Abstract
Asystasia gangetica and Pennisetum polystachion are the most troublesome weeds in oil palm plantations in Malaysia and establishment of cover crops under oil palms is an integral part of sustainable weed management in plantations. Replacement-series experiments were conducted to evaluate the mode and strength of competitiveness of four legume cover crop species, Calopogonium caeruleum, Centrosema pubescens, Mucuna bracteata and Pueraria javanica, and one soft grass species, Axonopus compressus, against Asystasia gangetica and Pennisetum polystachion. Relative yield, relative yield total and relative crowding coefficient were calculated with reference to shoot biomass. Asystasia gangetica was a consistently weaker competitor than Pennisetum polystachion. The legume cover crop species had higher relative yield when grown in mixtures, whereas the yield of Asystasia gangetica was lower in mixtures with legumes than in monoculture. The higher competitiveness of legumes was due to greater canopy height and leaf area index. Centrosema pubescens was a slightly weaker competitor than the other legumes. Although leaf area per plant of Centrosema pubescens did not differ from that of other three legumes, canopy height and blocking photosynthetically active radiation to the soil surface were lower. When grown with a cover crop for 12 weeks, dry weight per plant, leaf area and shoot number of Pennisetum polystachion increased as its proportion decreased in mixtures. Relative yield data indicated that Pennisetum polystachion is a strong competitor. Axonopus compressus was less susceptible to Pennisetum polystachion interference than the legumes. All cover crop species studied proved highly competitive against Asystasia gangetica, but none could compete against Pennisetum polystachion. The study confirms the feasibility of using a cover crop for management of Asystasia gangetica.
Introduction
Oil palm plantations cover more than 12 million ha worldwide, of which 4.69 million ha are located in Malaysia (MADI Citation2009/2010). The warm tropical climate and plentiful rainfall in Malaysia allow luxuriant growth of both crops and weeds. Pennisetum polystachion and Asystasia gangetica are frequently found weeds in oil palm plantations (Wibawa et al. Citation2009). Pennisetum polystachion is an invasive annual grass species (Baki Citation2004), while A. gangetica is a shade-tolerant broadleaved weed that can spread very rapidly in most Malaysian plantations and smallholdings. Weeds compete with oil palm for nutrients, particularly nitrogen (N) and potassium (K). When poorly managed, weeds can cause losses of between 25% and 100% of yield (Collins Citation1991). In one case, eradication of very dense stands of Asystasia (especially A. gangetica) in an oil palm plantation resulted in a 12% increase in fresh fruit bunch production (Kustyanti and Horne Citation1991). Weed control is an important and expensive component of plantation crop management that can account for 17–27% of the total upkeep costs for oil palm (Wibawa et al. Citation2010).
Cover crops are frequently planted in oil palm, rubber and coconut plantations in tropical Asia (Aldaba Citation1995). Cover crops check soil erosion, soil nutrient and organic matter losses (Lal et al. Citation1991), suppress weed growth and reduce soil moisture evaporation (Turner and Gillbanks Citation2003). In addition, legume cover crops can fix atmospheric N, recycle nutrients and improve soil nutrient availability (Lehmann et al. Citation2000). Establishment of cover crops in oil palm and rubber plantations is an integral part of plantation management in Malaysia. Cover crops such as Centrosema pubescens, Calopogonium caeruleum, Calopogonium muconoides and Pueraria phaseoloides have been widely tested and their effects on soil fertility and yield of major crops are well documented (Agamuthu and Broughton Citation1985). However, these traditional leguminous cover crops cannot compete successfully against growth of volunteer weeds, particularly Mikania, Asystasia and various grasses, which are usually controlled by hand weeding or herbicide application during the first year of plantation establishment (Kobayashi et al. Citation2003). Mucuna bracteata, which was first introduced into oil palm plantations in 1991, can control weeds effectively. The fast-growing nature of M. bracteata enables it to compete successfully with most weeds in oil palm plantations, but some competitive grasses and shade-tolerant weeds can co-exist with it (Goh and Chiu Citation2007). Axonopus compressus has the potential to be used as a cover crop to suppress weeds, especially troublesome broadleaved weeds, which generally comprise around 83% of the weed community (Mohamad et al. Citation2010). Senarathne and Perera (Citation2011) showed that application of glyphosate followed by the establishment of leguminous creeping cover crops was the most effective method of controlling P. polystachion.
Replacement-series designs are routinely used to characterize the competitive interactions of species in mixed stands, particularly in crop-weed interference studies (Fischer et al. Citation2000, Cralle et al. Citation2003). A replacement-series design includes pure stands and mixtures in which the proportions of the two species studied are varied, while the total plant density remains constant over all treatments (De Wit Citation1960). The shape of the curves in replacement-series diagrams is then used to interpret the nature of the interaction between the species (Cousens Citation1991, Radosevich et al. Citation2007). Relative yield (RY) is a commonly used variable that expresses the yield of a species in the mixture as a proportion of its yield in monoculture and thus measures interspecific and intraspecific competition (Hector Citation2006). Intraspecific competition occurs between individuals of the same species and can be expected in monoculture, whereas interspecific competition involves individuals of two or more species (Hector Citation2006). An increase in the competitiveness of a given species is reflected as an increase in RY value, resulting in a convex curve for the more competitive species, while the less competitive species produces a concave curve, indicating lower or slower rate of accumulation of that resource (Radosevich et al. Citation2007). This method is useful in evaluating relative growth in monoculture and interference effects between two species (or plant types) at a given total density, and in assessing interference between species in mixtures (Radosevich Citation1987). It could thus provide valuable insights into the dynamics of interference between cover crops and weeds in mixtures.
The present study sought to evaluate the interference dynamics between five cover crop species, Axonopus compressus, Calopogonium caeruleum, Centrosema pubescens, Mucuna bracteata and Pueraria javanica, and the weed species Asystasia gangetica and Pennisetum polystachion. The aim was to provide data support in selection of cover crop species for outcompeting weeds in oil palm plantations.
Materials and methods
Experimental site
Replacement-series competition experiments were performed in an open area at the Plant House of Universiti Putra Malaysia (UPM), Malaysia (3° 02′ N, 101° 42′ E; elevation 31 m a.s.l.) during January to April 2011. The climate at the experimental site is hot, humid and tropical, with high rainfall. During the experimental period, mean monthly maximum and minimum temperature were 33.75 and 23.2 °C, respectively, and relative humidity ranged from 93.4 to 96%, while sunshine hours ranged from 6.31 to 7.06 hours day−1. Growth medium for the experiment was prepared by mixing topsoil, sand and peat moss in proportions 8:2:1 (v/v).
Experimental treatments and design
The five cover crop (C) species (Axonopus compressus, Calopogonium caeruleum, Centrosema pubescens, Mucuna bracteata and Pueraria javanica) were combined with the two weed (W) species (Asystasia gangetica and Pennisetum polystachion) in the following replacement-series: C100:W0;C75:W25;C50:W50;C25:W75; and C0:W100. The treatments were arranged in a randomized complete design, with four replicates.
Crop husbandry
Polythene growbags (poly bags) measuring 30 cm×20 cm×25 cm were used for growing the cover crops. Eight plants were accommodated in each bag to maintain a density of 72 plants m−2 and the bags were arranged in rows with 15 cm inter-row spacing and 10 cm intra-row spacing. Mucuna bracteata seeds were clipped and other legume seeds were scarified with sandpaper to improve permeability to water and then treated with 0.2% Benomyl solution. Treated seeds were pre-germinated on filter paper for 3 days in the laboratory and then planted in the poly bags. Axonopus compressus cuttings were directly planted in the bags. At the same time, seeds of the weed species A. gangetica and P. polystachion were planted directly in the bags according to the spatial arrangement for the density, followed by thinning a week after emergence. After planting, each poly bag was treated with 0.2% Benomyl again. Cover crops were allowed to compete with the weeds for 12 weeks. Plants were watered daily and fertilized with 5 g NPK green® (N:P:K 15:15:15)/poly bag every 4 weeks. Weeds other than A. gangetica and P. polystachion were removed regularly during the experimental period.
Data collection
Ground cover by the canopy of cover crops was visually estimated every week. Before harvesting, photosynthetically active radiation (PAR) was measured using an illuminometer (Extech instruments, Model 407026) on the soil surface and above the canopy. The percentage of PAR penetrating the canopy was then calculated. Plant height from the soil surface to the top of the cover crop canopy and number of shoots on P. polystachion were measured. Leaves of cover crops and weeds were harvested from each individual poly bag, and the leaf area of each species was determined using an area meter (LI-3100, USA). Stems and leaves of cover crops and weeds were oven-dried at 70 °C for 72 hours to obtain shoot biomass and then converted to relative yield according to the following equations:
Regression of RYs for the different weed proportions 0, 0.25, 0.5, 0.75 and 1.0 were used to produce the replacement-series diagrams and determine the competitiveness in mixtures compared with monoculture (De Wit Citation1960, Cousens Citation1991, Weigelt and Jolliffe Citation2003). The shape of the RY replacement curve for shoot dry weight relative to expected yield was used as an indicator of the extent of interference between the two competing species (Harper Citation1977). Relative yield total (RYT) predicts the competition between the two test species for the same resources (Harper Citation1977) or is a measure of the performance of each under conditions of intra- and interspecific interactions relative to intraspecific interactions alone. Equivalent yield ratios (EYR) or the proportions at which both species growing in the mixture produce the same yield, was calculated for each mixture (Santos et al. Citation1997). RYT was calculated according to the equation (Harper Citation1977):
Aggressiveness of one species to another in the mixture was determined by the relative crowding coefficient (RCC), which serves as an index of competition when two species are mixed in equal proportions (Jolliffe et al. Citation1984). RCC was determined using the equation:
where W1m and W2m are shoot dry weight of crop species and weed species, respectively at C50:W50 mixture and W1p and W2p are shoot dry weight of crop species and weed species in pure culture (monoculture), respectively per poly bag.
Statistical analysis
All data were subjected to ANOVA using SAS statistical software package version 9.2 (SAS Citation2003) and values were further differentiated by Tukey test at p<0.05. All regressions were conducted using Sigma Plot version 11.
Results
Competition between cover crops and Asystasia gangetica
The de Wit competitiveness diagrams of relative shoot dry weight (RY) of cover crops and A. gangetica are presented in . The RY of A. compressus, C. caeruleum, C. pubescens and P. javanica (A, B, C, E) increased in a quadratic manner as their proportion in mixtures with A. gangetica increased, resulting in a convex curve. The response of M. bracteata was linear (D). The RY of A. gangetica was less than the pure stand level and produced a concave curve. The EYR in all diagrams was shifted away from 0.50, with the value for A. compressus, C. caeruleum, C. pubescens, M. bracteata and P. javanica being 0.68, 0.64, 0.62, 0.66 and 0.69, respectively. The RYT of M. bracteata was less than 1, that of C. caeruleum was more than 1 and that of A. compressus, C. pubescens and P. javanica was very close to 1 (). The RCC of all cover crops when grown in equal proportions with the weeds (C50:W50) was greater than that of A. gangetica on them (). The RCC of M. bracteata and P. javanica over A. gangetica was greater than that of the other cover crops. As regards the RCC of A. gangetica on cover crops, the highest values were recorded for C. pubescens ().
Figure 1. Interaction between cover crops and Asystasia gangetica in replacement-series experiments at 12 weeks after planting. (A) Axonopus compressus response: Y = −0.66X2−0.66X+0.91, R2 = 0.86; Asystasia gangetica response: Y = 0.71X2+0.18X+0.02, R2 = 0.88; (B) Calopogonium caeruleum response: Y = −0.94X2+0.02X+0.96, R2 = 0.97; Asystasia gangetica response: Y = 0.91X−0.004, R2 = 0.95; (C) Centrosema pubescens response: Y = −0.56X2−0.42X+0.98, R2 = 0.99; Asystasia gangetica response: Y = 0.37X2+0.61X−0.007, R2 = 0.98; (D) Mucuna bracteata response: Y = −1.03X+1.04, R2 = 0.99; Asystasia gangetica response: Y = 1.15X2−0.25X+0.03, R2 = 0.94; (E) Pueraria javanica response: Y = −0.72X2−0.15X+0.94, R2 = 0.92; Asystasia gangetica response: Y = 0.90X2+0.04X+0.02, R2 = 0.98.
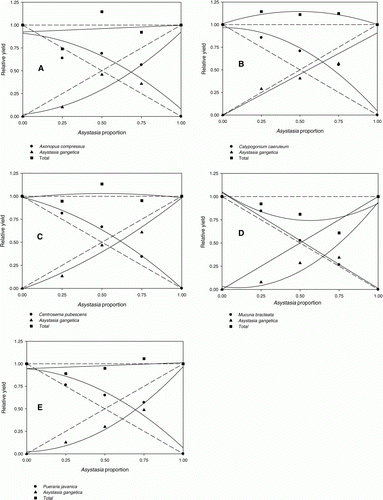
Table I. Relative crowding coefficient in interactions between different cover crop species and Asystasia gangetica and time of 80% coverage by cover crops.
Axonopus compressus and M. bracteata covered 80% of the surface at 4 weeks after planting (WAP), followed by C. pubescens at 5 WAP and C. caeruleum and P. javanica at 6 WAP (). Mean shoot dry weight per plant of A. gangetica decreased as the proportion of A. compressus, M. bracteata and P. javanica increased. The highest A. gangetica shoot dry weight was obtained in the pure stand of A. gangetica (W100) and the lowest in C75:W25 (). Calopogonium caeruleum and C. pubescens had no effect on A. gangetica shoot dry weight in any proportions. Mucuna bracteata and P. javanica reduced A. gangetica shoot dry weight when present in equal proportions (C50:W50). No statistical differences were observed between shoot biomass per plant of M. bracteata in different proportions. However, A. compressus, C. caeruleum, C. pubescens and P. javanica in monoculture (C100), C75:W25 and C50:W50 produced less shoot biomass per plant than in C25:W75 (). Canopy height of cover crops and A. gangetica was affected differently by the C:W proportions in the mixture (). Canopy height of different cover crops was unaffected by their proportions in mixtures. Asystasia gangetica had its highest canopy height in monoculture, but had reduced height in mixtures. Asystasia gangetica canopy height was similar when the proportion of cover crops to A. gangetica increased in mixtures, except with C. caeruleum. In general, canopy height of A. compressus, C. caeruleum, C. pubescens and P. javanica was lower than that of A. gangetica in monoculture, whereas that of M. bracteata and A. gangetica was similar (). Mucuna bracteata and P. javanica had the tallest canopy and C. caeruleum and C. pubescens the shortest. Leaf area of cover crops in association with A. gangetica responded in a similar way to shoot dry weight. Thus all the cover crops except M. bracteata had the highest leaf area in C25:W75. Asystasia gangetica leaf area decreased with increasing cover crop proportion in the mixture and was lowest in C75:W25. Light penetration in the C. pubescens treatments was higher than for the other legumes ().
Table II. Shoot dry weight, canopy height and leaf area of cover crop species and Asystasia gangetica in monoculture and mixtures at 12 weeks after planting.
Competition between cover crops and Pennisetum polystachion
The RY of P. polystachion increased in a quadratic manner as the proportion of P. polystachion in mixtures with C. caeruleum and P. javanica increased, which resulted in a concave curve for C. caeruleum and P. javanica (B, E). The response of P. polystachion with A. compressus and C. pubescens was linear and more than expected (A, C). However, the response of P. polystachion in interaction with M. bracteata was linear and near to the expected rate (D). The RY of A. compressus, C. pubescens and M. bracteata increased in a linear manner and was less than expected as their proportion in the mixture with P. polystachion increased, but the response of A. compressus and C. pubescens was greater than that of M. bracteata and was near to the expected rate. The RY of cover crops and P. polystachion was not equivalent when each comprised half the mixture () resulting in an EYR of 0.46, 0.21, 0.37, 0.39 and 0.33 for A. compressus, C. caeruleum, C. pubescens, M. bracteata and P. javanica, respectively. The RYT value of P. polystachion with M. bracteata was less than 1 for all mixtures, while with the other cover crops it was more than 1 (). The RCC of P. polystachion over cover crops when grown in equal proportions was more than the RCC of cover crops over P. polystachion ().
Figure 2. Interaction between cover crops and Pennisetum polystachion in replacement-series experiments at 12 weeks after planting. (A) Axonopus compressus response: Y = − 0.95X + 1.01, R2=0.98; Pennisetum polystachion response: Y = 0.90X + 0.14, R2=0.87; (B) Calopogonium caeruleum response: Y = 1.20X2−2.11X + 0.94, R2=0.95; Pennisetum polystachion response: Y = − 1.49X2+2.36X + 0.07, R2=0.92; (C) Centrosema pubescens response: Y = − 0.90X + 0.88, R2=0.91; Pennisetum polystachion response: Y = 0.91X + 0.19, R2=0.81; (D) Mucuna bracteata response: Y= −0.90X + 0.81, R2=0.83; Pennisetum polystachion response: Y = 0.81X + 0.11, R2=0.81; (E) Pueraria javanica response: Y = 0.31X2−1.19X + 0.94, R2=0.93; Pennisetum polystachion response: Y = − 1.04X2+1.96X + 0.04, R2=0.97.
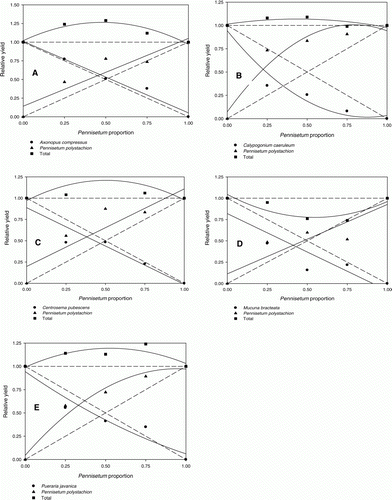
Table III. Relative crowding coefficient in interactions between different cover crop species and Pennisetum polystachion and time of 80% coverage by cover crops.
Canopy height, leaf area and shoot number of P. polystachion increased significantly in a linear manner when this weed was grown with cover crops compared with monoculture. The highest values of all three parameters at harvest occurred in the presence of maximum proportion of cover crops (C75:W25) (). Pennisetum polystachion was taller than any of the cover crops either when grown alone or in mixture. Pennisetum polystachion canopy height was affected by cover crop in the other mixtures as in C75:W25. Shoot dry weight, leaf area and canopy height of A. compressus and C. pubescens and canopy height of M. bracteata in interaction with P. polystachion did not vary for the different proportions. Other characteristics of C. pubescens, M. bracteata and P. javanica decreased with increasing P. polystachion proportion in mixtures, but sometimes showed an increase in C25:W75. However, in the equal proportions mixture (C50:W50), shoot dry weight, leaf area and shoot number of P. polystachion in interaction with M. bracteata were less than those recorded with the other cover crops ().
Table IV. Shoot dry weight, canopy height, leaf area of cover crop species and Pennisetum polystachion in monoculture and mixtures at 12 weeks after planting.
Discussion
As the proportion of A. gangetica in the mixtures increased, all the cover crop species except M. bracteata yielded over the expected rate and had a convex curve, whereas A. gangetica yielded under the expected rate and had a concave curve. Cover crops were more competitive than A. gangetica, as indicated by the concave curve for relative yield (RY) of A. gangetica as its proportion increased in the mixture. In general, a convex curve for one species and a concave curve for the other species in the series indicate that the species are competing for a common resource. Mutually stimulatory and antagonistic relations are indicated when both curves are convex and concave, respectively (Harper Citation1977). The RY of A. gangetica in interaction with M. bracteata was less than that of the pure stand, while the RY of M. bracteata was unaffected, indicating that M. bracteata did not benefit from its interaction with A. gangetica. This may be because the strong taproot system of M. bracteata grew extensively to take up nutrients in lower soil horizons, while the weak root system of A. gangetica absorbed nutrients in the upper soil horizon. Asystasia gangetica was more affected by interspecific interactions, as it grew better in pure stand than in mixtures. All five cover crop species produced a convex or linear response that resulted in EYR>0.50, suggesting that they were more competitive than A. gangetica. For EYR, the cover crop species performed in the order: P. javanica>A. compressus>M. bracteata>C. caeruleum>C. pubescens.
An RYT value around 1 indicates that the same resource or area is being used by the two competing species (overlap in resource utilization) (Jolliffe et al. Citation1984). An RYT>1 indicates some niche differentiation between the species, where competition is either avoided or minimized (Fischer et al. Citation2000). However, other processes can also produce RYT>1, indicating facilitation where one species benefits another (Hector Citation2006). An RYT<1 suggests mutual antagonism. In this experiment, underyielding (RYT<1) occurred with M. bracteata, over-yielding (RYT>1) with C. caeruleum and competition (RYT=1) with A. compressus, C. pubescens and P. javanica. This indicates that the dry weight of A. gangetica was strongly inhibited by the cover crops. The RYT value of C. caeruleum was>1, showing that it is more aggressive than A. gangetica and more competitive for resources. It also suggests that the two species make different demands on resources, leading to better growth of C. caeruleum, or that this crop:weed mixture is affected less by interspecific competition than by intraspecific competition, allowing for overyielding (RYT>1). Use of RYT has been suggested as a possible control of weeds by managing fertilizer application in the field, as RYT>1 indicates differences in species demand for potassium (Qasem Citation2010). Ong et al. (Citation2008) showed that improving nutrient availability in Azadirachta axcelsa trees by adding N fertilizer changed light penetration to A. gangetica and decreased its coverage.
Relative crowding coefficient (RCC) of A. compressus, C. caeruleum, C. pubescens, M. bracteata and P. javanica on A. gangetica when grown in equal proportions (C50:W50) exceeded the RCC of A. gangetica on them, indicating that the cover crops were generally more dominant than A. gangetica. The RCC was highest for P. javanica over A. gangetica, followed by M. bracteata, C. caeruleum, A. compressus and C. pubescens. Centrosema pubescens had the lowest RCC on A. gangetica, indicating that they were competing for the same resources and that C. pubescens reduced its growth more than the other crops for a given proportion. The highest RCC of A. gangetica on crops was recorded for C. pubescens. The hierarchy described by these competition experiments is: P. javanica>M. bracteata>C. caeruleum> A. compressus>C. pubescens.
All cover crops except M. bracteata produced more dry weight per plant when competing against the highest A. gangetica mixture (C25:W75) than in monoculture. However, shoot dry weight of A. compressus, C. caeruleum, C. pubescens and P. javanica remained unchanged from C100 to C50:W50, confirming that as the proportions changed over this range, intraspecific and interspecific competition counteracted each other. However, as lower densities of those plants were present in the C25:W75 mixture, it appeared that they were probably sufficiently far apart so that intraspecific competition was reduced. In such conditions, A. compressus, C. caeruleum, C. pubescens and P. javanica were more efficient in competing against A. gangetica. In contrast, A. gangetica shoot dry weight accumulation did not vary between C:W proportions, indicating low intraspecific competition. These findings suggest that A. compressus, C. caeruleum, C. pubescens and P. javanica responded plastically to competition, whereas A. gangetica did not. The observed RY values for those four cover crops, which were greater than the expected value, indicate that the mixture stands exceeded the average yield of monoculture stands.
The overyielding of these cover crops in the mixtures with A. gangetica can be explained by differences in their rooting system. Asystasia gangetica has a weak root system, so A. compressus, C. caeruleum, C. pubescens and P. javanica took advantage of the lower resistance belowground to produce profuse root mass. In those cover crops, the strong taproot system grew extensively to take up nutrients in lower soil horizons and the profuse lateral root system ramified the upper soil horizon to absorb the nutrients there. More competitive species produce a higher relative yield when grown in mixtures, whereas the yield of weak competitors is lower in mixture than in monoculture (Spehn et al. Citation2000, Gao et al. Citation2005). When one plant of basil (Ocimum sanctum) was competing with three of weed species, plant height and fresh weight/plant of basil increased (Singh and Singh Citation2009), and the increasing value of RTY for this ratio indicated some degree of resource complementarity. Overyielding has been associated with higher biomass density and light interception, but not increased canopy height (Vojtech et al. Citation2008). Overyielding is predicted to be smaller when species are not functionally different (Turnbull et al. Citation2005, Mwangi et al. Citation2007). Complementarity in plant architecture between grasses and herbs has been shown to be particularly important for increased light absorption, better three-dimensional space filling and greater biomass density in mixtures, whereas mixtures with only grasses show no increase in biomass accumulation and LAI (Spehn et al. Citation2000).
Cover crops and A. gangetica differed in their competitive abilities. Mechanisms by which weeds are suppressed include competition for resources, microclimate modification inhibiting weed seed germination and allelopathy. In addition, rapid canopy closure leads to reduced PAR for weed growth (Teasdale Citation1998). In pure and mixed stands, the cover crop tested here accumulated higher dry matter than A. gangetica. However, differences in growth, plant height and leaf area traits could explain the differences in interspecific competition. Mucuna bracteata grew faster than A. gangetica, producing taller plants and higher leaf area, which could have contributed to its better competitive performance. Pueraria javanica did not grow faster than M. bracteata but achieved a tall canopy and enough leaf area to suppress A. gangetica. However, C. pubescens had its maximum biomass in monoculture and covered the ground faster than P. javanica but the canopy was not high enough to shade A. gangetica. Furthermore, C. pubescens allowed more light to penetrate than the other legumes. Species of greater productivity in monoculture may not be effective competitors in mixtures (Harper Citation1977). The superior competitive ability of M. bracteata and P. javanica was most likely related to light exclusion (at the early stage of growth) associated with rapid establishment rate. Fast growth and taller stature result in a superior plant (Mosjidis and Wehtje Citation2011). Furthermore, plant size (plant weight and height) suggests a potential advantage for light capture and greater penetration of PAR to the soil surface, making the crop less competitive against weeds (Collins et al. Citation2007). It is likely that the mechanism of A. gangetica suppression by A. compressus is resource competition. Axonopus compressus became extremely dense by 6 WAP, thereby limiting the space available to the weed population and suppressing A. gangetica growth.
In the case of P. polystachion, its RY increased in a quadratic manner with C. caeruleum and P. javanica and linearly with A. compressus and C. pubescens to more than expected, indicating that P. polystachion was a superior competitor. The RY of cover crops in interaction with P. polystachion was concave or linear but less than expected, indicating that the cover crops were less competitive than P. polystachion. If the RY curves intersect at 50:50 proportions, the two competing species are relatively equal in competitiveness. The RY of cover crops increased in a linear or non-linear manner as their proportion in the mixture with P. polystachion increased, but their RY was not equivalent to that of P. polystachion when each comprised half the mixture. This resulted in EYR<0.50, suggesting that a large population of cover crop is needed to suppress a smaller population of P. polystachion. The RCC value demonstrates the aggressiveness of one species towards another. The greater RCC of P. polystachion over cover crops confirms the aggressiveness of this weed against the cover crops in terms of shoot dry weight production. When competing for limited resources, the species with the greater RCC in the mixture is the stronger competitor (Fischer et al. Citation2000). It appeared that not all cover crops had a negative impact on plant weight, leaf area and shoot numbers, since higher values of those parameters were found in the presence of increasing proportion of cover crops than in the pure P. polystachion stands. When P. polystachion was grown together with any of the five cover crops, resource capture from the limited resources available in these poly bag experiments favoured weed growth at the expense of cover crop growth. Thus even when the planting density of cover crops was threefold that of P. polystachion, the cover crops were not able to capture sufficient resources to promote their growth. Therefore, P. polystachion grew better in the mixture than in monoculture and was a better competitor than the other species.
One explanation for the superiority of P. polystachion over cover crops could be that no negative impact resulted from the interaction and that P. polystachion actually benefited from the interaction with A. compressus, C. caeruleum, C. pubescens and P. javanica, possibly because of the additional nitrogen contributed by these legumes. Pennisetum polystachion responded in a linear manner and nearly to the expected level as its proportion in the mixture with M. bracteata increased, indicating that it did not benefit from nitrogen fixation by M. bracteata. In the interaction of M. bracteata and P. polystachion allelopathy is likely to be involved, because of the lower growth increase of P. polystachion with M. bracteata compared with the other legumes. Furthermore, the RYT value of P. polystachion with M. bracteata was<1 for all proportions. This means that mutual antagonism was occurring, with the species producing less than expected when grown together. The occurrence of allelopathic interaction would have lowered the total yield in mixtures compared with monoculture (Cralle et al. Citation2003). Spitters (Citation1983) found niche differentiation between maize and peanut and noted that niche differentiation is very common when legume and grass plants are mixed. There was a lesser degree of competition between P. polystachion and A. compressus, as indicated by the large EYR, which was near to the expected RY of A. compressus, the lower RCC of P. polystachion over A. compressus compared with the other cover crops and the unaffected plant weight, height and leaf area of A. compressus when grown with P. polystachion. Oka Nurjaya (Citation2006) reported that in mixtures, A. compressus was more competitive than grass and legume species, while Rika et al. (Citation1990) observed that under coconut, a local cultivar of A. compressus produced higher yield than other grasses.
Pennisetum polystachion shoot dry weight production was greater than that of the cover crops and was strongly competitive and aggressive toward the crops. Our results indicate that the rapid growth and photosynthetic efficiency of P. polystachion permit it to avoid shading by cover crops. The superior competitiveness of P. polystachion relative to other species in our experiments was primarily due to efficient light utilization. Higher plant size (plant weight and height) offers advantages for light capture. All the cover crops tested were smaller than P. polystachion and caused a lesser degree of competition for the same resources in the mixture compared with the monoculture of each species (Crotser and Witt Citation2000). Light interception in P. polystachion bags was greater than in other bags, probably because P. polystachion established quickly and provided higher aboveground biomass than other species. It is also likely that the relatively open canopy of P. polystachion allowed rapid growth and reduced intraspecific competition. The amount of light intercepted or transmitted by any plant species is a measure of its canopy thickness, which is a function of leaf area index, phyllotaxy, form and size of leaves and branching. Clearly, P. polystachion had the highest canopy thickness relative to cover crops. Any plant that intercepts more light deprives other plants of light, leading to a drop in production/respiration ratio. Thus it is clear that any species that grows faster than the other species present produces a thicker canopy, occupies more space, becomes taller and ecologically dominant and is capable of inhibiting or even eliminating the other species (Asha Kumari et al. Citation2010). These results are in agreement with those of Kobayashi et al. (Citation2003), who showed that late ground-covering by the C. caeruleum shoot canopy increased light penetration under the canopy and caused perennial P. polystachion to be more competitive than C. caeruleum.
Pennisetum polystachion maintained its lateral spread through tillering, while the broadleaved species exhibited increasing values of canopy leaf area index. As Gealy et al. (Citation2003) stated, greater tiller production is one of the factors responsible for higher competitiveness. The aggressiveness of P. polystachion observed here can also be explained in terms of its prolific rooting system, which enabled it to capture more of the limited soil water and nutrients and resulted in rapid growth in terms of biomass accumulation, height and canopy development (Holt and Orcutt Citation1991).
All the cover crop species studied proved to be highly competitive against A. gangetica but none of them showed suppressive ability against P. polystachion because of their slow growth habit relative to that of P. polystachion. The practical implication of this study is that a cover crop can be used successfully for the management of A. gangetica. However, earlier planting and emergence of cover crops or early season weed control by pre-emergence herbicide application may help cause significant suppression of P. polystachion and reduce its competitiveness in the long run.
References
- Agamuthu , P. and Broughton , W. J. 1985 . Nutrient cycling within the developing oil palm–legume ecosystem . Agriculture, Ecosystems and Environment , 13 : 111 – 123 .
- Aldaba , F. R. 1995 . Coconut production in the Philippines: problems and prospects . Plantations, Recherche, Developpement , 2 : 15 – 21 .
- Asha Kumari , J. , Rama Chandra Prasad , P. and Reddy , K. B. 2010 . Competitive exclusion of Parthenium hysterophorus by other invasive species – A case study from Andhra Pradesh, India . Taiwania , 55 : 128 – 138 .
- Baki , H. B. 2004 . Invasive weed species in Malaysian agro-ecosystems: species, impacts and management . Malaysian Journal of Science , 23 : 1 – 42 .
- Collins , A. S. , Chase , C. A. , Stall , W. M. and Hutchinson , C. M. 2007 . Competitiveness of three leguminous cover crops with yellow nutsedge (Cyperus esculentus) and smooth pigweed (Amaranthus hybridus) . Weed Science , 55 : 613 – 618 .
- Collins , S. C. 1991 . Chemical control of grassy weeds . Tropical Grassy Weeds , 73 – 84 .
- Cousens , R. 1991 . Aspects of the design and interpretation of competition (interference) experiments . Weed Technology , 5 : 664 – 673 .
- Cralle , H. T. , Fojtasek , T. B. , Carson , K. H. , Chandler , J. M. , Miller , T. D. , Senseman , S. A. , Bovey , R. W. and Stone , M. J. 2003 . Wheat and Italian ryegrass (Lolium multiflorum) competition as affected by phosphorus nutrition . Weed Science , 51 : 425 – 429 .
- Crotser , M. P. and Witt , W. W. 2000 . Effect of Glycine max canopy characteristics, G. max interference, and weed-free period on Solanum ptycanthum growth . Weed Science , 48 : 20 – 26 .
- De Wit , C. T. 1960 . On competition . Versal. Landbouwk. Onderz , 66 : 1 – 82 .
- Fischer , A. J. , Messersmith , C. G. , Nalewaja , J. D. and Duysen , M. E. 2000 . Interference between spring cereals and Kochia scoparia related to environment and photosynthetic pathways . Agronomy Journal , 92 : 173 – 181 .
- Gao , Y. Z. , Wang , S. P. , Han , X. G. , Patton , B. D. and Nyren , P. E. 2005 . Competition between Artemisia frigida and Cleistogenes squarrosa under different clipping intensities in replacement series mixtures at different nitrogen levels . Grass and Forage Science , 60 : 119 – 127 .
- Gealy , D. R. , Wailes , E. J. , Estorninos , L. E. Jr and Chavez , R. S. C. 2003 . Rice cultivar differences in suppression of barnyardgrass (Echinochloa crus-galli) and economics of reduced propanil rates . Weed Science , 51 : 601 – 609 .
- Goh , K. J. and Chiu , S. B. 2007 . Mucuna bracteata. A cover crop living green manure , Petaling Jaya : Agricultural Crop Trust (ACT) .
- Harper , J. L. 1977 . Population biology of plants , London : Academic Press .
- Hector , A. 2006 . Overyielding and stable species coexistence . New Phytologist , 172 : 1 – 3 .
- Holt , J. S. and Orcutt , D. R. 1991 . Functional relationships of growth and competitiveness in perennial weeds and cotton (Gossypium hirsutum) . Weed Science , 39 : 575 – 584 .
- Jolliffe , P. A. , Minjas , A. N. and Runeckles , V. C. 1984 . A reinterpretation of yield relationships in replacement series experiments . Journal of Applied Ecology , 21 : 227 – 243 .
- Kobayashi , Y. , Ito , M. and Suwanarak , K. 2003 . Evaluation of smothering effect of four legume covers on Pennisetum polystachion ssp. setosum (Swartz) Brunken . Weed Biology and Management , 3 : 222 – 227 .
- Kustyanti , T. & Horne , P. 1991 . The effect of Asystasia on the growth of young rubber in polybags . pdf.usaid.gov/pdf_docs/PDABG454.pdf [Online] .
- Lal , R. , Regnier , E. , Eckert , D. J. , Edwards , W. M. , & Hammond , R. 1991 . Expectations of cover crops for sustainable agriculture . In W. L. Hargrove , Cover Crops for Clean Water . Conference of the soil and water conservation society .
- Lehmann , J. , Da Silva , J. P. , Trujillo , L. and Uguen , K. 2000 . Legume cover crops and nutrient cycling in tropical fruit tree production . Acta Horticulturae , 531 : 65 – 72 .
- Madi 2009/2010 . Malaysia Agriculture Directory and Index .
- Mohamad , R. B. , Wibawa , W. , Mohayidin , M. G. , Puteh , A. B. , Juraimi , A. S. , Awang , Y. and Lassim , M. B. M. 2010 . Management of mixed weeds in young oil-palm plantation with selected broad-spectrum herbicides . Pertanika Journal of Tropical Agricultural Science , 33 : 193 – 203 .
- Mosjidis , J. A. and Wehtje , G. 2011 . Weed control in sunn hemp and its ability to suppress weed growth . Crop Protection , 30 : 70 – 73 .
- Mwangi , P. N. , Schmitz , M. , Scherber , C. , Roscher , C. , Schumacher , J. , Scherer-Lorenzen , M. , Weisser , W. W. and Schmid , B. 2007 . Niche pre-emption increases with species richness in experimental plant communities . Journal of Ecology , 95 : 65 – 78 .
- Oka Nurjaya , I. M. 2006 . Competition between some grass and legume species with Axonopus compressus under reduced light intensity . Jurnal Biologi . http://www.ejournal.unud.ac.id . [Online] .
- Ong , K. H. , Lim , M. T. , Priscilla , P. and Keen , J. C. 2008 . Ground vegetation response to fertilization in an Azadirachta axcelsa stand in Johore, Malaysia . Journal of Agronomy 327 – 331 .
- Qasem , J. R. 2010 . Competitive relationship of cauliflower (Brassica oleracea l. var. botrytis) and nettle-leaved goosefoot (Chenopodium murale l.) and their differential responses to potassium level . Journal of Plant Nutrition , 33 : 2135 – 2147 .
- Radosevich , S. R. 1987 . Methods to study interactions among crops and weeds . Weed Technology , 1 : 190 – 198 .
- Radosevich , S. , Holt , J. and Ghersa , C. 2007 . Ecology of weeds and invasive plants: Relationship to agriculture and natural resource management , New York : John Wiley & Sons .
- Rika , I. K. , Mendra , I. K. , Gustioka , M. and Oka Nurjaya , I. M. 1990 . “ New forage species for coconut plantation in Bali ” . In Proceedings of a workshop on forage for plantation crops , Edited by: Shelto , H. M. and Stur , W. W. Canberra : ACIAR .
- Santos , B. M. , Bewick , T. A. , Stall , W. M. and Shilling , D. G. 1997 . Competitive interaction of tomato (Lycopersicon esculentum) and nutsedges (Cyperus spp.) . Weed Science , 45 : 229 – 233 .
- SAS 2003 . The SAS system for Windows, Version 9.1 . Cary , NC : SAS Inst. Inc .
- Senarathne , S. H. S. and Perera , K. C. P. 2011 . Effect of several weed control methods in tropical coconut plantation on weed abundance, coconut yield and economical value . International Research Journal of Plant Science , 2 : 25 – 31 .
- Singh , S. and Singh , M. 2009 . Interactions of basil (Ocimum sanctum L.) with some weed species – competition or allelopathy? . Indian Journal of Weed Science , 41 : 12 – 22 .
- Spehn , E. M. , Joshi , J. , Schmid , B. , Diemer , M. and Körner , C. 2000 . Above-ground resource use increases with plant species richness in experimental grassland ecosystems . Functional Ecology , 14 : 326 – 337 .
- Spitters , C. J. T. 1983 . An alternative approach to the analysis of mixed cropping experiments. 1. Estimation of competition effects . Netherlands Journal of Agricultural Science , 31 : 1 – 11 .
- Teasdale , J. R. 1998 . Weed suppression by hairy vetch residue . Proceedings of the Northeastern Weed Science Society , 42 : 73
- Turnbull , L. A. , Rahm , S. , Baudois , O. , Eichenberger-Glinz , S. , Wacker , L. and Schmid , B. 2005 . Experimental invasion by legumes reveals non-random assembly rules in grassland communities . Journal of Ecology , 93 : 1062 – 1070 .
- Turner , P. D. and Gillbanks , R. A. 2003 . Oil palm cultivation and management , Kuala Lumpur , , Malaysia : The Incorporated Society of Planters .
- Vojtech , E. , Loreau , M. , Yachi , S. , Spehn , E. M. and Hector , A. 2008 . Light partitioning in experimental grass communities . Oikos , 117 : 1351 – 1361 .
- Weigelt , A. and Jolliffe , P. 2003 . Indices of plant competition . Journal of Ecology , 91 : 707 – 720 .
- Wibawa , W. , Mohamad , R. , Juraimi , A. S. , Omar , D. , Ghazali Mohayidin , M. and Begum , M. 2009 . Weed control efficacy and short term weed dynamic impact of three non-selective herbicides in immature oil palm plantation . International Journal of Agriculture and Biology , 11 : 145 – 150 .
- Wibawa , W. , Mohayidin , M. G. , Mohamad , R. B. , Juraimi , A. S. and Omar , D. 2010 . Efficacy and cost-effectiveness of three broad-spectrum herbicides to control weeds in immature oil palm plantation . Pertanika Journal of Tropical Agricultural Science , 33 : 233 – 241 .