Abstract
Microdochium nivale (Fr.) Samuels and Hallett (teleomorph Monographella nivalis (Schafnitt) is one of the most damaging pathogens of cool-season amenity turfgrasses. The search for new or improved means to reduce susceptibility to M. nivale is an ongoing target for turfgrass research. From 2010 to 2012, field studies were conducted at the Curragh Golf Club, Co Kildare, Ireland on Agrostis canina L. ssp. canina and Poa annua L. swards, to evaluate M. nivale suppression by 14-day sequential applications of potassium phosphite (KH2PO3), applied alone and in combinations with a fungicide (iprodione) and a biostimulant. Treatment effect on turfgrass quality was also determined. Over two years of trials, phosphite treated plots, when compared with untreated controls, exhibited significantly lower percentages (p<0.01) of M. nivale incidence. The addition of the biostimulant did not enhance this effect. The iprodione treatments showed significantly higher suppression than the phosphite treatments (p<0.01), and the addition of phosphite significantly enhanced the fungicide control of M. nivale (p<0.01). This last combined treatment was most effective overall, with full inhibition on the treated plots. Turfgrass quality on all phosphite treated plots was significantly better (p<0.01) than either the control or iprodione treated plots.
Introduction
Disease prevention is a major factor, and is one of the more problematic and contentious areas in the successful management of amenity turfgrasses. Pathogenic fungi and oomycetes are the major causal agents (Beard and Oshikazu Citation1997, Vargas Citation2005). The ascomycete pathogen, Microdochium nivale (Fr.) Samuels and Hallett (teleomorph Monographella nivalis (Schafnitt) (Smiley et al. Citation1992) is regarded as the most important disease of temperate climates, infecting most cool-season species and affecting up to 100% of golf greens in the UK each year (Smiley et al. Citation1992, Beard and Oshikazu Citation1997, Mann Citation2002, Vargas Citation2005). M. nivale was until recently thought to be a single species, with two varieties, var. M. nivale nivale and var. M. nivale majus, but it is now recognized as two distinct species: M. nivale and M. majus (Glynn et al. Citation2005, Hofgaard et al. Citation2006). M. majus differs from M. nivale in conidial morphology and host specificity (Lees et al. Citation1995, Maurin et al. Citation1995) and is responsible for many disease complexes in species of wheat, barley and oat (Pettitt et al. Citation1993, Humphreys et al. Citation1995, Clement and Parry Citation1998, Diamond et al. Citation1998). M. nivale proliferates on forage or turfgrasses (Mahuku et al. Citation1998). Control of M. nivale in turfgrass is achieved by implementing Integrated Pest Management (IPM) programmes (Beard and Oshikazu Citation1997) which reduce disease incidence to acceptable levels. Chemical plant protectants are an integral part of IPM and while the efficacy and safety of these products is not disputed their use sometimes gives rise to a number of issues: adverse public opinion due to the perception of high frequency of use, associated costs of chemicals, possible inhibition of non-target beneficial microorganisms, development of fungicide-resistant populations, and possible legislative restrictions.
The search for new or improved means to reduce susceptibility to M. nivale is an ongoing target for turfgrass research. One possible means is the use of phosphite (PO3 3 −, Phi) as either a fungicide or biostimulant, as a component of an integrated approach to disease management. Phi is derived from phosphorous acid – H3PO3, which, to prevent phytotoxicity, is commonly modified prior to use with potassium hydroxide (KOH), forming potassium dihydrogen phosphite (KH2PO3) or dipotassium hydrogen phosphite (K2HPO3) (Ouimette and Coffey Citation1988). In turfgrass management these form the active substances in numerous products currently marketed worldwide as either fungicides or fertilizers (Landschoot and Cook Citation2005). In the UK and Ireland over the past number of years, they have been marketed as biostimulants or even called defence activators; this helped stimulate this current research, the aim of which was to determine the effect Phi treatment has on turfgrass, in particular its possible reduction of M. nivale incidence.
Throughout the literature there are numerous examples of successful inhibition of phytopathogens by Phi, although the means by which this is achieved is often debated (Abbasi and Lazarovits Citation2006). Phi has proven very effective in controlling, in a preventative mode of use, oomycete pathogens (Sanders Citation1983, Cook Citation2009) with studies reporting Phi inhibition of pathogens by both direct fungistatic means and indirectly through stimulation of plant defence processes (Fenn and Coffey Citation1984, Citation1987, Saindrenan et al. Citation1988, Grant et al. Citation1990, Jackson et al. Citation2000, McCarren et al. Citation2009). There are also a number of studies in the literature reporting the inhibitory properties of Phi against ascomycetes. Phi treatments reduced mycelial growth of Alternaria alternata (Fr.) Keissl (Reuveni et al. Citation2003) and in vitro sensitivity to Phi of Colletotrichum graminicola (Ces.) Wilson, has also been reported (Burpee Citation2005). Hofgaard et al. (Citation2010) concluded that development of M. majus in winter wheat (Triticum aestivum L.) was significantly reduced by potassium phosphite (K2HPO3) treatment. In turfgrass management Phi was first used for the control of oomycete pathogens such as Pythium spp. (Cook et al. Citation2006, Schroetter et al. Citation2006). It was subsequently combined with Mancozeb, a dithiocarbamate fungicide, to control summer decline of bentgrass (Beard and Oshikazu Citation1997). Reports of improved turf quality following sequential applications of Phi have also been reported (Vincelli and Dixon Citation2005, Cook et al. Citation2006). Dempsey and Owen (Citation2010) determined a significant improvement in Agrostis stolonifera L. quality and density following treatment with Phi.
What can be concluded from these studies is that Phi has a complex mode of action, acting both directly on the pathogen and indirectly by stimulating host defence responses. The research confirms that Phi can significantly reduce the occurrence of many oomycete pathogens in numerous plant species, and while there is some evidence for similar success in controlling ascomycete pathogens, there are no studies specifically on the effect Phi may have on M. nivale infection in turfgrass.
Therefore, the aims of these field trials were to determine whether Phi can reduce susceptibility to M. nivale, whether the addition of a biostimulant could increase the efficacy of Phi to reduce M. nivale infection and if the addition of Phi to a standard fungicide (iprodione) could enhance the suppression of M. nivale compared with the fungicide alone. The effect Phi treatments had on turfgrass quality were also assessed and quantified during these trials.
Materials and methods
Trial plots and experimental design
Trial plots were established in 2010 on a sandy/loam soil, pH 7.1, within the area of the turf nursery at the Curragh Golf Club, Co Kildare, Ireland (53.15° N/6.80° W 110 m a.s.l.). Kildare has a temperate oceanic climate, with rainfall annual mean of 711 mm and air temperature mean of 9.2 °C; gives monthly and 30-year mean weather data for the location (Met_Eireann Citation2011). This mild climate gives rise to ideal conditions for turfgrass, with growth almost throughout the entire year. The plots, 2×2 m square, were composed of Poa annua L., Agrostis canina L. ssp. canina, variety Avalon and Agrostis stolonifera L., variety Penn G-6 and were maintained throughout the trial periods at 6 mm height of cut, using a John Deere 220 pedestrian mower. The P. annua sward previously formed part of a now disused fairway and for the 6 years previous to the trials had been maintained as part of a turfgrass nursery. The A. canina canina sward was originally established from seed in 2005 and was moved to the trial location as turfgrass sod in 2009. The A. stolonifera sward was established from seed in April 2010, however, due to its poor establishment, year one of the trials comprised the P. annua and A. canina canina plots only, with trial extension to include the A. stolonifera plots in year two.
Table I. Meteorological data for trial location.
Nutritional inputs
Granular nutrient inputs (Andersons 21:3:21) were applied at the beginning of May and September 2010 and 2011, at a rate of 30 g m−1, giving annual nutritional inputs (ANI) of 126 kg N ha−1, 7.9 kg P ha−1 (in the form of P2O5), 105 kg K ha−1. Two applications per annum were considered sufficient, as the N source contained 84.25% polymer coated urea giving a sustained release. A late-season application of a 21% nitrogen product could be construed as excessive and not representative of best management practices as it could encourage M. nivale infection, however, as M. nivale infection was the focus of this study it was deemed appropriate. Further nutrient inputs were supplied as part of the treatment applications and are detailed below. No irrigation inputs were required.
Treatments
Treatments were arranged in a randomized complete block design with five replications and comprised sequential applications, applied biweekly from September 2010 to March 2011 and from September 2011 to March 2012, using 20 L knapsack sprayers operating at 6 bar, calibrated to deliver 800 L ha−1. Treatments comprised:
| 1. | PK Plus 3:7:18 (Grigg Bros, 14% KH2PO3, specific gravity 1.37, pH 7.05) applied at 20 L ha−1(ANI: 11.5 kg N ha−1, 11.7 kg P ha−1 (in the form of P2O5), 57.3 kg K ha−1), 53.7 kg PO3 3 − ha−1. | ||||
| 2. | PK Plus (20 L ha−1) + Ultraplex Biostimulant (Grigg Bros, 5-0-3, specific gravity 1.26, pH 2.5) applied at 10 l ha−1(ANI: 20.5 kg N ha−1, 11.7 kg P ha−1 (in the form of P2O5), 61.7 kg K ha−1), 53.7 kg PO3 3 − ha−1. | ||||
| 3. | Chipco Green (Bayer, 255 g L iprodione) applied at 20 L ha−1. | ||||
| 4. | Chipco Green (20 L ha−1) + PK Plus (20 L ha−1) (ANI: 11.5 kg N ha−1, 11.7 kg P ha−1 (in the form of P2O5), 57.3 kg K ha−1), 53.7 kg PO3 3 − ha−1. | ||||
| 5. | NPK control (3:7:18 to match nutritional input of PK Plus) applied at 20 L ha−1(ANI: 11.5 kg N ha−1, 11.7 kg P ha−1 (in the form of P2O5), 57.3 kg K ha−1). | ||||
| 6. | Untreated control. | ||||
Biweekly applications of iprodione to control M. nivale would be considered excessive for routine turfgrass management, as it could lead to resistance in the pathogen population; this rate was used during the trials to ensure maximum suppression.
Data collection and analysis
Trial plots were assessed monthly for M. nivale occurrence from September 2010 to March 2011 and from September 2011 to March 2012, and rated on a scale of 0–100, evaluation being percentage of plot area affected by disease: 0 being no incidence and 100 being 100% coverage (Bruneau et al. Citation2000). Turf quality (which excluded the impact of disease within each plot) was also measured during both trial periods; this was assessed visually and marked on a scale of 1–10 (1 = dead turf, 5 = acceptable, 10 = excellent) (Horvath et al. Citation2007). For both disease incidence and turf quality, data were subject to analysis of variance (ANOVA) using SPSS Statistics 19.0 and significant treatment differences were separated by Tukey least significant difference test (LSD) at p<0.01. Because the plots were of two turfgrass species each with different growth habits and susceptibilities to M. nivale, plus the fact that the A. canina canina plots were located in a significantly more shaded area compared with the P. annua plots, thus increasing the disease pressure, data analyses were carried out and reported separately for each species.
Results
Disease incidence
In both years of the study M. nivale occurrence developed naturally, with high levels of infection from October 2010 to February 2011 and from September 2011 to February 2012. Analysis of the data determined significant differences in treatment effect (p<0.01) ().
Table II. Analysis of variance of treatment effect on M. nivale incidence on trial plots.
In year one, M. nivale became active mid October 2010, with disease incidence progressively increasing and peaking, following a 3-week covering of snow, at the end of December 2010. Disease pressure declined from the end of January 2011 and became non-existent from mid February 2011, until environmental conditions allowed re-emergence in September 2011. shows treatment effect on percent M. nivale incidence on the trial plots (n = 5) for each of the months of the first year's trial.
Table III. Treatment effect on percent M. nivale incidence on trial plots in year one.
In year two disease pressure became evident earlier than in the previous year with M. nivale incidence beginning mid September and remaining at high levels to mid December 2011. From January 2012 disease pressure declined gradually and ceased in March 2012. shows treatment effect on percent M. nivale incidence on the trial plots (n = 5) for each of the months of the second year's trials. During the second year, the trials were extended to include A. stolonifera plots, but as only one year's data are available they are not included in the full analysis.
Table IV. Treatment effect on percent M. nivale incidence on trial plots in year two.
Data from both years’ trials, as shown in and , determined that in both turfgrass species the control and NPK control plots displayed the highest mean values of disease incidence (p<0.01) ranging from 32 to 59%, levels of disease cover which would be unacceptable on any golf green. The application of Phi (Griggs PK Plus) reduced the incidence of the disease by more than 50%, with mean values ranging from 14 to 21%, significantly lower (p<0.01) than the control and NPK control plots. The addition of the biostimulant and surfactant package (Ultraplex) to the PK Plus treatments, did not cause any significant reduction in disease incidence compared with the PK Plus treatment alone.
Figure 1. Treatment effect on percent occurrence of M. nivale on P. annua trial plots. Data are mean values from September 2010 to March 2011 and from September 2011 to March 2012 (n = 5), letters indicate significant differences at p<0.01according to Tukey least significant difference test (LSD).
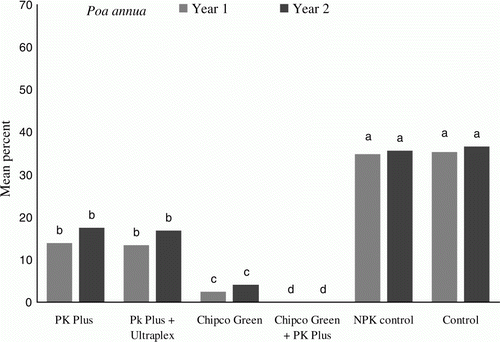
Figure 2. Treatment effect on percent occurrence of M. nivale on A. canina canina trial plots. Data are mean values from September 2010 to March 2011 and from September 2011 to March 2012 (n = 5), letters indicate significant differences at p<0.01 according to Tukey least significant difference test (LSD).
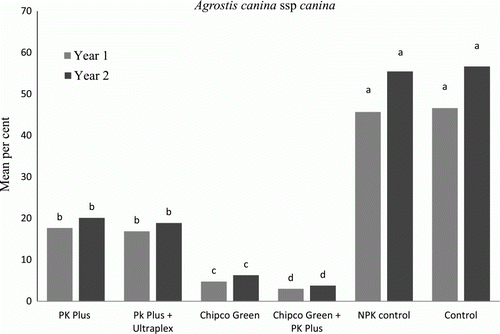
The plots which received iprodione (Chipco Green) treatments, as would be expected, displayed very high disease reductions, 92% less disease incidence than the control and NPK control and 33% less than the PK plus, PK Plus/Ultraplex treatments. However, despite the fact that the Chipco Green applications were applied at an extreme rate on a 2-week cycle, M. nivale was not fully inhibited. The treatments which were most effective at reducing M. nivale incidence (p<0.01) were the combination of Chipco Green and PK Plus. These fully inhibited M. nivale incidence on the P. annua plots and on the A. canina canina plots reduced it by 93% compared with the control plots. This total inhibition could indicate either differing modes of suppression or a possible synergistic effect.
Turfgrass quality
shows data on turfgrass quality from the P. annua and A. canina canina plots over the two trial periods. Turf quality at the beginning of both periods was uniform for all treatment plots, as the trials continued however, the quality of the control and NPK controls became progressively poorer, in comparison with the PK Plus and PK Plus/Ultraplex treatments. The Chipco Green treated plots, even with significantly less disease incidence than the PK Plus and PK Plus/Ultraplex treated plots, produced a poorer quality sward than the Phi treatments. The PK Plus/Chipco Green treatment produced the highest quality ratings overall. By the end of each trial period, the quality of all Phi treated swards was far superior to all other treatment regimes.
Table V. Treatment effect on turfgrass quality, assessed on a scale of 1–10.
Discussion
The data from the 2 years of these field trials have shown that, through periods of high disease pressure, as a result of sequential applications of Phi, there were significant and consistent reductions of M. nivale incidence in two commonly used cool-season turfgrasses. These trial results are supported by a previous study where significant disease reduction was reported in Phi-treated A. stolonifera (Dempsey and Owen Citation2010).
The mode of action of Phi in reducing M. nivale occurrence in these field trials has yet to be fully determined; there are numerous published reports showing Phi-mediated reduction of plant pathogens by both direct fungistatic means and indirectly through stimulation of plant defence processes (Fenn and Coffey Citation1987, Grant et al. Citation1990, Jackson et al. Citation2000, McCarren et al. Citation2009). Fenn and Coffey (Citation1987) for example, reported direct inhibition of Phytophthora spp. in Phi-treated Persea indica L.; while a study involving a number of Phytophthora species determined that Phi interfered with phosphate metabolism, by causing an accumulation of polyphosphate and pyrophosphate, resulting in a decrease in the pathogen's growth (Niere et al. Citation1994). In 2000, data were published showing that Phi inhibited enzymes of the glycolytic and phosphogluconate pathways, thus disrupting metabolism in Phytophthora spp. (Stehmann and Grant Citation2000). The possibility that Phi had a direct fungistatic effect on M. nivale in these field trials is a distinct possibility and in vitro research in this area is currently being carried out. Previously published in vitro studies into the inhibitory effect Phi has on the growth of ascomycetes have shown promising results. Reuveni et al. (Citation2003) reported mycelial growth of Alternaria alternata (Fr.) Keissl, was sensitive to Phi with an in vitro EC50 value (the effective dose which reduces growth to 50% of the growth of controls) of 278 µg ml−1 and Burpee (Citation2005) reported in vitro sensitivity to Phi of Colletotrichum graminicola (Ces.) Wilson, with an EC50 value of 121.9 mg ml−1 (EC50 values are of the active ingredients of the products used in both studies – no data available to calculate exact PO3 3 − values). The most relevant research to the results of these field trials was by Hofgaard et al. (Citation2010), who reported reduced in vitro growth of M. majus, when potassium phosphite (K2HPO3) was added to the growth medium (Hofgaard et al. Citation2010).
That disease suppression in these field trials was due to possible indirect inhibition of M. nivale by Phi, through stimulation or enhancement of inducible plant defence mechanisms is also a possibility. Jackson et al. (Citation2000) in a study of pathogen-infected Eucalyptus marginata L. concluded that Phi accumulations in planta led to stimulation of host defences. Phi treatment resulted in induced rapid defence responses, including release of Reactive Oxygen Species, localized cell death and an increase in defence-related phenolic compounds in Arabidopsis thaliana L. (Daniel and Guest Citation2006) and Saindrenan et al. (Citation1988) determined that Phi treatment enhanced the rate and magnitude of phytoalexin accumulation in Vigna unguiculata L. Analysis of Phi-treated turfgrass tissues with the aim to determine any increased synthesis of stress-related compounds would be beneficial.
The more likely hypothesis to explain the Phi-mediated disease reductions in these field trials is that M. nivale reduction was due to the combined effects of direct inhibition of the pathogen and enhanced host defence responses. Phi-induced effects on M. nivale metabolism may slow hyphal growth in planta, allowing increased time for the infected plant to respond. Phi may also induce the release of stress metabolites in M. nivale, enabling a more rapid recognition of the pathogen by the host plant. A more efficient recognition process allows a more rapid and hence more effective defence response, thus limiting the development of disease.
Turfgrass quality
Turfgrass quality is a vital aspect of turfgrass management and is determined by a combination of colour, density, uniformity and texture. The search for products or procedures which improve quality is constant. The data obtained over the 2 years of these trials, as shown in , determined there was significantly better visual quality on all Phi-treated plots. These data are supported by a previous study by Dempsey and Owen (Citation2010) who reported significant improvement of quality in Phi-treated A. stolonifera.
The reasons for the increase in turf quality could be attributed to a number of possible factors; firstly Phi treatments could inhibit pathogenic soil micro-organisms such as oomycetes and algae – while not displaying disease symptoms these could have a debilitating effect on turfgrass development. Phi has proven efficacy in reducing these organisms (Abbasi and Lazarovits Citation2006, Daniel and Guest Citation2006), thus allowing for the healthier development of the turfgrass. A second possibility is that the reduction of M. nivale allowed unrestricted growth and development, thus allowing for the improvement of the treated turfgrass. A further factor which could be considered is that Phi influenced a change in growth habit, Schroetter et al. in their 2006 study found that Phi applications to maize (Zea mays L.) led to an abnormality in growth habit, with the treated samples exhibiting an increase in lateral tillering, this could be a reason for the improvement, especially in the increase in turf density. Any of the possible factors above could have, either separately or in tandem, have led to the improvement in the turf quality; further study in this area is currently underway.
While the ability of Phi to lessen M. nivale incidence and improve turf quality was clearly demonstrated during these trials, a number of anomalies do require discussion. M. nivale incidence was consistently and significantly greater on the A. canina canina plots compared with the P. annua. This is due to a number of factors. The site of the A. canina canina sward, while in the same general area as the P. annua plots, was subject to extreme shading from trees, especially during the autumn and winter months. This ensured that environmental conditions for M. nivale occurrence were at an optimum. The P. annua plots had optimum light conditions and excellent air movement. A second factor to be considered is that A. canina canina, when not previously hardened by low temperatures, is highly susceptible to M. nivale infection (Espevig Citation2011), furthermore of six varieties of A. canina canina used in the study by Espevig, Avalon, the variety used in these field trials, proved more susceptible than all others.
There were no significant differences in treatment effect on either disease incidence or turf quality between the NPK control and untreated control plots in both trialled species. The N amount supplied by the treatment (11.5 kg ha−1) was relatively small, less than 10% of the total annual N input of 126 kg ha−1 and so it could be expected not to have a significant effect on either disease or quality. However, it could have been assumed that the application of relatively high amounts of K (57.3 kg ha−1) would have contributed to a reduction in disease incidence, but this was not the case. A more interesting and significant comparison is between the PK Plus and the NPK treatments. Both received identical nutritional inputs, the only difference being the Phi component of PK Plus (53.7 kg PO3 3− ha−1). This addition stimulated reduced disease incidence and a significant improvement in turfgrass quality in both turfgrass species.
These trial assessments are continuing and will gather further data to enhance the results from this present study. Further research to determine the processes involved in the Phi-mediated reduction of M. nivale is required and is presently underway. A wide range of laboratory procedures are currently being undertaken to determine the fungistatic properties of Phi and to evaluate Phi-mediated induction of secondary metabolic defence processes leading to induced resistance in treated turfgrasses.
In summary, what can be concluded from the results of these field trials is that routine and sequential applications of Phi, as part of an integrated disease management programme, while not reducing M. nivale to an acceptable level, will significantly reduce the incidence and severity of the disease. Furthermore, the addition of Phi to iprodione can significantly enhance suppression of M. nivale. Phi treatment also gave rise to significant improvement in turfgrass quality when compared with the untreated plots.
References
- Abbasi , P. A. and Lazarovits , G. 2006 . Seed treatment with phosphonate (AG3) Suppresses Pythium damping-off of cucumber seedlings . Plant Disease , 90 : 459 – 464 .
- Beard , J. and Oshikazu , T. 1997 . Colour atlas of turfgrass diseases , Chichester : John Wiley and Sons .
- Bruneau , A. H. , Newell , A. J. and Crossley , F. M. E. 2000 . Comparative performance of Bentgrass species and cultivars in close mown turf . Journal of Turfgrass Science , 76 : 63 – 69 .
- Burpee , L. L. 2005 . Sensitivity of Colletotrichium graminicola to phosphonate fungicides . International Turfgrass Society Research Journal , 10 : 163 – 169 .
- Clement , J. A. and Parry , D. W. 1998 . Stem-base disease and fungal colonisation of winter wheat grown in compost inoculated with Fusarium culmorum, F. graminearum and Microdochium nivale . European Journal of Plant Pathology , 104 : 323 – 330 .
- Cook , J. , Landschoot , P. J. , & Schlossberg , M. J. 2006 . Phosphonate products for disease control and putting green quality . Golf Course Management April , 93 – 96 .
- Cook , P. J. 2009 . Inhibition of Pythium spp. and suppression of Pythium blight and anthracnose with phosphonate fungicides . MSc dissertation . Altoona , PA : Pennsylvania State University .
- Daniel , R. and Guest , D. 2006 . Defence responses induced by potassium phosphonate in Phytophthora palmivora-challenged Arabidopsis thaliana . Physiological and Molecular Plant Pathology , 67 : 194 – 201 .
- Dempsey , J. & Owen , A. G. 2010 . The effect of phosphite treatments on the growth and disease susceptibility of Agrostis stolonifera L . 2nd European Turfgrass Society Conference , Angers , France , European Turfgrass Society .
- Diamond , H. , Cooke , B. M. and Cregg , B. 1998 . Morphology of sporodochia of Microdochium nivale var. nivale on cereal hosts using cryoscanning electron microscopy . Mycologist , 12 : 118 – 120 .
- Espevig , T. 2011 . Winter hardiness and management of velvet bentgrass (Agrostis canina) on golf greens in the Nordic climate . PhD dissertation . Norwegian University of Life Sciences .
- Fenn , M. E. and Coffey , M. D. 1984 . Studies on the in vitro and in vivo antifungal activity of Fosetyl-Al and phosphorus acid . Phytopathology , 74 : 606 – 611 .
- Fenn , M. and Coffey , M. D. 1987 . Phosphonate fungicides for control of diseases caused by Phytophthora . California Avocado Society 1987 Yearbook , 71 : 241 – 249 .
- Glynn , N. C. , Hare , M. C. , Parry , D. W. and Edwards , S. G. 2005 . Phylogenetic analysis of EF−1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties majus and nivale to species status . Mycological Research , 109 : 872 – 880 .
- Grant , B. R. , Dunstan , R. H. , Griffith , J. M. , Niere , J. O. and Smillie , R. H. 1990 . The mechanism of phosphonic (phosphorous) acid action in Phytophthora . Australasian Plant Pathology , 19 : 115 – 121 .
- Hofgaard , I. , Wanner , L. , Hageskal , G. , Henriksen , B. , Klemsdal , S. and Tronsmo , A. 2006 . Isolates of Microdochium nivale and M. majus differentiated by pathogenicity on perennial ryegrass (Lolium perenne L.) and in vitro growth at low temperature . Journal of Phytopathology , 154 : 267 – 274 .
- Hofgaard , I. S. , Ergon , Å. , Henriksen , B. and Tronsmo , A. M. 2010 . The effect of potential resistance inducers on development of Microdochium majus and Fusarium culmorum in winter wheat . European Journal of Plant Pathology , 128 : 269 – 281 .
- Horvath , B. J. , McCall , D. S. , Ervin , E. H. , & Zhang , X. 2007 . Physiological effects of phosphite formulations on turfgrass challenged with Pythium and heat stress . Virginia Turfgrass Journal January-February , 14 – 15 .
- Humphreys , J. , Cooke , B. M. and Storey , T. 1995 . Effects of seed-borne Microdochium nivale on establishment and grain yield of winter-sown wheat . Plant Varieties and Seeds , 8 : 107 – 117 .
- Jackson , T. J. , Burgessa , T. , Colquhounb , I. and Hardy , G. E. S. 2000 . Action of the fungicide phosphite on Eucalyptus marginata inoculated with Phytophthora cinnamomi . Plant Pathology , 49 : 147 – 154 .
- Landschoot , P. J. & Cook , J. 2005 . Sorting out the phosphonate products . Golf Course Management November , 73 – 77 .
- Lees , A. , Nicholson , P. , Rezanoor , H. and Parry , D. 1995 . Analysis of variation within Microdochium nivale from wheat evidence for a distinct sub-group . Mycological Research , 99 : 103 – 109 .
- Mahuku , G. , Hsiang , T. and Yang , L. 1998 . Genetic diversity of Microdochium nivale isolates from turfgrass . Mycological Research , 102 : 559 – 567 .
- Mann , R. 2002 . In vitro fungicide sensitivity of Microdochium nivale isolates from the UK . Journal of Turfgrass and Sports Surface Science , 78 : 25 – 30 .
- Maurin , N. , Rezanoor , H. N. , Lamkadmi , Z. , Somé , A. and Nicholson , P. 1995 . A comparison of biological, molecular and enzymatic markers to investigate variability within Microdochium nivale (Fries) Samuels and Hallett . Agronomie , 15 : 39 – 47 .
- McCarren , K. L. , Mc Comb , J. A. , Shearer , B. L. and Hardy , G. E. S. J. 2009 . In vitro influence of phosphite on chlamydospore production and viability of Phytophthora cinnamomi . Forest Pathology , 39 : 210 – 216 .
- Met_Eireann . 2011 . Monthly weather data . Available at : http://www.met.ie/climate/monthly-data.asp?Num=3723 (Accessed 14 December 2011 ).
- Niere , J. O. , Deangelis , G. and Grant , B. R. 1994 . The effect of phosphonate on the acid-soluble phosphorus components in the genus Phytophthora . Microbiology , 140 : 1661 – 1670 .
- Ouimette , D. G. and Coffey , M. D. 1988 . Quantitative analysis of organic phosphonates, phosphonate, and other inorganic anions in plants and soil by using high-performance ion chromatography . Phytopathology , 78 : 1150 – 1155 .
- Pettitt , T. R. , Parry , D. W. and Polley , R. W. 1993 . Improved estimation of the incidence of Microdochium nivale in winter wheat stems in England and Wales, during 1992, by use of benomyl agar . Mycological Research , 97 : 1172 – 1174 .
- Reuveni , M. , Sheglov , D. and Cohen , Y. 2003 . Control of moldy-core decay in apple fruits by β-aminobutyric acids and potassium phosphites . Plant Disease , 87 : 933 – 936 .
- Saindrenan , P. , Barchietto , T. , Avelino , J. and Bombeix , O. 1988 . Effects of phosphite on phytoalexin accumulation in leaves of cowpea infected with Phytophthora cryptogea . Physiological and Molecular Plant Pathology , 32 : 425 – 435 .
- Sanders , P. L. 1983 . Control of Pythium spp. and Pythium blight of turfgrass with fosetyl aluminum . Plant Disease , 67 : 1382 – 1383 .
- Schroetter , S. , Angeles-Wedler , D. , Kreuzig , R. and Schnug , E. 2006 . Effects of phosphite on phosphorus supply in corn (Zea mays) . Landbauforschung Volkenrode , 56 : 87 – 99 .
- Smiley , R. , Dernoeden , P. and Clarke , B. 1992 . Compendium of turfgrass diseases , St Paul , MN : APS Press .
- Stehmann , C. and Grant , B. 2000 . Inhibition of enzymes of the glycolytic pathway and hexose monophosphate bypass by phosphonate . Pesticide Biochemistry and Physiology , 67 : 13 – 24 .
- Vargas , J. 2005 . Management of turfgrass diseases , Chichester : John Wiley and Sons .
- Vincelli , P. & Dixon , E. 2005 . Performance of selected phosphite fungicides on greens . Golf Course Management July , 77 – 81 .