
ABSTRACT
Dental microwear (DMA) is a tool used for the palaeodiet reconstruction of animals in Archaeology. The use of this proxy on domestic ungulates provides valuable information to reconstruct livestock strategies, yet it presents several methodological limitations. Most studies have been carried out using low-magnification DMA and the interpretations often relied on comparisons with databases of extant wild ungulates. In addition, several studies have highlighted challenges in discerning diets in extant domestic caprines. In parallel, dental microwear texture analysis (DMTA) – a quantitative methodology based on 3D micro-texture height maps – has shown better discrimination. In this paper, we explore the capacity to distinguish four different management strategies of domestic goats (Capra hircus) and three species of wild ibexes (Capra nubiana, C. pyrenaica and C. ibex) using DMTA. Results revealed good discrimination among extant domestic goat populations and between wild and domestic goats. This new dataset was subsequently used to characterise the palaeodiet of archaeological goats from two Pre-Pottery Neolithic B sites in the southern Levant. Preliminary findings suggest evidence of human intervention in goats at least during the early 8th millennium BCE. In addition, incorporating various current management strategies has enhanced our understanding of early goat domestication in southern Levant.
Introduction
Sheep and goats played an important role in the late Prehistoric farming societies throughout the Mediterranean Basin. Since the Neolithic to the Iron Age, caprines are the predominant species – over pigs and cattle – in the archaeological faunal assemblages in many regions of the western and eastern Mediterranean (e.g. Halstead Citation1981, Citation2006; Saña Citation1998; Albarella et al. Citation2006; Albizuri et al. Citation2010; Trentacoste et al. Citation2018, Citation2021; Nieto-Espinet et al. Citation2020, Citation2021). Their major role in animal management since the Neolithic period makes caprines a valuable source of information to understand the husbandry practices in late Prehistory.
From traditional lines of evidence of zooarchaeology (e.g. taxonomic and anatomic representation and mortality profiles), it is possible to address issues such as the type of economical exploitation (e.g. meat or milk) (e.g. Stein Citation1987; Helmer and Vigne Citation2004; Helmer et al. Citation2005, Citation2007) or the composition of the herd (e.g. Grayson Citation1984, Davis Citation1987). However, some aspects of the livestock management practices such as animal food provisioning (e.g. type of pasture and foddering strategies), the type of exploitation (i.e. intensive or extensive) or the landscape use require further analyses.
The reconstruction of animal palaeodiet can provide relevant information about the strategies used by herders and farmers to feed animals and, therefore, on the type of pasture, the impact on the environment, the foddering strategies and the human intervention on animal diet in the early stages of animal domestication (e.g. Makarewicz et al. Citation2016; Yamada et al. Citation2018; Jiménez-Manchón et al. Citation2023). Stable isotope analysis, particularly of carbon isotope ratio (δ13C) from or dentine collagen, or tooth enamel, can be used to explore animal feeding strategies (e.g. Balasse Citation2002; Tornero et al. Citation2018; Navarrete et al. Citation2019). The carbon is incorporated from the food and water intake by the animal and the δ13C values vary according to the type of plants (e.g. Balasse Citation2002; Balasse et al. Citation2012). In addition, the integration of stable oxygen and carbon isotopes can provide insights into the seasonality of diet and water resources and climate (e.g. Makarewicz et al. Citation2017; Hadjikoumis et al. Citation2019; Trentacoste et al., Citation2020). There are, however, methodological limitations. This is a destructive method, requiring sampling preparation, which often results in a reduced number of analysed samples. The studies carried out on collagen are also constrained by its not always good preservation in e.g. arid regions like in the Near East (e.g. Grupe Citation1995; Miller et al. Citation2019). In contrast, studies performed on tooth enamel, which presents a better preservation than collagen, only record isotopic changes that occurred during the mineralisation of the tooth. Furthermore, isotopic signal can be affected by the enamel mineralisation process, which consists of two phases: secretion and maturation (Green et al. Citation2017).
Dental microwear analysis (DMA) is an additional powerful tool for palaeodietary reconstruction. This non-destructive tool is based on the analysis of microfeatures produced by ingested food items on the dental occlusal surface in the last weeks before the animal death (Walker et al. Citation1978; Teaford and Walker Citation1984; Gordon Citation1984; Grine Citation1986). This method is largely used for dietary reconstruction of wild ungulates from Pleistocene contexts to explore human hunting seasonality (e.g. Rivals and Deniaux Citation2005, Citation2005, Citation2015; Sánchez-Hernández et al. Citation2015), and ecological and environmental reconstruction (e.g. Calandra et al. Citation2016; Uzunidis Citation2020, Kubo and Fujita, Citation2021; Berlioz et al. Citation2022). However, the number of studies conducted on DMA on domestic ungulates is yet scarce, although in the last decade interest in palaeodiet reconstruction of domestic species has grown (e.g. Rivals et al. Citation2011; Rieau Citation2014; Mainland et al. Citation2016; Gallego et al. Citation2017, Citation2020; Jiménez-Manchón et al. Citation2019, Citation2020, Citation2023; Ibáñez et al. Citation2020; Lespes Citation2020; Sierra et al. Citation2021; Valenzuela-Suau et al. Citation2022). These works have shown several methodological limitations in the use of DMA on domestic ungulates. Firstly, most of these works have used Low-Magnification Dental Microwear Analysis (LMDA). This technique, based on the qualitative identification and quantification of microfeatures (i.e. scratches and pits) on the tooth surface, has been questioned about intra- and inter-observer error (Mihlbachler et al. Citation2012; De Santis et al. Citation2013), although this error is not statistically significant with well-trained observers (Semprebon et al. Citation2004; Semprebon and Rivals Citation2007). Secondly, most of the interpretations of the results obtained by LMDA have been carried out with a database spanning different taxa and groups including artiodactyls and perissodactyls (Solounias and Semprebon, Citation2002; Semprebon et al. Citation2004). In an attempt to overcome these limitations, several studies have tried to discriminate between different extant herd management strategies, mainly in domestic sheep, using reference collections (e.g. Rieau Citation2014; Ibáñez et al. Citation2020) or experimental works (Gallego et al. Citation2020). These studies have noted the high difficulty to discriminate extant diets in domestic species using this technique. In this sense, Ibáñez et al. (Citation2020) analysed two groups of extant ewes that were differently fed: one roaming on rangeland, combining Mediterranean forest and meadows, and the other on grasslands, using both LMDA and DMTA (Dental Microwear Texture Analysis). The results of this comparison showed better discrimination using DMTA compared to LMDA (Ibáñez et al. Citation2020).
DMTA is a technique based on quantitative analysis of the microtexture wear of the dental occlusal surface using confocal microscopy (Scott et al. Citation2005; De Santis et al. Citation2013). This method offers the advantage of gathering 3D surface texture representations at high magnifications from which it is possible to obtain quantitative parameters: ISO 25178 (International Organisation for Standardization 25178; Schulz et al. Citation2010; Purnell et al. Citation2012; Calandra et al. Citation2012) and SSFA parameters (Scale-Sensitive Fractal Analysis; Scott et al. Citation2005, Citation2006). Unlike previous techniques, (1) DMTA is a reproducible method that reduces intra- and inter-oberver errors (De Santis et al. Citation2013), and (2) it is a technique with which it is easier to discriminate different diets of domestic animals (Ibáñez et al. Citation2020).
In spite of the advantages that this technique offers to domestic animal palaeodiet reconstruction, its use on extant domestic ungulates is still uncommon (e.Merceron et al. Citation2016; Ramdarshan et al. Citation2017; Ackermans et al. Citation2020; Louail et al. Citation2021). These works were conducted in controlled-feeding experiments, primarily focusing on specific factors such as the effect of dust and grit on dental microwear. Consequently, the diet of these domestic ungulates was entirely controlled in experimental settings and does not reflect the diversity of animals in current or past management strategies. Therefore, studies that could characterise and discriminate diets of extant management strategies using DMTA are missing. If successful, such studies will then be applied to archaeological contexts.
In a previous work (Jiménez-Manchón et al. Citation2023), domestic goats that fed in wooded and overgrazed areas were successfully discriminated from various species of wild ibexes using DMTA. The use of a discriminant analysis to a collection of early Neolithic goats from Tell Qarassa North (southern Syria) and Kharaysin (northern Jordan) indicated that some individuals from the second site show similar pattern that the domestic goats. This information, besides the high variability in DMTA values for Kharaysin, suggested that human intervention in goat feeding could have taken place there at the beginning of the 8th millennium cal. BCE or even before. However, one major issue limited the scope of this preliminary, though promising, study. Only one type of domestic management was included in the predictive model, so it was not clear how various feeding strategies of domestic goats might affect the model’s ability to discriminate wild and domestic diets. Furthermore, it was not possible to determine the specific type of management strategy that herders may have been employed to feed goats.
The present work makes a significant contribution on this topic. In this paper, we explore the use of DMTA to characterise and discriminate different management strategies of four modern domestic goat populations with known diets. Our research focused on exploring how dental microwear patterns vary according to pasture system. In addition, the four management strategies of domestic goats were compared with the diet pattern of three different wild species. The main objective is to explore potential similarities and differences among these goat populations. The models derived from DMTA were then used to characterise the dietary patterns of goats (Capra cf. aegagrus) from two Pre-Pottery Neolithic sites in the southern Levant: Kharaysin (Zarqa, Jordan, ca. 8800–7500 cal. BCE) and Tell Qarassa (As-Suweida, Syria, ca. 8700–8200 cal. BCE).
We used two types of parameters: SSFA and ISO 25178. On the one hand, SSFA is a method to quantify the micro-texture surface by analysing the changes across scales. Three parameters have been considered: area-scale fractal complexity (Asfc), length-scale anisotropy of relief (epLsar) and heterogeneity of complexity (HAsfc). On the other hand, 30 standardised ISO parameters related to height, volume, area and function, used to analyse 3D surface texture, were considered. In this work, in addition to the methodology commonly used to characterise animal diet using SSFA and ISO parameters, we explored alternative methods to present ISO parameters such as bivariate graphs.
Consequently, this paper delves into the potential of DMTA in discriminating the dietary behaviours of modern wild and domestic goats with known diets and geographic origin. The development of this database through DMTA constitutes a valuable tool for interpreting dental microwear in archaeological and palaeontological contexts. Furthermore, we investigated the results obtained to characterise the diet of ancient goat populations, in order to enhance our understanding of the early husbandry practices in the Near East.
Material and methods
The reference collection
A total of 94 specimens have been analysed, of which 51 were domestic and 43 wild goats. The domestic goats (Capra hircus) were managed according to traditional systems but with different exploitations. They came from four different locations with different management strategies: the first group (CH1) of goats fed in the northern Saharan steppe (Algeria), the second group (CH2) in wooded and overgrazed areas in southern Catalonia (north-eastern Iberian Peninsula), the third one (CH3) on grasslands in the southern side of the Pyrenees (north-eastern Iberian Peninsula), and the last one (CH4) on wooded areas in the Causse of Larzac (southern France).
The wild specimens corresponded to three other species: Capra nubiana (CN) from the Near East; Capra ibex (CI) from the Alps; and Capra pyrenaica (CP) from the southern Iberian Peninsula (). No goats were deliberately killed for this study. The wild goats were from the osteological collection of several museums, and the domestic ones either naturally died or the mandibles were recovered from slaughterhouses or breeders.
Figure 1. Goat species, status, dietary regime and management strategy, geographical location and climate for each modern goat group. For CH1 and CH2, the circles in the dietary regime/management column indicate the intake of dust/grit.
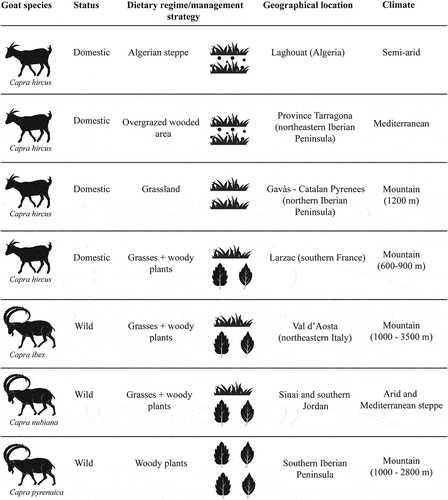
CH1. Extensive grazing on steppe vegetation (northern Sahara, Algeria)
The collection consisted of 18 adult female domestic goats of the Arabia breed, the most important local breed in Algeria (Manallah Citation2012). The specimens came from the region of Laghouat (northern Sahara, Algeria) (Ridouh Citation2021). The region is dominated by steppes with a semi-arid climate, experiencing increasing desertification and degradation of pastures (Belala et al. Citation2018), with average temperatures ranging from 10º to 34ºC. These goats are exploited for meat and they do not receive additional supplies of fodder. Regarding the diet, goats grazed in steppe vegetation which is mainly composed of grasses such as feather grass (Stipa tenacissima) and esparto grass (Lygeum spartum) (Dalila and Slimane Citation2008). The season of death is unknown for all individuals.
CH2. Extensive grazing on wooded and overgrazed areas (southern Catalonia, north-eastern Iberian Peninsula)
A collection of 15 domestic goats (four males and 11 females) of the Blanca de Rasquera breed was sampled. Four specimens are mixed with the Granadino-Murciana breed. The goats came from two close areas in southern Catalonia presenting the same environment (province of Tarragona). The vegetation of this environment is composed of woody plants: holm oak (Quercus ilex), hermes oak (Quercus coccifera), rosemary (Salvia rosmarinus), heather (Erica sp.), scorpion broom (Genista scorpius), shrubby hare’s-ear (Bupleurum fruticosum), wild madder (Rubia peregrina), juniper (Juniperus communis) and grass plants: nut grass (Cyperus rotundus) and spiny-head mat-rush (Lomandra longifolia) (Jiménez-Manchón et al. Citation2023). The flock grazed in a similar area throughout the year and, consequently, goats were fed in an overgrazed environment. These goats did not receive additional supplies of fodder and feed on available vegetation in wooded areas (500–1000 m). Given that the availability of woody plants is poor in this overgrazed environment, the intake of herbaceous plants by goats was dominant (Jiménez-Manchón et al. Citation2023). The specimens were collected in spring (n = 3), summer (n = 9), and autumn (n = 3) 2021.
CH3. Extensive mountain grazing on grasses (southern side of Pyrenees, northern Iberian Peninsula)
Three female domestic goats of Alpine breed were sampled. They originate from Gavàs, located on the southern side of the Pyrenees (Pallars Sobirà, northeastern Spain), and were devoted to milk exploitation. The goats died in winter 2021. During this season, they had seven hours per day of pasture in the meadow, feeding only on grasses (e.g. Arrhenatherion elatioris associations) in high mountain areas (>1200 m) with average winter temperatures ranging between -7º and 2°C (López-i-Gelats et al. Citation2015). They received a small amount of supplement of uncultivated meadow grasses.
CH4. Extensive browsing on wooded areas (Larzac, southern France)
A set of 15 domestic goats coming from the Causse of Larzac (southern France) was sampled. The sex and the season of death are unknown for all individuals. The flock browsed in wooded areas (600–900 m) on the plateau of Causse of Larzac, with a supramediterranean bioclimate. In this area, woody plants dominate, such as holm oak (Quercus ilex), pubescent oak (Quercus pubescens), beech (Fagus sylvativa), and hazelnut (Corylus avellane), along with the presence of grasses (Prioton Citation1970). The domestic goats did not receive additional supplies of fodder.
CN. Capra nubiana (Sinai Peninsula and southern Jordan)
We sampled 12 adult Nubian ibexes (three females, seven males and two of unknown sex) from three localities (i.e. Wadi Ihmed, Wadi Adani and Wadi Leithih) from the Sinai Peninsula (eastern Egypt) and the region of Petra (southern Jordan). The specimens, curated at the Zoological Museum of Strasbourg in France, were collected between 1893 and 1911 between spring (n = 2) and summer (n = 3). The season of death of the rest of the individuals (n = 7) is unknown. The region is characterised by heterogeneous landscapes, transitioning from arid to steppe Mediterranean woodlands. In the arid regions, vegetation is scarce, and it is dominated by shrubs (Haloxylon persicum and Retama raetam) with patches of Acacia. In the steppe regions, there is a cover of grasses and Artemisia plants, as well as woody plants such as Moringa, juniper (Juniperus), Pistacia and Phoenix (Palmer et al. Citation2007). The Nubian goat population had a flexible diet, with a tendency to grazing (Hackham and Ritte Citation1991).
CI. Capra ibex (Val d’Aosta, northern Italy)
Twelve adult Alpine ibexes (five females and seven males) from Val de Cogne in Val d’Aosta (north-western Italy) were analysed. The specimens, curated at the Zoological Museum of Strasbourg in France, were collected in 1897. The season of death is unknown for all specimens. Alpine ibexes can live at elevations ranging from 1000 to 3500 m. The vegetation is characterised by a gradient based on altitude. Above 1800 m, the vegetation is rich in woody plants such as Pinus sylvestris, Laric decidua and Betula alba. To a lesser extent, there are grasses and other herbaceous plants. Below 1800 m, meadows (Avenula pubescens, Gentiana lutea and Galium glaucum) and steppe-like communities (Centaurea scabiosa, Galium verum) are more common (Pini et al. Citation2017). This species has a diet dominated by grasses and other herbaceous plants, although their diets may vary according to the season (Couturier Citation1962; Zingg Citation2009; Parrini et al. Citation2009).
CP. Capra pyrenaica (southern Iberian Peninsula)
A set of 19 adult Iberian ibexes (14 females and five males) from Sierra Nevada and Sierra de Cazorla (southern Iberian Peninsula), curated at the Estación Experimental de Zonas Áridas-CSIC (Almería, Spain), the Estación Biológica de Doñana-CSIC (Jaca, Spain) and the Parque Nacional de Sierra Nevada (Granada, Spain), were sampled. The osteological collection was collected between 1987 and 2007 in spring (n = 7), summer (n = 7), and autumn-winter (n = 3). The season of death is unknown for three individuals. The vegetation depends on the altitude. Above 2700 m, woody plants such as Festuca clementei and Arenaria nevadensis dominate. Below, there are Juniperus communities, Pinus sylvestris, Thyme-pasture communities and herbaceous plants. This goat species tends to consume ligneous plants, such as Quercus ilex, Pinus nigra and Thymus zygis. Among the grasses, the consumption of Brachypodium stands out (Martinez et al. Citation1985; Cuartas and García-González Citation1992; Cuartas et al. Citation1996; Martínez Citation2000).
Archaeological materials
The archaeological material studied originated from two Pre-Pottery Neolithic sites situated in the southern Levant: Kharaysin (Zarqa, Jordan) and Tell Qarassa (As-Suweida, Syria) (). Kharaysin is a large site around 25 ha and it is located in northwestern Jordan (Ibáñez et al. Citation2015). Excavations conducted at Kharaysin have revealed four chronological sequences: 1) Late Pre-Pottery Neolithic A (PPNA, beginning of the 9th millennium cal. BCE), 2) Early Pre-Pottery Neolithic B (EPPNB, second half of the 9th mill. cal. BCE), 3) Middle Pre-Pottery Neolithic B (MPPNB, beginning of the 8th mill. cal. BCE), and 4) Late Pre-Pottery Neolithic B (LPPNB, end of the 8th mill. cal. BCE). The preliminary zooarchaeological study indicates the progressive increase in the frequency of goats at the expense of gazelles from the lower to the upper layers. In light of these findings, the results obtained from a DMTA analysis suggest the possible coexistence of managed and wild goats during the MPPNB phase (Jiménez-Manchón et al. Citation2023). In this work, we selected a total of 13 goats from Kharaysin, excluding specimens affected by taphonomical alterations.
Tell Qarassa, situated in the southern Syria, is an Early Pre-Pottery Neolithic B site occupied between c. 8700 and 8200 cal. BCE (Ibáñez et al. Citation2010). The excavations conducted in 2009 and 2010 revealed buildings dating from the mid-9th millennium cal. BCE. According to the zooarchaeological analysis, goats prevailed, followed by other ungulates such as gazelles, wild boars and aurochs (Ibáñez et al. Citation2010). DMTA was performed on three goat specimens that did not exhibit taphonomical alterations.
The palaeoenvironmental data available for the southern Levant revealed the existence of varied ecological niches (e.g. Asouti et al. Citation2015). Kharaysin and Qarassa regions were characterised by a combination of wooded steppe components and riparian taxa (Arranz-Otaegui et al., Citation2017). Notably, eudicotyledonous plants like Pistacia, Amygdalus and Quercus, and, to a lesser extent, grasses were present in the surrounding vegetation.
Dental microwear analysis
The occlusal surface of the mandible teeth was carefully cleaned using acetone and 96% ethanol. For this purpose, we used cotton wool to avoid damaging the surface. Then, we moulded the occlusal surface using high-precision polyvinylsiloxane (President Light Body, Coltene Waledent). Two moulds were taken: the first one was made to remove the remaining dust and was discarded, and the second was kept for DMTA analysis.
DMTA was conducted on the mesio-buccal facet of the second lower molar (M2) protoconid (). A single facet was selected since the inclusion of other may increase the sample variance and may make differenciation between dietary groups difficult (Ramdarshan et al. Citation2017). When it was not possible, the analysis was extended to the hypoconid. The moulds were analysed with a white-light scanning confocal microscope (Sensofar Plu Neox) using a ×50 (0.80 NA) objective housed at the Institució Milà i Fontanals-CSIC (Barcelona, Spain). For each facet, an area of 150 × 150 μm was selected and measured using MountainsMap 7 software (Digital Surf) (). The extreme measured points were eliminated (upper and lower 0.5%) to remove the outliers. The tooth occlusal surface generally has a curved morphology. The least-square (LS) plan method was used to correct the lack of horizontality of the sample. Consequently, the form was eliminated using a polynomial function (power 8; Francisco et al. Citation2018). Archaeological teeth with taphonomic features were excluded from the study based on previous studies (Uzunidis et al. Citation2021; Weber et al., Citation2022).
Figure 2. Exemplary occlusal view from a right hemimandible of goat, and topography texture enamel surface (false colour elevation map) of mesiobuccal facet of a second lower molar protoconid.
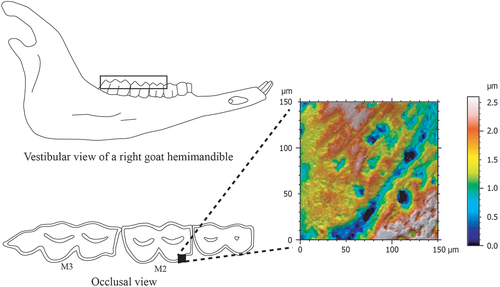
SSFA and ISO 25178 parameters were obtained from Mountains 7 (). Firstly, three SSFA parameters were measured: complexity (Asfc), anisotropy (epLsar) and heterogeneity of complexity calculated through 9 cells (HAsfc9) (Scott et al. Citation2005, Citation2006). Secondly, a total of 30 ISO parameters were considered (Table S1). These parameters are pooled to height (e.g. Sq, Sp and Sa), function (Smr), space (e.g. Str), volume (e.g. Vv, Vmp) and area (Sda and Sha).
Table 1. Description and functional groups of the ISO 25,178 and SSFA parameters.
Statistical analysis
According to the objectives of this study, two datasets have been analysed: the first focused only on the diet discrimination of four domestic populations, while the second included both the four domestic and the three wild populations. For both datasets, the statistical procedure was as follows.
These values were explored using bivariate plots, indicating the mean and standard error of mean, similarly to previous works (e.g. Ramdarshan et al. Citation2017). The presence of significant differences was tested through ANOVA’s test and Tukey’s pairwise comparison. ISO Parameters were log-transformed to increase normality and homoscedasticity (Francisco et al. Citation2018). The presence of significant differences (p ≤ 0.05) among the four groups of domestic goats, and the seven groups of wild and domestic goats was tested using an analysis of variance (ANOVA). When homogeneity of variance (Levene’s test) showed significant differences, Welch’s ANOVA was used. A Linear Discriminant Analysis (LDA) with cross-validation, based on parameters that present significant differences, was constructed. When Box’s M was violated (p ≤ 0.05), a Quadratic Discriminant analysis (QDA) was performed. The significance of discriminant analysis was assessed using Wilk’s Lambda (p ≤ 0.05). Consequently, the predictive model obtained for wild and domestic goats was used to classify the archaeological specimens from Kharaysin and Qarassa sites.
We explored the use of bivariate graphs based on 25178 ISO parameters. For this purpose, a Tukey’s post-hoc test based on ISO variables was carried out on the seven groups of modern wild and domestic goats (Table S5). The parameters showing the largest differences in the four main categories (height, function, volume and feature) of 25178 ISO parameters were selected. The statistical procedures were carried out using IBM SPSS 26 statistics software®.
Results
Dental microwear pattern of domestic goat management strategies
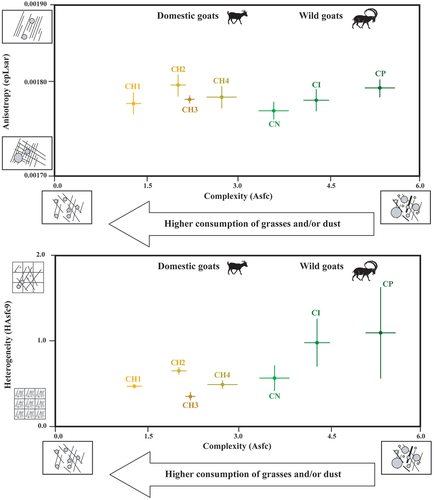
SSFA parameters were explored using two bivariate plots (). ANOVA’s test detected significant differences in complexity (Asfc) (Table S1.1). No significant differences were found in heterogeneity (HAsfc9) and anisotropy (epLsar). Goats managed in the Laghouat steppe (CH1) presented the lowest complexity values (Mean Asfc = 1.14), whereas goats managed in the more wooded vegetation (CH4) presented the highest (Mean Asfc = 2.6). The groups CH2 and CH3 (i.e. goats managed in overgrazed and grassland areas) presented intermediate values of complexity (). These results indicate that the higher consumption of woody plants by goats from the Causse of Larzac increases complexity (Asfc), and the higher intake of grasses and dust/grist by goats from the Laghouat steppe reduces it. Concerning heterogeneity (HAsfc9) and anisotropy (epLsar), although no significant differences were found, goats managed in grasslands (Pyrenees) showed the lowest values for both parameters. It is worth noting that goats grazing in overgrazed vegetation (CH2) presented higher values than the population fed on steppe areas (CH1) ( and ).
Figure 3. Biplots (mean and standard error of the mean) of complexity (Asfc), anisotropy (epLsar), heterogeneity of complexity (HAsfc9) of the four management strategies of domestic goats (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac).
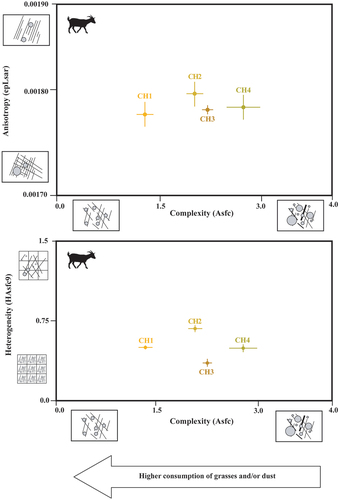
Table 2. Mean (Mn), standard error of the mean (SEM) and standard deviation (SD) values of SSFA textural parameters of wild and domestic extant goats. Asfc: complexity; epLsar: anisotropy; HAsfc9: heterogeneity of complexity.
A Quadratic Discriminant Analysis was performed on the basis of the ISO parameters that significantly differ (Table S1.2). The predictive model showed a correct cross-validation classification of 74.5% of the individuals. Goats managed in grassland in the Pyrenees (100%) were the best classified. The other groups were similarly classified between 72.2% and 73.3% ().
Table 3. Classification of extant domestic specimens using the classificatory rule obtained from a Quadratic Discriminant Analysis (QDA).
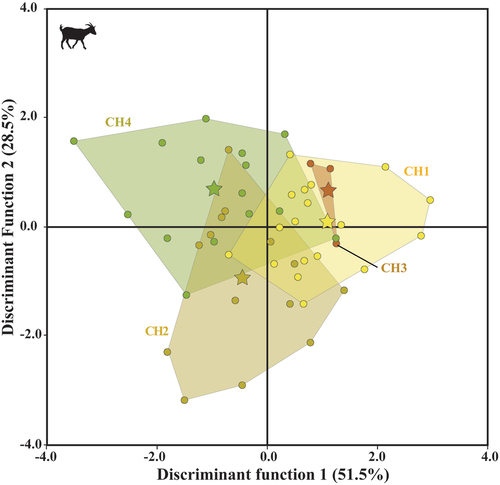
In the QDA texture-dietary plot (Discriminant Function 1 = 51.5%, Discriminant Function 2 = 28.5%), goats managed in Algerian steppe and overgrazed areas generally occupied positive values along the Discriminant Function 1 (), while goats that grazed in grassland vegetation (Pyrenees) and in the Causse of Larzac had negative values. According to the discriminant coefficients (Table S1.2), the texture surface of goats managed in Laghouat steppe and overgrazed vegetation is characterised by less volume (Vm) and roughness (e.g. Sq).
Figure 4. Quadratic discriminant analysis of ISO parameters of domestic modern goats. The star represents the mean for each group (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac).
Dental microwear pattern of wild ibexes and goat management strategies
SSFA values of wild ibexes and domestic goats are presented in . ANOVA’s test detected significant differences in complexity (Asfc) and heterogeneity of complexity (HAsfc9), with no differences in anisotropy (epLsar) (Table S1.3). All three groups of wild ibexes showed higher complexity (Asfc) values than domestic goats (3.34 ≤ Mean ≥ 5.0). Among the wild ibexes, CN had the lowest complexity values and CP presented the highest ones. Concerning heterogeneity, ibexes from the Near East (CN) and the Alps (CI), characterised by a great intake of ligneous plants compared to the other wild goat populations, presented the lowest values. In contrast, domestic goat managed in grassland (CH3) had the lowest values. The other goat groups displayed intermediate values ( and ). The Tukey’s pairwise comparison revealed significant differences in complexity (Asfc) among various groups of and domestic goats (S5), such as CI and CH2, CN and CH2, CP and CH4. Significant differences have only been found between two groups of domestic goats (CH4 and CH1). Regarding heterogeneity (HAsfc9), differences have been observed between modern groups of wild and domestic goats (i.e. CN and CH3, CP and CH3, CP and CH4, CN and CH4).
Figure 5. Biplots (mean and standard error of the mean) of Complexity (Asfc), anisotropy (epLsar), heterogeneity of complexity (HAsfc9) of the four management strategies of domestic goats and wild ibexes (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac; CI= Capra ibex; CN= Capra nubiana; CP= Capra pyrenaica).
A Quadratic Discriminant analysis was performed based on the ISO parameters that significantly differ between the 7 groups (Table S1.3). 72.3% of individuals were correctly classified according to the QDA (). Domestic goats managed in grasslands (CH3) and wild goats from the Near East (CN) were the best classified 100% and 83.3% respectively). In contrast, domestic goats managed in the Laghouat steppe (61.1%) and in overgrazed areas (66.7%) were more poorly classified ().
Table 4. Classification of extant wild and domestic specimens using the classificatory rule obtained from a Quadratic Discriminant Analysis (QDA).
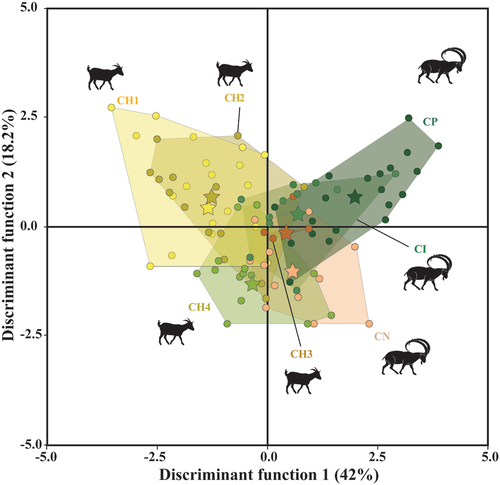
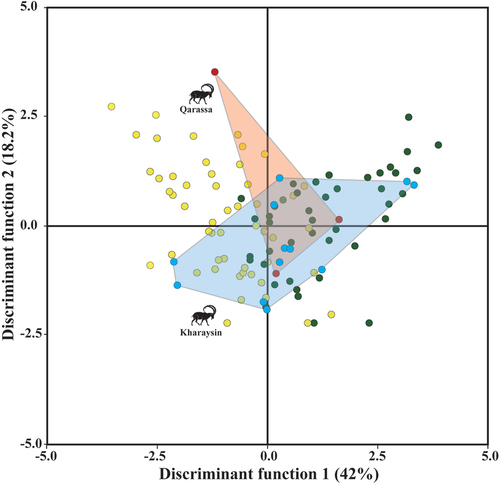
The goat groups occupied different areas of the QDA texture-dietary plot (Discriminant Function 1 = 42%, Discriminant Function 2 = 18.2%), some of them overlapping each other (). Overall, the mean of the four domestic groups presented negative values along axis 1 (excepting CH3), while the three ibex groups had positive values. Goats managed in the Laghouat steppe and overgrazed areas presented the lowest values. According to the discriminant coefficients (Table S1.4), specimens with negative values showed less rough dental texture surface than individuals that plot in positive values. The ibexes generally had more positive values than domestic goats, suggesting a rougher and more complex surfaces. Among the wild goats, the ibexes from the southern Iberian Peninsula displayed the roughest dental surface compared to the other groups.
Figure 6. Quadratic discriminant analysis of ISO parameters of domestic and wild extant goats. The star represents the mean for each group (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac; CI= Capra ibex; CN= Capra nubiana; CP= Capra pyrenaica).
Differentiation of dietary regimes using ISO bivariate graphs
According to the Tukey’s post-hoc analysis based on ISO 25178 (Table S1.5), the variables with the most significant differences across the four main categories of ISO parameters (height, function, volume and feature) were selected. There are Spk (function), Sq (height), Vmp (volume) and Spc (feature). Subsequently, they were combined in six bivariate graphs ().
Figure 7. Biplots of the mean five most discriminant ISO parameters of domestic and wild extant goats and their respective standard errors (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac; CI= Capra ibex; CN= Capra nubiana; CP= Capra pyrenaica).
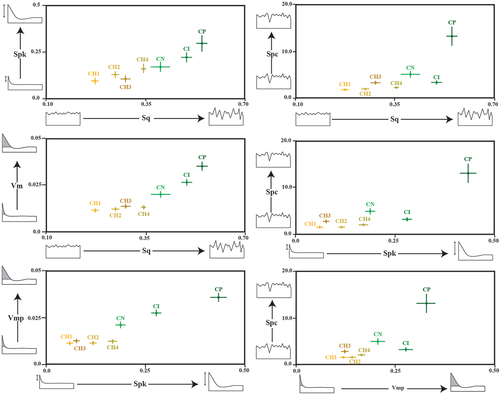
Domestic goats had low values for height (Sq), volume (Vmp), function (Spc) and feature (Spk), indicating more flattened dental texture surfaces than those of wild goats. Among the four groups of domestic goats, those managed in Laghouat (CH1) had the lowest values for all four parameters. In contrast, goats that grazed in wooded areas (CH4) showed higher values than the previous ones. Goats managed in grasslands (CH3) and overgrazed areas (CH2) showed intermediate values. These data indicate that the higher intake of ligneous plants increases the values of height, volume, feature and function. Among the three groups of wild ibexes, CP showed the highest values for all four parameters. Excepting for Spc parameter, CN displayed the lowest ones, and CI intermediate values.
Archaeological application
SSFA parameters (Asfc, epLsar and HAsfc9) were extracted from the archaeological specimens of Tell Qarassa (n = 3) and Kharaysin (n = 13) (). The obtained values were incorporated into the reference database (). In terms of complexity (Asfc), the archaeological goat values are very close to those of the domestic goats with a more diverse diet (CH4) and those fed on grasses (CH3). Heterogeneity values (HAsfc9) are higher compared to the domestic groups and closer to the wild ibexes, particularly to CP and CN. As for anisotropy (epLsar), the values from Kharaysin resemble those of both domestic and wild goats. However, it is worth noting that the values of Qarassa are much lower than any of the modern goats studied.
Figure 8. Biplots (mean and standard error of the mean) of Complexity (Asfc), anisotropy (epLsar), heterogeneity of complexity (HAsfc9) of the four management strategies of domestic goats, wild ibexes and archaeological specimens from Kharaysin and Qarassa (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac; CI= Capra ibex; CN= Capra nubiana; CP= Capra pyrenaica).
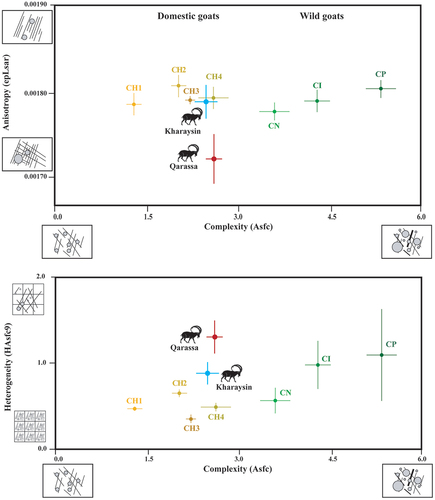
Table 5. Mean (Mn), standard error of the mean (SEM) and standard deviation (SD) values of SSFA textural parameters of archaeological specimens from Tell Qarassa and Kharaysin. Asfc: complexity; epLsar: anisotropy; HAsfc9: heterogeneity of complexity.
Subsequently, the archaeological specimens were incorporated into the QDA obtained from the reference collections of modern wild and domestic goats (). Among the goats from Qarassa, one exhibits negative values for DF1, another displays positive values, and the third one shows intermediate values. Regarding the goats from Kharaysin, the specimens showed a high degree of variability. Specifically, three individuals have negative values for DF1, another three displayed positive values, and the remaining seven plot near ‘0’.
Figure 9. Quadratic discriminant analysis based on modern domestic (yellow) and wild goats (green), and archaeological specimens from Kharaysin (blue) and Qarassa (red) (CH1= goats managed in the Algerian steppe; CH2= in wooded and overgrazed areas in the northeastern Iberian Peninsula; CH3= in grasslands in the Pyrenees; CH4= in wooded areas in the Larzac; CI= Capra ibex; CN= Capra nubiana; CP= Capra pyrenaica).
The predictive model obtained for the four domestic goat groups and the three wild goat species using the QDA was used to classify the archaeological specimens (). 58.3% of individuals from Kharaysin were classified within one of the wild goat groups. Specifically, four individuals were classified with dental abrasions comparable as those of Capra nubiana (CN), and three were classified as Capra ibex (CI), indicating a mixed diet with tendency to grasses (see 2.1.5 and 2.1.6). 41.7% of individuals were classified within the domestic groups. Among these, three showed a diversified diet similar to CH4, two have tooth abrasions comparable to those found in goats that fed in wooded and overgrazed areas (CH2), and one was classified as goats fed in steppe (CH1). Concerning the archaeological specimens from Qarassa, two individuals were each classified within a different wild species group (i.e. Capra nubiana and Capra ibex), and only one within one domestic goat group. Specifically, it was classified within the group of domestic goats that fed in mixed vegetation (CH4).
Table 6. Classification of each archaeological specimen using the classificatory rule obtained from a Quadratic Discriminant Analysis (QDA).
Discussion
This study presents the dental microwear texture analysis of 51 domestic goats and 43 wild ibexes. Domestic goats came from four different localities and were managed differently with contrasted grassland-woodland intakes; and the wild ibexes came from three geographical areas where they fed on the vegetation available in their environments. In this work, we explored whether, and how, DMTA can discriminate these modern goat groups. For this purpose, we combined SSFA and ISO parameters, which allow a correct differentiation among diet groups (Ackermans et al. Citation2020). This work represents the largest database of domestic ungulates, excluding controlled feeding experiments, for palaeodiet reconstruction using DMTA.
Discrimination of domestic goats
The results presented in this work demonstrate that tooth wear textures vary according to management strategies. Domestic goats fed in the Laghouat steppes (CH1) showed the lowest values for complexity (Asfc), height (e.g. Sq) and volume (e.g. Vmp), and presented the roundest and smallest peaks (Spc and Spk). This degree of dental abrasion can be explained by their high consumption of grass plants, rich in phytolithes (see section 2.1.1). These results are consistent with previous works, which have shown that a diet composed of monocot plants (e.g. grasses) generates low values of complexity and a less rough dental texture wear (Scott et al. Citation2005, Citation2006, Citation2012; Ungar et al. Citation2012; Merceron et al. Citation2016; Ramdarshan et al. Citation2016; Francisco et al. Citation2018). In addition, the Laghouat steppe is an arid environment (e.g. Belala et al. Citation2018) and, consequently, goats potentially consumed high proportions of dust and/or grit. The presence of small exogenous particles in the diet can lead to less complex and less rough surface (e.g. Ramdarshan et al. Citation2016; Merceron et al. Citation2016; Winkler et al. Citation2020; Ackermans et al. Citation2020; Louail et al. Citation2022). Therefore, goats herding in environments such as the steppe without supplies of fodder, where grasses and dust/grit consumption is important, display dental wear with very little roughness and complexity, and rather a flattened surface.
The groups of domestic goats grazing in an overgrazed environment (CH2) and grassland areas (CH3) also exhibited relatively low values for complexity, volume and roughness, although higher than those of the previous group of domestic goats (CH1). The differences observed between goats fed in Laghouat steppe (CH1) and those in grasslands and overgrazed areas (CH2 and CH3) can be attributed to the higher loads of dust and grit in the steppe environment. Another explanation can be the variation in the degree of humidity among different vegetation types. The limited moisture content of plants in the Laghouat steppe may lead to a more abrasive dental microwear pattern (Winkler et al. Citation2021). In contrast, domestic goats from the Pyrenees (CH3) and the northeastern Iberian Peninsula (CH2) were managed in pastures with higher humidity levels. There is not available reference collection with animals fed with dry and wet food. However, despite the difference in the materials analysed, a texture analysis of lithic 3D use-wear conducted on a reference collection revealed that harvesting dry cereals generated more abrasive use-wear compared to wet cereals (Ibáñez et al. Citation2021). Future research could encompass domestic caprines managed pastures with different degrees of humidity to explore the potential impact of this factor on DMTA.
Goats managed in the Causse of Larzac (CH4) had a mixed diet consisting of monocotyledonous and eudicotyledonous plants. This is thus the group of domestic goats which ingested the highest proportion of eudicotyledonous plants among the groups analysed in this work. Their diet resulted in a tooth wear more complex and rougher than the other domestic goat groups. These results are coherent with previous works that attested that the intake of eudicotyledone herbaceous plants (clover fodder) causes dental wear rougher and more complex (e.g. Merceron et al., Citation2016).
Nevertheless, caution must be exercised as the sample size of CH3 is reduced. In addition, our results have some limitations since DMA can vary according to the season, sex and age (Merceron et al. Citation2010; Rivals et al. Citation2015; Berlioz et al. Citation2017, Uzunidis, Citation2020; Jiménez-Manchón et al. Citation2023). In this study, these factors could not be considered because they were not available for most specimens. Future research will need to consider them to improve our understanding of the effect of these factors on DMTA.
Discrimination of wild and domestic goats
Dental texture microwear patterns of three wild goat populations (C. pyrenaica, C. ibex and C. nubiana) have been compared to those of four management strategies of domestic goats. The results obtained by SSFA and ISO parameters indicate that the three wild goat groups had a dental texture that was more complex (Asfc), rougher (height and functional ISO parameters such as Sq and Sk) and with more volume (e.g. Vmp) compared to the four domestic goat groups. In a previous work (Jiménez-Manchón et al. Citation2023), the texture dental wear of these three wild goat populations was compared to that of the domestic goat group fed in an overgrazed environment in the northeastern Iberian Peninsula (CH2). The preliminary results of this previous work indicated a more abraded tooth texture surface in domestic goats than in wild ibexes. In the present paper, the inclusion of three management strategies (i.e. CH1, CH3 and CH4) supports the results obtained by our previous study (Jiménez-Manchón et al. Citation2023). These differences can be explained by the higher consumption of woody plants by wild goats (see sections 2.1.5 to 2.1.7) compared to domestic goats.
A dental microwear texture dominated by pits and scratches of different sizes tends to produce more complex surfaces (Ungar et al., Citation2007). In this sense, Asfc values for wild ibexes are more variable than in domestic goats (), suggesting the consumption of both hard and soft plants. Other hypotheses, including mechanical and physiological factors, can also explain that (Jiménez-Manchón et al. Citation2023). Domestic goats would have taken more small abrasive particles and/or soft food than wild goats. Domestic goats were fed in human-made environments and, therefore, the availability of grasses and the intake of dust/grit were probably higher. However, other factors can contribute to these differences between wild and domestic caprines. Physiological factors, such as the difference in muscle force, may play a role in this (e.g. Harbers et al. Citation2020; Neaux et al. Citation2022). The possible differences between rumination processes between wild and domestic caprines (García-González Citation1987; Cuartas and García-González Citation1992; Clauss et al. Citation2006) could also explain that since it affects DMA (Hatt et al. Citation2019, Citation2020). Dust and grit are washed off by the liquid of rumen and this process varies according to the diet, the sex of animals and between ruminants (Clauss et al. Citation2009).
The higher variability found in wild goats compared to domestic goats could be explained by the fact that the mobility of wild taxa is not controlled by humans. Consequently, the resources available in their diet are more diverse than in domestic goats. Other factors that could contribute to this high variability may include differences in feeding behaviours between males and females in extant ibex populations (e.g. Cuartas et al. Citation1996), as well as seasonal variations. Considering that DMTA reflects the last food intakes before death, seasonality can influence the results (e.g. Merceron et al., Citation2021). In future work, other techniques, such as dental cementum, could be employed to address this issue (Naji et al. Citation2022; Jiménez-Manchón et al. Citation2023). In regard to variability among wild goats, Capra nubiana (CN) exhibits lower values for complexity (Asfc), height (e.g. Sq) and volume (e.g. Vm) when compared to Capra pyrenaica (CP) and Capra ibex (CI). CN is the only studied species that inhabits a dry environment (i.e. desert). This can be attributed to the substantial intake of dust and grit, resulting in a more abrasive dental wear. Moreover, different degrees of humidity within plants, as previously suggested in the case of domestic goats (see 4.1), may provide an additional explanation.
It is worth mentioning the similarities between dental microwear texture between the domestic group with a rougher and more complex dental texture wear, CH4, i.e. herding goats in mixed vegetation in southern France, and the Nubian ibexes (CN). Nubian goats were fed in an arid environment and, therefore, it is the species with the highest grass consumption among the wild populations studied in this work and the availability of resources in their environment is lower than for the other wild goat species (i.e. CP and CI). Even so, the dental microwear pattern was rougher and more complex than those of domestic goats.
Our results show a gradient in dental microwear texture based on the intake of grasses, dust/grit, and woody plants among domestic goats fed on Laghouat steppes (CH1), domestic goats with a mixed dietary regime (CH4), Nubian goats with a significant intake of grass plants and the wild ibex from the southern Iberian Peninsula (CP) and the Alps (CI).
Exploring the use of ISO 25,178 parameters
In DMTA, Scale-Sensitive Fractal Analysis (SSFA) is broadly used for the analysis of ungulate and primate palaeodiet reconstructions (e.g. Krueger, Citation2015; Martínez-Martínez et al., Citation2022). The main variables are: complexity (Asfc), anisotropy (epLsar), heterogeneity of complexity (HAsfc) and textural fill volume (Tfv). These parameters can be easily presented in bivariate or univariate plots because of the low number of variables.
More recently, researchers have extended the use of ISO 25178 parameters. These parameters are used for animal diet reconstruction, but also in lithic use-wear (e.g. Ibáñez et al. Citation2019). These variables are powerful because they can measure orientation of micro-features, anisotropy, height, volume and area at the same time for a given sample using confocal microscopy. Due to the high number of parameters (>30 parameters), statistical procedures to reduce the number of parameters have been proposed in several works (Francisco et al. Citation2018; Ackermans et al. Citation2021), with the aim to ease visualisation of the results. In this work, we explored the visualisation of the ISO 25178 in bivariate plots, as proposed in previous works (Ackermans et al. Citation2020). For this purpose, a post-hoc pairwise analysis was performed. We selected the most discriminant variables of four parameter groups: Vmp (material volume of the core at given material ratio, for volume), Sq (root mean square height, for height), Spc (arithmetic mean peak curvature, for feature) and Spk (peak reduced height, for function). Previous works have already demonstrated the effectiveness of these parameters to discriminate animal diets. Purnell et al. (Citation2013) and Ibáñez et al. (Citation2020) attested that the consumption of woody plants increases the volume (Vmp) of the texture of dental wear. Winkler et al. (Citation2020) observed that Spc can be used to discriminate diets according to the size and shape of the particles consumed. Finally, Sq is a parameter broadly used to distinguish rough and flattened surfaces in DMTA (e.g. Yamada et al. Citation2018; Francisco et al. Citation2018; Louail et al. Citation2022).
The results presented in bivariate plots in are very consistent with those obtained by SSFA () and by discriminant analysis (). The goat group with higher intake of ligneous plants presented high values for volume (Vmp), height (Sq), peaks (Spk) and pointed-shaped peaks (Spc). More specifically, among domestic goats, those fed in the Laghouat steppe presented the lowest values for these four parameters. In contrast, goats fed in mixed areas presented the highest ones. The same trend was observed with wild goats. They presented higher values for these parameters than the domestic ones. Among wild ibexes, C. pyrenaica had the highest values, and C. nubiana the lowest. These bivariate plots, although they could also be presented in univariate plots, could be an easier alternative to visualise the different diets in goats using ISO 25178 parameters.
Reconstructing goat palaeodiet in the neolithic near east
Several zooarchaeological findings support the early goat management in the southern Levant since Early Pre-Pottery Neolithic B (c. 8500–8100 cal. BCE) and Middle Pre-Pottery Neolithic B (c. 8100–7500 cal. BCE) periods (Horwitz et al., Citation1999; Munro et al., Citation2018; Gourichon and Horwitz, Citation2021). However, determining the status of goats (i.e. wild or domestic) in this region is challenging due to difficulties in morphological distinction. To address this issue, palaeodiet reconstruction is a powerful approach, as dietary behavioural changes may have occurred before morphometric differences emerged. In this sense, a previous study demonstrated the potential of DMTA to tackle this matter (Jiménez-Manchón et al. Citation2023). Moreover, this technique allows the exploration of feeding strategies, pasture types (e.g. woodland or open areas), and foddering practices during the Early Neolithic period. In the aforementioned work (Jiménez-Manchón et al. Citation2023), palaeodiet reconstruction was carried out on archaeological goats from the early Neolithic sites of Tell Qarassa and Kharaysin. This investigation utilised a comparative reference collection of wild goats (Capra nubiana, Capra ibex and Capra pyrenaica) alongside a single management strategy in domestic goats (extensive grazing on wooded and overgrazed areas).
In this paper, we analysed the same individuals from these two sites using a broader comparative database (i.e. three wild species and four management strategies). The results obtained in this study show that half of the archaeological goat specimens from Kharaysin are classified in one of the wild goat groups, specifically C. nubiana and C. ibex, while the other half fall into one of the groups of domestic goats. The results are coherent with the previous work (Jiménez-Manchón et al. Citation2023), suggesting the occurrence of animal husbandry control during Middle Pre-Pottery Neolithic B at this site. Additionally, it is important to note the high variability found in Kharaysin, which may again indicate the coexistence of both domestic and wild goats (Jiménez-Manchón et al. Citation2023). The inclusion of a large number of management strategies in this paper allowed us to refine the type of management strategy. Archaeological goats are predominantly classified in the CH4 group, indicating that they fed in mixed vegetation (eudicots and grass plants). These results could indicate that these goats could have grazed in the surrounding of the site, which were dominated by wooded steppe vegetation, with a prevalence of eudicots such as Quercus and Pistacia, as well as grasses (e.g. Asouti et al. Citation2015).
Regarding Tell Qarassa, two individuals were classified as wild ibex (C. nubiana and C. ibex) and one was classified within one of the groups of domestic goats. As for Kharaysin, this specimen was classified in the CH4 groups, indicating a mixed diet consisting of eudicots and grass plants, as found in the surroundings of the site (Arranz-Otaegui et al., Citation2017). Notably, for the goats from Tell Qarassa, high values of heterogeneity (HAsfc9) and low values of anisotropy (epLsar) are observed. These values could indicate a heterogeneous wear pattern across the dental surface, suggesting high variability in diet and predominant consumption of eudicot plants (Scott et al. Citation2012; Merceron et al. Citation2016). However, these interpretations should be taken with caution due to the reduced number of specimens studied.
The application of this new comparative reference collection demonstrates the potential of DMTA to characterise palaeodiet in domestic and wild ungulates in Archaeology, enabling the exploration of broader topic such as feeding strategies employed by herders, as well as more specific issues, such as differences in feeding behaviours between wild and domestic ungulates. In future research, additional feeding strategies, such as types of fodder, will be incorporated to the database.
Conclusion
The palaeodiet reconstruction of domestic ungulates is a valuable source of information on livestock management, as it is possible to assess the food management, the foddering strategies and landscape use in late Prehistory. Dental microwear analysis is a non-destructive powerful approach for palaeodiet reconstruction. Unlike wild ungulates, the number of works dealing with the analysis of palaeodiet in domestic mammals is still scarce. These works have been mainly carried out using low-magnification dental microwear analysis (LMDA) and the discrimination of palaeodiets, i.e. browsers and grazers, of domestic animals with this technique is not well adapted. A previous work showed that the use of texture quantitative analysis (DMTA) allows better discrimination in domestic ungulates than LMDA (Ibáñez et al. Citation2020).
In this paper, we built a reference collection of modern domestic goats with different exploitations and diets using DMTA. More precisely, we analysed goats fed in the Laghouat steppe, on grasslands of the southern side of the Pyrenees, in wooded and overgrazed areas of the northeastern Iberian Peninsula, and in wooded areas of the Causse of Larzac in southern France. This study shows that it is possible to discriminate quantitatively different dietary regimes through DMTA.
The groups of modern domestic goats presented a texture dental wear linked to the type of pasture and diet. Thus, the group of domestic goats with a higher intake of grasses and dust/grit (goats fed in the Laghouat steppe) presented a less complex and very abraded texture dental wear. In contrast, the group with a more diversified diet (goats fed in wooded areas in the Causse of Larzac in southern France) presented a higher roughness and complexity. The two other groups, fed in grasslands and overgrazed areas, presented intermediate values. The comparison between the tooth wear of these domestic goat groups and three wild goat populations shows significant differences. Domestic goats generally present a more abraded and less complex dental wear than that of wild ibexes. Our data support the results obtained in previous work (Jiménez-Manchón et al. Citation2023). These differences could be explained by mechanical factors, i.e. a higher intake of soft food and small abrasive particles by domestic goats, although physiological and behavioural differences between domestic and wild goats cannot be ruled out, especially because the study deals with four different species (one domestic and three wild). Additionally, we explored the reduction of ISO 25178 parameters to easily visualise the results obtained by this type of parameter. Our results indicate that the most discriminant parameters are Vmp, Sq, Spc and Spk. These variables can be used to discriminate among goat palaeodiets using bivariate and/or univariate graphs. This comparative reference collection comprises wild and domestic goats and was used to characterise the palaeodiet of archaeological goats from two Pre-Pottery Neolithic sites in the southern Levant, namely Tell Qarassa and Kharaysin. The results reinforce those obtained in a previous study (Jiménez-Manchón et al. Citation2023), suggesting human feeding control for part of the goats. In this paper, the incorporation of additional management strategies suggests that early managed goats in the southern Levant were potentially fed in a mixed environment dominated by wooded plants.
This paper demonstrates the potential of DMTA to differentiate wild and goat populations and to discriminate various management strategies. However, it is important to note that this database does not encompass all the diversity that exists currently or might have existed in the past. This is particularly evident in the fact that a few of the archaeological specimens studied fall outside the reference models (domestic and wild) used (). Furthermore, caution must be exercised as the modern specimens were collected in different seasons, potentially affecting sample variability. Other factors to consider include sex, much of which was unknown in modern specimen databases. Additionally, sample size has to be considered, which is limited in some modern goat populations and archaeological assemblages. Future research will incorporate bioarchaeological approaches to address these issues, such as cementum analyses, new wild goat species, management strategies, and archaeological data to further enhance our understanding of the impacts of known diets in DMTA for palaeodiet reconstruction in Archaeology and Palaeontology.
Supplemental Material
Download Zip (69.5 KB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2024.2359466.
Additional information
Funding
References
- Ackermans NL, Winkler DE, Martin LF, Kaiser TM, Clauss M, Hatt J-M. 2020. Dust and grit matter: abrasives of different size lead to opposing dental microwear textures in experimentally fed sheep (Ovis aries). J Exp Biol. 223:jeb220442. doi: 10.1242/jeb.220442.
- Ackermans NL, Winkler DE, Schulz-Kornas E, Kaiser TM, Martin LF, Hatt J-H, Clauss M. 2021. Dental wear proxy correlation in a long-term feeding experiment on sheep (Ovis aries). J R Soc Interface. 18(180):20210139. doi: 10.1098/rsif.2021.0139.
- Albarella U, Tagliacozzo A, Dobney K, Rowley-Conwy P. 2006. Pig hunting and husbandry in prehistoric italy: a contribution to the domestication debate. Proc Prehist Soc. 72:193–227. doi: 10.1017/S0079497X00000827.
- Albizuri S, Nieto-Espinet A, Valenzuela-Lamas S. 2010. Canvis en l’alimentació càrnia a Catalunya entre els segles XII i III aC. Sagvntvm Extra. 9:161–188.
- Arranz-Oategui A, López-S’áez JA, Araus JL, Portillo M, Balbo A, Iriarte E, Gourichon L, Braemer F, Zapata L, Ibáñez JJ. 2017. Landscape transformations at the dawn of agriculture in southern Syria (10.7-9.9 ka cal. BP): plant-specific responses to the impact of human activities and climate change. Quat Sci Rev. 158:145–163.
- Asouti E, Kabukcu C, White CE, Kuijt I, Finlayson B, Makarewicz C. 2015. Early Holocene woodland vegetation and human impacts in the arid zone of the southern Levant. Holocene. 25(10):1565–1580. doi: 10.1177/0959683615580199.
- Balasse M. 2002. Reconstructing dietary and environmental history from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Intl J Osteoarchaeol. 12(3):155–165. doi: 10.1002/oa.601.
- Balasse M, Obein G, Ughetto-Monfrin J, Mainland I. 2012. Investigating seasonality and season of birth in past herds: a reference set of sheep enamel stable oxygen isotope ratios. Archaeometry. 54(2):349–368. doi: 10.1111/j.1475-4754.2011.00624.x.
- Belala F, Hirche A, Duller SD, Tourki M, Salamani M, Grandi M, Ait Hamouda T, Boughani M. 2018. Rainfall patterns of algerian steppes and the impacts on natural vegetation in the 20th century. J Arid Land. 10(4):561–573. doi: 10.1007/s40333-018-0095-x.
- Berlioz E, Azorit C, Blondel C, Tellado Ruiz MS, Merceron G. 2017. Deer in an arid habitat: dental microwear textures track feeding adaptability. Hystrix. 28:222–230.
- Berlioz E, Leduc C, Hofman-Kaminska E, Bignon-Lau O, Kowalcyk R, Merceron G. 2022. Dental microwear foraging ecology of a large browsing ruminant in Northern Hemisphere: The European moose (Alces alces). Palaeogeogr Palaeoclimatol Palaeoecol. 586:110754. doi: 10.1016/j.palaeo.2021.110754.
- Calandra I, Labonne G, Schulz-Kornas E, Kaiser TM, Montuire S. 2016. Tooth wear as a means to quantify intra-specific variations in diet and chewing movements. Sci Rep. 6(1):34037. doi: 10.1038/srep34037.
- Calandra I, Schulz E, Pinnow M, Krohn S, Kaiser TM. 2012. Teasing apart the contributions of hard dietary items on 3D dental microtextures in primates. J Hum Evol. 63(1):85–98. doi: 10.1016/j.jhevol.2012.05.001.
- Clauss M, Hofmann RR, Fickel J, Streich WH, Hummel J. 2009. The intraruminal papillation gradient in wild ruminants of different feeding types: implications for rumen physiology. J Morphol. 270(8):929–942. doi: 10.1002/jmor.10729.
- Clauss M, Hummel J, Streich WJ. 2006. The dissociation of the fluid and particle phase in the forestomach as a physiological characteristic of large grazing ruminants: an evaluation of available, comparable ruminant passage data. Eur J Wildl Res. 52(2):88–98. doi: 10.1007/s10344-005-0024-0.
- Couturier MAJ. 1962. Le bouquetin des Alpes Capra aegagrus ibex. Grenoble: Couturier.
- Cuartas P, García-González R. 1992. Quercus ilex browse utilization by Caprini in Sierra de Cazorla and Segura (Spain). Vegetatio. 99-100(1):317–330. doi: 10.1007/BF00118239.
- Cuartas P, García-González R, Alados CL, Escós J. 1996. Selección de la dieta ycomportamiento alimentario. In: Alados CL, Quilez JE, editors. Ecología y comportamiento de la cabra mont´es. Consideraciones para su gesti´on. Monografíasdel Museo Nacional de Ciencias Naturales. Madrid. p. 151–198.
- Dalila N, Slimane B. 2008. La désertification dans les steppes algériennes : causes, impacts et actions de lutte. VertigO. 8(1). doi: 10.4000/vertigo.5375.
- Davis SJM. 1987. The archaeology of Animals. London: Routledge.
- De Santis LRG, Scott JR, Schubert BW, Donohue SL, Mccray BM, Van Stolk CA, Winburn AA, Greshko MA, O’Hara MC, Ungar P. 2013. Direct Comparisons of 2D and 3D Dental Microwear Proxies in Extant Herbivorous and Carnivorous Mammals. PLoS One. 8(8):e71428. doi: 10.1371/journal.pone.0071428.
- Francisco A, Blondel C, Brunetière N, Ramdarshan A, Merceron G. 2018. Enamel surface topography analysis for diet discrimination. A methodology to enhance and select discriminative parameters. Surf Topography: Metrol Prop. 6(1):015002. doi: 10.1088/2051-672X/aa9dd3.
- Gallego A, Colominas L, Burguet-Coca A, Aguilera M, Parlet J-M, Tornero C. 2020. What is on the menu today? Creating a microwear reference collection through a controlled-food trial to study feeding management systems of ancient agropastoral societies. Quaternary International. 557:3–11. doi: 10.1016/j.quaint.2020.02.020.
- Gallego A, Rivals F, Colominas L, Palet J-M. 2017. Pastando en las marismas. Una aproximación desde la técnica del desgaste dentario a la alimentación del ganado ovino en el Empordà romano (noreste de la Península Ibérica). Pyrenae. 48:93–113.
- García-González R. 1987. Estudio del crecimiento postnatal en corderos de raza Rasa Aragonesa. Monografías del Instituto de Estudios Altoaragoneses 13. Huesca: CSIC, Diputación Provincial de Huesca.
- Gordon KD. 1984. Hominoid dental microwear: complications in the use of microwear analysis to detect diet. J Dent Res. 63(8):195–215. doi: 10.1177/00220345840630080601.
- Gourichon L, Kolska-Horwitz L. 2021. An inter-regional comparison of animal domestication in the northern and southern levant. In: Vieugué J, Mazzucco N, editors. Dietary Practices of the First Mediterranean Farmers Producing, Storing, Preparing and Consuming Foodstuffs in the Neolithic Period. Turnhout: Brepols Publishers; p. 33–68.
- Grayson DK. 1984. Quantitative Zooarchaeology. New York: Academic Press.
- Green DR, Green GM, Colman AS, Bidlack FB, Tafforeau P, Smith TM. 2017. Synchrotron imaging and Markov Chain Monte Carlos reveal tooth mineralization patterns. PLOS ONE. 12(10):e0186391. doi: 10.1371/journal.pone.0186391.
- Grine FE. 1986. Dental evidence for dietary differences in australopithecus and paranthropus: a quantitative analysis of permanent molar microwear. J Hum Evol. 15(8):783–822. doi: 10.1016/S0047-2484(86)80010-0.
- Grupe G. 1995. Preservation of collagen in bone from dry, sandy soil. J Archaeolog Sci. 22(2):193–199. doi: 10.1006/jasc.1995.0021.
- Hackham E, Ritte U. 1991. Foraging pressure of the Nubian ibex Capra ibex nubiana and its effect on the indigenous vegetation of the En Gedi Nature Reserve, Israel. Biol Conserv. 63(1):9–21. doi: 10.1016/0006-3207(93)90068-C.
- Hadjikoumis A, Vigne JD, Simmons A, Guilaine J, Fiorillo D, Balasse M. 2019. Autumn/winter births in traditional and Pre-Pottery Neolithic caprine husbandry in Cyprus: evidence from ethnography and stable isotopes. J Anthropol Archaeol. 53:102–111. doi: 10.1016/j.jaa.2018.12.001.
- Halstead PLJ. 1981. Counting sheep in neolithic and bronze age Greece. In: Hodder I, Isaac G, Hammond N, editors. Pattern of the past: studies in honour of David Clarke. Cambridge: Cambridge University; p. 307–339.
- Halstead PLJ. 2006. Sheep in the Garden: the Integration of Crop and Livestock Husbandry in Early Farming Regimes of Greece and Southern Europe. In: Serjeantson D, Field D, editors. Animals in the Neolithic of Britain and Europe. Oxford: Oxbow Books; p. 42–55.
- Harbers H, Zanolli C, Cazenave M, Theil J-C, Ortiz K, Blanc B, Locatelli Y, Schafberg R, Lecompte F, Baly I, et al. 2020. Investigating the impact of captivity and domestication on limb bone cortical morphology: an experimental approach using a wild boar model. Sci Rep. 10(1):19070. doi: 10.1038/s41598-020-75496-6.
- Hatt J-M, Codron D, Ackermans NL, Martin LF, Richter H, Kircher PR, Gerspach C, Hummel J, Clauss M. 2020. Differences in the rumen washing me- chanism in sheep fed diets with abrasives of various concentrations and sizes. Palaeogeogr Palaeoclimatol Palaeoecol. 550:109728. doi: 10.1016/j.palaeo.2020.109728.
- Hatt J-M, Codron D, Müller DWH, Ackermans NL, Martin LF, Kircher PR, Hummel J, Clauss M. 2019. The rumen washes off abrasives before heavy-duty chewing in ruminants. Mamm Biol. 97:104–111. doi: 10.1016/j.mambio.2019.06.001.
- Helmer D, Gourichon L, Sidi Maamar H, Vigne J-D. 2005. L’élevage des caprinés néolithiques dans le sud-est de la France : saisonnalité des abattages, relations entre grottes‐bergeries et sites de plein air. Anthropozool. 40:167‐189.
- Helmer D, Gourichon L, Vila E. 2007. The development of the exploitation of products from Capra and Ovis (meat, milk and fleece) from the PPNB to the Early Bronze in the northern Near East (8700 to 2000 BC cal.). Anthropozool. 42:41–69.
- Helmer D, Vigne J-D. 2004. La gestion des caprinés domestiques dans le midi de la France. In: Bodu P, Constantin C, editors. Approches fonctionnelles en Préhistoires. Actes de XXVème colloque du Congrès Préhistorique de France. Nanterre: Société Préhistorique Française; p. 297–407.
- Horwitz LK, Tchernov E, Ducos P, Becker C, von den Driesch A, Martin L, Garrard A. 1999. Animal domestication in the southern Levant. Paleorient. 25(2):63–80.
- Ibáñez JJ, Anderson PC, Arranz-Otaegui A, González-Urquijo JE, Jörgensen-Lindahl A, Mazzuco N, Pichon F, Richet T. 2021. Sickle gloss texture analysis elucidates long-term change in plant harvesting during the transition of agriculture. J Archaeolog Sci. 136:105502. doi: 10.1016/j.jas.2021.105502.
- Ibáñez JJ, Balbo A, Braemer F, Gourichon F, Gourichon L, Iriarte E, Santana J, Zapata L. 2010. The early PPNB levels of Tell Qarassa North (Sweida, southern Syria). Antiquity. 84:325.
- Ibáñez JJ, Jiménez-Manchón S, Blaise É, Nieto-Espinet A, Valenzuela-Lamas S. 2020. Discriminating management strategies in modern and archaeological domestic caprines using low-magnification and confocal dental microwear analyses. Quat Int. 557:23–38. doi: 10.1016/j.quaint.2020.03.006.
- Ibáñez JJ, Lazuen T, González-Urquijo J. 2019. Identifying experimental tool use through confocal microscopy. J Archaeol Method Theory. 26(3):1176–1215. doi: 10.1007/s10816-018-9408-9.
- Ibáñez JJ, Muñiz J, Iriarte E, Monik M, Santana J, Teira L, Corrada M, Lagüera MÁ, Lendakova Z, Regalado E, et al. 2015. Kharaysin: a PPNA and PPNB site by the Zarqa river 2014 and 2015 field seasons. Neo-Lithics. 2(15):11–19.
- Jiménez-Manchón S. 2020. Pratiques d’élevage entre l’Empordà et le Languedoc à l’âge du Fer. Archéozoologie, alimentation animale et saisonnalité [ PhD thesis 3]. de Montpellier, Montpellier, France: Université Paul-Valéry.
- Jiménez-Manchón S, Gourichon L, Muñiz J, Ibáñez JJ. 2023. Discriminating dietary behaviour between wild and domestic goats using dental microwear texture: first results from a modern reference set and early Neolithic goat exploitation in the southern Levant. J Archaeolog Sci. 155:105779. doi: 10.1016/j.jas.2023.105779.
- Jiménez-Manchón S, Rivals F, Gourichon L, De Prado G, Codina F, Castanyer P, Tremoleda J, Santos M, Gardeisen A. 2023. A combined approach to reconstruct livestock management in Iron Age north-eastern Iberia: estimating the season of death and palaeodiet using cementochronology and dental micro- and mesowear analyses. Archaeofauna. 31(1):161–177. doi: 10.15366/archaeofauna2023.32.1.010.
- Jiménez-Manchón S, Valenzuela-Lamas S, Cáceres I, Orengo H, Gardeisen A, López D, Rivals F. 2019. Reconstruction of caprine management and landscape use through dental microwear analysis: the case of the Iron age site of el Turó de la Font de la Canya (Barcelona, Spain). Environ Archaeol. 24(3):303–316. doi: 10.1080/14614103.2018.1486274.
- Krueger KL. 2015. Reconstructing diet and behavior in bioarchaeological groups using incisor microwear texture analysis. J Archaeolog Sci: Rep. 1:29–37.
- Kubo MO, Fujita M. 2021. Diets of Pleistocene insular dwarf deer revealed by dental microwear texture analysis. Palaeogeogr Palaeoclimatol Palaeoecol. 562:110098. doi: 10.1016/j.palaeo.2020.110098.
- Lespes C. 2020. Pratiques alimentaires et agropastorales à la fin de l’Âge du Bronze et aux débuts de l’Âge du Fer en Languedoc : du littoral aux premiers contreforts méridionaux du Massif Central [ PhD thesis]. de Montpellier, Montpellier, France: Université Paul-Valéry 3.
- López-I-Gelats F, Rivera-Ferre MG, Madruga-Andreu C, Bartolomé-Filella J. 2015. Is multifunctionality the future of mountain pastoralism? Lessons from the management of semi-natural grasslands in the Pyrenees. Span J Agric Res. 13(4):e0307. doi: 10.5424/sjar/2015134-6960.
- Louail M, Caner L, Neaux D, Ortiz K, Locatelli Y, Cucchi T. 2022. Identifying the impact of soil ingestion on dental microwear textures using a wild boar experimental model. J Archaeol Method Theory. 30(3):855–875. doi: 10.1007/s10816-022-09574-6.
- Louail M, Ferchaud S, Souron A, Walker AEC, Merceron G. 2021. Dental microwear texture differ in pigs with overall similar diets but fet with different seeds. Palaeogeogr Palaeoclimatol Palaeoecol. 573:110515. doi: 10.1016/j.palaeo.2021.110415.
- Mainland IL, Towers J, Ewens V, Davis G, Montgomery J, Batey C, Card N, Downes J. 2016. Toiling with teeth: an integrated dental analysis of sheep and cattle dentition in iron age and viking late norse orkney. J Archaeolog Sci: Rep. 6:837–855. doi: 10.1016/j.jasrep.2015.12.002.
- Makarewicz CA, Arbuckle BS, Öztan A. 2017. Vertical transhumance of sheep and goats identified by intra-tooth sequential carbon (δ13C) and oxygen (δ18O) isotopic analyses: evidence from Chalcolithic Köşk Höyük, central Turkey. J Archaeolog Sci. 86:68–80. doi: 10.1016/j.jas.2017.01.003.
- Makarewicz C, Horwitz LK, Goring-Morris AN. 2016. Local adoption of animal husbandry in the southern Levant: an isotopic perspective from the Pre-Pottery Neolithic B funerary site of Kfar HaHoresh. Environ Archaeol. 21(3):199–213. doi: 10.1179/1749631415Y.0000000024.
- Manallah K. 2012. Techniques de mesure hyperfréquences large bande appliquées aux matériaux hétérogènes solides [ master thesis]. Algeria: Université Ferhat Abbas.
- Martínez T. 2000. Diet selection by Spanish ibex in early summer in Sierra Nevada. Acta Theriol (Warsz). 45(3):335–346. doi: 10.4098/AT.arch.00-33.
- Martínez-Martínez LM, Estebaranz-Sánchez F, Romero A, Ibáñez JJ, Hidalgo-Trujillo L, Avià Y, Pérez-Pérez A. 2022. Effectiveness of buccal dental-microwear texture in African Cercopithecoidea dietary discrimination. Am J Biol Anthropol. 179(4):678–686.
- Martinez T, Martínez E, Fandos P. 1985. Composition of the food of the Spanish Wild Goat in Sierras de Cazorla and Segura, Spain. Acta Theriol (Warsz). 30(29):461–494. doi: 10.4098/AT.arch.85-31.
- Merceron G, Berlioz ´E, Vonhof H, Green D, Garel M, Tütken T. 2021. Tooth tales told by dental diet proxies: an alpine community of sympatric ruminants as a model to decipher the ecology of fossil fauna. Palaeogeogr Palaeoclimatol Palaeoecol. 562:110077. doi: 10.1016/j.palaeo.2020.110077.
- Merceron G, Escarguel G, Angibault J-M, Verheyden-Tixier H, Begun D. 2010. Can dental microwear textures record inter-individual dietary variations? PLoS One. 5(3):1–9. doi: 10.1371/journal.pone.0009542.
- Merceron G, Ramdarshan A, Blondel C, Boisserie J-R, Brunetiere N, Francisco A, Gautier D, Milhet X, Novello A, Pret D. 2016. Untangling the environmental from the dietary: dust does not matter. Proc R Soc B. 283(1838):20161032. doi: 10.1098/rspb.2016.1032.
- Mihlbachler MC, Beatty BL, Caldera-Siu A, Chan D, Lee R. 2012. Error Rates and Observer Bias in Dental Microwear Analysis Using Light Microscopy. Palaeontol Electronica. 15:12A. doi: 10.26879/298.
- Miller H, Baird D, Pearson J, Lamb AL, Grove M, Martin M, Garrard A. 2019. The origins of nomadic pastoralism in the eastern Jordanian steppe: a combined stable isotope and chipped stone assessment. Levant. 50(3):281–304. doi: 10.1080/00758914.2019.1651560.
- Munro N, Bar-Oz G, Meier J, Sapir-Hen L, Stiner MC, Yeshurun R. 2018. The emergence of animal management in the southern levant. Nat Sci Rep. 8:9279.
- Naji S, Rendu W, Gourichon L. 2022. Dental cementum in Anthropology. Cambridge University Press.
- Navarrete V, Tornero C, Balasse M, Saña M. 2019. Food management of early introduced caprine and bovine herds in the early Neolithic site of La Draga (Banyoles): An isotopic approach. Int J Osteoarchaeol. 29(6):986–998. doi: 10.1002/oa.2812.
- Neaux D, Harbers H, Blanc, Barbara B, Ortiz K, Locatelli Y, Herrel A, Debat V, Cucchi T. 2022. The effect of captivity on craniomandibular and calcaneal ontogenetic trajectories in wild boar. J Exp Zool B Mol Dev Evol. 338(8):575–585. doi: 10.1002/jez.b.23130.
- Nieto-Espinet A, Huet T, Trentacoste A, Guimarães S, Orengo H, Valenzuela-Lamas S, Zerboni A. 2021. Resilience and livestock adaptations to demographic growth and technological change: A diachronic perspective from the Late Bronze Age to Late Antiquity in NE Iberia. PLOS ONE. 16(2):e0246201. doi: 10.1371/journal.pone.0246201.
- Nieto-Espinet A, Valenzuela-Lamas S, Bosch D, Gardeisen A. 2020. Livestock production, politics and trace: A glimpse from Iron Age and Roman Languedoc. J Archaeolog Sci: Rep. 30:102077. doi: 10.1016/j.jasrep.2019.102077.
- Palmer C, Gilbertson D, el-Rishi H, Hunt C, Grattan J, McLaren S, Pyatt B, Mattingly. 2007. The Wady Faynan today: landscape, environment, people. In: Barker G, Gilbertson D, editors. Archaeology and Desertification: the Wady Faynan Landscape Survey, southern Jordan. Oxford: Oxbow Books; p. 25–57.
- Parrini F, Cain JW, Krausman PR. 2009. Capra ibex (Artiodactyla: Bovidae). Mamm Species. 830:1–12. doi: 10.1644/830.1.
- Pini R, Ravazzi C, Raiteri L, Guerreschi A, Castellano L, Comolli R, Gill J. 2017. From pristine forests to high-altitude pastures: an ecological approach to prehistoric human impact on vegetation and landscapes in the western Italian Alps. J Ecol. 105(6):1580–1597. doi: 10.1111/1365-2745.12767.
- Prioton J. 1970. Flore forestière du Larzac. Causses et Cévennes. 11(1):415–422.
- Purnell MA, Crumpton N, Gill PG, Jones G, Rayfield EJ. 2013. Within-guild dietary discrimination from 3-D textural analysis of tooth microwear in insectivorous mammals. J Zool. 291(4):249–257. doi: 10.1111/jzo.12068.
- Purnell MA, Seehausen O, Galis F. 2012. Quantitative three-dimensional microtextural analyses of tooth wear as a tool for dietary discrimination in fishes. J R Soc Interface. 9(74):2225–2233. doi: 10.1098/rsif.2012.0140.
- Ramdarshan A, Blondel C, Brunetière N, Francisco A, Gautier D, Surault J, Merceron G. 2016. Seeds, browse, and tooth wear: a sheep perspective. Ecol Evol. 6(16):5559–5569. doi: 10.1002/ece3.2241.
- Ramdarshan A, Blondel C, Gautier D, Surault J, Merceron G. 2017. Overcoming sampling issues in dental tribology: insights from an experimentation on sheep. Palaeontol Electr. 30(3):1–19. doi: 10.26879/762.
- Ridouh MD. 2021. Étude comparative des caractéristiques morphométriques et génitales chez la chèvre [ master thesis]. Algeria: Université l’arbi ben mhidi Oum El Bouaghi.
- Rieau C. 2014. Étude de la gestion du bétail à l’aide de la méthode des microusures dentaires dans le Midi méditerranéen entre l’âge du Bronze et la période galloromaine [ PhD thesis]. de Montpellier, Montpellier, France: Université Paul-Valéry 3.
- Rivals F, Deniaux B. 2005. Investigation of human hunting seasonality through dental microwear analysis of two Caprinae in late Pleistocene localities in Southern France. J Archaeolog Sci. 32(11):1603–1612. doi: 10.1016/j.jas.2005.04.014.
- Rivals F, Gardeisen A, Cantuel J. 2011. Domestic and Wild Ungulates Dietary Traits at Kouphovouno (Sparte, Greece): Implications for Livestock Management and Paleoenvironment in the Neolithic. J Archaeolog Sci. 38(3):528–537. doi: 10.1016/j.jas.2010.10.007.
- Rivals F, Prignano L, Semprebon G, Lozano S. 2015. A tool for determing duration of mortality events in archaeological assemblages using extant ungulate microwear. Sci Rep. 5(1):17330. doi: 10.1038/srep17330.
- Saña M. 1998. Arqueozoologia i faunes neolítiques a Catalunya. Problemàtica platejada entorn a la dinàmica del procés de domesticació animal. Cypsela. 12:99–110.
- Sánchez-Hernández C, Rivals F, Blasco R, Rosell J. 2015. Short, but repeated Neanderthal visits to Teixoneres Cave (MIS 3, Barcelona, Spain): a combined analysis of tooth microwear patternsand seasonality. J Archaeolog Sci. 49:317–325. doi: 10.1016/j.jas.2014.06.002.
- Schulz E, Calandra I, Kaiser TM. 2010. Applying tribology to teeth of hoofed mammals. Scanning. 32(4):162–182. doi: 10.1002/sca.20181.
- Scott R, Teaford M, Ungar PS. 2012. Dental microwear texture and anthropoid diets. Am J Biol Anthropol. 147(4):551–579. doi: 10.1002/ajpa.22007.
- Scott RS, Ungar PS, Bergstrom TS, Brown CA, Childs BE, Teaford MF, Walker A. 2006. Dental microwear texture analysis: technical considerations. J Hum Evol. 51(4):339–349. doi: 10.1016/j.jhevol.2006.04.006.
- Scott RS, Ungar PS, Bergstrom TS, Brown CA, Grine FE, Teaford MF, Walker A. 2005. Dental microwear texture analysis reflects diets of living primates and fossil hominins. Nature. 436(7051):693–695. doi: 10.1038/nature03822.
- Semprebon G, Godfrey L, Solounias N, Sutherland MR, Jungers WL. 2004. Can low-magnification stereomicroscopy reveal diet? J Hum Evol. 47(3):115–144. doi: 10.1016/j.jhevol.2004.06.004.
- Semprebon G, Rivals F. 2007. Was grass more prevalent in the pronghorn past? An assessment of the dietary adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeogr Palaeoclimatol Palaeoecol. 253(3–4):332–347. doi: 10.1016/j.palaeo.2007.06.006.
- Sierra A, Balasse M, Rivals F, Fiorillo D, Utrilla P, Saña M. 2021. Sheep husbandry in the early Neolithic of the Pyrenees: New data on feeding and reproduction in the cave of Chaves. J Archaeolog Sci: Rep. 37:102935. doi: 10.1016/j.jasrep.2021.102935.
- Solounias N, Semprebon G. 2002. Advances in the reconstruction of ungulate ecomorphology with application to early fossil equids. Am Mus Novit. 3366:1–49.
- Stein GJ. 1987. Regional Economic Integration in Early State Societies. Third Millenium B.C. Patoral Production at Gritille, Southeast Turkey. paleo. 13(2):101–111. doi: 10.3406/paleo.1987.4432.
- Teaford MF, Walker A. 1984. Quantitative Differences in Dental Microwear Between Primate Species with Different Diets and a Comment on the Presumed diet of Sivapithecus. Am J Phys Anthropol. 64(2):191–200. doi: 10.1002/ajpa.1330640213.
- Tornero C, Aguilera M, Ferrio JP, Arcusa H, Moreno-García M, Garcia-Reig S, Rojo-Guerra M. 2018. Vertical sheep mobility along the altitudinal gradient through stable isotope analyses in tooth molar bioapatite, meteoric water and pastures: A reference from the Ebro valley to the Central Pyrenees. Quaternary International. 484(10):94–106. doi: 10.1016/j.quaint.2016.11.042.
- Trentacoste A, Lightfood E, Le Roux P, Buckley M, Kansa SW, Esposito C, Gleba M. 2020. Heading for the hills? A multi-isotope study of sheep management in first-millenniumm BC Italy. J Archaeolog Sci: Rep. 102036.
- Trentacoste A, Nieto-Espinet A, Guimarães S, Wilkens B, Petrucci G, Valenzuela-Lamas S. 2021. New trajectories or accelerating change? Zooarchaeological evidence for Roman transformation of animal husbandry in Northern Italy. Archaeol Anthropol Sci. 13(1):1–22. doi: 10.1007/s12520-020-01251-7.
- Trentacoste A, Nieto-Espinet A, Valenzuela-Lamas S, Biehl PF. 2018. Pre-Roman improvements to agricultural production: Evidence from livestock husbandry in late prehistoric Italy. PLOS ONE. 13(12):e0208109. doi: 10.1371/journal.pone.0208109.
- Ungar PS. 2007. Dental Functional Morphology: The Known, the Unknown and the Unknowable. In: Ungar PS, editor. Early Hominin Diets: The Known, the Unknown and the Unknowable. Oxford: Oxford University Press; p. 39–55.
- Ungar P, Scott JR, McNulty KP, Harcourt-Smith WEH, Lehman T, Dunsworth HM. 2012. Environments of early Miocene Rusinga Island and Songhor: evidence from the dental microwear of tragulids. Am J Phys Anthropol. 147:289–290.
- Uzunidis A. 2020. Dental wear analyses of Middle Pleistocene site of Lunel-Viel (Hérault, France): did Equus and Bos live in a wetland? Quat Int. 557:39–46.
- Uzunidis A, Pineda A, Jiménez-Manchón S, Xafis A, Ollivier V, Rivals F. 2021. The impact of sediment abrasion on tooth microwear analysis: an experimental study. Archaeol Anthropol Sci. 13(8):134. doi: 10.1007/s12520-021-01382-5.
- Valenzuela-Suau L, Rivals F, Ramis D, Valenzuela-Lamas S. 2022. Caprine dental microwear reveals livestock management and exploitation of landscape during the Middle and Late Bronze Age of the Balearic Islands (ca. 1500–850 cal. BC). Archaeol Anthropol Sci. 14(2):37. doi: 10.1007/s12520-021-01487-x.
- Walker A, Hoeck HN, Perez L. 1978. Microwear of mammalian teeth as an indicator of diet. Science. 201(4359):908–910. doi: 10.1126/science.684415.
- Weber K, Winkler D, Schulz-Kornas E, Kaiser TM, Tütken T. 2022. Post-mortem enamel surface texture alteration during taphonomic processes-do experimental approaches reflect natural phenomena?. PeerJ. 10:e12635.
- Winkler DE, Clauss M, Rölle M, Schulz-Kornas E, Codron D, Kaiser TM, Tütken T. 2021. Dental microwear texture gradients in guinea pigs reveal that material properties of the diet affect chewing behaviour. J Exp Biol. 224(13):jeb242446. doi: 10.1242/jeb.242446.
- Winkler DE, Tütken T, Schulz-Kornas E, Kaiser TM, Müller J, Leichliter J, Weber K, Hatt J-M, Clauss M. 2020. Shape, size, and quantity of ingested external abrasives influence dental microwear texture formation in guinea pigs. Proc Natl Acad Sci USA. 117(36):22264–22273. doi: 10.1073/pnas.2008149117.
- Yamada E, Kubo MO, Kohno N. 2018. Three-dimensional tooth surface texture analysis on stall-fed and wild boars (Sus scrofa). PLOS ONE. 10.1371/journal.pone.0204719.
- Zingg A. 2009. Seasonal variability in the diet composition of Alpine ibex (Capra ibex ibex L.) in the Swiss National Park [ master thesis]. Switzerland: University of Zurich.