Abstract
Improvement of agricultural water use efficiency is of major concern with drought problems being one of the most important factors limiting grain production worldwide. Effective management of water for crop production in water-scarce areas requires efficient approaches. Increasing crop water use efficiency and drought tolerance by genetic improvement and physiological regulation may be a means to achieve efficient and effective use of water. A limited water supply inhibits the photosynthesis of plants, causes changes of chlorophyll contents and components and damage to photosynthetic apparatus. It also inhibits photochemical activities and decreases the activities of enzymes in plants. Water stress is one of the important factors inhibiting the growth and photosynthetic abilities of plants through disturbing the balance between the production of reactive oxygen species and the antioxidant defence, causing accumulation of reactive oxygen species which induce oxidative stress to proteins, membrane lipids and other cellular components. A number of approaches are being used to enhance water use efficiency and to minimize the detrimental effect of water stress in crop plants. Proper plant nutrition is a good strategy to enhance water use efficiency and productivity in crop plants. Plant nutrients play a very important role in enhancing water use efficiency under limited water supply. In this paper we discuss the possible effective techniques to improve water use efficiency and some macronutrients (nitrogen, phosphorus, potassium, calcium and magnesium), micronutrients (zinc, boron, iron, manganese, molybdenum and chloride), and silicon (a beneficial nutrient) in detail to show how these nutrients play their role in enhancing water use efficiency in crop plant.
Introduction
Water use efficiency (WUE) can be described on various scales from the leaf to the field (Sinclair et al., Citation1984). In its simplest terms, WUE refers to the ratio of yield to the water used during crop growth (Waraich et al., Citation2008) whereas the ratio of yield to transpiration is termed as transpiration efficiency (TE). Water use efficiency provides a simple way of assessing whether yield is limited by water supply or other factors. A good understanding of crop management effects on WUE may provide researchers with opportunities to identify and select appropriate crop management practices for improved water use efficiency.
The world's population (6000 million) of today is expected to reach about 8100 million by 2030, an increase of 35%. The growing population will result in considerable additional demand for food. Simultaneously, water demand from non-agricultural sectors will keep growing in both developed and developing countries. About 40% of the land in the world is under arid and semi-arid climatic conditions (Gamo, Citation1999). An FAO analysis (FAO, Citation2003) of 93 developing countries expects an increase of agricultural production over the period 1998–2030 by 49% in rain fed and by 81% in irrigated regions. Therefore, much of the additional food production is expected to come from irrigated land, three-quarters of which is located in developing countries. The irrigated area, in developing countries, in 1998 has nearly doubled since 1962. The FAO estimates that the irrigated area in the selected 93 developing countries will only grow by 23% over the 1998–2030 period. However, the effective harvested irrigated area (considering the increase in cropping intensity) is expected to increase by 34%. This means that more area needs to be brought under irrigation to produce more crops with the present available water resources (Ali & Talukder, Citation2008). In the dry areas, water, not land, is the most limiting resource for improving agricultural production. Maximization of yield per unit of water (water use efficiency), and not yield per unit of land (land productivity), is therefore a better strategy for dry farming systems. Under such conditions, more efficient water management techniques must be adopted (Ali & Talukder, Citation2008).Water deficiency and low availability of nutrients often limit crop growth and production potential in agro-ecosystems because most crops are sensitive to water and nutrient deficits during different critical stages. On the other hand, excessive use of water can increase production costs and pollute the environment (enhancing fertilizer leaching). So improvement of water use efficiency (also termed water productivity) in agriculture is very important. It is widely believed that an increase in agricultural water use efficiency is the key approach to mitigate water shortages and to reduce environmental problems.
This paper therefore discusses the role of mineral nutrition and possible mechanisms to improve water use efficiency.
Crop and soil management approaches to enhance water use efficiency
Enhancing agricultural WUE by crop and soil management approaches refers to a farming practice that is able to take full advantage of the natural rainfall and irrigation facilities. The core problem that water-saving agricultural research has to solve is how to raise the water utilization rate and water use efficiency, that is to achieve a high yield on irrigated farmland with the minimum input of water and in rainfed agriculture to maximize rainfall use efficiency. What we stress here is that water-saving agriculture is not simply water-saving irrigation. It is a comprehensive exercise using every possible water-saving measure in whole-farm production, including the full use of natural precipitation as well as the efficient management of an irrigation network and plant mineral nutrition. The different management approaches by which we can improve WUE are given below.
Water-saving irrigation
Due to increased water scarcity, the irrigated area is unlikely to expand in the dry-land region. Supplemental irrigation, the combination of dry-land farming and limited irrigation, is an ideal choice for improving crop yields (Bai & Dong, Citation2001). Good irrigation scheduling requires the timing of irrigation and the amount of water applied to match actual field conditions. This requires information on soil-moisture conditions at the time of irrigation and, when using irrigation, close cooperation among farmers to be effective. Further, water-saving irrigation depends on adopting water-saving techniques in the transportation and application of the water. On-farm WUE can be improved by moving to a more efficient irrigation system. There are three main types of irrigation systems available: border or furrow flood irrigation, sprinkler irrigation and drip irrigation, with drip irrigation being more efficient than sprinkler irrigation and sprinkler irrigation being more efficient than border/furrow irrigation. An investigation into the irrigation of maize showed that 210 mm of irrigation applied by border irrigation under a mulch of wheat straw gave a grain yield, total evapotranspiration and WUE of 8000 kg ha−1, 390 mm and 2.2 kg m−3, respectively (Zhang et al., Citation2002). Increasing the amount of irrigation to 270 mm increased the yield to 8834 kg ha−1, but WUE decreased. Straw mulching reduced the total evaporation by 50 mm showing that wheat straw mulching can reduce the amount of irrigation required by maize in the North China Plain (Zhang et al., Citation2002). Irrigation of cotton in Xinjiang indicated that the flowering and budding stages were the most suitable times to supply limited irrigation water, thus significantly improving the WUE by 57% (Hu et al., Citation2002). In the northern part of China, Liu et al. (Citation2003) reported that the yield and WUE of winter wheat under sprinkler irrigation conditions was increased by 28 and 48% and 636 m3 ha−1 water was saved compared with that under border irrigation conditions. Zhang & Cai, (Citation2001) conducted a cotton irrigation experiment that demonstrated that irrigation under the surface of a plastic mulch is an effective way to protect against soil evaporation and that a mild water deficit during the budding stage could significantly enhance lint yield and improve water use efficiency. These examples show that water-saving irrigation can really save water and need to be used at a regional scale. However, to improve the standard of irrigation design and irrigation efficiency requires (i) enforcement or regulation – requiring irrigation to be designed to a standard, and (ii) education – informing water users of the benefits of good design to encourage them to expect a high standard of design from irrigation equipment suppliers (Zhou, Citation2003).
Terracing and contour farming
With frequent farming activities and a high degree of cultivation, sloping land with an angle of 10–258 is highly susceptible to soil erosion. Cultivation on such slopes can lead to erosion of 0.43 cm and 48 t ha−1 of fertile topsoil (Wei et al., Citation2000). Changing such sloping land into contour terraces prevents water and soil erosion, raises land quality and grain yield. Sloping land with an angle of 6–108 can be improved by planting crops along the contour using a 0.5-m deep and 1-m wide trench or ridge to conserve soil and water, improve soil fertility and facilitate sustainable development. In the semiarid Loess Plateau, the building of level terraces has enhanced water infiltration, raised the rainfall utilization rate and created high-yielding farmland while also conserving the soil and water. Combined with other agricultural techniques, it has played a major role in increasing the productivity and sustainability of the region (Deng et al., Citation2000).
Mulching
Mulching with crop residues can improve water use efficiency by 10–20% through reduced soil evaporation and increased plant transpiration. Mulching with crop residues during the summer fallow can increase soil water retention (Feng, Citation1999). Straw mulching can be easily implemented by local farmers because material is easily accessible, low cost and does not contaminate the soil. Combined with N, P and potassium (K) fertilizers, mulching of residues can improve yields by at least 1500 kg ha−1 (Sun & Wang, Citation2001). Plastic film has also been widely used to mulch the soil surface and promote crop growth during early growth when temperatures are low. Several methods of using plastic film have been adopted, including sowing wheat and rice through holes in the plastic, sowing maize and wheat in rows in the furrow with plastic between the rows, and mulching two subsequent crops with the same plastic film. Wang et al. (Citation2004) conducted a field experiment in a loess soil in central Shaanxi Province to identify the effects of rainwater harvesting on WUE and yield of winter wheat. They used ridge-furrow tillage, the ridge being mulched by plastic for rainwater harvesting in the furrows. They demonstrated that mulching significantly increased the harvesting of rainwater and significantly increased yield. Biomass and grain yield in the mulched plots were 39.5 and 28.9% higher than in the corresponding treatments without mulching. The highest WUE of 4.4 kg m−3 for biomass and 2.2 kg m−3 for grain yield was achieved when N fertilizer was added and plant density was kept low.
Plant mineral nutrition
Proper nutrition is the basic need of every living organism. There are now 17 elements which are considered essential for plants to complete their life cycle. These essential plant nutrients are divided into two categories; macronutrients and micronutrients. Macronutrients include carbon (C), hydrogen (H), oxygen (O), nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg) and sulphur (S). Micronutrients are zinc (Zn), copper (Cu), iron (Fe), manganese (Mn), boron (B), molybdenum (Mo), chlorine (Cl) and nickel (Ni). Although silicon (Si) is not essential, it is considered as a beneficial plant nutrient. Adequate nutrition is essential for the integrity of plant structure and key physiological processes such as light interception by chlorophyll (N, Mg), energy for carbohydrate build-up (P), and osmotic regulation of the stomata (K), among others. Therefore, a well-nourished plant is expected to produce more biomass per unit of transpired water than an under-nourished one. Radin and Mathews (Citation1989) also found that N and P deficient plants strongly reduced the hydraulic conductivity of the root cortical cells. These plant nutrients are not only required for better plant growth and development, but also helpful to improve agricultural WUE. Increasing evidence suggests that mineral-nutrient status of plants plays a critical role in enhancing water use efficiency by proper nutrition. This review is an effort to highlight the the role of essential mineral nutrients in improving agricultural WUE in crop plants.
Macro-nutrients
Nitrogen
Nitrogen is important in improving WUE and soil water use. Increased yields and WUE from added N were observed in several dry-land areas where crops were grown on the same land for several years (Shan & Chen, Citation1993). Liu et al. (Citation1998) reported maximum yield and highest WUE under optimum fertilizer inputs of 90 kg N ha−1 and 13 kg P ha−1 in the semiarid field conditions of a loess hilly area in Ningxia. Increased N fertilizer application was positively correlated with yield (correlation coefficient, 0.96) and WUE (correlation coefficient, 0.89) of spring wheat. They also reported that N fertilizer applied in spring wheat improved the development of the root system and especially enhanced the root growth in the cultivated 0–20 cm soil layer. The increased root system in the fertilized plants was able to improve crop water use and nutrient absorption and hence crop yield and WUE were increased. He et al. (Citation1999) conducted experiments to clarify the effects of water and N fertilizer and animal manures on WUE of potatoes. The results showed that both N and water supply very significantly increased WUE. Xu and Zhao (Citation2001) reported that in northern China, the crop WUE increased steadily over 10 years from 0.22 to 0.90 kg m−3 due to technological developments. The increase in WUE was mainly caused by the establishment of water conservation measures, soil improvement, the adoption of new crop varieties and the continuous use of increasing amounts of N fertilizers. Fertilizers including nitrogen have a great potential to further increase the efficiency of water use, especially for crops that grow in autumn when the water supply is plentiful.
Possible mechanisms to enhance WUE by improving N nutrition in crop plants are presented in .
Figure 1. Possible mechanisms through which nitrogen improves water use efficiency in crop plants. Please see text for abbreviations.
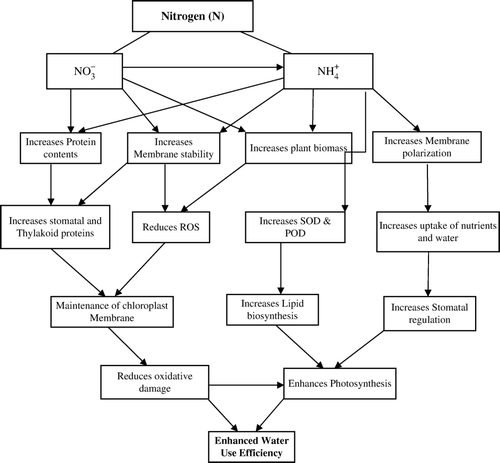
Nitrogen, an important macronutrient, affects photosynthetic capacity of leaves by increasing stromal and thylakoid proteins in leaves (Makino et al., Citation1992; Bungard et al., Citation1997) and being an integral part of DNA, RNA, chlorophyll and other molecules, it plays an important role in cell metabolism. Absorption and utilization of nutrients like N by plants growing under water stress is very critical for plant growth and productivity. Water stress results in a depression in nutrient uptake particularly of N which may contribute towards reduced yield. In addition, although the crop may have roots penetrating the deeper and wetter parts of the soil profile, nutrients concentrated in a dry surface may be unavailable to plants (Wright, Citation1982).
Availability of N and consequently the response to N applied as fertilizers or in other forms is closely related to the ability of plant roots to absorb water from soil. When water inside the plant declines below a threshold level, stomata close which causes a decrease in transpiration resulting in a reduction in water transport through the plant. This in turn, affects the roots’ ability to absorb water and nutrients as effectively as under normal transpiration. Inhibition of nutrient uptake by plants in dry soils also relates to the nutrient transport in soil by mass flow and diffusion (Mackay & Barber, Citation1985; Seiffert et al., Citation1995), which may diminish nutrient availability at the root surface as well as cause a decrease in mineralization of organically bound nutrients (Bloem et al., Citation1992; Walworth, Citation1992). Severe drought may further decrease nutrient transport to the root surface by inducing root shrinkage and which thus causes loss of soil–root contact (North & Nobel, Citation1997).
Inorganic fertilization has been reported to mitigate the adverse effects of water stress by increasing the WUE of crop growth and development (Marschner, Citation1995; Payne et al., Citation1995; Raun & Johnson, Citation1999). Under mild water stress low doses of N increased grain yield in winter wheat, however under severe water conditions high N application proved to be detrimental (Nielsen & Halvorson, Citation1991). Eghball and Maranville (Citation1993) observed a non-significant interaction between drought and N supply for root/shoot ratio, root length, root dry weight, average root radius and nitrogen flux. Kathju et al. (Citation1990) observed that when wheat plants were grown under low (N0P0) and high (N80P80) fertility conditions and water stress was imposed at various stages of a plant's life cycle, increasing intensities of stress adversely affected leaf metabolism and plant performance. However, the performance of plants was better under high fertility conditions, at all stages, under different intensities of water stress. Ashraf et al. (Citation2001) reported that N application minimizes the adverse effect of drought on dry matter and grain yield in pearl millet. This indicates that in dry land agriculture, where water is a limiting factor, fertilizer application to a reasonable extent can be considered for drought mitigation management.
Phosphorus
Phosphorus being a constituent of nucleic acids, phospholipids, phosphor-proteins, dinucleotides and adenosine triphosphate is required for processes including the storage and transfer of energy, photosynthesis, the regulation of some enzymes and the transport of carbohydrates (Hu & Schmidhalter, Citation2001). Soils in arid areas are often calcareous and have high pHs (e.g. those in Mediterranean regions). In the semi-arid tropics, soils are often rich in aluminium and iron oxides, and the pHs are low. Both of these soil types show a strong tendency for P fixation (Oertli, Citation1991). It is generally accepted that the uptake of P by crop plants is reduced in dry-soil conditions (Pinkerton & Simpson, Citation1986). For example, the translocation of P to the shoots is severely restricted even under relatively mild drought stress (Rasnick, Citation1970). However, Liebersbach et al. (Citation2004) reported that the large amount of molecular exudates (i.e. mainly mucilage) from plants in dry soil counteract the reduced mobility of P under such conditions. Turner (Citation1985) pointed out that P deficiency appears to be one of the earliest effects of mild to moderate drought stress in soil-grown plants.
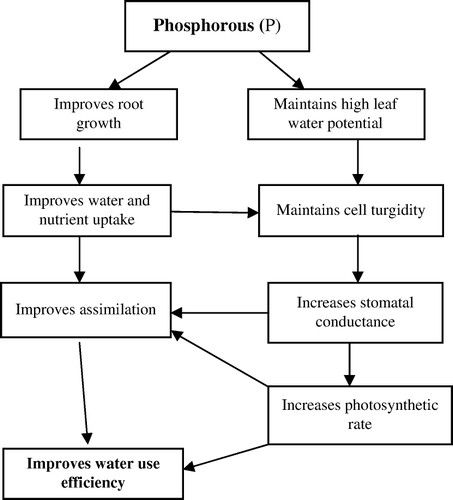
The application of P fertilizer can improve plant growth considerably under drought conditions (Ackerson, Citation1985; Studer, Citation1993; Garg et al., Citation2004). The positive effects of P on plant growth under drought have been attributed to an increase in stomatal conductance (Brück et al., Citation2000), photosynthesis (Ackerson, Citation1985), higher cell-membrane stability, water relations (Sawwan et al., Citation2000) and water-use efficiency. Ajouri et al. (Citation2004) reported that priming seeds with solutions containing the limiting nutrients under drought conditions (such as P and Zn) can improve barley establishment. Smith (Citation2002) suggested that strategies for increasing nutrient uptake by overexpressing genes encoding for high-affinity P transporters are likely to be an important strategy in the future, especially in light of the increasing problems caused by P-deficient soils of the semi-arid tropics. Possible mechanisms to enhance the WUE by improving P nutrition in crop plants are presented in .
Figure 2. Possible mechanisms through which phosphorus improves water use efficiency in crop plants.
Potassium
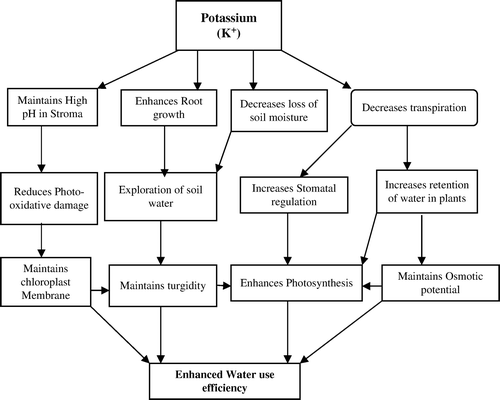
Potassium is an essential factor in protein synthesis, glycolytic enzymes and photosynthesis; an osmoticum mediating cell expansion and turgor-driven movements; and a competitor of Na+ under saline conditions (Marschner, Citation1995). Numerous studies have shown that the application of K fertilizer mitigates the adverse effects of drought on plant growth (Andersen et al., Citation1992; Studer, Citation1993; Sangakkara et al., Citation2001). Potassium increases the plant's drought resistance through its functions in stomatal regulation, osmoregulation, energy status, charge balance, protein synthesis and homeostasis (Beringer & Trolldenier Citation1978; Marschner, Citation1995). It also maintains turgor pressure (Mengel & Arneke, Citation1982) and reduces transpiration under drought conditions (Andersen et al., Citation1992). In plants coping with drought stress, the accumulation of K+ may be more important than the production of organic solutes during the initial adjustment phase, because osmotic adjustment through ion uptake like K+ is more energy efficient (Hsiao, Citation1973). Working with wheat, Morgan (Citation1992) showed that the lines displaying high osmotic adjustments had a high accumulation of K+ in their tissues. In these lines, K+ accounted for about 78% of all osmotica. By contrast, amino acids, which were the only other important contributor, constituted only about 22%. A recent study by Ma et al. (Citation2004) showed that K+ accumulation in the expanding leaves in three genotypes of Brassica napus oilseeds accounted for about 25% of drought-induced changes in osmotic adjustment. Possible mechanisms to enhance WUE by improving K nutrition in crop plants are presented in . Potassium nutrition increases crop WUE by utilizing the soil moisture more efficiently than in K-deficient plants. The positive effects of K on water use efficiency may be through promotion of root growth accompanied by a greater uptake of nutrients and water by plants (Rama Rao, Citation1986) and through the reduction of transpirational water loss (Beringer & Trolldenier, Citation1978). Also, K maintains the osmotic potential and turgor of the cells (Hsiao, Citation1973; Lindhauer ,Citation1985) and regulates stomatal functioning under water stress conditions (Umar et al., Citation1993; Nandwal et al., Citation1988; Kant & Kafkafi, Citation2002), which is reflected in improved crop yield under water limited conditions (Umar & Bansal, Citation1997; Umar & Moinuddin, Citation2002). Besides, it takes part in many essential processes in plants (Marschner, Citation1995) and enhances photosynthetic rate, plant growth and yield under stress conditions (Sharma et al., Citation1996; Tiwari et al., Citation1998; Egila et al., Citation2001; Umar & Moinuddin, Citation2002). The protective role of K in plants suffering from water stress has been attributed to the maintenance of a high pH in stroma and against the photo-oxidative damage to chloroplasts (Cakmak, Citation1997).
Figure 3. Possible mechanisms through which potassium improves water use efficiency in crop plants.
Calcium
Calcium plays a vital role in regulating many physiological processes that influence both growth and responses to environmental stresses. Included among these are water and solute movement as influenced through the effects of Ca2 + on membrane structure and stomatal function, cell division and cell-wall synthesis, direct or signalling roles in systems involved in plant defence and repair of damage from biotic and abiotic stress, and rates of respiratory metabolism and translocation (McLaughlin & Wimmer, Citation1999).
As for many other elements, the addition of supplemental Ca2 + has beneficial effects. For instance, supplemental Ca2 + in the growth medium increased the relative growth rate of barley under abiotic stress conditions (Cramer et al., Citation1990). Similarly, Hu and Schmidhalter (Citation1997) reported that although a biotic stress significantly reduced the Ca2 + accumulation in wheat leaves, an increase in the Ca2 + concentration in the nutrient solution enhanced Ca2 + accumulation. However, Hu and Schmidhalter (Citation1998) later found no difference in Ca2 + accumulation between the control and stress treatments in growing leaves of wheat plants grown in soil. Supplemental Ca2 + has also been reported to alleviate the adverse effects of stress on the germination and vegetative growth of bean (Awada et al., Citation1995). By contrast, various Ca2 +:Na+ ratios had no significant effect on the uptake of Na+ by rice (Yeo & Flowers, Citation1985).
In recent years, intracellular Ca2 + has been found to regulate the responses of plants to drought and has also been implicated in the transduction of drought stress signals in plants, which play an essential role in osmoregulation under this condition (Knight et al., Citation1997; Bartels & Sunkar, Citation2005). Key discoveries include drought-induced expression of Ca2 +-dependent protein kinases (Urao et al., Citation1994) and hyperosmotic shock-induction of putative Ca2 +-binding proteins (Ko & Lee, Citation1996). Sadiqov et al. (Citation2002) found that Ca2 + participates in signalling mechanisms of drought-induced proline accumulation for osmotic adjustment. They supported the findings of Knight et al. (Citation1997), who indicated that this was due to the induction of a transcript for proline biosynthetic enzyme (P5CS) being inhibited in the presence of Ca2 + channels in Arabidopsis (also shown in wheat and maize; Nayyar, Citation2003).
Magnesium and sulphur
Little information is available on the role of Mg and S nutrition in enhancing water use efficiency of plants. However, we expect that drought reduces both Mg and S uptake because Mg deficiency is occurring more frequently as cropping intensity increases and because the decrease in atmospheric S during the past decade has been associated with the appearance of S-deficiency symptoms (Scherer, Citation2001) and has had severe consequences for S nutrition and crop production.
Magnesium is involved in numerous physiological and biochemical processes in plants affecting growth and development. Magnesium is exceptional in activating more enzymes than any other mineral nutrient (Epstein & Bloom, Citation2004). Some examples of Mg-activated enzymes include ATPases, ribulose-1, 5-bisphosphate (RuBP) carboxylase, RNA polymerase and protein kinases (Marschner, Citation1995; Shaul, Citation2002). The role of Mg as the central atom of the chlorophyll (Chl) molecule is perhaps the best-known function of Mg in plants in that it is associated with the development of leaf chlorosis, typically interveinal, under Mg deficiency stress. Depending on the Mg status of the plant, between 6% and 35% of the total Mg may be bound in the chloroplasts, a higher proportion being associated with a lower Mg status (Scott & Robson, Citation1990). With the onset of Mg deficiency, there is firstly a decline in the concentration of Chl b, which is followed by that of Chl a (Hermans et al., Citation2004). An initial decrease in Chl concentration in Mg deficient plants has been ascribed to early accumulation of sugars in leaves, rather than to low levels of Mg (Hermans et al., Citation2004). According to Hermans et al. (Citation2004), high accumulation of sugars may repress the expression of the cab2 gene that is responsible for encoding Chl a and b protein. Magnesium deficiency symptoms vary between plant species, but there are some similarities. As Mg is a relatively mobile nutrient in plants, deficiency always appears first in the older leaves (Bergmann, Citation1992). As discussed below in detail, development of Mg deficiency symptoms is highly affected by light intensity. By increasing light intensity, plants under low Mg supply rapidly develop interveinal chlorosis and reddish spots on the leaves (Marschner & Cakmak, Citation1989). Moreover, for leaves with the same low concentration of Mg, development of leaf chlorosis and necrosis is significantly accentuated by increasing light intensity (Marschner & Cakmak, Citation1989). In crops, Mg deficiency is especially associated with strongly leached sandy acid soils with a low cation exchange capacity. One of the early reactions of Mg deficiency stress in plants is its impact on partitioning of dry matter between roots and shoots. An increasing body of evidence indicates that Mg plays a fundamental role in phloem export of photosynthates from the source to the sink organs, and its deficiency results in dramatic increases in accumulation of carbohydrates in the source leaves (Cakmak et al., Citation1994a, Citation1994b; Marschner et al., Citation1996; Hermans et al., Citation2004). Reduced transport and hence accumulation of carbohydrates in Mg-deficient leaves cause alterations in photosynthetic carbon metabolism and restrict CO2 fixation. Impairment of the photosynthetic electron transport to CO2 through photosynthetic membranes may cause an accumulation of non-utilized electrons and absorbed energy. Under such conditions, the electrons and excitation energy not used in photosynthetic CO2 fixation is channelled to molecular O2, leading to the generation of highly reactive O2 species (ROS) and consequently to damage of chloroplast constituents such as Chl and membrane lipids (Marschner & Cakmak, Citation1989; Mittler, Citation2002; Asada, Citation2006).
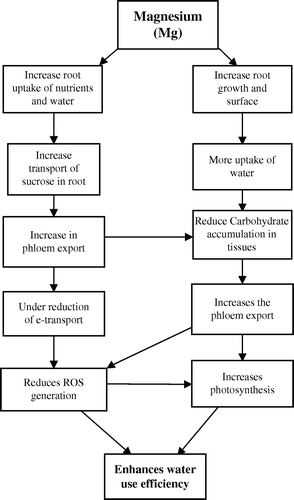
Possible mechanisms to enhance water use efficiency in crop plants by improving Mg nutrition are presented in . Magnesium increases the root growth and root surface area which helps to increase uptake of water and nutrients by roots and transport of sucrose from leaves to roots (). Magnesium improves carbohydrate translocation by increasing phloem export and reduces ROS generation and photo-oxidative damage to chloroplast under water-limited conditions. Maintenance of chloroplast structure by improving Mg nutrition enhances the photosynthetic rate under water stress which in turn improves the water use efficiency.
Figure 4. Possible mechanisms through which magnesium improves water use efficiency in crop plants.
Micro nutrients
The contributions of micro nutrients in enhancing water use efficiency are not well-defined. Micronutrients help the macro nutrients in enhancing WUE by activation of certain physiological, biochemical and metabolic processes within the plant body. The role of micronutrients in drought alleviation is discussed below.
Boron
Boron is directly or indirectly involved in several physiological and biochemical processes during plant growth such as cell elongation, cell division, cell wall biosynthesis, membrane function, nitrogen (N) metabolism, leaf photosynthesis and uracil synthesis (Marschner, Citation1995; Zhao & Oosterhuis, Citation2002).
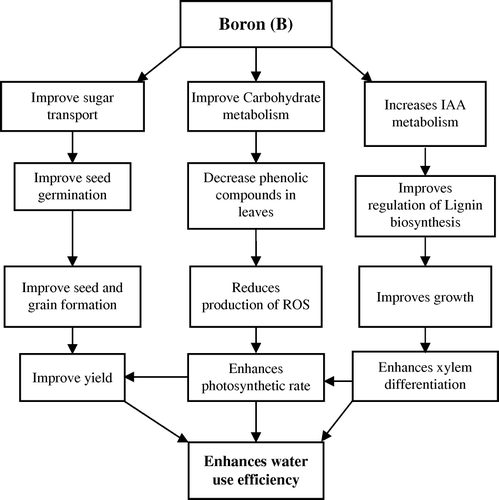
Boron nutrition improves cell enlargement in growing tissues because of its structural role in the cell wall (O'Neill et al., Citation2004). In higher plants, B application induces changes in carbohydrate metabolism (Camacho-Cristobal & Gonzalez-Fontes, Citation1999), which in turn is responsible for the decrease in concentration of phenolic compounds in leaves (Blevins & Lukaszewski, Citation1998; Camacho-Cristobal et al., Citation2002). The low level of these phenolics oxidized to derivatives such as quinones, reduce the production of extremely reactive oxygen species (ROS) and cause reduction in peroxidative damage to vital components of cell membrane such as lipids and proteins, which eventually leads to repair of several cellular functions (Cakmak & Romheld, Citation1997; El-Shintinawy, Citation1999; del Rio et al., Citation2003). Thus, B may also protect plasma membrane against peroxidative damage by toxic O2 species. The role of B in reproduction of higher plants is well established, and its deficiency is responsible for creating male sterility and inducing floral abnormalities (Sharma, Citation2006). Apart from this, B deficiency is known to influence indirectly the activities of numerous enzymes such as peroxidase, ribonuclease, indole acetic acid (IAA) oxidase, and polyphenol oxidase in several crop plants. A key role for B has also been suggested in IAA metabolism and in regulation of lignin biosynthesis and xylem differentiation (Marschner, Citation1995). Boron nutrition improves sugar transport in the plant body which helps to improve seed germination and seed grain formation. This in turn improves yields and enhances water use efficiency (). Boron application also improves carbohydrate metabolism and decreases the phenolic compounds in leaves. This in turn reduces the production of ROS and enhances the photosynthetic rate which results in improved yield and water use efficiency ().
Figure 5. Possible mechanisms through which boron improves water use efficiency in crop plants.
Zinc
Zinc is an essential nutrient for plant growth and development. Crop productivity is limited due to its deficiency (Irshad et al., Citation2004; Rashid & Ryan, Citation2004). Zinc is involved in cellular functions such as protein metabolism, photosynthetic carbon metabolism and indole acetic acid (IAA) metabolism (Marschner, Citation1995). Nutritional stress of zinc may affect plant water status, net CO2 assimilation rate, stomatal conductance and transpiration rate (Ben-Rouina et al., Citation2006). Zinc is a component of plant carbonic anhydrase (CA) enzyme (Tobin, Citation1970) and it is located in the chloroplast and cytoplasm in C3 plants (Okhi, Citation1976). A decline in CA activity has been reported in plants due to Zn deficiency (Okhi, Citation1976). Earlier researchers (Nelson et al., Citation1969; Hatch & Slack, Citation1970) reported that CA may be directly involved in photosynthesis, facilitating the diffusion of CO2 through the liquid phase of the cell to the chloroplast. Sharma et al. (Citation1995) observed that Zn deficiency caused decrease in CA activity, stomatal aperture and transpiration in leaves of cauliflower.
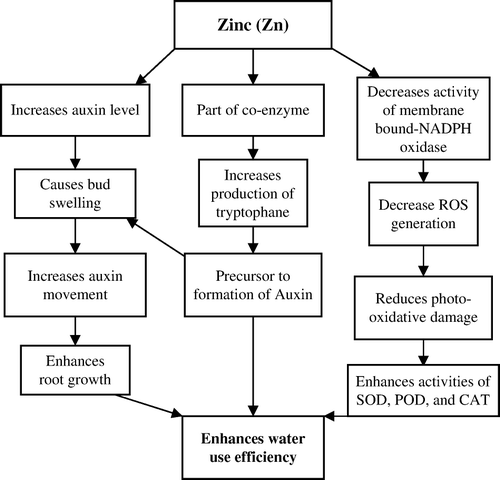
Possible mechanisms to minimize detrimental effects of drought in crop plants by improving Zn nutrition are presented in . Zinc is important for its ability to influence auxin levels and has long been known to be a co-enzyme for production of tryptophan, a precursor to the formation of auxin (Bennett & Skoog, Citation2002). In most trees, the general sequence of activity in early spring is auxin production, bud swell, auxin movement, auxin-directed transport, root growth, followed by phloem and xylem differentiation (Seeley, Citation2006). As indicated above, normal auxin functions are likely to be disrupted in drought conditions. Maintaining adequate hormone levels gives a competitive advantage to withstand adverse conditions of all kinds. In another mechanism, Zn application reduces the activity of membrane-bound NADPH oxidase which in turn decreases the generation of ROS (Cakmak & Marschner, Citation1988) and photo-oxidation processes reduce (Marschner & Cakmak, Citation1989), while the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) enhance (Yu et al., Citation1998). Zinc plays an important role in lowering ROS generation and defending cells against ROS attack (Marschner, Citation1995; Cakmak, Citation2000).
Figure 6. Possible mechanisms through which zinc improves water use efficiency in crop plants. Please see text for abbreviations.
Silicon
Silicon (Si) has been regarded as an essential element in a number of species of the Poaceae and Cyperaceae, but it has not been possible to demonstrate that it is essential to all higher plants because direct evidence is still lacking that it is part of a molecule of an essential plant constituent or metabolite (Epstein, Citation1994, Citation1999). Recently, the role of Si in plant metabolism has received increasing attention. Liang et al. (Citation2003) strongly suggest that Si may be involved in metabolic or physiological and/or structural activity in higher plants exposed to abiotic and biotic stresses. It has been reported that Si increases plant tolerance to high manganese (Mn) concentrations (Horst & Marschner, Citation1978), drought (Lux et al., Citation2002), heavy metals (Neumann & zur Nieden, Citation2001) and plant resistance to pests and pathogens (Belanger et al., Citation2003; Richmond & Sussman, Citation2003). However, the use of Si and its relationships with plant growth and with mineral nutrition of plants under water stress have not been studied in such detail.
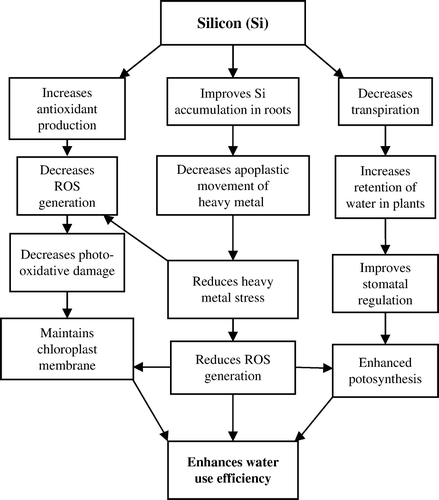
The possible mechanisms to enhance WUE in crop plants by improving silicon nutrition are presented in . Silicon enhances WUE by reducing the detrimental effects of drought (Epstein, Citation1999; Ma & Takahashi, Citation2002, Richmond & Sussman, Citation2003; Ma, Citation2004). Silicon deposition in cell walls of roots reduces apoplastic bypass flow and provides binding sites for metals, resulting in decreased uptake and translocation of toxic metals and reduces heavy metal stress injury to the roots. This will improve the root activity and enhances the WUE in plants. Deposition of Si in the culms, leaves and hulls enhances the strength and rigidity of cell walls and decreases transpiration from the cuticle and, thus, increases the resistance to lodging and enhances WUE. Under water-limited conditions, Si-alleviated effects have been associated with an increase in antioxidant defence abilities (Liang et al., Citation2003; Zhu et al., Citation2004). However, this might be a beneficial result of Si as opposed to a direct effect because it is unlikely that Si affects the activity of antioxidant enzymes. Silicon nutrition increases antioxidant production and reduces ROS generation which in turn reduces photo-oxidative damage and maintains the integrity of chloroplast membrane. This enhances the photosynthetic rate of the plants which in turn enhances productivity and water use efficiency in plants.
Figure 7. Possible mechanisms through which silicon improves water use efficiency in crop plants.
Other nutrients
The role of micro nutrients (iron, manganese, molybdenum and chloride) in enhancing water use efficiency is not well-defined. The indirect role of these nutrients in enhancing water use efficiency is described below.
Iron (Fe) is involved in the production of chlorophyll. It is a component of many enzymes associated with energy transfer, nitrogen reduction and fixation, and lignin formation. Iron is associated with sulphur in plants to form compounds that catalyse other reactions. Drought-induced deficiency of Fe causes yellowing of leaves due to low levels of chlorophyll. Leaf yellowing first appears on the younger upper leaves in interveinal tissues. Severe Fe deficiencies cause leaves to turn completely yellow or almost white, and then brown as leaves die. Uptake of Fe decreases with increased soil pH, and is adversely affected by high levels of available phosphorus, manganese and zinc in soils.
Manganese (Mn) is necessary in photosynthesis, nitrogen metabolism and to form other compounds required for plant metabolism. Interveinal chlorosis is a characteristic Mn-deficiency symptom. In very severe Mn deficiency cases, brown necrotic spots appear on leaves, resulting in premature leaf drop. Delayed maturity is another deficiency symptom in some species. Manganese has no direct role in enhancing WUE. It can reduce the adverse effects of drought and improve WUE indirectly by enhancing the photosynthetic rate, and nitrogen metabolism in the plant body. Manganese nutrition reduces interveinal chlorosis, brown necrotic spots on leaves and reduces premature leaf drop.
Molybdenum (Mo) can reduce the adverse effects of water stress and improve WUE indirectly due to its involvement in enzyme systems relating to nitrogen fixation by bacteria growing symbiotically with legumes, nitrogen metabolism, protein synthesis and sulphur metabolism. It has a significant effect on pollen formation, so fruit and grain formation are affected by Mo nutrition in plants.
Chloride (Cl) is a mobile anion in plants, most of its functions relate to salt effects (stomatal opening) and electrical charge balance in physiological functions in plants. Chloride also indirectly affects plant growth by stomatal regulation of water loss. Chloride enhances WUE by reduction in wilting and restricting highly branched root systems in cereal crops. The role of Cl in decreasing the incidence of various diseases in small grains is perhaps more important than its nutritional role from a practical viewpoint.
Conclusions
Improving agricultural water use efficiency will continue to increase because of the demand for increased grain production. Greater yield per unit of water is one of the most important challenges in water-limited agriculture. In plants, drought can cause reduction in photosynthetic rate, chlorophyll content, stomatal conductance, transpiration rate and relative leaf water content, and destruction of some physiological processes, which ultimately reduce plant growth and development. An environmental stress like drought contributes significantly to reduce crop yields well below the potentially maximum yields which ultimately results in the reduction in WUE. Due to these risks, it is necessary to minimize the detrimental effects of drought in plants (primary producers) below permissible limits. The management of plant nutrients is very helpful to enhance WUE by reducing the detrimental effects of drought. Better plant nutrition can be helpful to utilize the available water more efficiently by a number of mechanisms. Application of nutrients like N, K, Mg, B, Zn and Si reduce the toxicity of reactive oxygen species (ROS) produced under water-limited conditions by increasing the concentration of antioxidants like superoxide dismutase (SOD); Catalase (CAT) and peroxidase (POD) in the plant cells. These antioxidants scavenge ROS and reduce photo-oxidation and maintain the integrity of chloroplast membrane and increase the photosynthetic rate in the crop plants which in turn enhances the WUE. Some plant nutrients like P, K, Mg and Zn improve root growth. Improvement in root growth increases the intake of water which helps in stomatal regulation and enhances the photosynthetic rate and increases water use efficiency. Potassium, calcium and chloride (K, Ca, Cl) applications help in osmotic adjustment. These nutrients help to maintain high tissue water potential and improve the WUE under water stress condition. The micronutrients like Fe, B, Mn and Mo improve the WUE by alleviating the adverse effects of drought indirectly by activating the physiological, biochemical and metabolic processes in the plants. The literature available on this aspect is insufficient to fully understand the role of plant micronutrients to enhance the WUE. Therefore, more future research is required for better understanding of interactions between WUE and plant micro nutrients in soil–plant systems.
References
- Ackerson , R. C. 1985 . Osmoregulation in cotton in response to water-stress. 3. Effects of phosphorus fertility . Plant Physiology , 77 : 309 – 312 .
- Ajouri , A. , Asgedom , H. and Becker , M. 2004 . Seed priming enhances germination and seedling growth of barley under conditions of P and Zn deficiency . Journal of Plant Nutrition and Soil Science , 167 : 630 – 636 .
- Ali , M. H. and Talukder , M. S. U. 2008 . Increasing water productivity in crop production – A synthesis . Agricultural Water Management , 95 : 1201 – 1213 .
- Andersen , M. N. , Jensen , C. R. and Lösch , R. 1992 . The interaction effects of potassium and drought in field-grown barley. 1. Yield, water-use efficiency and growth . Acta Agriculturae Scandinavica Section B – Soil & Plant Science , 42 : 34 – 44 .
- Asada , K. 2006 . Production and scavenging of reactive oxygen species in chloroplast and their functions . Plant Physiology , 141 : 391 – 396 .
- Ashraf , M. , Shehbaz , M. and Ashraf , M. Y. 2001 . Influence of nitrogen supply and water stress on growth and nitrogen, phosphorous, potassium and calcium contents in pearl millet . Biologia Planterum , 44 : 459 – 462 .
- Awada , S. , Campbell , W. F. , Dudley , L. M. , Jurinak , J. J. and Khan , M. A. 1995 . Interactive effects of sodium-chloride, sodium-sulfate, calcium-sulfate, calcium-chloride on snapbean growth, photosynthesis, ion uptake . Journal of Plant Nutrition , 18 : 889 – 900 .
- Bai , Q. J. and Dong , S. T. 2001 . Agricultural high-efficient water usage and sustainable development of water saving agriculture . Journal of Shandong Agriculture University , 32 : 331 – 335 .
- Bartels , D. and Sunkar , R. 2005 . Drought and salt tolerance in plants . Critical Reviews in Plant Sciences , 24 : 23 – 58 .
- Belanger , R. R. , Benhamou , N. and Menzies , J. G. 2003 . Cytological evidence of an active role of silicon in wheat resistance to powdery mildew (Blumeria graminis f. sp. tritici) . Phytopathology , 93 : 402 – 412 .
- Bennett , J. P. and Skoog , F. 2002 . Preliminary experiments on the relation of growth-promoting substances to the rest period in fruit trees . Plant Physiology , 13 : 219 – 225 .
- Ben-Rouina , B. , Chedli , B. A. , Rehman , H. A. and Muhammad , B. 2006 . Water relations, proline accumulation and photosynthetic activity in olive tree (Olea europaea L. cv. “Chemlali”) in response to salt stress . Pakistan Journal of Botany , 38 : 1397 – 1406 .
- Bergmann , W. 1992 . Nutritional disorders of plants –development, visual and analytical diagnosis , Stuttgart : Fischer Verlag .
- Beringer , H. and Trolldenier , G. 1978 . “ Influence of K nutrition on the response to environmental stress ” . In Potassium research – review and trends. Proceedings of the 11th Congress of the International Potash Institute , 189 – 222 . Bern : International Potash Institute .
- Blevins , D. G. and Lukaszewski , K. M. 1998 . Boron in plant structure and function . Annual Review of Plant Physiology and Molecular Biology , 49 : 481 – 500 .
- Bloem , J. , De Ruiter , P. C. , Koopman , G. J. and Brussaard , L. 1992 . Microbial numbers and activity in dried and rewatered arable soil under integrated and conventional management . Soil Biology and Biochemistry , 24 : 655 – 665 .
- Brück , H. , Payne , W. A. and Sattelmacher , B. 2000 . Effects of phosphorus and water supply on yield, transpirational water-use efficiency, and carbon isotope discrimination of pearl millet . Crop Science , 40 : 120 – 125 .
- Bungard , R. A. , McNeil , A. and Morton , J. D. 1997 . Effect of nitrogen on the photosynthetic apparatus of Clematis vitalba grown at several irradiances . Australian Journal of Plant Physiology , 24 : 205 – 214 .
- Cakmak , I. 1997 . “ Role of potassium in protecting higher plants against photo-oxidative damage ” . In Food security in the WANA region, the essential need for balanced fertilization , Edited by: Johnston , A. E. 345 – 352 . Basel : International Potash Institute .
- Cakmak , I. 2000 . Possible roles of zinc in protecting plant cells from damage by reactive oxygen species . New Phytologist , 146 : 185 – 205 .
- Cakmak , I. and Marschner , H. 1988 . Zinc-dependent changes in ESR signals, NADPH oxidase and plasma membrane permeability in cotton roots . Physiologia Plantarum , 73 : 132 – 186 .
- Cakmak , I. and Romheld , V. 1997 . Boron deficiency-induced impairments of cellular functions in plants . Plant and Soil , 193 : 71 – 83 .
- Cakmak , I. , Hengeler , C. and Marschner , H. 1994a . Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency . Journal of Experimental Botany , 45 : 1245 – 1250 .
- Cakmak , I. , Hengeler , C. and Marschner , H. 1994b . Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants . Journal of Experimental Botany , 45 : 1251 – 1257 .
- Camacho-Cristobal , J. J. and Gonzalez-Fontes , A. 1999 . Boron deficiency causes a drastic decrease in nitrate content and nitrate reductase activity . and increases the content of carbohydrates in leaves from tobacco plants. Planta , 209 : 528 – 536 .
- Camacho-Cristobal , J. J. , Anzellotti , D. and Gonzalez-Fontes , A. 2002 . Changes in phenolic metabolism of tobacco plants during short-term boron deficiency . Plant Physiology and Biochemistry , 40 : 997 – 1002 .
- Cramer , G. R. , Epstein , E. and Läuchli , A. 1990 . Effects of sodium, potassium and calcium on salt-stressed barley. 1. Growth analysis . Physiologia Plantarum , 80 : 83 – 88 .
- del Rio , L. A. , Corpas , F. J. , Sandalio , L. M. , Palma , J. M. and Barroso , J. B. 2003 . Plant peroxisomes, reactive oxygen metabolism, and nitric oxide . IUBMB Life , 55 : 71 – 81 .
- Deng , X. P. , Wang , S. Q. and Peng , K. S. 2000 . Assessment of food productive potential on the different land type in the middle-scale eco-agricultural research area of Yan'an . Journal of Chinese Soil Water Conservation , 14 : 87 – 91 .
- Eghball , B. and Maranville , J. W. 1993 . Root development and nitrogen influx of corn genotypes grown under combined drought and nitrogen stresses . Agronomy Journal , 85 : 147 – 152 .
- Egila , J. N. Jr , Davies , F. T. and Drew , M. C. 2001 . Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. Leprechaun: plant growth, leaf macro and micronutrient content and root longevity . Plant and Soil , 229 : 213 – 224 .
- El-Shintinawy , F. 1999 . Structural and functional damage caused by boron deficiency in sunflower leaves . Photosynthetica , 36 : 565 – 573 .
- Epstein , E. 1994 . The anomaly of silicon in plant biology . Proceedings of the National Academy of Sciences USA , 91 : 11 – 17 .
- Epstein , E. 1999 . Silicon . Annual Review in Plant Physiology and Molecular Biology , 50 : 641 – 664 .
- Epstein , E. and Bloom , A. J. 2004 . Mineral nutrition of plants: Principles and perspectives , 2nd edn , Sunderland, MA : Sinauer Associates .
- Food and Agriculture Organization 2003 . World agriculture: towards 2015/2030 . London : Earthscan Publications .
- Feng , H. C. 1999 . Effects of straw mulching on soil conditions and grain yield of winter wheat . Chinese Bulletin and Soil Science , 30 : 174 – 175 .
- Gamo , M. 1999 . Classification of arid regions by climate and vegetation . Journal of Arid Land Studies , 1 : 9 – 17 .
- Garg , B. K. , Burman , U. and Kathju , S. 2004 . The influence of phosphorus nutrition on the physiological response of moth bean genotypes to drought . Journal of Plant Nutrition and Soil Science , 167 : 503 – 508 .
- Hatch , M. D. and Slack , C. R. 1970 . Photosynthetic CO2 fixation pathways . Annual Review in Plant Physiology , 21 : 141 – 162 .
- He , H. , Cheng , G. L. and Zhao , S. W. 1999 . Effect of different water and fertilizer conditions on water use efficiency of potato . Agricultural Research in Arid Areas , 17 : 59 – 66 .
- Hermans , C. , Johnson , G. N. , Strasser , R. J. and Verbruggen , N. 2004 . Physiological characterization of magnesium deficiency in sugar beet: acclimation to low magnesium differentially affects photosystems I and II . Planta , 220 : 344 – 355 .
- Horst , W. J. and Marschner , H. 1978 . Effect of silicon and manganese tolerance of bean plants (Phaseolus vulgaris L.) . Plant and Soil , 50 : 287 – 303 .
- Hsiao , T. C. 1973 . Plant responses to water stress . Annual Review in Plant Physiology , 24 : 519 – 570 .
- Hu , S. J. , Song , Y. D. , Zhou , H. F. and Tian , C. Y. 2002 . Experimental study on water use efficiency of cotton in the Tarim River Basin . Agricultural Research in Arid Areas , 20 : 65 – 70 .
- Hu , Y. and Schmidhalter , U. 1997 . Interactive effects of salinity and macronutrient level on wheat. 2. Composition . Journal of Plant Nutrition , 20 : 1169 – 1182 .
- Hu , Y. and Schmidhalter , U. 1998 . Spatial distributions and net deposition rates of mineral elements in the elongating wheat (Triticum aestivum L.) leaf under saline soil conditions . Planta , 204 : 212 – 219 .
- Hu , Y. and Schmidhalter , U. 2001 . Effects of salinity and macronutrient levels on micronutrients in wheat . Journal of Plant Nutrition , 24 : 273 – 281 .
- Irshad , M. , Maqsood , A. G. , Tariq , A. , Rehmat , U. and Iftikhar , A. 2004 . Growth response of cotton lines contrasting for QTL markers associated with drought tolerance . Crop Science , 38 : 835 – 842 .
- Kant , S. and Kafkafi , U. 2002 . “ Potassium and abiotic stresses in plants ” . In Role of potassium in nutrient management for sustainable crop production in India , Edited by: Pasricha , N. S. and Bansal , S. K. Gurgaon, Haryana : Potash Research Institute of India .
- Kathju , S. , Vyas , S. P. , Garg , B. K. , & Lahiri , A. N. 1990 . Fertility induced improvement in performance and metabolism of wheat under different intensities of water stress . Proceedings of the International Congress of Plant Physiology , 88 , 854 858 . New Delhi, India .
- Knight , H. , Trewavas , A. J. and Knight , M. R. 1997 . Calcium signalling in Arabidopsis thaliana responding to drought and salinity . Plant Journal , 12 : 1067 – 1078 .
- Ko , J. H. and Lee , S. H. 1996 . Biochemical studies of purified 23 kD calcium-binding protein in Dunaliella salina and its cDNA cloning . Plant Physiology , 111 : 714 – 720 .
- Liang , Y. C. , Chen , Q. , Liu , Q. , Zhang , W. H. and Ding , R. X. 2003 . Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.) . Journal of Plant Physiology , 160 : 1157 – 1164 .
- Liebersbach , H. , Steingrobe , B. and Claassen , N. 2004 . Roots regulate ion transport in the rhizosphere to counteract reduced mobility in dry soil . Plant and Soil , 260 : 79 – 88 .
- Lindhauer , M. G. 1985 . Influence of K nutrition and drought and water stressed sunflower plants differing in K nutrition . Journal of Plant Nutrition , 10 : 1965 – 1973 .
- Liu , H. J. , Kang , Y. H. and Liu , S. P. 2003 . Regulation of field environmental condition by sprinkler irrigation and its effect on water use efficiency of winter wheat . Transactions of the Chinese Society of Agricultural Engineering , 19 : 46 – 51 .
- Liu , Z. M. , Shan , L. , Deng , X. P. , Inanaga , S. , Sunohara , W. and Harada , J. 1998 . Effects of fertilizer and plant density on the yields, root system and water use of spring wheat . Research in Soil Water Conservation , 5 : 70 – 75 .
- Lux , A. , Luxova , M. , Hattori , T. , Inanaga , S. and Sugimoto , Y. 2002 . Silicification in sorghum (Sorghum bicolor) cultivars with different drought tolerance . Physiologia Plantarum , 115 : 87 – 92 .
- Ma , J. F. 2004 . Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses . Soil Science & Plant Nutrition , 50 : 11 – 18 .
- Ma , J. F. and Takahashi , E. 2002 . Soil, fertilizer, and plant silicon research in Japan , Amsterdam : Elsevier Science .
- Ma , Q. F. , Turner , D. W. , Levy , D. and Cowling , W. A. 2004 . Solute accumulation and osmotic adjustment in leaves of Brassica oilseeds in response to soil water deficit . Australian Journal of Agricultural Research , 55 : 939 – 945 .
- Mackay , A. D. and Barber , S. A. 1985 . Soil moisture effects on root growth and phosphorus uptake by corn . Agronomy Journal , 177 : 519 – 523 .
- Makino , A. , Sakashita , H. , Hidema , J. , Mae , T. , Ojima , K. and Osmond , B. 1992 . Distinctive responses of ribulose-1, 5-bisphosphate carboxylase and carbonic anhydrase in wheat leaves to nitrogen nutrition and their possible relationships to CO2 transfer resistance . Plant Physiology , 100 : 1737 – 1743 .
- Marschner , H. 1995 . Mineral nutrition of higher plants , 2nd edn , San Diego, CA : Academic Press .
- Marschner , H. and Cakmak , I. 1989 . High light intensity enhances chlorosis and necrosis in leaves of zinc potassium, and magnesium deficient bean (Phaseolus vulgaris L.) plants . Journal of Plant Physiology , 134 : 308 – 315 .
- Marschner , H. , Kirkby , E. A. and Cakmak , I. 1996 . Effect of mineral nutritional status on shoot-root partitioning of photo assimilates and cycling of mineral nutrients . Journal of Experimental Botany , 47 : 1255 – 1263 .
- McLaughlin , S. B. and Wimmer , R. 1999 . Transley Review No. 104 – Calcium physiology terrestrial ecosystem processes . New Phytologist , 142 : 373 – 417 .
- Mengel , K. and Arneke , W. W. 1982 . Effect of potassium on the water potential, the pressure potential, the osmotic potential and cell elongation in leaves of Phaseolus vulgaris . Physiologia Plantarum , 54 : 402 – 408 .
- Mittler , R. 2002 . Oxidative stress, antioxidants, and stress tolerance . Trends in Plant Science , 7 : 405 – 410 .
- Morgan , J. M. 1992 . Osmotic components and properties associated with genotypic differences in osmoregulation in wheat . Australian Journal of Plant Physiology , 19 : 67 – 76 .
- Nandwal , A. S. , Hooda , A. and Datta , D. 1988 . Effect of substrate moisture and potassium on water relation and C, N and K distribution in Vigan radiata . Biologia Plantarum , 41 : 149 – 153 .
- Nayyar , H. 2003 . Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists . Environmental and Experimental Botany , 50 : 253 – 264 .
- Nelson , E. B. , Cenedella , A. and Tolbert , N. E. 1969 . Carbonic anhydrase in Chlamydomonas . Phytochemistry , 8 : 2305 – 2306 .
- Neuman , D. and zur Nieden , U. 2001 . Silicone and heavy metal tolerance of higher plants . Phytochemistry , 56 : 685 – 692 .
- Nielsen , D. C. and Halvorson , A. D. 1991 . Nitrogen fertility influence on water stress and yield of winter wheat . Agronomy Journal , 83 : 1065 – 1070 .
- North , G. B. and Nobel , P. S. 1997 . Root–soil contact for the desert succulent Agava deserti in wet and drying soil . Journal of Experimental Botany , 38 : 2068 – 2081 .
- Oertli , J. J. 1991 . Nutrient management under water and salinity stress Proceedings of the symposium on nutrient management for sustained productivity , pp. 138 165 . Department of Soils, Punjab Agricultural University Ludhiana, , India .
- O'Neill , M. A. , Ishii , T. , Albusheim , P. and Darvill , A. G. 2004 . Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharide . Annual Review of Plant Biology , 55 : 109 – 139 .
- Ohki , K. 1976 . Effect of zinc nutrition on photosynthesis and carbonic anhydrase activity in cotton . Physiologia Plantarum , 38 : 300 – 304 .
- Payne , W. A. , Hossner , L. R. , Onken , A. B. and Wedt , C. W. 1995 . Nitrogen and phosphorus uptake in pearl millet and its relation to nutrient and transpiration efficiency . Agronomy Journal , 87 : 425 – 431 .
- Pinkerton , A. and Simpson , J. R. 1986 . Interactions of surface drying and subsurface nutrients affecting plant-growth on acidic soil profiles from an old pasture . Australian Journal of Experimental Agriculture , 26 : 681 – 689 .
- Radin , J. W. and Mathews , M. A. 1989 . Water transport properties of cortical cells in roots of nitrogen and phosphorus deficient cotton seedlings . Plant Physiology , 89 : 264 – 268 .
- Rama Rao , N. 1986 . Potassium nutrition of pearl millet subjected to moisture stress . Journal of Potassium Research , 2 : 1 – 12 .
- Rashid , A. and Ryan , J. 2004 . Micronutrient constraints to crop production in soils with mediterranean type characteristics: A review . Journal of Plant Nutrition , 27 : 959 – 975 .
- Rasnick , M. 1970 . Effect of mannitol and polyethylene glycol on phosphorus uptake by maize plants . Annals of Botany , 34 : 497 – 502 .
- Raun , W. R. and Johnson , G. V. 1999 . Improving nitrogen use efficiency for cereal production . Agronomy Journal , 91 : 357 – 363 .
- Richmond , K. E. and Sussman , M. 2003 . Got silicon? The non-essential beneficial plant nutrient . Current Opinion in Plant Biology , 6 : 268 – 272 .
- Sadiqov , S. T. , Akbulut , M. and Ehmedov , V. 2002 . Role of Ca2 + in drought stress signaling in wheat seedlings . Biochemistry (Moscow) , 67 : 491 – 497 .
- Sangakkara , U. R. , Frehner , M. and Nösberger , J. 2001 . Influence of soil moisture and fertilizer potassium on the vegetative growth of mungbean (Vigna radiata L. Wilczek) and cowpea (Vigna unguiculata L. Walp) . Journal of Agronomy and Crop Science , 186 : 73 – 81 .
- Sawwan , J. , Shibli , R. A. , Swaidat , I. and Tahat , M. 2000 . Phosphorus regulates osmotic potential and growth of African violet under in vitro-induced water deficit . Journal of Plant Nutrition , 23 : 759 – 771 .
- Scherer , H. W. 2001 . Sulphur in crop production – Invited paper . European Journal of Agronomy , 14 : 81 – 111 .
- Scott , B. J. and Robson , A. D. 1990 . Changes in the content and form of magnesium in the first trifaliatelaef of subterranean clover under altered or constant root supply . Australian Journal of Agricultural Research , 41 : 511 – 519 .
- Seeley , S. 2006 . Hormonal transduction of environmental stresses . HortScience , 25 ( 11 ) : 13
- Seiffert , S. , Kaselowsky , J. , Jungk , A. and Claassen , N. 1995 . Observed and calculated potassium uptake by maize as affected by soil water content and bulk density . Agronomy Journal , 87 : 1070 – 1077 .
- Shan , L. and Chen , G. L. 1993 . The principles and practices of dry land farming on the Loess Plateau , Beijing : Chinese Academic Press .
- Sharma , C. P. 2006 . Plant micronutrients , Enfield, NH : Science Publishers .
- Sharma , K. D. , Nandwal , A. S. and Kuhad , M. S. 1996 . Potassium effects on CO2 exchange, NRA and yield of clusterbean cultivars under water stress . Journal of Potassium Research , 12 : 412 – 423 .
- Sharma , P. N. , Tripathi , A. and Bisht , S. S. 1995 . Zinc requirement for stomatal opening in cauliflower . Plant Physiology , 107 : 751 – 756 .
- Shaul , O. 2002 . Magnesium transport and function in plants: the tip of the iceberg . Biometals , 15 : 309 – 323 .
- Sinclair , T. R. and Ludlow , M. M. 1984 . Who taught plants thermodynamics? The unfulfilled potential of plant water potential . Australian Journal of Plant Physiology , 33 : 213 – 217 .
- Smith , F. W. 2002 . The phosphate uptake mechanism . Plant and Soil , 245 : 105 – 114 .
- Studer , C. 1993 . Interactive effects of N-P-K-nutrition and water stress on the development of young maize plants Ph.D. Thesis, ETHZ, Zurich, Switzerland
- Sun , J. and Wang , Y. B. 2001 . Effect of straw cover on wheat yield and soil environment in dryland field . Translation of Chinese Society of Agriculture Engineers , 17 : 53 – 55 .
- Tiwari , H. S. , Agarval , R. M. and Bhatt , R. K. 1998 . Photosynthesis, stomatal resistance and related characters as influenced by potassium under normal water supply and water stress condition in rice (Oryza sativa L.) . Indian Journal Plant Physiology , 3 : 314 – 316 .
- Tobin , A. J. 1970 . Carbonic anhydrase from parsley leaves . Journal of Biological Chemistry , 245 : 2656 – 2666 .
- Turner , L. B. 1985 . Changes in the phosphorus content of Capsicum annuum leaves during water-stress . Journal of Plant Physiology , 121 : 429 – 439 .
- Umar , S. & Moinuddin 2002 . Genotypic differences in yield and quality of groundnut as affected by potassium nutrition under erratic rainfall conditions . Journal of Plant Nutrition , 25 , 1549 1562 .
- Umar , S. and Bansal , S. K. 1997 . Effect of potassium application on water stressed groundnut . Fertilizer News , 42 : 27 – 29 .
- Umar , S. , Rama Rao , N. and Sekhon , G. S. 1993 . Differential effects of moisture stress and potassium levels on growth and K uptake in sorghum . Indian Journal of Plant Physiology , 36 : 94 – 97 .
- Urao , T. , Katagiri , T. , Mizoguchi , T. , Yamaguchishinozaki , K. , Hayashida , N. and Shinozaki , K. 1994 . Two genes that encode Ca2 +-dependent protein-kinases are induced by drought and high-salt stresses in Arabidopsis thaliana . Molecular Genetics and Genomics , 244 : 331 – 340 .
- Walworth , J. L. 1992 . Soil drying and rewetting, or freezing and thawing, affects soil solution composition . Soil Science Society of American Journals , 56 : 433 – 437 .
- Wang , C. R. , Tian , X. H. and Li , S. X. 2004 . Effects of plastic sheet-mulching on ridge for water-harvesting cultivation on WUE and yield of winter wheat . Scientia Agricultura Sinica , 37 : 208 – 214 .
- Waraich , E. A. , Ahmad , R. and Ahmad , S. S. 2008 . Water use efficiency and yield performance of wheat (Triticum aestivum L.) under different levels of irrigation and nitrogen . Caderno de Pesquisa série Biologia , 20 : 22 – 34 .
- Wei , X. P. , Wang , Q. J. and Wang , W. Y. 2000 . “ The loss of soil nutrients on Loess Plateau affected by precipitation ” . In Soil erosion and dryland farming , Edited by: Laflen , J. M. , Tian , J. and Huang , C. 176 – 184 . New York : CRC Press .
- Wright , D. 1982 . “ Crop physiology ” . In Agricultural note book , Edited by: Halley , R. J. 69 – 82 . London : Butterworth Scientific .
- Xu , F. A. and Zhao , B. Z. 2001 . Development of crop yield and water use efficiency in Fengqiu Country, China . Acta Pedologia Sinica , 38 : 491 – 497 .
- Yeo , A. R. and Flowers , T. J. 1985 . The absence of an effect of the Na + /Ca2 + ratio on sodium-chloride uptake by rice (Oryza sativa L.) . New Phytologist , 99 : 81 – 90 .
- Yu , Q. , Osborne , L. and Rengel , Z. 1998 . Micronutrient deficiency changes activities of superoxide dismutase and ascorbate peroxidase in tobacco plants . Journal of Plant Nutrition , 21 : 1427 – 1437 .
- Zhang , X. Y. , Chen , S. Y. and Liu , M. Y. 2002 . Evapotranspiration, yield and crop coefficient of irrigated maize under straw mulch conditions . Progress in Geography , 21 : 583 – 592 .
- Zhang , Z. H. and Cai , H. J. 2001 . Effects of regulated deficit irrigation on plastic-mulched cotton . North West Agriculture and Forestry University , 29 : 9 – 12 .
- Zhao , D. and Oosterhuis , D. M. 2002 . Cotton carbon exchange, non-structural carbohydrates, and boron distribution in tissues during development of boron deficiency . Field Crops Research , 78 : 75 – 87 .
- Zhou , W. B. 2003 . Review on the study of water resources utilization efficiency in irrigation district in arid and semiarid areas of China . Journal of Arid Land Research and Environment , 17 : 91 – 95 .
- Zhu , Z. , Liang , Y. C. and Sun , W. 2004 . Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.) . Plant and Science , 167 : 527 – 533 .