
ABSTRACT
Knowledge of the Campanian-Maastrichtian titanosaurian sauropods from southern South America has increased markedly in recent years. Several taxa have been reported from northern Patagonia, as well as few taxa from southern Patagonia that were described in the last years. However, titanosaurs from the end-Cretaceous of Central Patagonia are poorly known. Here we report the associated remains of a new species of a titanosaurian sauropod from the La Colonia Formation found in northern Chubut Province. Titanomachya gimenezi gen. et sp. nov. is represented by a caudal vertebra and appendicular elements. Titanomachya, is interpreted as a member of Lithostrotia, sharing derived features with saltasaurids and bearing unique characteristics in the astragalus. The astragalus morphology of Titanomachya display an intermediate form between Colossosauria and Saltasauroidea, allowing articulation between zeugopodium and autopodium with contributions from both the tibia and fibula. The type specimen is a small-sized adult, estimated at 5.8–9.8 tons. Comparative analyses reveal distinct sauropod faunas in different Patagonian regions and Brazilian formations during the Campanian – Maastrichtian. In northern Patagonia, aeolosaurines and saltasaurines predominate, while in southern Patagonia predominate colossosaurians and other large titanosaurs. The less well-known sauropod fauna from central Patagonia includes aeolosaurines, eutitanosaurs, and the addition of small-bodied saltasauroids such as Titanomachya.
Introduction
Titanosaurian sauropods were diverse in the latest Cretaceous (Campanian – Maastrichtian) ecosystems of Patagonia (Santucci and Filippi Citation2022). The current fossil record is heavily biased towards the better sampled Neuquén Group in northern Patagonia, where two major titanosaur lineages are recorded, Colossosauria and Saltasauroidea. The fossil record in southern Patagonia (Austral Basin) includes colossosaurian titanosaurs, such as Puertasaurus reuili Novas et al. Citation2005 (Cerro Fortaleza Formation, Campanian) and possibly Nullotitan glaciaris Novas et al. Citation2019 (Chorrillo Formation, Campanian – Maastrichtian), as well as the large saltasauroid Dreadnoughtus schrani Lacovara et al. Citation2014 (Cerro Fortaleza Formation).
In Central Patagonia, the colossosaurian Aeolosaurus colhuehuapensis Casal et al. Citation2007 (Lago Colhue Huapi Formation, Campanian – Maastrichtian) is one of the few named titanosaur taxa from the latest Cretaceous. Other findings in the same formation include the eutitanosaurs Argyrosaurus superbus (Lydekker Citation1893) and Elaltitan lilloi (Mannion and Otero Citation2012). However, recent studies indicate that Elaltitan comes from lower levels in this formation (= Coniacian) and is not contemporaneous with Argyrosaurus (Casal et al. Citation2016).
Previous studies had already noted the presence of fragmentary remains of saltasauroids from the Maastrichtian of La Colonia Formation in northern Chubut Province. These remains were collected in the Cerro Bayo Norte area (Gasparini et al. Citation2015; Sterli et al. Citation2022; ) and consist of an anterior caudal vertebra (MPEF-PV 10827) and a posterior caudal centrum (MPEF-PV 10828). Recent fieldwork in this area has yielded associated and partially articulated remains of a more complete titanosaur specimen from the La Colonia Formation that are described here ().
Figure 1. Location map of the site of the discovery of the holotype of Titanomachya gimenezi. Modified from Sterli et al. (Citation2022)
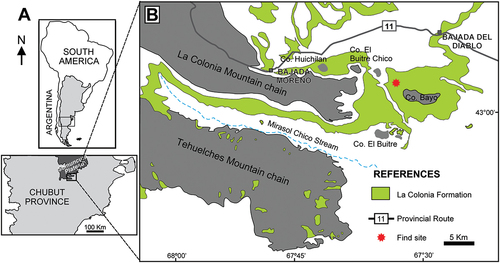
Figure 2. Geological columns of the La Colonia Formation, at Norte de Cerro Bayo, the area of the discovery of the holotype of Titanomachya gimenezi. Modified from Sterli et al. (Citation2022)
The new specimen represents a new saltasauroid taxon, the first to be recognised from Central Patagonia for the end-Cretaceous. These remains are described in this work and are the basis for the erection of a new species, Titanomachya gimenezi (). These materials confirm the presence of saltasauroids in the La Colonia Formation. Along with Dreadnoughtus, they are, up to now, the only two saltasauroids recorded south of the North Patagonian Massif.
Figure 3. Skeletal reconstruction of Titanomachya gimenezi. Preserved bones shown in light blue. Reconstruction by Gabriel Lio.

Institutional abbreviations
IANIGLA, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Colección de Paleo- vertebrados, Mendoza, Argentina; MACN, Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’, Ciudad Autónoma de Buenos Aires, Argentina; MAU-PV- (MRS), Vertebrate Paleontology collection, Museo ‘Argentino Urquiza’, Rincón de Los Sauces, Neuquén, Argentina; MLP, Museo de La Plata, La Plata, Argentina; MPCA, Muse Provincial ‘Carlos Ameghino’, Cipolletti, Río Negro, Argentina; MPEF-PV, Vertebrate Paleontology collection, Museo Paleontológico ‘Egidio Feruglio’ Trelew, Chubut, Argentina; MUCPv, Vertebrate Paleontology collection, Museo de Geología y Paleontología de la Universidad Nacional del Comahue, Neuquén, Argentina; PVL, Instituto ‘Miguel Lillo’, Tucumán, Argentina; UNCUYO-LD, Universidad Nacional de Cuyo, Laboratorio y Museo de Dinosaurios, Mendoza, Argentina; UNPSJB, Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina.
Anatomical abbreviations
4t, fourth trochanter; asp, ascending process; cc, cnemial crest; cf, cnemial fossa; dc, deltopectoral crest; fh, femoral head; fs, fibular surface; gt, greater trochanter; hb, hemispherical bulge; hh, humeral head; isped, ischial peduncle; lb, lateral bulge; lc, lateral condyle; ld, latissimus dorsi insertion scar; lec, lateral epicondyle; lt, lateral tuberosity; mc, medial condyle; pc, posterior condyle; poap, postacetabular process; pr, posterior ridge; scf, supracondylar fossa; suc, supracoracoid insertion scar; ts, tibial surface
Materials and methods
Description nomenclature
Robustness Index (RI) was calculated as the average of the greatest widths of the proximal end, mid-shaft and distal end of the element/length of the element (Wilson and Upchurch Citation2003).
Osteological comparisons were made mainly with sauropods from the Campanian−Maastrichtian of Patagonia, such as Rocasaurus muniozi, Aeolosaurus rionegrinus, Bonatitan reigi, Menucocelsior arriagadai (Salgado and Azpilicueta Citation2000; Powell Citation1987; Martinelli and Forasiepi Citation2004; Aranciaga Rolando et al. Citation2022; Allen Formation, Río Negro, North Patagonia), the specimen MPCA−Pv 862 from Salitral Moreno (Allen Formation), Aeolosaurus colhuehuapensis (Colhue Huapi Formation, Chubut, Central Patagonia), Dreadnoughtus (Cerro Fortaleza Formation, Santa Cruz, South Patagonia), and Nullotitan (Chorrillo Formation, Santa Cruz, South Patagonia), as well as with other phylogenetically related sauropods, according to the present analysis, such as Saltasaurus loricatus, Uberabatitan riberoi, Baurutitan britoi, Yamanasaurus lojaensis (Bonaparte and Powell Citation1980; Kellner et al. Citation2005; Salgado and Carvalho Citation2008; Apesteguía et al. Citation2020), which comes also from Maastrichtian levels.
Phylogenetic analysis
In order to test the phylogenetic affinities of Titanomachya gimenezi, an equally weighted parsimony analysis was carried out. We used the matrix of Pérez Moreno et al. (Citation2023), consisting of 432 characters and 103 taxa, with some modifications (see Supplementary Information). We conducted the analysis using the ‘New Technology Search’, using the command ‘xmult = hits 100’. Under this command, the New Technology Search algorithms (‘Sectorial Search’, ‘Ratchet’, ‘Drift’ and ‘Tree Fusing’) are applied together with the traditional search procedures, such as Wagner Trees, Tree Branch Reconnection (TBR) and Subtree Pruning- Regrafting (SPR) algorithms, to find the Minimum Length Trees (MLTs). The trees obtained were subjected to another round of TBR, which resulted in trees that exceeded the memory limit (>400,000 MPTs found). Node supports for the MPT’s were calculated using Bootstrap and Jackknife (Felsenstein Citation1985; Farris et al. Citation1996), and Bremer Support (=Decay Index; Bremer Citation1994). Additionally, the effect of unstable taxa in support values was explored using the pcrjak script for TNT (Pol and Goloboff Citation2020) to assess if low support values were caused by the inclusion of fragmentary taxa, such as Titanomachya. The resampling support analyses consisted in 1000 pseudo-replicates in TNT with the same tree search strategy employed for the parsimony analysis, in which the resulting trees were summarised through absolute frequencies. Bremer support was calculated using the TNT script bremsup.run that combines heuristic searches of suboptimal trees allied to tree searches under negative constraints. The obtained topologies were processed in FigTree v1.3.1.
Body mass estimation
We estimated the body mass of Titanomachya using the quadratic equations for quadrupeds proposed by Campione (Citation2017). All analyses were carried out in R, version 4.0.3 with functions from xlsx 0.5.7, MASSTIMATE 1.2, qpcR 1.4–0, ape 3.3, Geiger 2.0.6, and nlme 3.1–122 (Paradis Citation2012; Adrian Citation2014; Andrej-Nikolai Citation2014; Campione et al. Citation2014; Pennell et al. Citation2014; Pinheiro et al. Citation2015). Quadratic equations for quadrupeds and bipeds are integrated into the MASSTIMATE 1.3 QE and cQE functions, respectively, using the argument ‘quadratic¼’ (see Campione Citation2017 Supplementary Material for full R script).
Systematic palaeontology
DINOSAURIA Owen, 1842
SAUROPODA Marsh, Citation1878
NEOSAUROPODA Bonaparte, Citation1986
TITANOSAURIFORMES Salgado, Coria, and Calvo, Citation1997
TITANOSAURIA Bonaparte and Coria, Citation1993
TITANOMACHYA GIMENEZI gen. et sp. nov.
Derivation of name
The Titanomachy, in Greek mythology, is the battle fought by the Olympians against the Titans, in which the latter are defeated. Titanomachya honours that battle since it comes from the time when the titanosaurs became extinct. The specific epithet gimenezi is in honour of the late Dr. Olga Giménez, who was the first female palaeontologist that studied the dinosaurs from Chubut Province.
Diagnosis
Sauropod characterised by the following unique combination of characters and autapomorphies (marked with an asterisk): haemal canal approximately 25% of the total length of the chevron; humeral deltopectoral attachment with prominent development; humeral lateral margin almost straight until the half-length or even more; humeral deltopectoral crest markedly expanded distally; humerus, strong posterolateral bulge around the level of the most distal portion of the deltopectoral crest; astragalus with symmetrical development of the articular surfaces, with the lateral surface for articulation of fibula and calcaneum as developed as the surface for the tibia (*); ratio of mediolateral expansion of the tibia/tibial facet of the astragalus nearly 75% (*); astragalus with a hemispherical posterior bulge (*); astragalus without posterior fossa; mediolateral width of the astragalus four times its proximodistal height (*).
Holotype
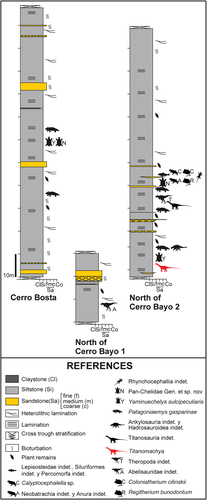
MPEF Pv 11547, a partial skeleton consisting of: MPEF Pv 11547/10, a posterior caudal vertebra (incomplete); MPEF Pv 11547/14/1–6 incomplete ribs; MPEF Pv 11547/11/12, haemal arches; 11547/7, left humerus; MPEF Pv 11547/15 left iliac fragment; MPEF Pv 11547/13, right pubic fragment; MPEF Pv 11547/1, right femur; MPEF Pv 11547/2, left femur (distal); MPEF Pv 11547/4, right tibia; MPEF Pv 11547/6, left tibia; MPEF Pv 11547/3, right fibula; MPEF Pv 11547/5, left fibula; MPEF Pv 11547/8, complete right astragalus; MPEF Pv 11547/9, incomplete left astragalus ().
Figure 4. Spatial distribution of Titanomachya gimenezi skeletal elements.
Locality and horizon
The specimen here described come from the north of Cerro Bayo area (see Gasparini et al. Citation2015; Sterli et al. Citation2022; ). Precise GPS data has been deposited at the MPEF collection. This area is located approximately 10 km south-west of Bajada del Diablo, Chubut Province, Patagonia, Argentina. This area exposes outcrops of the La Colonia Formation (Maastrichtian – Danian; Clyde et al. Citation2021). This stratigraphic unit is composed mostly of laminated mudstones, pelitic fine sandstones, and banded siltstones and claystones deposited in an estuary, tidal flat or coastal plain environment, influenced by occasional high freshwater stream flow from the continent and tidal currents from the sea (Pascual et al. Citation2000; Gasparini et al. Citation2015; Sterli et al. Citation2022). This facies association has been deposited under a seasonal climate with alternating periods of humidity and aridity (Ardolino and Delpino Citation1987; Ardolino et al. Citation1995; Gasparini et al. Citation2015). The holotype of Titanomachya gimenezi comes from the middle section of the La Colonia Formation (see Gasparini et al. Citation2015; ), in levels like those of other dinosaurs (abelisaurids, hadrosaurs, ankylosaurs), turtles, and mammals reported from this formation.
Description
Axial skeleton
Caudal vertebrae
MPEF 11,547/10 is a posterior caudal vertebra (). It preserves the complete centrum and the base of the neural arch from the right side with the right prezygapophysis. The neural spine is not preserved.
Figure 5. Titanomachya gimenezi, holotype. MPEF Pv 11547/10. Posterior caudal vertebra in A, posterior; B, right lateral; C, anterior; D, dorsal and E, ventral views. Abbreviations: lr, longitudinal ridge. Scale bar = 5 cm.
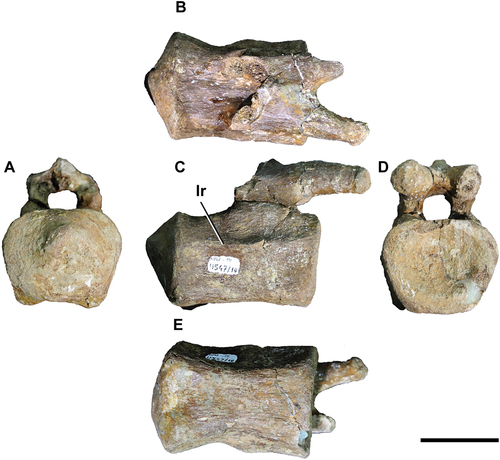
In lateral view (), the centrum is 50% longer than its height (measured anteriorly; ) and has straight anterior and posterior margins (disregarding the posterior articulation of the centrum). Likewise, both margins are inclined anteriorly 5° with respect to the ventral surface of the centrum. At mid-height of the lateral face of the centrum, a anteroposteriorly orientated subtle ridge is observed. The edge of the lateral ridge on the left side is more pronounced on the anterior part, just below the neural arch. In turn, that of the right side is equally pronounced throughout the centrum length. Dorsal and ventral to this ridge, the lateral face of the vertebral centrum is dorsoventrally concave. This is more apparent on the right lateral face. The lower margin of the centrum is slightly concave. The posterior articulation has its apex displaced somewhat dorsally. The dorsal and ventral margins of the posterior articulation are straight, being the ventral margin the longer one.
Table 1. Measurements of the elements of the axial skeleton of Titanomachya gimenezi.
The vertebral centrum is wider than it is high (). In fact, the maximum width of the centrum equals the height taken from the ventral margin of the centrum to the dorsal margin of the preserved base of the neural arch. In posterior view (), the vertebral centrum is hexagon shaped. Viewed from the posterior end, the ventral margin of the centrum is slightly concave, whereas the dorsal margin is approximately straight. In dorsal view (), the lateral margins of the vertebral centrum are nearly straight and subparallel. The posterior articulation is pointed, and the lateral margins of the posterior articulation are straight to slightly concave. The neural canal is wider than it is tall.
The ventral face of the centrum is slightly concave transversely and anteroposteriorly, but it does not form a distinct groove (). This concavity becomes slightly deeper towards the posterior part of the centrum. The ventral face becomes narrower at the mid-length of the centrum. The longitudinal borders of the ventral face are little pronounced, therefore there are no marked ventrolateral ridges. The centra of the vertebra is broken in such way that the internal tissues can be seen. Based on the damaged area, the internal structure of the vertebra appears to be solid, not camellate.
The little that has been preserved of the neural arch of the vertebra is in the anterior half of the centrum (). Anteriorly and posteriorly to the neural arch, the dorsal surface of the vertebral centrum is almost flat, slightly concave transversely.
Compared with the posterior caudal of other lithostrotians of the Campanian-Maastrichtian interval, Titanomachya gimenezi (MPEF 11547/10) exhibits notable differences. The vertebral centrum of the sauropod from the La Colonia Formation is not dorsoventrally compressed as in the species Rocasaurus and Pellegrinisaurus powelli (Salgado Citation1996); its lateral ventral margin is much less concave, the posterior articulation notably more pointed, and it has a slight longitudinal ridge at the mid height of the lateral face of the vertebral centrum that is not observed in the species from the Allen Formation (e.g. Bonatitan, Rocasaurus, and Pellegrinisaurus).
In addition, Titanomachya gimenezi (MPEF 11547/10) does not have the ventral depression present in Rocasaurus, although this absence could be due to its position in the sequence. In Titanomachya gimenezi, the base of the neural arch is shorter anteroposteriorly relative to the length of the vertebral centrum than in Rocasaurus, and the prezygapophyses are relatively longer.
In lateral view, the caudal centrum of Titanomachya is very similar to that of Rapetosaurus krausei (Curry Rogers and Forster Citation2001), although the posterior articulation is more pointed and somewhat dorsally located in the Patagonian sauropod. On the other hand, the longitudinal ridge on the lateral face is not present in the sauropod from Madagascar, whose posterior articulation is, in posterior view, more subcircular than in the Patagonian sauropod. It is also very similar to that of Isisaurus colberti (Jain and Bandyopadhyay Citation1997), although in the species from India the neural arch is proportionally longer and the ventral margin of the centrum more concave in lateral view (Jain and Bandyopadhyay Citation1997, figs. 13, 14).
In the posterior caudal vertebra of Titanomachya gimenezi the base of the neural arch is less anteriorly located than in the tenth? caudal vertebra of Aeolosaurus sp (MPCA−Pv 27174; García and Salgado Citation2013, fig. 2D).
Proportionally, the vertebral centrum of Titanomachya gimenezi is similar to that of the distal mid-caudal vertebrae of Menucocelsior, although in the latter, the vertebral centra are, in antero/posterior views, more subcircular than in Titanomachya gimenezi, where the centrum is rather square in these views. Likewise, both resemble each other in the position of the base of the neural arch (Aranciaga Rolando et al. Citation2022, fig. 4R—S) and in the lack of a ventral groove.
Proportionally, the only preserved caudal vertebra of Titanomachya gimenezi (MPEF 11547/10) is very similar to the caudal vertebra N° 23 of Dreadnoughtus (Lacovara et al. Citation2014, fig. 1M). In Dreadnoughtus, the ratio length (counting the condyle)/height of the vertebral centrum is 1.88 (Lacovara et al. Citation2014, Supplementary Information), whereas in Titanomachya (MPEF 11547/10) it is ~1.81. Similarly, Dreadnoughtus caudal vertebra N° 23 appears to have ventral margins as straight as in Titanomachya (MPEF 11547/10). In any case, Dreadnoughtus does not present the lateral longitudinal ridge observed in Titanomachya (MPEF 11547/10).
Titanomachya is similar to the specimen MPCA−Pv 862 from Salitral Moreno (Río Negro Province) (García and Salgado Citation2013, fig. 12), although in this last specimen the caudal centrum is slightly taller proportionally (centrum length/height = 1.66). However, beyond their matching proportions, there are important morphological differences. In MPCA−Pv 862, the ventral and dorsal margins of the vertebral centrum are very concave, whereas in Titanomachya (MPEF 11547/10) they are only slightly concave.
The posterior margin of the centrum of MPCA−Pv 862 (not counting the posterior articulation) is convex in lateral view (García and Salgado Citation2013, fig. 12A1), whereas in Titanomachya (MPEF 11547/10) it is straight. In lateral view, the anterior margin of the centrum of MPCA−Pv 862 is convex, and that of Titanomachya (MPEF 11547/10) is straight. In addition, the lateral ridge (present on both caudal vertebrae) is much more pronounced in Titanomachya than in MPCA-Pv 862.The posterior articulation is pointed in both specimens, but it is more pronounced in MPCA−Pv 862. In posterior view, the morphology of both is very similar, but in MPCA−Pv 862 the ventral face is slightly more excavated (García and Salgado Citation2013, fig. 12 A3), which could be indicating a difference in position in the sequence.
In MPCA−Pv 862, the posterior articular condyle is more pronounced than in Titanomachya gimenezi (MPEF 11547/10) (García and Salgado Citation2013, fig. 12 A2). The neural arch is similar, but in MPCA−Pv 862 the neural canal appears to be taller than it is wide, while in Titanomachya gimenezi (MPEF 11547/10) it appears to be wider than it is tall. As can be seen in ventral view, the ventral face of MPCA−Pv 862 is broader, flatter, and their lateral margins more concave.
The only preserved caudal vertebra of Titanomachya gimenezi (MPEF 11547/10) resembles the most posterior caudal of Nullotitan (holotype, MPM 21542) (Novas et al. Citation2019, fig. 17 R and S), which, according to Novas et al. (op. cit.), corresponds to the caudal N° 21 (here, the length/height ratio of the vertebral centrum is approximately 1.45, which means that the caudal vertebra from Santa Cruz is proportionally a little taller and shorter). In MPM 21542, the longitudinal rim that runs along the lateral face of the caudal vertebra of Titanomachya gimenezi (MPEF 11547/10) is not observed, and the ventral edge of the latter, is considerably less concave in lateral view than in MPM 21542.
Haemal arches
Two complete chevrons (MPEF Pv 11547/11 and 12) are preserved (). As in other sauropods, such as Shunosaurus lii (Dong et al. Citation1983), basally branching members of Diplodocoidea, most macronarians and several titanosaurs (e.g. Isisaurus, Saltasaurus, Andesaurus delgadoi, Malawisaurus dixeyi, Epachthosaurus sciuttoi, Bonitasaura salgadoi; Haughton Citation1928; Powell Citation1990; Calvo and Bonaparte Citation1991; Apesteguía Citation2004), all Titanomachya gimenezi chevrons are type OY (open ‘Y’-shaped; Otero et al. Citation2012), being of the subtype SOY (straight and open Y-shaped). Based on the morphology of these subtypes these elements would correspond to the most anterior region of the tail.
Figure 6. Titanomachya gimenezi, holotype. MPEF pv 11547/11 and 12. Middle haemal arch A, right lateral; B, anterior views. Anterior haemal arch C, right lateral; D, anterior views. Scale bar = 10 cm.

The haemal canals constitutes approximately 25% of the total chevron length in MPEF-Pv -11547/11, whereas it comprises approximately 30% in MPEF-Pv -11547/12. (). This condition is not common among titanosaurs, which have a haemal canal representing 50% of the total length of the chevrons. The condition seen in Titanomachya is traceable in Saltasaurus, diplodocoids and in non-titanosaurian somphospondyians (e.g. Diplodocus carnegii, Giraffatitan brancai; Hatcher Citation1901; Janensch Citation1914). The proximal rami are strongly compressed anteroposteriorly with this compression becoming more pronounced distally as in Andesaurus. Moreover, the proximal rami are not a different structure from the blade but run smoothly to the distal end. The articular heads are transversely thickened, and have a simple, flat articular surface, as in Malawisaurus, Saltasaurus and Bonitasaura. The articular surfaces are ventromedially inclined at an angle of approximately 45°.
Dorsal ribs
Numerous pieces of ribs have been recovered from the Titanomachya quarry, but few provide important information on the costal grill of this taxon. Elements MPEF Pv 11547/14/from 1 to 6 represent six fragmented right thoracic vertebrae ().
Figure 7. Titanomachya gimenezi, holotype. MPEF Pv 11547/14/1–6. Thoracic ribs fragments. MPEF Pv 11547/1 in A, external and B, internal views. MPEF Pv 11547/2 in C, external view. MPEF Pv 11547/3 in D, external and E, internal views. MPEF Pv 11547/4 in F, external and G, internal views, MPEF Pv 11547/5 in H, external and I, internal views, MPEF Pv 11547/6 in J, internal; K, proximal; L, anterior; M, external and N, posterior views. Scale bar = 30 cm.

Both elements MPEF Pv 11547/14/1 () and MPEF Pv 11547/14/2 () represent the most distal portions of anterior to mid ribs (probably 3rd to 5th). These are decidedly flattened bilaterally, and their distal ends are expanded antero-posteriorly. A semicircular process resembling an uncinate process can be seen on the anterior border of MPEF Pv 11547/14/1 ().
The elemment MPEF Pv 11547/14/5 () represents a fragment of the middle third of a probable 5th to 7th dorsal rib. In lateral view it is strongly convex both anteroposterior as proximodistally. In cross-section the rib is subcylindrical and it is slightly broader anteroposteriorly than lateromedially.
Elements MPEF Pv 11547/14/3 () and MPEF Pv 11547/14/4 () lack proximal (capitulum and tuberculum) and distal ends, however, their morphology allows them to be interpreted as possible middle to posterior ribs (probably 7th to 8th). As in the mid-posterior ribs of Opisthocoelicaudia skarzynskii (Borsuk-Bialynicka Citation1977) and Overosaurus paradasorum (Coria et al. Citation2013), these ribs are thin and strongly curved proximodistally. Despite their marked compactness, in cross-section these ribs are suboval or even rounded in the transverse section.
Dorsal rib MPEF Pv 11547/14/6 differs from the other ribs by being anteroposteriorly wider and more compressed, with the proximal and distal cross-sections being triangular (). In lateral view, this rib has sigmoidal outlines with distal ends curved backwards (). In medial (internal) view, the rib has a proximodistal short and robust keel which is in connection with the capitulum as seen in the dorsal ribs of Overosaurus (). The capitulum projects medially perpendicular to both the tuberculum and shaft. The tuberculum projects more dorsally than the capitulum. In both anterior and posterior sides, a closed depression is present between capitulum and tuberculum. Therefore, we think this rib probably be the 9th or 10th dorsal rib of Titanomachya.
Appendicular skeleton
Humerus
MPEF 11547/7 is a left humerus (, ). It is a robust bone, with a robustness index (RI) of 0.33 being therefore less robust than that of Opisthocoelicaudia (RI = 0.37), but it is within the range recorded for Neuquensaurus australis (Lydekker Citation1893; RI = 0.30 − 0.33) (Otero Citation2010). It is morphologically similar and with a similar RI to that of Dreadnoughtus (Lacovara et al. Citation2014, fig. 2D), somewhat more robust than that of Rapetosaurus (RI = 0.29, Otero et al. Citation2020), Nullotitan and Patagotitan mayorum (both with a RI = 0.28; Carballido et al. Citation2017), Bonatitan (both with a RI = 0.27, Curry Rogers Citation2009, ; Salgado et al. Citation2014), Aeolosaurus sp. (RI = 0.26, APM personal observation), Isisaurus colberti (RI = 0.25, Jain and Bandyopadhyay Citation1997, ), and much more than in Narambuenatitan palomoi (Filippi et al. Citation2011; RI = 0,20, APM, personal observation).
Figure 8. Titanomachya gimenezi, holotype. MPEF Pv 11547/7, left humerus in A, posterior; B, proximal; C,distal; D, medial and E, anterior views. Abbreviations: cb, coracobrachialis origin scar; dc, deltopectoral crest; hh, humeral head; lc, lateral condyle; ld, latissimus dorsi insertion scar; mc, medial condyle; scf, supracondylar fossa; suc, supracoracoid insertion scar. Scale bar = 10 cm.
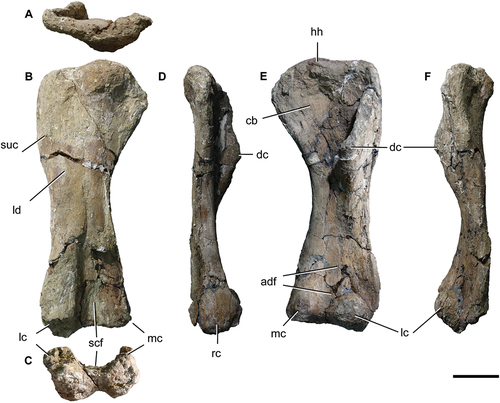
Table 2. Measurements of the forelimb of Titanomachya gimenezi.
Table 3. Measurements of femora and tibiae of Titanomachya gimenezi.
In general, the humerus of Titanomachya is similar to that of Dreadnoughtus. The humeral head is quite straight, not very sigmoid. The lateroproximal corner of the humerus forms a right angle (). The deltopectoral crest, site of insertion of the supracoracoideus and deltoideus clavicularis muscles (Otero Citation2018), widens distally. Around the level of the distal portion of the deltopectoral crest there is a strong posterolateral bulge: the supracoracoideus insertion scar (Otero Citation2018; Otero et al. Citation2020, fig. 3G, H). This feature described here as representing supracoracoideus is present in almost all titanosaurs (e.g. see C280 in Mannion et al. Citation2019). A similar structure was also recorded in Abditosaurus kuehnei (Vila et al. Citation2022). Ventromedial to this, there is the latissimus dorsi insertion scar (), as in Patagotitan (Otero et al. Citation2020, fig. 3G, H).
On the medial part of the proximal anterior fossa of the humerus, there are small parallel ridges that, following to Otero (Citation2018), are interpreted as the coracobrachialis origin scar (). In this sense, Titanomachya differs from Patagotitan where there are two scars in the same position (an autapomorphic character of that giant sauropod, Otero et al. Citation2020, fig. 3).
The humeral distal articular surfaces are anteriorly exposed (), as is characteristic of saltasaurines and some non-South American lithostrotians (González Riga et al. Citation2019). The lateral or radial condyle is undivided, as in most titanosaurs (e.g. Neuquensaurus, Nullotitan, Aeolosaurus rionegrinus, Isisaurus, Patagotitan). The lateral and medial condyles are about at the same height, being the lateral one slightly lower (). The anterodistal fossa (Otero Citation2018, fig. 12) is shallow (). On the posterior face of the distal end of the humerus, there is the supracondylar fossa (). Although this area is not well preserved, this fossa occupies approximately a quarter of the total length of the humerus.
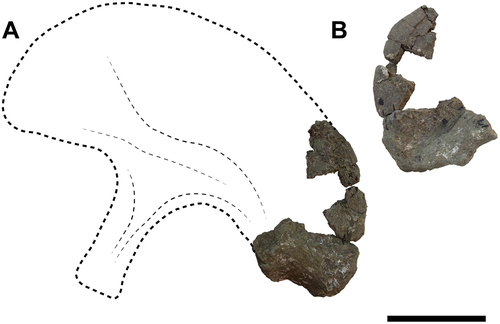
Ilium
A fragment of a left ilium has been preserved (), in which a very small part of the acetabulum, the ischial peduncle and the postacetabular process can be observed. Dorsally, the markedly curved posterodorsal outline is preserved as in most sauropods. There is no evidence of pneumaticity in either the ilium fragments
Figure 9. Titanomachya gimenezi, holotype. MPEF Pv 11547/15, left ilia fragment in A, external and B, internal views. Abbreviations: isped, ischial peduncle; poap, postacetabular process. Scale bar = 15 cm.
Pubis
A proximal fragment of a right pubis is preserved (). On its proximal part, the ventral margin of the pubic foramen is observed, which would have been anteroposteriorly elongated. The entire distal process and part of the proximal sector of the pubis have not been preserved. The laterodorsal margin of the pubis is curved. The lateroventral surface of the pubis (external) is smoothly convex. There is no evidence of pneumaticity in the pubis fragment.
Figure 10. Titanomachya gimenezi, holotype. MPEF Pv 11547/13, indeterminated pube fragment in external view. Scale bar = 20 cm.

Femur
A complete right femur has been preserved, MPEF 11547/1 (RI = 0.24), and one distal end of a left femur, MPEF 11547/2 (; ). Although the femur of Titanomachya (MPEF 11547/1) is somewhat diagenetically compressed, it is possible to see that it is straighter than in Dreadnoughtus, where it is more sigmoid.
Figure 11. Titanomachya gimenezi, holotype. MPEF Pv 11547/1, right femur in A, anterior; B, proximal; C, medial; D, posterior and E, distal views. Abbreviations: fh, femoral head; gt, greater trochanter; lb, lateral bulge; lec, lateral epicondyle; lc, lateral condyle; mc, medial condyle; pr, posterior ridge; 4t, fourth trochanter. Scale bar = 20 cm.
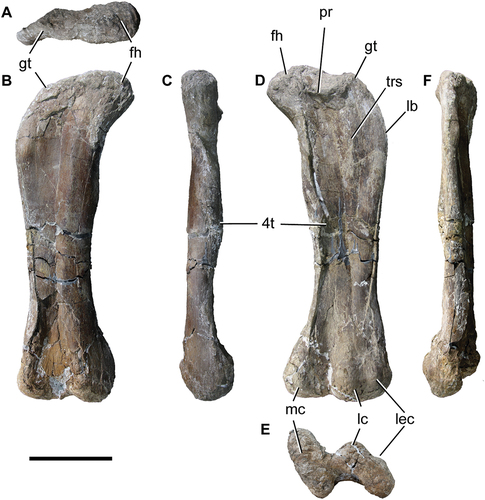
As with the humerus, the femur of Titanomachya is robust (RI = 0.24); a bit more robust than those of Rocasaurus (MPCA-Pv 46/6, RI = 0.23) and Titanosauria gen. et sp. indet. (MPCA-Pv 33/2, RI = 0.23), but less than the other femur recorded in Salitral Moreno: Aeolosaurus sp (MPCA-PV 27174, RI = 0.26; García and Salgado Citation2013, figs. 5). In this respect, it occupies an intermediate position between the saltasaurine Neuquensaurus (RI = 0.249 − 0.28, Otero Citation2010) and Dreadnoughtus (RI = 0.27), and the more gracile Bonatitan, Patagotitan (both with RI = 0.23), Mendozasaurus neguyelap (González Riga Citation2003) and Bonitasaura (RI = 0.2 and 0.22 respectively; Salgado et al. Citation2014; Gallina and Apesteguía Citation2015; Otero et al. Citation2020).
In Titanomachya, the femoral head seems to be less developed than in Dreadnoughtus, where it is not only more developed, but also more dorsally projected. In the sauropod from the La Colonia Formation, the dorsal margin of the proximal end of the femur is more horizontal and concave than in Dreadnoughtus. In the latter, this margin is dorsomedially more inclined than in the sauropod from the La Colonia Formation. In Titanomachya gimenezi, there is a pronounced ridge on the posterior surface between greater trochanter and head (, pr). Also, distal to the greater trochanter, there is a ridge that is here interpreted as the remainder trochanteric shelf.
The transverse width of the femur gradually increases from the ventral constriction towards the lateral bulge. This expansion is not so noticeable in Dreadnoughtus, precisely because its femur is sigmoid. The outermost part of the lateral bulge is located more proximally in Dreadnoughtus (Lacovara et al. Citation2014, fig. 2F) and in the three femora from Salitral Moreno (García and Salgado Citation2013, fig. 5) than in Titanomachya gimenezi ().
Titanomachya lacks the constriction of the femoral shaft at the distal end of the lateromedial expansion that is present in Dreadnoughtus (Lacovara et al. Citation2014, fig. 2F), although this could be due to the crushing suffered by the material. In the sauropod from Chubut, the tibial condyle is more developed than the fibular condyle, and the lateral epicondyle does not have a very important development and is not as lateralised as in Bonatitan (Salgado et al. Citation2014, ) and Bonitasaura (Gallina and Apesteguía Citation2015, ).
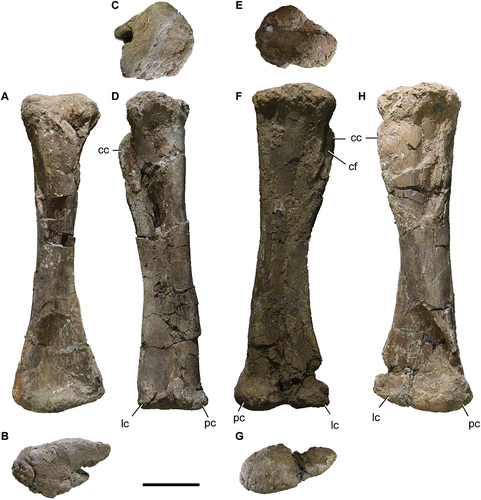
Tibia
The two tibiae are preserved (). These are somewhat crushed, as are the other appendicular bones. Although both bones have roughly the same degree of preservation, the cnemial crest area of the left tibia (MPEF 11547/6) is somewhat better preserved than the right one (MPEF 11547/4; ).
Figure 12. Titanomachya gimenezi, holotype. Tibiae: (a-d) left tibia MPEF 11547/6 in A, anterior; B, distal; C, lateral and D, proximal views. Abbreviations: cc, cnemial crest; cf, cnemial fossa; lc, lateral condyle; pc, posterior condyle. Scale bar = 10 cm.
The tibia of Titanomachya is gracile and slightly sigmoid (). Both tibiae (MPEF 11547/4 [RI = 0.245] and MPEF 11547/6 [RI = 0.225]) are more graceful than that of Neuquensaurus (RI = 0.30 − 0.36, Otero Citation2010), Bonatitan (RI = 0.266 and 0.257, Salgado et al. Citation2014) and Laplatasaurus araukanicus (Huene Citation1929; RI = 0.22, Gallina and Otero Citation2015). Like Epachthosaurus it has a tibia/femur ratio greater than 0.6 (0.69). This ratio is also traceable to diplodocid and titanosauriformes (Mannion et al. Citation2019).
The proximal articulation of the left tibia of Titanomachya gimenezi (MPEF 11547/6) has a subcircular contour in proximal view (), as is characteristic in titanosaurs, while the distal end is expanded lateromedially (). The distal expansion is slightly greater than the anteroposterior expansion of the proximal end (). In this sense, it is very different from the Nullotitan tibia where both ends are strongly expanded, especially the proximal one (Novas et al. Citation2019, fig. 22).
The cnemial crest of Titanomachya gimenezi is anterolaterally directed. It does not seem to be as expanded as in other titanosaurs, like Bonatitan, among others. The cnemial fossa between the cnemial crest and the tibial head is deep (), unlike Nullotitan (Novas et al. Citation2019, fig. 22).
The medial surface of the tibial diaphysis is flat in its middle part. The anterior surface of the diaphysis in its middle part is slightly convex. In both tibiae, the anterior surface of the distal end of the shaft is similarly fractured and depressed. The posterior condyle is more developed than the lateral condyle (), proportionally more developed than in individual B of Bonatitan (MACN-PV RN 821, Salgado et al. Citation2014, fig. 11b). The distal surface of the posterior condyle is rough. The posterior condyle is slightly below the level of the lateral condyle, as in other titanosaurs. The groove between the lateral and posterior condyles is not well preserved, but it seems not to have been much closed (), as observed in other titanosaurs, such as Mendozasaurus (Gonzalez Riga et al. Citation2018, fig. 20L). The proximal process of the astragalus was inserted into this groove (Otero Citation2010, ).
Fibula
Both fibulae have been recovered, being the right fibula (MPEF 11547/3) better preserved than the left one (MPEF 11547/5; ). It is a straight element, straighter than in Nullotitan, where the fibula is more sigmoid.
Figure 13. Titanomachya gimenezi, holotype. Fibulae. MPEF 11547/5 left fibula in A, lateral; B, distal; C, proximal, and D, posterior views. MPEF 11547/3 right fibula in E, medial; F, proximal and G, lateral views. Abbreviations: lt, lateral tuberosity. Scale bar = 10 cm.
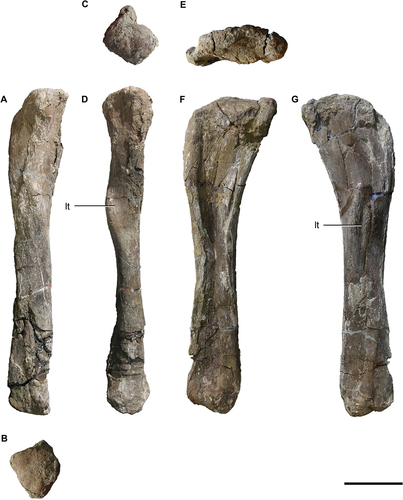
Both bones have different degrees of robustness (), presumably due to preservational deformation, which makes it difficult to establish comparisons (left fibula RI = 0.125; right fibula RI = 0.18). Even so, there can be seen that the fibula of Titanomachya gimenezi is more gracile than that of Uberabatitan (RI = 0.6, Novas et al. Citation2019), Nullotitan (RI = 0.4, Novas et al. Citation2019), and Neuquensaurus (RI = 0.7 − 0.23). In Laplatasaurus (RI = 0.16, Gallina and Otero Citation2015) and Bonatitan (RI = 0.149, Salgado et al. Citation2014) the robustness indexes are within the range comprised by both fibulae of Titanomachya gimenezi.
Table 4. Measurements of fibulae and astragalus of Titanomachya gimenezi.
The proximal end of the left fibula of Titanomachya appears subquadrangular in proximal view (), although this is due to deformation, whereas the distal end is subtriangular in distal view (). The proximal end of the right fibula is lateromedially compressed (). This is the less deformed of both fibulae.
The anteroproximal fossa is present but it is poorly developed; it is not as wide as in Laplatasaurus (Gallina and Otero Citation2015, fig. 3.1, 8) or Uberabatitan (Salgado and Carvalho Citation2008, fig. 19).
The medial face of the right fibula of Titanomachya gimenezi is collapsed, but the lateral face is well preserved. On this last face, the lateral protuberance or iliofibularis tubercle is observed (). The posterior rim of this protuberance is better developed. This rim forms an angle with the long axis of the bone of approximately 20°, which is lower than that observed in Uberabatitan (Salgado and Carvalho Citation2008, fig. 19G). In Nullotitan, the anterior rim of the protuberance is the one that seems to be more developed (Novas et al. Citation2019, fig. 23B). In this last sauropod, the anterior rim is subparallel to the long axis of the bone, as in Titanomachya.
In anteroposterior view, the protuberance or lateral tubercle is slightly asymmetrical in Titanomachya gimenezi with the most laterally projected point of the tuberosity being located on the proximal part of it (). In contrast, in Menucocelsior (Aranciaga Rolando et al. Citation2022, fig. 5F, G) and other sauropods, such as Laplatasaurus (Gallina and Otero Citation2015) and Uberabatitan (Salgado and Carvalho Citation2008, fig. 19H), the tuberosity is rather symmetrical, and its most laterally projected point placed at the middle of the tuberosity.
In Titanomachya, the posterior surface to the posterior rim of the protuberance is slightly concave. In this area, there is not a tuberosity fossa like the one that exists in Laplatasaurus (Gallina and Otero Citation2015, fig. 3.4, 11). In the middle of the diaphysis, below the protuberance, the lateral face of the diaphysis is flat and oriented somewhat anteriorly.
Astragalus
Both astragali are preserved: MPEF 11547/9 (left, incomplete, lacking part of the medial surface to articulate with the tibia) (), and MPEF 11547/8 (right, complete) (). The astragalus is wedge-shaped, transverselly wider than anteroposteriorly long and proportionally low, being three times wider than high (). It lacks foramina at the base of the ascending process. The astragalus of Titanomachya gimenezi is unusual in that its lateral surface for articulation of fibula and calcaneum (fibular surface, fs in ) is as developed as the surface for the tibia (tibial surface, ts in ). The fibular surface is wide, gently concave, a little more than the tibial surface. There is a dorsal process on both astragalus (clearly observed in anterior view), oriented slightly laterally. In the right one, it is better preserved (). We interpret this structure as the ascending process observed in other dinosaurs.
Figure 14. Titanomachya gimenezi, holotype. MPEF 11547/9. Left astragalus in A, anterior; B, dorsal; C, posterior; D, ventral; E, fibular and F, medial views. asp, ascending process. Scale bar = 5 cm.

Figure 15. Titanomachya gimenezi, holotype. MPEF 11547/8. Right astragalus in A, anterior; B, dorsal; C, posterior; D, ventral; E, fibular and F, tibial views. Abbreviations: asp, ascending process; fs, fibular surface; hb, hemispherical bulge; ts, tibial surface. Scale bar = 5 cm.

In describing the astragalus of Neuquensaurus, Salgado et al. (Citation2005, p. 632) wrote: ‘The fact that the titanosaur astragalus is transversely reduced means that the transverse expansion of the distal end of the titanosaur tibia was not accompanied’. In fact, in Neuquensaurus (Salgado et al. Citation2005, Table I), the width of the astragalus MCS-5/29 that was found articulated to the tibia MCS-5/25 is 56% of the width of the distal end of the tibia. In turn, in Opisthocoelicaudia, the same relationship is about 54%, and in Uberabatitan 69% (Salgado and Carvalho Citation2008). In the right astragalus of Titanomachya, the tibial surface of the astragalus is 16 cm, and the lateromedial expansion of the tibia is 22 cm, which gives a ratio of 76%.
A mediolaterally short astragalus was considered by D’emic (Citation2012) to be a synapomorphy of Saltasauridae; in Titanomachya gimenezi, the fact that it is so mediolaterally extensive may be considered an autapomorphy (ch. 378/2).
In the ventral half of the posterior face of the astragalus there is an ovoid, hemispherical protuberance (whose longer axis is laterodorsally/centrally oriented), which is not observed in the astragalus of other titanosaurs. In this regard, this feature is considered an autapomorphy of Titanomachya.
In dorsal view, the astragalus of Titanomachya has a relatively flat surface, which would be oriented slightly medially, separating the slightly concave, posteromedially oriented tibial articulation from the more concave, posterolaterally oriented, fibular/calcaneal articulation. The ventral surface of the astragalus is rugose, and the anterior surface is smooth.
Discussion
Phylogenetic analysis
Phylogenetic analysis resulted in 237 MPTs of 1608 steps that resulted in a partially resolved Strict Consensus Tree (RI = 0.71; CI = 0.35) (). Strict consensus tree exhibits several polytomies in different clades (). To evaluate the causes of this low resolution in those points of the phylogeny and detect if it is due to a combination of taxa and characters, or if, on the contrary, it is due to the presence of one or more unstable taxa (‘wildcard’ sensu Nixon and Wheeler Citation1992), the iterative PCR method was used (‘Positional Congruent Reduced’; iterPCR [Pol and Escapa Citation2009]). With this method it was possible to detect different alternative positions for the taxa Isanosaurus attavipachi, Rayososaurus agrioensis, Lusotitan atalaiensis, Malarguesaurus florenciae, Nemegtosaurus mongoliensis, Andesaurus, Isisaurus, Baurutitan, Epachthosaurus, Puertasaurus and Notocolossus gonzalezparejasi (Lapparent and Zbyszewski Citation1957; Nowinski Citation1971; Bonaparte Citation1996; Buffetaut et al. Citation2000; González Riga et al. Citation2009; González Riga et al. Citation2016). Most of the polytomies are resolved by pruning these taxa from the MPTs (). The result of our phylogenetic analysis recovers Titanomachya as a taxon deeply nested within Saltasauroidea. The strict consensus tree topology obtained is, in general terms, similar to that obtained in previous iterations of this dataset.
Figure 16. Strict consensus of the phylogenetic analysis. Matrix based on that of Pérez Moreno et al. (Citation2023).
As in previous iterations of this dataset, Lithostrotia is recovered here as a clade reduced in taxonomic content with respect to than in its original formulation definition and nested within Saltasauroidea. Lithostrotians here supported by eight synapomorphies: 1) presence of paroccipital process with ventral non-articular process (cha. 69/1); 2) presence of axis with centrum twice as long as high (cha. 121/1); 3) presence of cervical vertebrae with height/width ratio less than 0.7 (cha. 132/3); 4) presence of anterior cervical vertebrae with prespinal lamina (cha. 137/1); 5) presence of anterior caudal neural arches with well-developed SPRL lamina extending to the anterior or anterolateral borders of the neural spine (cha. 243/2); 6) presence of mid-caudal vertebrae with vertical neural spines (cha. 257/1); 7) presence of posterior caudal vertebrae with vertical neural spine (cha. 260/0) and 8) presence of mid-caudal vertebrae whose prezygapophyses length with respect to the length of the centrum vary between 40% and 50% (cha. 423/1). Titanomachya is recovered within Lithostrotia and shares three derived characters with saltasaurids: caudal bone texture solid (221/1), humerus with a strong posterolateral bulge on around level of the deltopectoral crest (298/1) and humeral deltopectoral crest markedly expanded distally (302/1). In turn, the analysis reveals three characters that are autapomorphies for the new species within the context of Lithostrotia: chevron haemal canal short, approximately 25% chevron length (270/0), humeral deltopectoral attachment prominent (301/0) and fibular facet of the astragalus faces posterolaterally, revealing the anterior margin in posterior view (374/1).
Jackknife and Bootstrap values are below 50% in most of the internal nodes of Macronaria. This low support for macronarians was already mentioned by Carballido and Sander (Citation2014) and, more recently, by Carballido et al. (Citation2020). Support values for Saltasauroidea and other titanosaur clades are generally low (see Supplementary Information), but this is at least partially caused by the presence of several incompletely known taxa that decrease nodal support measures because they can be easily placed in different nodes. For instance, Bremer support values are minimal and jackknife frequency values are below 50% for almost all nodes within Titanosauria. The pcrjak script identifies 20 titanosaurian taxa that decrease jackknife values (see Supplementary Information). Ignoring the alternative positions of these taxa in the jackknife replicates results in the identification of more than 15 titanosaurian nodes that have moderately high support (between 54% and 95%). Titanomachya is rather incompletely known and therefore it is highlighted as one of the titanosaurian taxa that decrease nodal support and forcing Titanomachya in different positions within Saltasauroidea leads to only moderately suboptimal topologies. A more basal position within Saltasauroidea (as the most basal branch of this clade or as the sister group of Lithostrotia) implies three extra steps and a more derived position (as the sister group of Neuquensaurus+Saltasaurus) implies two extra steps. In sum, currently available information helps placing Titanomachya in a phylogenetic context although more material is needed to place this taxon with high levels of certainty Saltasauroidea.
Titanosaurian biotas composition of the Campanian–Maastrichtian of Patagonia
The composition of sauropod assemblages recorded in the Campanian – Maastrichtian of South America varies latitudinally, and several regions have been noted to contain distinct assemblages (Santucci and Filippi Citation2022). This includes the Brazilian assemblages (Marilia, Serra da Galga, and Adamantina formations), northern Patagonian (Allen, Anacleto, Los Alamitos and Angostura Colorada formations), central Patagonia (Lago Colhué Huapi and La Colonia formations) and southern Patagonian (Chorrillo, Dorotea, and Cerro Fortaleza formations).
The sauropod assemblages during the latest Cretaceous period in northern Patagonia are predominantly characterised by the prevalence of aeolosaurines and saltasaurines. Within the Allen Formation, a minimum of four distinct titanosaurs has been identified across various locations, primarily falling outside the clade Saltasauroidea. The saltasaurine taxon Rocasaurus was originally recognised at the Salitral Moreno locality (Salgado and Azpilicueta Citation2000), but a second specimen was reported from the Salitral Ojo de Agua (Aranciaga Rolando et al. Citation2022). Both sites have yielded additional titanosaur remains (García and Salgado Citation2013; Aranciaga Rolando et al. Citation2022), including the recently named Menucocelsior (a non-saltasaurine of uncertain affinities; Aranciaga Rolando et al. Citation2022). In proximity to the stone quarries of Lago Pellegrini, another saltasauroid, Pellegrinisaurus, has been identified and further west (close to the city of San Patricio del Chañar), the aeolosaurine Panamericansaurus schroederi (Porfiri and Calvo Citation2010) has been documented. Finally, at the Bajo de Santa Rosa, another non-saltasaurine taxon, Bonatitan has been identified.
Underlying the Allen Formation, the Anacleto Formation also presents a wealth of titanosaurs discovered across various sites. In this formation, the assemblage includes both aeolosaurines and saltasaurines (although the former group is less diverse) and representatives of the clade Colossosauria (e.g. Pitekunsaurus macayai; Filippi and Garrido Citation2008). Taxa such as Laplatasaurus and Neuquensaurus originate from the vicinity of Cinco Saltos, while a specimen of Antarctosaurus wichmannianus was unearthed 15 kilometres southwest of General Roca. From the outskirts of Rincon de los Sauces, taxa such as Pitekunsaurus and Narambuenatitan emerge, the latter considered a probable aeolosaurine (Pérez Moreno et al. Citation2023). Barrosasaurus casamiquelai (Salgado and Coria Citation2009) comes from Sierra Barrosa, northeast of Plaza Huincul.
Other units from northern Patagonia with more limited exposure, such as the Los Alamitos Formation and Angostura Colorada Formation have also yielded sauropod materials that include records of the genus Aeolosaurus (Powell Citation1987; Salgado and Coria Citation1993) and the specimen MACB-RN-147 (Powell Citation1987). In sum, the sauropod fauna in northern Patagonia during the late Campanian and Maastrichtian has long been recognised as predominantly composed by saltasaurines and aeolosaurines (Santucci and Filippi Citation2022), as well as other titanosaurians with uncertain affinities (Leanza et al. Citation2004; Salgado and Bonaparte Citation2007; García and Salgado Citation2013; Aranciaga Rolando et al. Citation2022). This assemblage was recognised as unique within South America and distinct from the faunas of southern Patagonia and those from Brazil. Within the same temporal context, Brazilian formations like Marilia, Serra da Galga, and Adamantina, which are considered either Campanian or Maastrichtian, appear to exclusively feature aeolosaurines. These taxa include Uberabatitan, Baurutitan, Caieiria allocaudata (Silva Junior et al. Citation2022), Adamantisaurus mezzalirai (Santucci and Bertini Citation2006), Gondwanatitan faustoi (Kellner and Azevedo Citation1999), and Arrudatitan maximus (Santucci and Arruda-Campos Citation2011).
The sauropod record in southern Patagonia has also noted to be distinct from the other regions, as argued by Novas et al. (Citation2019) and Aranciaga Rolando et al. (Citation2022). These authors indicated that the sauropod record of the Chorrillo Formation concurs with that of the Cerro Fortaleza Formation, in the existence of large body-sized titanosaurs showing affinities with Colossosauria (e.g. Puertasaurus) and more basal taxa (e.g. Dreadnoughtus). The recently described Nullotitan lacks characteristics of Aelosaurini and Saltasauridae, and may be related to Colossosauria (Novas et al. Citation2019). Reports on sauropods from the Dorotea Formation in southern Chile are scarce (Leppe et al. Citation2014) but seems to also include large-bodied titanosaurs, as in the Cerro Fortaleza and Chorrillo formations.
The sauropod fauna of the uppermost Cretaceous of Central Patagonia was until now little known, with the exception of the aeolosaurine Aeolosaurus colhuehuapensis (Casal et al. Citation2007), and the eutitanosaurs Argyrosaurus and Elaltitan from the Colhue Huapi Formation. Titanomachya gimenezi and the very fragmentary sauropod remains described by Gasparini et al. (Citation2015), are the sauropod records from the La Colonia Formation that contribute to the poorly sampled sauropod fauna from central Patagonia. During the latest Cretaceous of central Patagonia, there was a coexistence of small bodied saltasauroids (such as Titanomachya), aeolosaurines, and larger bodied titanosaurs of uncertain affinities. In this context, the sauropod record of central Patagonia approaches more closely the phylogenetic and ecological diversity of northern Patagonia than that of southern Patagonia. Further sampling is required, however, to test if the sauropod fauna from central Patagonia is sufficiently distinct from that of northern Patagonia.
According to current knowledge, the distribution of titanosaur faunas during the Campanian-Maastrichtian interval in Patagonia would have followed a north-south gradient. In the north, saltasauroids, aeolosaurines, and colossosaurians would have coexisted. Moving southward, aeolosaurines would have decreased until they were absent in the extreme south, where only saltasauroids and giant colossosaurians coexisted.
Body mass estimation
The known specimen of Titanomachya indicates a small adult size. We estimated body mass that ranges between 5.79 and 9.79 tons (considering the 25.4% of percent prediction error [PPE]: Campione Citation2017), with a mean body mass of 7.79 tons for this species. Estimates of < 10 tons are rare among titanosaurs. Within the Saltasauroidea there are a few examples. The lightest titanosaurs are Magyarosaurus dacus (Nopcsa Citation1915) and Lirainosaurus astibiae (Sanz et al. Citation1999) with a mass of less than a ton and two tons respectively. Among Saltasauridae, the heaviest is Ophistocoelicaudia with a mass of 25 tons, while within the Saltasaurinae the body mass varies between five and six tons.
Although the average body mass of Titanomachya is higher than that of Saltasaurus and Neuquensaurus, it does not reach the body mass of Opisthocoelicaudia. In this sense, it could be concluded that Titanomachya was a titanosaur with a similar body size as Saltasaurus and Neuquensaurus but heavier. The superior body mass compared to titanosaurs of similar size, such as Neuquensaurus and Saltasaurus, may stem from ontogenetic stage factors. However, supporting this hypothesis requires clarification of the specimen’s ontogenetic stage through histological studies. It is likely that this greater body mass influenced the peculiar anatomy of Titanomachya‘s astragalus to support such weight, but there is insufficient evidence to support this hypothesis.
Morphological diversity of the astragalus within Titanosauria
Within Titanosauria there are few taxa that have their astragalus preserved, however, there is a variety of forms within Colossosauria, different from that found in Saltasauroidea ().
Figure 17. Schematic drawings of the morphologies of astragalus within Titanosauria in all its views. A, diagram of the typical colossosaurianian astragalus and its articulation with tibia and fibula. B, diagram of the typical saltasaurioid astragalus and its articulation with tibia and fibula. C, diagram of the astragalus of Titanomachya and its articulation with the tibia and fibula.
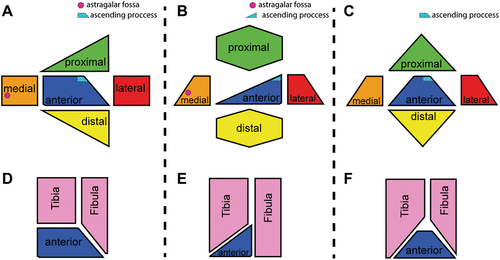
A typical ian astragalus can be recognised by the following morphology (): In anterior view the astragali are trapezoidal (not wedge-shaped), with the dorsal and lateral margin practically flat and the ventral margin curved. In lateral view, a subquadrangular perimeter is observed, with the dorsal margin flat and the rest slightly curved. On this face is developed the contact surface with the fibula, which can be flat or concave and covers almost the entire exposed area. The ascending process has a sharp edge directed upward. In posterior view the astragalus is tapering and presents an extremely reduced and somewhat rough surface. On the medial aspect there is an undivided fossa with a deep foramen. In medial view, the dorsal surface, flat, falls to end in the medial process, which is oriented obliquely. The anterior border is flat proximally and curves distally in a continuous line with the distal margin. The fossa and medial foramen, located in the posterior half of the astragalus, are well evident in this view. In proximal view the astragalus has a triangular outline, with its lateral and anterior sides forming an angle of nearly 90°. Although the proximal side is flat, there are two lines that separate it from the lateral and medial sides, which fall towards their respective sides. On the left side of the ascending process, part of the fossa and medial foramen can be observed. Finally, the distal view shows a rough and strongly convex surface contacting area for the proximal surfaces of, at least, the metatarsals II and IV. This type of morphology allows the articulation between the zeugopodium and the autopodium to have a preponderant contribution of the tibia and a minor contribution of the fibula.
On the other hand, a typical saltasauroid astragalus can be recognised by the following morphology (): in anterior view the astragali are scalene triangular (wedge-shaped), with the dorsal and lateral margins practically flat and the ventral margin curved. In lateral view a trapezoidal or wedge-shaped perimeter is observed, with the dorsal margin flat or slightly concave and the rest strongly curved. On this face is developed the contact surface with the fibula, which is usually concave and covers almost the entire exposed area. The ascending process is a robust structure. In posterior view the astragalus is tapering and presents an extremely reduced and somewhat rough surface. On the medial aspect there is an undivided fossa with a deep foramen. In medial view, the dorsal surface, concave, falls to end in the medial process, which is oriented obliquely. The fossa and medial foramen, located in the posterior half of the astragalus, are well evident in this view. In proximal view the astragalus has a quadrangular to discoidal perimeter. The proximal aspect is concave except where the ascending process is located. The ascending process is restricted to the most posterior region, being high and quadrangular in outline. Finally, the distal view shows a rough and strongly convex surface, contact area for the proximal surfaces of at least metatarsals II and III or III and IV. Unlike what occurs in Colossosauria, in Saltasauroidea there is no contribution of the fibula in the articulation between the zeugopodium and the autopodium.
The case of Titanomachya seems to be an intermediate case between the astragalus seen in Colossosauria and that seen in Saltasauroidea (). The astragalus of Titanomachya has a modified saltasauroid morphology. Its main modifications are its trapezoidal outline in an anterior view and its isosceles triangular outline in a proximal (or distal) view. Both outlines are so due to its symmetrical double wedge shape, not seen in any other titanosaur. This unique morphology of Titanomachya allows an articulation between zeugopodium and autopodium like that of Colossosauria, but with a contribution of the tibia and fibula similar to each other ().
Conclusions
Despite being represented only by limb elements, Titanomachya exhibits a unique combination of characteristics for a titanosaur from the Upper Cretaceous. Here, are highlighted its outstanding morphological features such as a haemal canal comprising 25% of the anterior chevron’s total length, prominent development of the humeral deltopectoral attachment, a straight or slightly curved humeral lateral margin, extremely reduced cnemial crests on the tibiae, a symmetrical development of the articular surfaces on the astragalus and a strong posterior bulge on the astragalus, among other distinguishing features.
During the Campanian-Maastrichtian interval, Patagonia exhibited a diverse range of titanosaurs. Northern Patagonia was dominated by aeolosaurines and saltasaurines, whereas Southern Patagonia was populated by large-bodied forms, such as Nullotitan (identified as a putative colossosaurian), Puertasaurus and Dreadnogthus. The presence of taxa such as Titanomachya to the south of the Nordpatagonian massif (Central Patagonia) could indicate the existence of a fauna composed of small titanosaurs (aeolosaurines and saltasaurines), coexisting with a fauna of giant titanosaurs confined to the south of the continent, resembling more closely the faunal association present in northern Patagonia.
Titanomachya, based on the known specimens, indicates a small adult size. Its estimated body mass ranges from 5.79 to 9.79 tons, with a mean body mass of 7.79 tons. This places Titanomachya in a size range similar to Saltasaurus and Neuquensaurus but heavier. The peculiar anatomy of Titanomachya‘s astragalus may be influenced by its greater body mass, although further evidence is needed to support this hypothesis.
The astragalus morphology varies among different titanosaur taxa. Colossosauria and Saltasauroidea exhibit distinct forms. Colossosaurian astragali are trapezoidal in anterior view, with a flat dorsal and lateral margins and a curved ventral margin. They have a subquadrangular perimeter in lateral view, with a sharp upward-directed ascending process. Saltasauroid astragali, on the other hand, are scalene triangular in anterior view, with a flat or slightly concave dorsal margin and a strongly curved ventral margin. They have a trapezoidal or wedge-shaped perimeter in lateral view and a robust ascending process. The astragalus of Titanomachya shows an intermediate morphology between Colossosauria and Saltasauroidea, with a modified Saltasauroid morphology characterised by a trapezoidal outline in anterior view and an isosceles triangular outline in proximal or distal view.
In conclusion, Titanomachya exhibits unique morphological characteristics among titanosaurian sauropods, and its small adult size places it in a similar range as Saltasaurus and Neuquensaurus but heavier. The astragalus of Titanomachya displays an intermediate morphology between Colossosauria and Saltasauroidea, allowing for an articulation between zeugopodium and autopodium similar to Colossosauria but with contributions from both the tibia and fibula. Further research is needed to fully understand the diversity and evolutionary significance of titanosaurian sauropods in Patagonia during the Upper Cretaceous and to confirm the relationship between body mass and the unique astragalus morphology observed in Titanomachya. Overall, Titanomachya stands out as an intriguing specimen with its distinctive characteristics and occupies a significant place in the evolutionary landscape of sauropod dinosaurs in Patagonia during the Late Cretaceous period.
Supplemental Material
Download MS Word (392.2 KB)Acknowledgments
We thank the students of the Tecnicatura en Paleontología of the Universidad del Chubut for preparing the fossil materials at the laboratory of the Museo Paleontológico Egidio Feruglio. A special thanks to Karen Panzeri, José O’Gorman, Agustín Ruella, Patricio Santamarina, Vince, and Becky Bruscas for technical assistance during the field work. The project was financed by PICT 2018-0605 and NGS 928922 R-22 (to DP). The paper was improved by the thoughtful comments and suggestions of Juan Canale, Pablo Gallina and Philip Mannion, for which we deeply appreciate their valuable contributions.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary data
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2024.2332997.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Adrian A. 2014. xlsx: read, write, format Excel 2007 and Excel 97/2000/XP/2003 files. Version 0.5.7. https://cran.r-project.org/web/packages/xlsx/index.html.
- Andrej-Nikolai S. 2014. qpcR: modelling and analysis of real-time PCR data. Version 1.4-0. https://cran.r-project.org/web/packages/qpcR/index.html.
- Apesteguía S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften. 91(10):493–497. doi: 10.1007/s00114-004-0560-6.
- Apesteguía S, Luzuriaga JES, Gallina PA, Granda JT, Jaramillo GAG. 2020. The first dinosaur remains from the Cretaceous of Ecuador. Cretaceous Res. 108:104345. doi: 10.1016/j.cretres.2019.104345.
- Aranciaga Rolando M, García Marsà JA, Agnolín FL, Motta MJ, Rozadilla S, Novas F. 2022. The sauropods record of Salitral Ojo de Agua: an Upper Cretaceous (Allen Formation) fossiliferous locality from northern Patagonia, Argentina. Cretaceous Res. 129(2022):105029. doi: 10.1016/j.cretres.2021.105029.
- Ardolino A, Busteros A, Cucchi R, Franchi M, Lema H, Remesal M. 1995. Cuerpos alcalinos básicos Paleógenos del sur de Somún Cura (Argentina) y su marco estratigráfico. Paleógeno de América del Sur. Asociación Paleontológica Argentina, Buenos Aires, Publicación Especial. 3:7e22.
- Ardolino A, Delpino D. 1987. Senoniano (continental-marino). Comarca Nordpatagónica, Provincia del Chubut, Argentina. X Congreso Geológico Argentino (Tucumán). Actas. 3:193e196.
- Bonaparte JF. 1986. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétosauridés) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentina). Annales de Paléontologie (Vert.-Invert.) 72(3):325–386.
- Bonaparte JF. 1996. Cretaceous tetrapods of Argentina. Münchner Geowissenschaften Abhandlungen. 30:73–130.
- Bonaparte JF, Coria RA. 1993. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Rio Limay (Albanio-Cenomaniano) de la Provincia del Neuquén, Argentina [A new and huge titanosaur sauropod from the Rio Limay Formation (Albian-Cenomanian) of Neuquén Province, Argentina]. Ameghiniana. 30(3):271–282.
- Bonaparte JF, Powell JE. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Mémoires de la Société Géologique de France, Nouvelle Série. 139:19–28.
- Borsuk-Bialynicka M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarzynskii gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontol Pol. 37:5–64.
- Bremer K. 1994. Branch support and tree stability. Cladistics. 10(3):295–304. doi: 10.1111/j.1096-0031.1994.tb00179.x.
- Buffetaut E, Suteethorn V, Cuny G, Tong H, Le Loeuff J, Khansubha S, Jongautchariyakul S. 2000. The earliest known sauropod dinosaur. Nature. 407(6800):72–74. doi: 10.1038/35024060.
- Calvo JO, Bonaparte JF. 1991. Andesaurus delgadoi gen. et. sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquen, Argentina. Ameghiniana. 28:303–310.
- Campione NE. 2017. Extrapolating body masses in large terrestrial vertebrates.Paleobiology. 43(4):693e699. doi:10.1017/pab.2017.9.
- Campione NE, Evans DC, Brown CM, Carrano MT. 2014. Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions. Methods Ecol Evol. 5:913e923.
- Carballido JL, Pol D, Otero A, Cerda IA, Salgado L, Garrido AC, Krause JM. 2017. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. Proc Royal Soc B Biol Sci. 284(1860):20171219. doi: 10.1098/rspb.2017.1219.
- Carballido JL, Sander PM. 2014. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. J Syst Paleontol. 12(3):335–387. doi: 10.1080/14772019.2013.764935.
- Carballido JL, Scheil M, Knötschke N, Sander PM. 2020. The appendicular skeleton of the dwarf macronarian sauropod Europasaurus holgeri from the Late Jurassic of Germany and a re-evaluation of its systematic affinities. J Syst Paleontol. 18(9):739–781. doi: 10.1080/14772019.2019.1683770.
- Casal G, Martínez RD, Luna CM, Ibiricu LM. 2016. Ordenamiento y caracterización faunisitica del Cretacico superior del Grupo Chubut, Cuenca del Golfo San Jorge, Argentina. Rev Bras Paleontol. 19(1):53–70. doi: 10.4072/rbp.2016.1.05.
- Casal G, Martínez R, Luna M, Sciutto JC, Lamanna M. 2007. Aeolosaurus colhuehuapensis sp.nov. (Sauropoda, Titanosauria) de la Formación Bajo Barreal, Cretácico Superior de Argentina. Rev Bras Paleontol. 10(1):53–62. doi: 10.4072/rbp.2007.1.05.
- Clyde WC, Krause JM, De Benedetti F, Ramezani J, Cúneo NR, Gandolfo MA, Haber P, Whelan C, Smith T. 2021. New South American record of the Cretaceous–Paleogene boundary interval (La Colonia Formation, Patagonia, Argentina). Cretaceous Res. 126:104889. doi: 10.1016/j.cretres.2021.104889.
- Coria RA, Filippi LS, Chiappe LM, Garcia R, Arcucci AB. 2013. Overosaurus paradasorum gen. et sp. nov., a new sauropod dinosaur (Titanosauria: Lithostrotia) from the Late Cretaceous of Neuquén, Patagonia, Argentina. Zootaxa. 3683(4):357–376. doi: 10.11646/zootaxa.3683.4.2.
- Curry Rogers K. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. J Vertebr Paleontol. 29(4):1046–1086. doi: 10.1671/039.029.0432.
- Curry Rogers KA, Forster CA. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature. 412:530–534.
- D’emic M. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zool J Lin Soc. 166:624–671. doi: 10.1111/j.1096-3642.2012.00853.x.
- Dong Z, Zhou S, Zhang H. 1983. Dinosaurs from the Jurassic of Sichuan. Palaeontologica Sinica, New Series C, Whole Number 162(23):1–136.
- Farris JS, Albert VA, Källersjö M, Lipscomb D, Kluge AG. 1996. Parsimony jackknifing outperforms neighbor-joining. Cladistics. 12(2):99–124. doi: 10.1111/j.1096-0031.1996.tb00196.x.
- Felsenstein J. 1985. Phylogenies and the comparative method. Am Nat. 125(1):1–15. doi: 10.1086/284325.
- Filippi LS, García RA, Garrido AC. 2011. A new titanosaur sauropod dinosaur from the Upper Cretaceous of North Patagonia, Argentina. Acta Palaeontol Pol. 56(3):505–520. doi: 10.4202/app.2010.0019.
- Filippi LS, Garrido AC. 2008. Pitekunsaurus macayai gen. et sp. nov., new titanosaur (Saurischia, Sauropoda) from Upper Cretaceous Neuquén Basin, Argentina. Ameghiniana. 45(3):575–590.
- Gallina PA, Apesteguía S. 2015. Postcranial anatomy of Bonitasaura salgadoi (Sauropoda, Titanosauria) from the Late Cretaceous of Patagonia. J Vertebr Paleontol. 35:e924957. doi: 10.1080/02724634.2014.924957.
- Gallina PA, Otero A. 2015. Reassessment of Laplatasaurus araukanicus (Sauropoda: Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. Ameghiniana. 52(5):487–501. doi: 10.5710/AMGH.08.06.2015.2911.
- García RA, Salgado L. 2013. The titanosaur sauropods from the late Campanian–early Maastrichtian Allen Formation of Salitral Moreno, Río Negro, Argentina. Acta Palaeontol Pol. 58(2):269–284.
- Gasparini Z, Sterli J, Parras A, O’Gorman JP, Salgado L, Varela J, Pol D. 2015. Late Cretaceous reptilian biota of the La Colonia Formation, central Patagonia, Argentina: occurrences, preservation and paleoenvironments. Cretaceous Res. 54:154–168. doi: 10.1016/j.cretres.2014.11.010.
- González Riga BJ. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana. 40(2):155–172.
- González Riga BJ, Lamanna MC, Ortiz David LD, Calvo JO, Coria JP. 2016. A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot. Sci Rep. 6(19165):1–15. doi: 10.1038/srep19165.
- González Riga BJ, Lamanna MC, Otero A, Ortiz David LD, Kellner AWA, Ibiricu LM. 2019. An overview of the appendicular skeletal anatomy of South American titanosaurian sauropods, with definition of a newly recognized clade. An Acad Bras Cienc. 91(Suplemento 2):e20180374. doi: 10.1590/0001-3765201920180374.
- Gonzalez Riga BJ, Mannion PD, Poropat SF, Ortiz David LD, Coria JP. 2018. Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships. Zool J Linn Soc. 184(1):136–181. doi: 10.1093/zoolinnean/zlx103.
- González Riga BJ, Previtera E, Pirrone CA. 2009. Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Cretaceous Res. 30(1):135–148. doi: 10.1016/j.cretres.2008.06.006.
- Hatcher JB. 1901. Diplodocus Marsh; its osteology, taxonomy, and probate habits, with a restoration of the skeleton. Mem Carnegie Mus. 1:1–63. doi: 10.5962/p.234818.
- Haughton SH. 1928. On some reptilian remains from the dinosaur beds of Nyasaland. Trans R Soc S Afr. 16(1):67–75. doi: 10.1080/00359192809519658.
- Huene FV. 1929. Los sauriquios y ornitisquios del Cretáceo argentino. Anales del Museo de La Plata, serie 2 3:1–196.
- Jain SL, Bandyopadhyay S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of central India. J Vertebr Paleontol. 17(1):114–136. doi: 10.1080/02724634.1997.10010958.
- Janensch W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten [Overview of the vertebrate fauna of the Tendaguru beds]. Archiv für Biontologie. 3:81–110.
- Kellner AW, Azevedo SD. 1999. A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. Natl Sci Mus Ser Mono. 15(111):e142.
- Kellner AWA, D. A. Campos D, Trotta MNF. 2005. Description of a titanosaurid caudal series from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museu Nacional, Rio de Janeiro. 63(3):529–564.
- Lacovara KJ, Lamanna MC, Ibiricu LM, Poole JC, Schroeter ER, Ullmann PV, Voegele KK, Boles ZM, Carter AM, Fowler EK, et al. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Sci Rep. 4:6196. doi: 10.1038/srep06196.
- Lapparent AFD, Zbyszewski G. 1957. Les dinosauriens du Portugal [The dinosaurs of Portugal]. Mémoires des Services Géologiques du Portugal, nouvelle série 2:1–63.
- Leanza HA, Apesteguıa S, Novas FE, de la Fuente MS. 2004. Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages. Cretaceous Res. 25(1):61–87. doi: 10.1016/j.cretres.2003.10.005.
- Leppe M, Stinnesbeck W, Frey H, Mansilla H, Vogt M, González E, Manríquez L, Cisternas K, Mihoc M, Jujihara T. 2014. Late Cretaceous terrestrial biota from Las Chinas-Cerro Guido complex, Magallanes Region, southern Chile: a key area for the Antarctic-South American biogeography. 23rd Latin American Colloquium on Earth Sciences, Abstracts; p. 101.
- Lydekker R. 1893. The dinosaurs of Patagonia. Anales del Museo de la Plata. 2:1–14.
- Mannion PD, Otero A. 2012. A reappraisal of the Late Cretaceous Argentinean sauropod dinosaur Argyrosaurus superbus, with a description of a new titanosaur genus. J Vertebr Paleontol. 32(3):614–638. doi: 10.1080/02724634.2012.660898.
- Mannion PD, Upchurch P, Schwarz D, Wings O. 2019. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zool J Linn Soc. 185(3):784–909. doi: 10.1093/zoolinnean/zly068.
- Marsh OC. 1878. Principal characters of American Jurassic dinosaurs. Am J Sci. 3(95):411–416. doi: 10.2475/ajs.s3-16.95.411.
- Martinelli A, Forasiepi A. 2004. Upper Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Río Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae). Rev Mus Argent Cienc Nat. 6(2):257–305. doi: 10.22179/REVMACN.6.88.
- Nixon KC, Wheeler QD. 1992. Extinction and the origin of species. Extinction and Phylogeny. 119:143.
- Nopcsa BF. 1915. Die dinosaurier der Siebenbürgischen landesteile Ungarns [Dinosaurs of the Transylvanian regions of Hungary]. Mitteilungen aus den Jahrbuch der Königlich Ungarnischen Geologischen Reichsanstalt. 23:1–24.
- Novas F, Agnolín F, Rozadilla S, Aranciaga-Rolando A, Brisson-Egli F, Motta M, Cerroni M, Ezcurra M, Martinelli A, D´angelo J, et al. 2019. Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina. Rev Mus Argent Cienc Nat. 21(2):217–293. doi: 10.22179/REVMACN.21.655.
- Novas F, Salgado L, Calvo JO, Agnolín F. 2005. Giant Titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. Rev Mus Argent Cienc Nat. 7(1):37–41. doi: 10.22179/REVMACN.7.344.
- Nowinski A. 1971. Nemegtosaurus mongoliensis n. gen., n. sp. (Sauropoda) from the uppermost Cretaceous of Mongolia. Palaeontol Pol. 25:57–58.
- Otero A. 2010. The appendicular skeleton of Neuquensaurus, a Late Cretaceous saltasaurine sauropod from Patagonia, Argentina. Acta Palaeontol Pol. 55(3):399–426. doi: 10.4202/app.2009.0099.
- Otero A. 2018. Forelimb musculature and osteological correlates in Sauropodomorpha (Dinosauria, Saurischia). PLoS One. 13(7):e0198988. doi: 10.1371/journal.pone.0198988.
- Otero A, Carballido JL, Pérez Moreno A. 2020. The appendicular osteology of Patagotitan mayorum (Dinosauria, Sauropoda). J Vertebr Paleontol. doi: 10.1080/02724634.2020.1793158.
- Otero A, Gallina PA, Canale JI, Haluza A. 2012. Sauropod haemal arches: morphotypes, new classification and phylogenetic aspects. Hist Biol. 24(3):243–256. doi: 10.1080/08912963.2011.618269.
- Paradis E. 2012. Analysis of Phylogenetics and Evolution with R. Vol. 2. Springer.
- Pascual R, Goin FJ, González P, Ardolino A, Puerta PF. 2000. A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals. Geodiversitas. 22:395e414.
- Pennell MW, Eastman JM, Slater GJ, Brown JW, Uyeda JC, FitzJohn RG, Alfaro ME, Harmon LJ, geiger v2. 2014. 0: an expanded suite of methods for fitting macroevolutionary models to phylogenetic trees. Bioinformatics. 30(15):2216e2218.
- Pérez Moreno A, Otero A, Carballido JL, Salgado L, Calvo JO. 2023. The appendicular skeleton of Rinconsaurus caudamirus (Sauropoda: Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. Cretaceous Res. 142:105389. doi: 10.1016/j.cretres.2022.105389.
- Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team. 2015. nlme: linear and nonlinear mixed effects models. R Package Version 3. 1e122. https://cran.rproject.org/web/packages/nlme/index.html.
- Pol D, Escapa IH. 2009. Unstable taxa in cladistic analysis: identification and the assessment of relevant characters. Cladistics. 25:515–527.
- Pol D, Goloboff PA. 2020. The impact of unstable taxa in coelurosaurian phylogeny and resampling support measures for parsimony analyses.
- Porfiri J, Calvo JO. 2010. Panamericansaurus schroederi gen. nov. sp. nov. un nuevo Sauropoda (Titanosauridae-Aeolosaurini) de la Provincia del Neuquén, Cretácico Superior de Patagonia, Argentina. Braz J Geol. 1(1):4.
- Powell JE. 1987. The Late Cretaceous Fauna from Los Alamitos, Patagonia, Argentina. Part. VI. The titanosaurids. Revista Museo Argentino Ciencias Naturales. 3(3):147–153.
- Powell JE. 1990. Epachtosaurus sciutto, un dinosaurio sauropodo del cretácico de Patagonia (Provincia de Chubut, Argentina). V Congreso Argentino de Paleontología y Bioestratigrafía. Universidad Nacional de Tucumán; p. 123–128.
- Salgado L. 1996. Pellegrinisaurus powelli nov. Gen. et sp. (Sauropoda, Titanosauridae) from the Upper Cretaceous of lago Pellegrini, northwestem Patagonia, Argentina. Ameghiniana. 33(4):355–365.
- Salgado L, Apesteguía S, Heredia SE. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous saltasaurine titanosaur from north Patagonia. J Vertebr Paleontol. 25(3):623–634. doi: 10.1671/0272-4634(2005)025[0623:ANSONA]2.0.CO;2.
- Salgado L, Azpilicueta C. 2000. Un nuevo saltasaurino (Sauropoda, Titanosauridae) de la provincia de Río Negro (Formación Allen, Cretácico Superior), Patagonia, Argentina. Ameghiniana. 37(3):259–264.
- Salgado L, Bonaparte JF. 2007. Sauropodomorpha. 188–228. In: Gasparini Z, Salgado L, Coria RA, editors. Patagonian Mesozoic reptiles. Bloomington (IN): Indiana University Press; p. 374.
- Salgado L, Carvalho IS. 2008. Uberabatitan ribeiroi, a new titanosaur from the Marília Formation (Bauru Group, Upper Cretaceous), Minas Gerais, Brazil. Palaeontology. 51(4):881–901. doi: 10.1111/j.1475-4983.2008.00781.x.
- Salgado L, Coria RA. 1993. El género Aeolosaurus (SAUROPODA, TITANOSAURIDAE) en la Formación Allen (CAMPANIANO-MAASTRICHTIANO), de la Provincia de Río Negro, Argentina. Ameghiniana. 30(2):1–10. Buenos Aires.
- Salgado L, Coria RA. 2009. Barrosasaurus casamiquelai gen. et sp. nov., a new titanosaur (Dinosauria, Sauropoda) from the Anacleto Formation (Late Cretaceous: early Campanian) of Sierra Barrosa (Neuquén, Argentina). Zootaxa. 2222(1):e16. doi: 10.11646/zootaxa.2222.1.1.
- Salgado L, Coria RA, Calvo JO. 1997. Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana. 34(1):3–32.
- Salgado L, Gallina PA, Paulina Carabajal A. 2014. Redescription of Bonatitan reigi (Sauropoda: Titanosauria), from the Campanian-Maastrichtian of the Río Negro Province (Argentina). Hist Biol. 27(5):525–548. doi: 10.1080/08912963.2014.894038.
- Santucci RM, Arruda-Campos AD. 2011. A new sauropod (Macronaria, Titanosauria) from the Adamantina Formation, Bauru Group, Upper Cretaceous of Brazil and the phylogenetic relationships of Aeolosaurini. Zootaxa. 3085(1):1–33. doi: 10.11646/zootaxa.3085.1.1.
- Santucci RM, Bertini RJ. 2006. A new titanosaur from western São Paulo State, Upper Cretaceous Bauru Group, south-east Brazil. Palaeontology. 49(1):59–66.
- Santucci RM, Filippi LS. 2022. Last Titans: titanosaurs From the Campanian–Maastrichtian Age. In: Otero A, Carballido J, Pol D, editors. South American Sauropodomorph Dinosaurs: record, Diversity and Evolution. Heidelberg: Springer; p. 341–392.
- Sanz JL, Powell JE, Le Loeuff J, Martinez R, Pereda-Suberbiola X. 1999. Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain). Titanosaur phylogenetic relationships. Estudios del Museo de Ciencias Naturales de Alava. 14(especial 1):235–255.
- Silva Junior JC, Martinelli AG, Marinho TS, da Silva JI, Langer MC. 2022. New specimens of Baurutitan britoi and a taxonomic reassessment of the titanosaur dinosaur fauna (Sauropoda) from the Serra da Galga Formation (Late Cretaceous) of Brazil. PeerJ. 10:e14333. doi: 10.7717/peerj.14333.
- Sterli J, Parras A, Albino A, Becerra MG, Carballido JL, Gouiric-Cavalli S, Muzzopappa P, Oriozabala C, Panzeri KM, Pérez Moreno A, et al. 2021 2022. Vertebrados continentales de la Formación la Colonia (Campaniano-Maastrichtiano). In: En Giacosa R, editor. Relatorio XXI CGA Geología y Recursos Naturales de la provincia del Chubut; p. 834–865.
- Vila B, Selles A, Moreno-Azanza M, Razzolini NL, Gil-Delgado A, Canudo JI, Galobart À. 2022. A titanosaurian sauropod with Gondwanan affinities in the latest Cretaceous of Europe. Nat Ecol Evol. 6(3):288–296. doi: 10.1038/s41559-021-01651-5.
- Wilson JA, Upchurch P. 2003. A revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution. J Syst Paleontol. 1:125–160. doi: 10.1017/S1477201903001044.