Abstract
Recent years have witnessed a rapid growth in organic farming in Lithuania. We aimed to compare the occurrence of Brevicoryne brassicae L., its primary parasitoid Diaeretiella rapae (McIntosh) and predators from Coccinellidae and Cecidomyiidae families in non-fertilized, organically (with cow manure) and synthetically fertilized white cabbage crops. Occurrence of B. brassicae was higher on organically fertilized plants. Parasitoid D. rapae had the strongest effect in reducing the aphid population, as other predators occurred only in small densities. Quantity of parasitized aphids by D. rapae was higher on manure fertilized plants. In non-fertilized cabbage plants seasonal parasitization per cent of B. brassicae by D. rapae increased along with the expected lower aphid pressure compared with fertilized plants and reached on average 28.5%. D. rapae reduced the population of cabbage aphid by on average 15.5% in manure fertilized and −12.9% in synthetically fertilized plants. The regression showed a positive relationship between the increase of the number of B. brassicae and activity of D. rapae. No coccinellids were found in aphid colonies on synthetically fertilized plants. Different fertilization did not have any significant effect on the incidence of Aphidoletes aphidimyza Rond. In conclusion, organic fertilization increased the population of cabbage aphid more than its natural enemies and the abundance of natural enemies was too small to provide effective reduction of this pest.
Introduction
The cabbage aphid, Brevicoryne brassicae L. (Aphidae), is a colonial aphid infesting crucifer plants in temperate areas all over the world and causes serious losses of yield in Brassica crops and reduces their marketable value (Liu et al., Citation1994; Costello & Altieri, Citation1995; Shaltiel & Ayal, Citation1998). B. brassicae prefers feeding not only on younger plant tissues (Costello & Altieri, Citation1995) but also on young leaves, which makes the pest very harmful to white cabbage. Cruciferous plants contain glucosinolates as cabbage aphid main attractant (Ellis, Citation1984; Zabża, Citation1989). The Braconid wasp Diaeretiella rapae is an important primary parasitoid of a wide range of aphids including B. brassicae (Hagvar & Hofsvang, Citation1991; Elliot et al., Citation1994; Pike et al., Citation1999; Jankowska & Wiech, Citation2003). In some cases the natural enemies are influenced by the host plant, crop cultural practices and environmental conditions (Tamaki et al., Citation1981). To localize their prey in natural habitats, entomophagous beneficials use numerous chemical cues emitted by prey and host plants, each alone or in association (Vet & Dicke, Citation1992). Furthermore, parasitoids and prey prefer the same host plant, possibly because aphids and D. rapae positively respond to the volatile compounds produced by the plants (Bundemberg, Citation1990), and honeydew, that is emitted by aphids and used by D. rapae as a kairomone (Shaltiel & Ayal, Citation1998). Lopez et al. (Citation1990) found that scattered B. brassicae are more likely to be attacked by D. rapae than B. brassicae in colonies. Higher abundance and activity of parasitoids of cabbage aphid D. rapae in ecologically grown cabbage crops has been observed since 2003 in the experimental fields of the Institute of Horticulture (Duchovskiene & Raudonis, Citation2008).
Coccinellidae are the best known predators of aphids (Gadomski et al., Citation1998). Another predator, Cecidomyiid – Aphidoletes aphidimyza Rond., plays an important role in reducing the number of cabbage aphids (Jankowska, Citation2005). The current control of aphids in Brassica crop fields is based mainly on insecticides (Ellis et al., Citation1996; Nunnenmacher & Goldbach, Citation1996). A potential consequence within integrated crop management systems is that aphicides take out susceptible genotypes and thereby reduce the population sufficiently to allow biological control of the resistant individuals by surviving natural enemies (Umoru & Powell, Citation2002). Also the use of insecticides on vegetable crops has caused increasing concern among growers, markets and consumers (Ellis et al., Citation1996). Because of the restrictions imposed on organic cabbage production and the limited availability of the biopesticides, it is difficult to make this production system economically reasonable in Lithuania (Starkute, Citation2008). As a result, it is relevant to estimate the input of natural enemies and possibilities of bioagent use.
The present study was designed to examine the effect of different cabbage fertilizing systems on the occurrence of Brevicoryne brassicae L., its parasitoid Diaeretiella rapae (McIntosh), predators from Coccinellidae and Cecidomyiidae families, and the relationship between cabbage aphid and parasitoid.
Materials and methods
We carried out experiments in the experimental fields of the Institute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry (Central Lithuania 55° 60′ N 23° 48′ E) in 2004–2005. The soil of the experimental site is Epicalcari-endohypogleyic Cambisol with a texture of sandy loam on light loam and a humus content of 1.67%. Cabbage seedlings of cv. BielorusiŠka Dotnuvos were planted 70×50 cm, each replicate covered an area of 12 m2, and the treatment was replicated four times at a systemic plot distribution. No pesticides were used in all treatments and weeds were removed mechanically.
The trial included the following treatments:
| 1. | Control – non-fertilized. | ||||
| 2. | Fertilized with manure from cows. Cow manure at a rate of 60 t ha−1 was applied before planting, as in the organic growing technology. | ||||
| 3. | Fertilized with synthetic fertilizers. A separate block (11 m from the manure-fertilized treatment, according to Rutkoviene et al., Citation2002) contained a treatment applied with synthetic fertilizers before cabbage planting (N – 200 kg ha−1, P – 120 kg ha−1, K – 120 kg ha−1), as in the conventional growing technology. | ||||
Observations started when the first winged forms of B. brassicae appeared and were continued every 4–5 days. Ten plants per plot were inspected. Winged and non-winged forms of aphids, mummies of parasitized aphids, larvae of lady beetles and cecidomyiids were counted. Parasitized aphids were left on vegetables to allow the emergence of the next generations of the parasitoid. Caterpillars of chewing pests were collected and discarded. The identification of Diaeretiella rapae adults was based on the identification key by Kopaneva (1982). The identification of Coccinelidae adults was based on the identification key by Pileckis (Citation1976). The Cecidomyiidae larvae were collected, reared and identified using the key by Mamaev (1969).
Meteorological data (air temperature and amount of precipitation) were recorded using Metos D equipment ().
Table I. Meteorological conditions of the experimental years and long-term averages for the 1924–2000 period.
Parasitization was determined as: parasitization (%) = parasitized aphids×100/(parasitized aphids +non-parasitized aphids). The number of aphids, their parasitoids and predators were compared among treatments in this study by a single factor analysis of variance (ANOVA). Specific differences were identified with Duncan's multiple range test (p=0.05). The relationships between the number of cabbage aphids and intensity of D. rapae parasitization were analysed using the correlation analysis from the EXCEL program package.
Results
The experimental years differed in the weather conditions (). In 2004, lower temperatures in June and especially July, which is the main aphid development period, adversely affected cabbage aphid development, which resulted in a lower population of B. brassicae compared with that in 2005. Rainfall during August was significantly higher than the long-term average in both years. August was colder in 2005 than in 2004 and the long-term average. This negatively affected not only aphids but also their parasitoid development. However, the abundance of aphids parasitized by D. rapae decreased in the second 10-day period of August in relation to the decrease in aphid abundance in both years. Temperature effect on the abundance of D. rapae is not clear.
In all treatments, cabbage plants were colonized by B. brassicae at the same time, but the number of aphids on plants differed (). In 2004–2005, the highest mean number of aphids was observed in cabbage treatments fertilized with manure (). Results from 2004 show that aphid population was low (maximum B. brassicae per plant = 21) and there were no significant differences between the organic and conventional treatments, but the abundance of aphids in manure-fertilized plots was 2.4 and 2.1 times higher than in non-fertilized and synthetically fertilized plots, respectively. In 2005, the abundance of aphids in manure-fertilized plots was 4.1 and 3.2 times higher than in non-fertilized and synthetically fertilized plots.
Figure 1. Dynamic of cabbage aphids and their parasitization by Diaeretiella rapae on non-fertilized (A, B), manure-fertilized (C, D) and synthetically fertilized (E, F) cabbage plants.
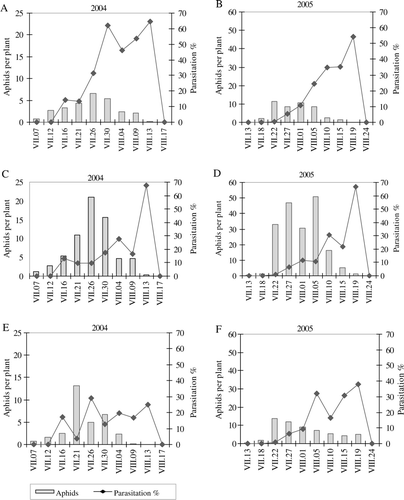
Table II. The occurrence of B. brassicae and D. rapae on differently fertilized white cabbage in 2004–2005.
During the experimental period, D. rapae occurred each year with some delay subject to aphid emergence (). First mummies of aphids were found in mid July in 2004 and at the end of July in 2005. At the beginning of occurrence, the number of parasitized aphids was low (averaged over all treatments −0.4 in 2004 and in 2005 −0.1 parasitized aphids per plant). Higher parasitization was observed during the periods with lower aphid abundance. When the weather conditions were favourable for rapid growth of aphid population, despite the growth of parasitoid population, the degree of parasitization decreased. The degree of parasitization varied from plot to plot. The highest parasitization of aphids was observed at the beginning of August 2004, in non-fertilized and manure-fertilized plots it was 64.6 and 67.5%, respectively (A, 1C); and in mid August 2005 it was 54.2 and 66.6%, respectively (B, 1D). The parasitization was lower in the plots fertilized with synthetic fertilizers; at the end of aphids’ occurrence on plants it made up 25% (E) and 38% (F), respectively. In 2004, the counts of parasitized aphids in manure-fertilized plots were 1.2 and 2.4 times higher than in non-fertilized and synthetically fertilized plots, respectively (). In 2005, the number of parasitized aphids in manure-fertilized plots was 1.8 and 1.6 times higher than in non-fertilized and synthetically fertilized plots, respectively. D. rapae reduced the population of cabbage aphid by on average 28.5% in non-fertilized plants. The percentage of parasitized aphids was on average 13.0% and 15.6% lower respectively in manure fertilized and synthetically fertilized than in non-fertilized plants.
The occurrence of the Coccinellidae in aphid colonies is presented in . During our research, no coccinellids were found in aphid colonies on cabbage plants treated with synthetic fertilizers. During the experimental period, we collected lady beetle adults of only one Coccinellidae species – sevenspotted lady beetle (Coccinella septempunctata L.).
Table III. The abundance of Coccinella septempunctata L. and Aphidoletes aphidimyza Rond. in the cabbage aphid (Brevicoryne brassicae L.) colonies in 2004–2005 (at the time of occurrence on plants).
The larvae of A. aphidimyza were recorded on cabbage plants only in the 2005 growing season (). Comparison of the incidence of Aphidoletes aphidimyza Rond. on differently fertilized cabbage crops in 2005 did not reveal any significant differences between its quantities. We did not calculate any correlations and regression for these two predators, because they occurred on single plants and for a short time.
To assess the relationship between the number of cabbage aphids and activity of D. rapae, we calculated the correlation and regression coefficients (). Statistically significant correlation was found for all treatments in our study. The correlation between the mean number of parasitized aphids and the mean number of aphids was the strongest in 2004 when the infestation of aphids was lower. The regression showed a positive relationship between the increase in aphid number and D. rapae parasitization. The increase of the number of parasitized aphids subject to the increase in the number of aphids was the highest for non-fertilized cabbage.
Table IV. The relationship between the number of B. brassicae and D. rapae on cabbage as affected by fertilization.
Discussion
Air temperature, leaf temperature and soluble nitrogen are important factors influencing aphid growth and reproduction (Burgess et al., Citation1996). Optimal nitrogen levels are important for plant growth and development (Muzika & Pregizer, 1992). We can suggest that in our study nitrogen supply was higher in fertilized treatments, because plants were bigger, than in non-fertilized. High nitrogen fertility can increase insect populations (Muzika & Pregizer, 1992). According to Staley et al. (Citation2010), nitrogen concentration was greater for conventionally fertilized than organically fertilized B. oleracea. Glucosinolate concentrations were up to three times higher in Brassica plants grown in the organic treatments and the Brassica specialist B. brassicae was more abundant on organically fertilized plants, which concurs with our study. Costello and Altieri (Citation1995) did not find any significant differences between fertilizer (compost or synthetic fertilizer) effects on B. brassicae and D. rapae population growth. Predators and parasitoids in cabbage fields were also important for the control of cabbage aphid, but their role was also low in early season, and the natural population of parasitoids could not control the aphid population (Nunnenmacher & Goldbach, Citation1996; Jankowska & Wiech, Citation2003). The natural population of parasitoids and predators was low at the beginning of aphid occurrence in our research. According to Zhang and Hassan (2003), one of the ways to increase parasitization at this time is the transplantation of cabbage plants that include both D. rapae mummies at different development stages and unparasitized aphids providing a continuous supply of parasitoids for about 2 weeks (Zhang & Hassan, 2003). According to some authors, D. rapae is attracted by allyl isothiocyanate (Zabża, Citation1989; Titayavan & Altieri, Citation1990) released by damaged plants. This might suggest that fertilized cabbage plants (especially manure-fertilized ones) were damaged more severely or contained more of this substance than non-fertilized, and this caused an increase in the activity and abundance of the D. rapae. According to Shaltiel and Ayal (Citation1998) the wasp D. rapae uses honeydew emitted by its host cabbage aphid as a kairomone. The findings of these authors indicate that honeydew level is used by D. rapae as a cue for assessment of the number of aphids in the colony. In our study, higher aphid population and more parasitized aphids were found in manure-fertilized treatments. This could suggest that more honeydew was produced in manure-fertilized plots with a higher aphid population. It is likely that the amount of honeydew increased not as rapidly as the abundance of aphids, therefore the number of parasitized aphids was lower in non-fertilized and only a little higher in fertilized treatments in 2005 compared with 2004 (). According to Zhang and Hassan (2003) the parasitoid D. rapae at a wasp aphid ratio 1.2:1 resulted in successful control of the aphids by the second generation of the parasitoid. This might suggest that in 2004 the highest parasitization of aphids was the result of second generation of D. rapae parasitization when the number of aphids was the lowest and rated at 1.2:1. Zhang and Hassan (2003) found that parasitization by D. rapae reduced the number of offspring of the cabbage aphid and shortened its reproduction period. In 2004–2005, D. rapae reduced the population of cabbage aphid by 12.4–28.5% (). The results obtained demonstrate the significant role D. rapae plays in limiting the aphid population. However, the effectiveness of parasitoids as biological control agents can be constrained by insecticides being used to control the same or other target pests in the crop, not only through direct mortality but also as a result of sublethal effects on biological attributes such as fecundity, longevity, egg viability and sex ratio (Habtu & Samsoe-Petersen, 1987). Thus, when applying insecticides against aphids one should consider their influence on a parasitoid population.
The impact of lady beetles on reducing the number of cabbage aphid may be significant, if they feed on some cruciferous plants, like oilseed rape crop, oilseed rape self-sown and white mustard (Gadomski et al., Citation1998). Lady beetles appeared at the onset of aphid occurrence, but twice in 2004 and three times in 2005 they were found only sporadically. This agrees with the findings of Jankowska (Citation2005) that cabbage aphids on cabbage plants as food are of little interest for lady beetles. Females of A. aphidimyza start laying eggs on leaves where there is enough food for larval development. The larvae of this predatory midge often occurred only on single plants with abundant colonies of aphid (Jankowska, Citation2005). It also confirmed the fact that although in 2005 aphid midge larvae were not observed before August, when aphid colonies markedly increased, in 2004 A. aphidimyza larvae were not found, because the aphid colonies were too small. A. aphidimyza is successfully used in biological control of aphids in greenhouses on a commercial scale, and it presents a good perspective for biological aphid control in the field, too (Jankowska, Citation2005).
In our research, the complex of natural enemies of cabbage aphid on organically fertilized cabbage was more abundant compared with synthetically fertilized plants. However, the population of natural enemies was too small to provide effective reduction of this pest. To get a more consistent view, this issue needs to be researched further. Brevicoryne brassicae L. was more abundant on organically fertilized cabbage plants. Quantity of parasitized aphids by primary parasitoid Diaeretiella rapae (McIntosh) was higher on manure fertilized plants. In non-fertilized cabbage plants seasonal parasitization per cent of B. brassicae by D. rapae increased along with the expected lower aphid pressure compared with fertilized plants and reached on average 28.5%. D. rapae reduced the population of cabbage aphid by on average 15.5% in manure fertilized and −12.9% in synthetically fertilized plants. Parasitoid D. rapae had the strongest effect reducing the aphid population, as other predators occurred only in small densities. The regression showed a positive relationship between the increase of the number of B. brassicae and activity of D. rapae in all treatments. No coccinellids were found in aphid colonies on synthetically fertilized plants. Different fertilization did not have any significant effect on the incidence of Aphidoletes aphidimyza Rond.
References
- Ayal , Y. 1987 . The foraging strategy of Diaeretiella rapae. I. (The concept of the elementary unit of foraging . Journal of Animal Ecology , 56 : 1057 – 1068 .
- Bundemberg , W. J. 1990 . Honeydew as a contact kairomone for aphid parasitoids . Entomologia Experimentalis et Applicata , 55 : 139 – 148 .
- Burgess , A. J. , Warington , S. and Allen-Williams , L. 1996 . Cabbage aphid (Brevicoryne brassicae. L.) ‘performance’ on oilseed rape (Brassica napus. L.) experiencing water deficits: roles of temperature and food quality. Brassica symposium – IX Crucifer Genetics Workshop . Acta Horticulture , 407 : 245 – 251 .
- Costello , M. J. and Altieri , M. A. 1995 . Abundance, growth rate and parasitism of Brevicoryne brassicae and Myzus persicae (Homoptera: Aphididae) on broccoli grown in living mulches . Agriculture, Ecosystems and Environment , 52 : 187 – 196 .
- Duchovskienė , L. and Raudonis , L. 2008 . Seasonal abundance of Brevicoryne brassicae L. and Diaeretiella rapae (M'Intosh) under different cabbage growing systems . Ekologija , 54 : 260 – 264 .
- Ellis , P. R. 1984 . Can factors which influence the relationship between Brevicoryne brassicae and its host plants be exploited in integrated pest management? International Control in Field Vegetable Crops . IOBS/WPRS Bulletin , 17 ( 8 ) : 90 – 95 .
- Ellis , P. R. , Tatchell , G. M. , Collier , R. H. and Parker , W. E. 1996 . Assessment of several components that could be used in an integrated programme for controlling aphids on field crops of lettuce . IOBC/WPRS Bulletin , 19 : 91 – 97 .
- Elliot , N. C. , Reed , D. K. , French , B. W. and Kindler , S. D. 1994 . Aphid host effects on the biology of Diaeretiella rapae . Southwestern Entomologist , 19 : 279 – 283 .
- Gadomski , H. , Kelm , M. and Klukowski , Z. 1998 . “ Predatory insects in colonies of Brevicoryne brassicae (L.) on different cruciferous plants ” . In Aphids and other homopterous insects , Edited by: Cichocka , E. , Goszczynski , W. and Wiech , K. 97 – 106 . Skierniewice, , Poland : Graf-Sad .
- Habtu , F. F. and Samsøe-Petersen , L. 1987 . Semi-field method for testing side effects of pesticides on adults of the parasitic wasp Encarsia formosa (Gahan) (Hym., Aphelenidae) . Journal of Applied Entomology , 104 : 473 – 479 .
- Hagvar , E. B. and Hofsvang , T. 1991 . Aphid parasitoids (Hymenoptera, Aphidiidae): biology, host selection and use in biological control . Biological News and Information , 12 : 13 – 41 .
- Jankowska , B. 2005 . The effect of cruciferous vegetables on occurrence of Cecidomyiidae, Coccinellidae and Chrysopidae predators in colonies of cabbage aphid (Brevicoryne brassicae L.) on different cabbage vegetables . Journal of Plant Protection Research , 45 : 301 – 308 .
- Jankowska , B. and Wiech , K. 2003 . Occurrence of Diaretiella rapae (M'Intosh) (Aphidiidae) in cabbage aphid (Brevicoryne brassicae L.) colonies on different crucifer crops . Sodininkystė ir daržininkystė , 22 : 155 – 164 .
- Kopaneva , L. M. 1982 . Key to harmful and useful insects and mites of vegetables cultures and potato in the USSR (In Russian) . Leningrad : Kolos .
- Liu , S. S. , Hommes , M. and Hildenhagen , R. 1994 . Damage to white cabbage by the aphid Brevicoryne brassicae (L.): influence of aphid density and stage of plant growth . IOBC/WPRS Bulletin , 17 : 75 – 89 .
- Lopez , E. R. , van Driesche , R. G. and Elkinton , J. S. 1990 . Rates of parasitism by Diaeretiella rapae (Hymenoptera: Braconidae) for cabbage aphids (Homoptera: Aphididae) in and outside colonies: why do they differ? . Journal of the Kansas Entomological Society , 63 : 158 – 165 .
- Mamaev , B. M. 1969 . Cecidomyiidae (Itonididae) . In G. Bei–Bienko , Ya . (eds.), Keys to the insects of the European part of the USSR Volume 5, Part 1: Diptera & Siphonaptera . Nauka Publishing House , Moscow, , Russia . pp. 356 – 420 . (In Russian) .
- Muzika , R. M. and Pregitzer , K. S. 1992 . Effect of nitrogen fertilization on leaf phenolic production of grand fir seedlings . Journal of Experimental Botany , 54 : 1931 – 1939 .
- Nunnenmacher , L. and Goldbach , H. E. 1996 . Aphids on lettuce: the effect of excluding aphid predators . IOBC/WPRS Bulletin , 19 : 38 – 47 .
- Pike , K. S. , Stary , P. , Miller , T. , Allison , D. , Graf , G. , Boydston , L. , Miller , R. and Gillespie , R. 1999 . Host range and habitats of the aphid parasitoid Diaeretiella rapae (Hymenoptera: Aphididae) in Washington State . Environmental Entomology , 28 : 61 – 71 .
- Pileckis , S. 1976 . Lietuvos vabalai , p. 124 . In Lithuanian . Vilnius, , Lithuania : Mokslas .
- Rutkoviene , V. , Bakutis , B. , Rutkovaite , I. , Slavinskiene , V. , Šaluchaite , A. , & Zemeckis , R. , 2002 Rules of ecological agriculture: production, processing, realization, marking . p. 63 . (In Lithuanian) . Vilnius, , Lithuania , Ekoagros .
- Shaltiel , L. and Ayal , Y. 1998 . The use of kairomones for foraging decisions by an aphid parasitoid in small host aggregations . Ecological Entomology , 23 : 319 – 329 .
- Staley , J. T. , Stewart-Jones , A. , Pope , T. W. , Wright , D. J. , Leather , S. R. , Hadley , P. , Rossiter , J. T. , van Emden , H. F. and Poppy , G. M. 2010 . Varying responses of insect herbivores to altered plant chemistry under organic and conventional treatments . Proceedings of the Royal Society , 277 : 779 – 786 .
- Starkutė , R. 2008 . Ecological vegetable growing investigations at the Lithuanian Institute of Horticulture . Sodininkystė ir daržininkyste 27 , 199 – 204 . (In Lithuanian) .
- Tamaki , G. , Annis , B. and Weiss , M. 1981 . Response of natural enemies to the green peach aphid in different plant cultures . Environmental Entomology , 10 : 375 – 378 .
- Titayavan , M. and Altieri , M. A. 1990 . Synomone-mediated interactions between the parasitoid Diaretiella rapae and Brevicoryne brassicae under field conditions . Entomophaga , 35 : 499 – 507 .
- Umoru , P. A. and Powell , W. 2002 . Sub-lethal effect of the insecticides Pirimicarb and Dimethoate on the aphid parasitoid Diaretiella rapae (Hymenoptera: Braconidae) when attacking and developing in insecticide-resistant hosts . Biocontrol Science and Technology , 12 : 605 – 614 .
- Vet , L. E. M. and Dicke , M. 1992 . Ecology of infochemical use by natural enemies in a tritrophic context . Annual Review of Entomology , 37 : 141 – 172 .
- Zabza , A. 1989 . Chemiczne podstawy oddzialywania roslina-owad . Kosmos , 36 : 155 – 177 .
- Zhang , W. Q. and Hassan , S. A. 2003 . Use of the parasitoid Diaretiella rapae (McIntosh) to control the cabbage aphid Brevicoryne brassicae (L.) . Journal of Applied Entomology , 127 : 522 – 526 .