev Mus Argent Cienc Nat. 6(1):67–103. doi: 10.22179/REVMACN.6.74.
- Novas FE, Agnolin FL, Lio GL, Rozadilla S, Suárez M, de la Cruz R, de Souza Carvalho I, Rubilar-Rogers D, Isasi MP. 2021. New transitional fossil from late Jurassic of Chile sheds light on the origin of modern crocodiles. Sci Rep. 11(1):14960. doi: 10.1038/s41598-021-93994-z.
- Novas FE, Agnolin F, Rozadilla S, Aranciaga-Rolando A, Brissón-Eli F, Motta M, Cerroni M, Ezcurra M, Martinelli A, D´angelo J, et al. 2019. Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina. Rev Mus Argent Cienc Nat. 21:217–293. doi: 10.22179/REVMACN.21.655.
- Novas FE, Brissón Egli F, Agnolin FL, Gianechini FA, Cerda I. 2018. Postcranial osteology of a new specimen of Buitreraptor gonzalezorum (Theropoda, Unenlagiidae). Cret Res. 83:127–167. doi: 10.1016/j.cretres.2017.06.003.
- Novas FE, Cambiaso AV, Ambrioso A. 2004. A new basal iguanodontian (Dinosauria, Ornithischia) from the Upper Cretaceous of Patagonia. Ameghiniana. 41(1):75–82.
- Novas FE, de Valais S, Vickers-Rich P, Rich T. 2005. A large Cretaceous theropod from Patagonia, Argentina, and the evolution of carcharodontosaurids. Naturwissenschaften. 92(5):226–230. doi: 10.1007/s00114-005-0623-3.
- Ortega F, Escaso F, Sanz JL. 2010. A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature. 467(7312):203–206. doi: 10.1038/nature09181.
- Osborn HF. 1923. Two Lower Cretaceous dinosaurs from Mongolia. Am Mus Novit. 95:1–10.
- Ősi A, Prondvai E, Butler R, Weishampel DB, Evans AR. 2012. Phylogeny, Histology and Inferred Body Size Evolution in a New Rhabdodontid Dinosaur from the Late Cretaceous of Hungary. PloS One. 7(9):e44318. doi: 10.1371/journal.pone.0044318.
- Ostrom JH. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin Area, Wyoming and Montana. Bull Peabody Mus Nat Hist. 35:1–234. doi: 10.2307/j.ctvxkn7tk.
- Owen R. 1842. Report on British fossil reptiles. Part II. In: Report of the Eleventh Meeting of the British Association for the Advancement of Science Held at Plymouth in July 1841. London: John Murray; p. 60–204.
- Parméra TCC, Gallo V, Silva HMAD, Figueiredo FJD. 2019. Distributional patterns of Aptian-Albian paleoichthyofauna of Brazil and Africa based on Track Analysis. An Acad Bras Ciênc. 91(suppl 2):e20160456. doi: 10.1590/0001-3765201720160456.
- Paul GS. 1988. Predatory Dinosaurs of the World. New York: Simon & Schuster; p. 472.
- Paul GS. 2002. Dinosaurs of the Air. Baltimore: The Johns Hopkins University Press; p. 460.
- Pellegrini RA, Callahan WR, Hastings AK, Parris DC, McCauley JD. 2021. Skeletochronology and Paleohistology of Hyposaurus rogersii (Crocodyliformes, Dyrosauridae) from the Early Paleogene of New Jersey, USA. Animal. 11(11):3067. doi: 10.3390/ani11113067.
- Pereda-Suberbiola X, Galton PM, Mallison H, Novas F. 2013. A plated dinosaur (Ornithischia, Stegosauria) from the Early Cretaceous of Argentina, South America: an evaluation. Alcheringa. 37(1):65–78. doi: 10.1080/03115518.2012.702531.
- Pereira PVLGDC, Ribeiro TB, Brusatte SL, CRDA C, Marinho TDS, Bergqvist LP. 2020. Theropod (Dinosauria) diversity from the Açu Formation (mid-Cretaceous Potiguar Basin, Northeast Brazil. Cret Res. 114:104517. doi: 10.1016/j.cretres.2020.104517.
- Persons WI, Currie PJ. 2011. Dinosaur speed demon: The caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids. PloS One. 6(10):e25763. doi: 10.1371/journal.pone.0025763.
- Peterson OA, Gilmore CW. 1902. Elosaurus parvus: a new genus and species of the Sauropoda. Ann Carnegie Mus. 1(3):490–499. doi: 10.5962/p.78087.
- Pol D, Rauhut OWM. 2012. A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proc R Soc B. 279(1741):3170–3175. doi: 10.1098/rspb.2012.0660.
- Pol D, Rauhut OWM, Becerra M. 2011. A Middle Jurassic heterodontosaurid dinosaur from Patagonia and the evolution of heterodontosaurids. Naturwissenschaften. 98(5):369–379. doi: 10.1007/s00114-011-0780-5.
- Poropat SF, Kundrát M, Mannion PD, Upchurch P, Tischler TR, Elliott DA. 2021. Second specimen of the Late Cretaceous Australian sauropod dinosaur Diamantinasaurus matildae provides new anatomical information on the skull and neck of early titanosaurs. Zoo J Linn Soc. 192(2):610–674. doi: 10.1093/zoolinnean/zlaa173.
- Poropat SF, Nair JP, Syme CE, Mannion PD, Upchurch P, Hocknull SA, Cook AG, Tischler TR, Holland T. 2017. Reappraisal of Austrosaurus mckillopi Longman, 1933 from the Allaru Mudstone of Queensland, Australia’s first named Cretaceous sauropod dinosaur Alcheringa. Australas J Palaeontol. 41(4):543–580. doi: 10.1080/03115518.2017.1334826.
- Powell JE. 1987. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina part VI—the titanosaurids. Rev Mus Argent Cienc Nat. 3(3):147–153.
- Powell JE. 1992. Osteologia de Saltasaurus loricatus (Sauropoda-Titanosauridae) del Cretacico Superior del Noroeste argentino. In: Sanz J, Buscalioni A, editors. Los dinosaurios y su entorno biotico. Cuenca: Instituto Juan de Valdes, Serie Actas Academicas; p. 165–230.
- Powell JE. 2003. Revision of South American titanosaurid dinosaurs: paleobiological, palaeobiogeographical and phylogenetic aspects. Rec Queen Vict Mus. 111:1–173.
- Prates I, Fernandez R. 2015. Bacia do Recôncavo – Sumário geológico e setores em oferta. Brasília: Agência Nacional de Petroleo (ANP) - Superintendência de Definição de Blocos (SDB); p. 24.
- Prieto-Marquez A, Salinas GC. 2010. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. J Vert Palaeontol. 30(3):813–837. doi: 10.1080/02724631003763508.
- Rauhut OWM. 2011. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Spec Pap Palaeontol. 86:195–239. doi: 10.1111/j.1475-4983.2011.01084.x.
- Rauhut OWM, Carballido JL, Pol D. 2015. A diplodocid sauropod dinosaur from the Late Jurassic Cañadón Calcáreo Formation of Chubut, Argentina. J Vert Paleontol. 35(5):e982798. doi: 10.1080/02724634.2015.982798.
- Rauhut OWM, Carballido JL, Pol D. 2020. First osteological record of a stegosaur (Dinosauria, Ornithischia) from the Upper Jurassic of South America. J Vert Paleontol. 40(6):e1862133. doi: 10.1080/02724634.2020.1862133.
- Rauhut OWM, Carrano MT. 2016. The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania. Zoo J Linn Soc. 178(3):546–610. doi: 10.1111/zoj.12425.
- Rauhut OWM, Hübner T, Lanser K. 2016. A new megalosaurid theropod dinosaur from the late Middle Jurassic (Callovian) of north-western Germany: Implications for theropod evolution and faunal turnover in the Jurassic. Palaeontol Elec. 19(2):1–65. doi: 10.26879/654.
- Raven TJ, Barrett PM, Pond SB, Maidment SCR. 2020. Osteology and taxonomy of British Wealden Supergroup (Berriasian–Aptian) ankylosaurs (Ornithischia, Ankylosauria). J Vert Palaeontol. 40(4):e1826956. doi: 10.1080/02724634.2020.1826956.
- Remes K. 2006. Revision of the Tendaguru sauropod dinosaur Tornieria africana (Fraas) and its relevance for sauropod paleobiogeography. J Vert Paleontol. 26(3):651–669. doi: 10.1671/0272-4634(2006)26[651:ROTTSD]2.0.CO;2.
- Rich TH, Kear BP, Sinclair R, Chinner B, Carpenter K, McHugh ML, Vickers-Rich P. 2014. Serendipaceratops arthurcclarkei Rich & Vickers-Rich, 2003 is an Australian Early Cretaceous ceratopsian. Alcheringa: Australas J Palaeontol. 38(4):456–479. doi: 10.1080/03115518.2014.894809.
- Rich TH, Rich VP. 1989. Dinosaurs and biotas of the Early Cretaceous of Southeastern Australia. Nat Geogr Res. 5(1):15–53.
- Riguetti FJ, Apesteguía S, Pereda-Suberbiola X. 2022. A new Cretaceous thyreophoran from Patagonia supports a South American lineage of armoured dinosaurs. Sci Rep. 12(1):11621. doi: 10.1038/s41598-022-15535-6.
- Riguetti FJ, Pereda-Suberbiola X, Ponce D, Salgado L, Apesteguía S, Rozadilla S, Arbour V. 2022. A new small bodied ankylosaurian dinosaur from the Upper Cretaceous of North Patagonia (Río Negro Province, Argentina). J Syst Palaeontol. 20(1):2137441. doi: 10.1080/14772019.2022.2137441.
- Rodrigues T, Kellner A. 2010. Note on the pterosaur material described by Woodward from the Recôncavo Basin, Lower Cretaceous, Brazil. Rev Bras Paleontol. 13(2):159–164. doi: 10.4072/rbp.2010.2.08.
- Romer AS. 1956. Osteology of Reptiles. Chicago: University of Chicago Press; p. 772.
- Rozadilla S, Agnolín FL, Novas FE. 2019. Osteology of the Patagonian ornithopod Talenkauen santacrucensis (Dinosauria, Ornithischia). J Syst Palaeontol. 17(24):2043–2089. doi: 10.1080/14772019.2019.1582562.
- Rozadilla S, Agnolin FL, Novas FE, Aranciaga Rolando AM, Motta MJ, Lirio JM, Isasi MP. 2016. A new ornithopod (Dinosauria, Ornithischia) from the Upper Cretaceous of Antarctica and its palaeobiogeographical implications. Cret Res. 57:311–324. doi: 10.1016/j.cretres.2015.09.009.
- Rozadilla S, Egli FB, Agnolín FL, Aranciaga-Rolando AM, Novas FE. 2021. A new hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous of northern Patagonia and the radiation of South American hadrosaurids. J Syst Palaeontol. 19(17):1207–1235. doi: 10.1080/14772019.2021.2020917.
- Rozhdestvensky AK. 1966. Novye igyanodonti ie tsentrallinoy Asii. Phillogeneticheskye y taksonomicheskye veaimoothoshenia poednich Iguanodontidae y rannich Hadrosauridae [New iguanodonts from central Asia. Phylogenetic and taxonomic relationships of late Iguanodontidae and early Hadrosauridae]. Paleontol Zh. 1966(3):103–116.
- Rubilar-Rogers D, Soto-Acuña S, Otero RA, Yury-Yáñez R. 2013. First evidence of a dinosaur from Upper Cretaceous levels of the Dorotea Formation Sierra Baguales, Southernmost Chile. Geosu. R2013:345–348.
- Ruiz-Omeñaca JI, Canudo JI, Cruzado-Caballero P, Infante P, Moreno-Azanza M. 2005. Baryonychine teeth (Theropoda: Spinosauridae) from the Lower Cretaceous of La Cantalera (Josa, NE Spain). Kaupia. 14(5):59–63.
- Sales MAF, Liparini A, de Andrade MB, Aragão PRL, Schultz CL. 2017. The oldest South American occurrence of Spinosauridae (Dinosauria, Theropoda). J South Am Earth Sci. 74:83–88. doi: 10.1016/j.jsames.2016.10.005.
- Sales MAF, Schultz CL, Joger U. 2017. Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil. PloS One. 12(11):e0187070. doi: 10.1371/journal.pone.0187070.
- Salgado L, Azpilicueta C. 2000. A new saltasaurine dinosaur (Sauropoda, Titanosauridae) from the Rio Negro Province (Allen Formation, Upper Cretaceous), Patagonia, Argentina. Ameghiniana. 37(3):259–264.
- Salgado L, Canudo J, Garrido A, Moreno-Azanza M, Martínez LCA, Coria RA, Gasca JM. 2017. A new primitive neornithischian dinosaur from the Jurassic of Patagonia with gut contents. Sci Rep. 7(1):42778. doi: 10.1038/srep42778.
- Salgado L, Carvalho ISD. 2008. Uberabatitan ribeiroi, a new titanosaur from the Marília Formation (Bauru Group, Upper Cretaceous), Minas Gerais, Brazil. Palaeontol. 51(4):881–901. doi: 10.1111/j.1475-4983.2008.00781.x.
- Salgado L, Coria RA, Calvo JO. 1997. Evolution of titanosaurid sauropods: phylogenetic analysis based on the postcranial evidence. Ameghiniana. 34:3–32.
- Salgado L, García R. 2002. Variación morfológica en la secuencia de vértebras caudales de algunos saurópodos titanosaurios. Rev Española Paleontol. 17(2):211–216. doi: 10.7203/sjp.17.2.21568.
- Salgado L, Garrido A, Cocca SE, Cocca JR. 2004. Lower Cretaceous rebbachisaurid sauropods from Cerro Aguada Del León, Neuquén Province, northwestern Patagonia, Argentina. J Vert Paleontol. 24(4):903–912. doi: 10.1671/0272-4634(2004)024[0903:LCRSFC]2.0.CO;2.
- Salisbury SW, Frey E. 2001. A biomechanical transformation model for the evolution of semi-spheroidal articulations between adjoining vertebral bodies in crocodilians. In: Grigg G, Seebacher F, Franklin C, editors. Crocodilian Biology and Evolution. Chipping Norton: Surrey Beatty; p. 85–134.
- Samathi A, Chanthasit P, Sander P. 2019. Two new basal coelurosaurian theropod dinosaurs from the Early Cretaceous Sao Khua Formation of Thailand. Act Palaeontol Pol. 64(2):239–260. doi: 10.4202/app.00540.2018.
- Sampson SD, Carrano MT, Forster CA. 2001. A bizarre predatory dinosaur from the Late Cretaceous of Madagascar. Nature. 409(6819):504–506. doi: 10.1038/35054046.
- Sánchez-Hernández B, Benton MJ, Naish D. 2007. Dinosaurs and other fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NE Spain. Palaeogeo Palaeoclim Palaeoeco. 249(1–2):180–215. doi: 10.1016/j.palaeo.2007.01.009.
- Sander P, Tückmantel C. 2003. Bone lamina thickness, bone apposition rates, and age estimates in sauropod humeri and femora. Paläontol Z. 77(1):161–172. doi: 10.1007/BF03004566.
- Santucci RM, Arruda-Campos ACD. 2011. A new sauropod (Macronaria, Titanosauria) from the Adamantina Formation, Bauru Group, Upper Cretaceous of Brazil and the phylogenetic relationships of Aeolosaurini. Zootaxa. 3085(1):1–33. doi: 10.11646/zootaxa.3085.1.1.
- Sanz J, Powell J, Le Loeuff J, Martínez R, Pereda Suberbiola X. 1999. Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain) and the titanosaur phylogenetic relationships. Estud Mus Cienc Nat Alava. 14:235–255.
- Sayão JM, Saraiva AÁF, Brum AS, Bantim RAM, De Andrade RCLP, Cheng X, De Lima FJ, De Paula Silva H, Kellner AWA. 2020. The first theropod dinosaur (Coelurosauria, Theropoda) from the base of the Romualdo Formation (AlbianAraripe Basin, Northeast Brazil). Sci Rep. 10(1):10892. doi: 10.1038/s41598-020-67822-9.
- Scheetz RD. 1999. Osteology of Orodromeus makelai and the phylogeny of basal ornithopod dinosaurs [ Doctoral thesis]. Montana, USA: Montana State University; p. 186.
- Schobbenhaus C, Neves BBDB. 2003. A geologia do Brasil no contexto da Plataforma Sul-Americana. In: Bizzi L, Schobbenhaus C, Vidotti R, Gonçalves J, editors. Geologia, Tectônica e Recursos Minerais do Brasil: texto, mapas & SIG. Brasília: CPRM - Serviço Geológico do Brasil; p. 5–25.
- Schwarz-Wings D, Frey E, Martin T. 2009. Reconstruction of the bracing system of the trunk and tail in hyposaurine dyrosaurids (Crocodylomorpha; Mesoeucrocodylia. J Vert Paleontol. 29(2):453–472. doi: 10.1671/039.029.0228.
- Seeley HG. 1888. I. On the classification of the fossil animals commonly named Dinosauria. Proc R Soc Lond. 43(258–265):165–171. doi: 10.1098/rspl.1887.0117.
- Sereno PC. 1986. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). Nat Geogr Res. 2:234–256.
- Sereno PC, Beck AL, Dutheil DB, Gado B, Larsson HCE, Lyon GH, Marcot JD, Rauhut OWM, Sadleir RW, Sidor CA, et al. 1998. A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science. 282(5392):1298–1302. doi: 10.1126/science.282.5392.1298.
- Sereno PC, Brusatte SL. 2008. Basal Abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger. Acta Palaeontol Pol. 53(1):15–46. doi: 10.4202/app.2008.0102.
- Sereno PC, Chao S. 1988. Psittacosaurus xinjiangensis (Ornithischia: Ceratopsia), a new psittacosaur from the Lower Cretaceous of northwestern China. J Vert Paleontol. 8(4):353–365. doi: 10.1080/02724634.1988.10011724.
- Sereno PC, Dutheil DB, Iarochene M, Larsson HCE, Lyon GH, Magwene PM, Sidor CA, Varricchio DJ, Wilson JA. 1996. Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science. 272(5264):986–991. doi: 10.1126/science.272.5264.986.
- Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, Larsson HCE, Kemp T. 2008. Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PloS One. 3(9):e3303. doi: 10.1371/journal.pone.0003303.
- Sereno PC, Wilson JA. 2005. Structure and evolution of a sauropod tooth battery. In: Curry-Rogers K, Wilson J, editors. The sauropods: evolution and paleobiology. Berkeley: University of California Press; p. 157–177.
- Serrano-Martínez A, Ortega F, Sciscio L, Tent-Manclús JE, Fierro Bandera I, Knoll F. 2015. New theropod remains from the Tiourarén Formation (?Middle Jurassic, Niger) and their bearing on the dental evolution in basal tetanurans. Proc Geol Assoc. 126(1):107–118. doi: 10.1016/j.pgeola.2014.10.005.
- Shipman P. 1981. Life History of a Fossil: an Introduction to Taphonomy and Paleoecology. Cambridge: Harvard University Press; p. 22.
- Silva JCG Jr, Marinho TS, Martinelli AG, Langer MC. 2019. Osteology and systematics of Uberabatitan ribeiroi (Dinosauria: Sauropoda): a Late Cretaceous titanosaur from Minas Gerais, Brazil. Zootaxa. 4577(3):401–438. doi: 10.11646/zootaxa.4577.3.1.
- Soto-Acuña S, Vargas AO, Kaluza J, Leppe MA, Botelho JF, Palma-Liberona J, Simon-Gutstein C, Fernández RA, Ortiz H, Milla V, et al. 2021. Bizarre tail weaponry in a transitional ankylosaur from subantarctic Chile. Nature. 600(7888):259–263. doi: 10.1038/s41586-021-04147-1.
- Soto M, Perea D, Cambiaso A. 2012. First sauropod (Dinosauria: Saurischia) remains from the Guichón Formation, Late Cretaceous of Uruguay. J South Am Earth Sci. 33(1):68–79. doi: 10.1016/j.jsames.2011.08.003.
- Souza LG. 2019. Systematic revision of the postcranial bones from crown Crocodylia group (Crocodyliformes: Eusuchia) and its significance for fossil record. [ Doctoral thesis]. Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro; p. 588.
- Souza RGD, Campos DDA. 2018. New Crocodyliform specimens from Recôncavo-Tucano Basin (Early Cretaceous) of Bahia, Brazil. An Acad Bras Ciênc. 91(2):e20170382. doi: 10.1590/0001-3765201720170382.
- Souza RGD, Figueiredo RG, Azevedo SAK, Riff D, Kellner AWA. 2019. Systematic revision of Sarcosuchus hartti (Crocodyliformes) from the Recôncavo Basin (Early Cretaceous) of Bahia, north-eastern Brazil. Zoo J Linn Soc. 188(2):552–578. doi: 10.1093/zoolinnean/zlz057.
- Souza JD, Kosin M, Teixeira LR, Martins AAM, Bento RV, Borges VP, Leite CA, Arcanjo JB. 2004. Folha Salvador SD-24. In: Schobbenhaus C, Gonçalves J, Santos J, Abram M, Leão NR, Matos G, Vidotti R, Ramos M, and Jesus J, editors. Carta Geológica do Brasil ao Milionésimo, Sistema de Informações Geográficas - SIG, Programa Geologia do Brasil. Brasília: Serviço Geológico do Brasil - Companhia de Pesquisa em Recursos Minerais (CPRM).
- Souza RGD, Riff D, Kellner AWA. 2015. Taxonomic revision of Thoracosaurus bahiensis Marsh, 1869, a supposed Gavialoidea (Reptilia, Crocodylia) from Cretaceous deposits of the Recôncavo Basin, Brazil. Rev Bras Paleontol. 18(3):565–568. doi: 10.4072/rbp.2015.3.17.
- Stovall JW, Langston W. 1950. Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. Am Midland Nat. 43(3):696–728. doi: 10.2307/2421859.
- Stromer E. 1915. Das original des theropoden Spinosaurus aegyptiacus nov. gen., nov. spec. Mathematisch-physikalische Klasse Abhandlung. 28:1–31.
- Sues H-D, Evans DC, Galton PM, Brown CM. 2023. Anatomy of the neornithischian dinosaur Parksosaurus warreni (Parks, 1926) from the Upper Cretaceous (lower Maastrichtian) Horseshoe Canyon Formation of Alberta, Canada. Cret Res. 141:105369. doi: 10.1016/j.cretres.2022.105369.
- Sues H-D, Frey E, Martill DM, Scott DM. 2002. Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil. J Vert Palaeontol. 22(3):535–547. doi: 10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- Tapia A. 1918. Una mandibula de dinosaurio procedente de Patagonia. Physis. 4:369–370.
- Taquet P. 1972. Il giacimento di Gadoufaoua [The locality of Gadoufaoua]. I Dinosauri del Teneré. 41:51–60.
- Tennant JP, Mannion PD, Upchurch P, Sutton MD, Price GD. 2017. Biotic and environmental dynamics through the Late Jurassic–Early Cretaceous transition: evidence for protracted faunal and ecological turnover. Biol Rev. 92(2):776–814. doi: 10.1111/brv.12255.
- Tschopp E, Mateus O, Benson RBJ. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ. 3:e857. doi: 10.7717/peerj.857.
- Tumarkin‐Deratzian AR. 2009. Evaluation of long bone surface textures as ontogenetic indicators in centrosaurine ceratopsids. Anat Rec. 292(9):1485–1500. doi: 10.1002/ar.20972.
- Tumarkin-Deratzian AR, Vann DR, Dodson P. 2006. Bone surface texture as an ontogenetic indicator in long bones of the Canada goose Branta canadensis (Anseriformes: Anatidae). Zoo J Linn Soc. 148(2):133–168. doi: 10.1111/j.1096-3642.2006.00232.x.
- Upchurch P, Barrett PM, Dodson P. 2004. Sauropoda. In: Weishampel D, Dodson P, Osmolska H, editors. The Dinosauria. 2nd ed. Berkeley: University of California Press; p. 259–322.
- Upchurch P, Hunn CA, Norman DB. 2002. An analysis of dinosaurian biogeography: evidence for the existence of vicariance and dispersal patterns caused by geological events. Proc R Soc B. 269(1491):613–621. doi: 10.1098/rspb.2001.1921.
- Upchurch P, Mannion PD, Barrett PM. 2011. Sauropod dinosaurs. In: Batten D, editor. Field guide to English Wealden fossils. London: Palaeontological Association; p. 476–525.
- Veloso RL, Bandeira KLN, Souza L, Gallo V, Carvalho LB. 2023. Derby Hyposaurus: a historical review and update of its whereabouts. Hist Biol. 35(6). doi: 10.1080/08912963.2022.2073546.
- Veloso RL, Bandeira KLN, Souza L, Gallo V, De Carvalho LB. 2023. Derby Hyposaurus: a historical review and update of its whereabouts. Hist Biol. 35(6):1002–1010. doi: 10.1080/08912963.2022.2073546.
- Verdú FJ, Royo-Torres R, Cobos A, Alcalá L. 2015. Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain). Cret Res. 56:250–264. doi: 10.1016/j.cretres.2015.05.010.
- Viana CF, Gama Junior EG, Simões IDA, Moura JA, Fonseca JR, Alves RJ. 1971. Revisão estratigráfica da Bacia do Reconcavo. Bol Téc Petrobras. 14(3/4):157–192.
- Wang X, Bandeira KLN, Qiu R, Jiang S, Cheng X, Ma Y, Kellner AWA. 2021. The first dinosaurs from the Early Cretaceous Hami Pterosaur Fauna, China. Sci Rep. 11(1):14962. doi: 10.1038/s41598-021-94273-7.
- Wang X, Pan R, Butler RJ, Barrett PM. 2010. The postcranial skeleton of the iguanodontian ornithopod Jinzhousaurus yangi from the Lower Cretaceous Yixian Formation of western Liaoning, China. Earth Environ Sci Trans R Soc Edinb. 101(2):135–159. doi: 10.1017/S1755691010009266.
- Weishampel DB, Björk PR. 1989. The first indisputable remains of Iguanodon (Ornithischia: Ornithopoda) from North America: Iguanodon lakotaensis, sp. nov. J Vert Palaeontol. 9(1):56–66. doi: 10.1080/02724634.1989.10011738.
- Weishampel DB, Jianu CM, Csiki Z, Norman DB. 2003. Osteology and phylogeny of Zalmoxes (ng), an unusual euornithopod dinosaur from the latest Cretaceous of Romania. J Syst Palaeontol. 1(2):65–123. doi: 10.1017/S1477201903001032.
- White MA, Benson RBJ, Tischler TR, Hocknull SA, Cook AG, Barnes DG, Poropat SF, Wooldridge SJ, Sloan T, Sinapius GHK, et al. 2013. New Australovenator hind limb elements pertaining to the holotype reveal the most complete neovenatorid leg. PloS One. 8(7):e68649. doi:10.1371/journal.pone.0068649.
- Whitlock JA. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoo J Linn Soc. 161(4):872–915. doi: 10.1111/j.1096-3642.2010.00665.x.
- Wilson JA. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoo J Linn Soc. 136(2):215–275. doi: 10.1046/j.1096-3642.2002.00029.x.
- Wilson JA, D’Emic MD, Ikejiri T, Moacdieh EM, Whitlock JA, Farke A. 2011. A Nomenclature for Vertebral Fossae in Sauropods and Other Saurischian Dinosaurs. PloS One. 6(2):e17114. doi: 10.1371/journal.pone.0017114.
- Wilson JA, Sereno PC. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. J Vert Paleontol. 18(sup002):1–79. doi: 10.1080/02724634.1998.10011115.
- Wilson JA, Upchurch P. 2003. A revision of Titanosaurus Lydekker (dinosauria ‐ sauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution. J Syst Palaeontol. 1(3):125–160. doi: 10.1017/S1477201903001044.
- Woodward AS. 1888. XIV. Notes on some vertebrate fossils from the Province of Bahia, Brazil, collected by Joseph Mawson. Ann Mag Nat Hist. 2(8):132–136. doi: 10.1080/00222938809460891.
- Woodward AS. 1891. Evidence of the occurrence of pterosaurians and plesiosaurians in the Cretaceous of Brazil, discovered by Joseph Mawson, Esq., FGS. Ann Mag Nat Hist. 6(8):314–317. doi: 10.1080/00222939008694040.
- Xu X, Wang X-L, You H-L. 2000. A primitive ornithopod from the Early Cretaceous Yixian Formation of Liaoning. Vert PalAs. 38(4):318–325.
- You H-L, Ji Q, Li D. 2005. Lanzhousaurus magnidens gen. et sp. nov. from Gansu Province, China: the largest-toothed herbivorous dinosaur in the world. Geol Bull China. 24:785–794.
- You H-L, Luo Z-X, Shubin NH, Witmer LM, Tang ZL, Tang F. 2003. The earliest-known duck-billed dinosaur from deposits of late Early Cretaceous age in northwest China and hadrosaur evolution. Cret Res. 24(3):347–355. doi: 10.1016/S0195-6671(03)00048-X.
- Zaher H, Pol D, Carvalho AB, Nascimento PM, Riccomini C, Larson P, Juarez-Valieri R, Pires-Domingues R, Da Silva NJ, Campos DDA, et al. 2011. A complete skull of an early Cretaceous sauropod and the evolution of advanced titanosaurians. PloS One. 6(2):e16663. doi:10.1371/journal.pone.0016663.
- Zaher H, Pol D, Navarro BA, Delcourt R, Carvalho AB. 2020. An Early Cretaceous theropod dinosaur from Brazil sheds light on the cranial evolution of the Abelisauridae. C. R. Palevol. 6(6):101–115. doi: 10.5852/cr-palevol2020v19a6.
- Zan SQ, Chen J, Jin LY, Li T. 2005. A primitive ornithopod from the Early Cretaceous Quantou Formation of Central Jilin, China. Vert PalA. 43(3):182–193.
- Zheng W, Jin X, Shibata M, Azuma Y, Yu F. 2012. A new ornithischian dinosaur from the Cretaceous Liangtoutang Formation of Tiantai, Zhejiang Province, China. Cret Res. 34:208–219. doi: 10.1016/j.cretres.2011.11.001.
- Zurriaguz V, Cerda I. 2017. Caudal pneumaticity in derived titanosaurs (Dinosauria: Sauropoda). Cret Res. 73:14–24. doi: 10.1016/j.cretres.2017.01.001.