Abstract
The Colorado potato beetle (Leptinotarsa decemlineata) is probably the most dangerous pest species of potatoes. This study assessed the influence of expected climate warming on the number of Colorado potato beetle generations in the Wielkopolska region of Poland using two simulation models, SimLep 1 and NumoGen 1. Two types of data sets were utilised in the simulations: real meteorological data recorded in Winna Góra (52.2N; 17.4E) in the period 2002–2011 and virtual data obtained after transformation of the real data to include an increase in the recorded temperature values between 1 and 6°C. The threat to potato crops from Colorado potato beetle was found to increase in response to expected climate change. A temperature increase of 2°C above that recorded during the last few years would allow the development of two full generations of the beetle, and further temperature increases may even permit the occurrence of three generations.
Introduction
The Intergovernmental Panel on Climate Change (IPCC, Citation2007) projects increases in global mean surface temperatures of 1.1–6.4°C by the end of the twenty-first century. Similar to the global temperature the average temperature over Europe is also projected to continue increasing. Summer temperatures are projected to increase by up to 7°C in Southern Europe and 5°C in Northern Europe in the period 2080–2100 compared with the 1961–1990 average (van der Linden & Mitchell, Citation2009). It means that maximal temperature increase in Poland is projected to be about 6°C. Such changes in climate are likely to affect many natural processes that are temperature-dependent (Deutsch et al., Citation2008; Inouye, Citation2008). Insects are cold-blooded organisms and hence temperature is one of the most important environmental factors influencing their development (Petzoldt & Saeman, Citation2006). Temperature increases may therefore result in alterations in the geographical distribution of different insect species, variability in overwintering and population growth rates, increases in the number of generations, extension of the developmental period, effects on crop–pest synchrony, changes in interspecies interactions and increased risk of invasion by migrant pests (Porter et al., Citation1991). The close relationship between temperature and the development of cold-blooded organisms explains why insects are appropriate model candidates on which to study the effects of climate change (Chen et al., Citation2011).
The influence of temperature on the speed of development of a single generation in multivoltine species has been studied by many researchers (Collier et al., Citation1991; Ungerer et al., Citation1999; Mezeyová et al., Citation2008; Amiri et al., Citation2010). The objects of these studies have usually been insects of economic importance.
The Colorado potato beetle (Leptinotarsa decemlineata (Say)) is the most destructive insect pest of potatoes in North America and Europe (Ferro et al., Citation1985). If left uncontrolled, the pest can cause serious yield losses (Cranshaw & Radcliffe, Citation1980; Hare, Citation1980; Ferro et al., Citation1983). Experiments conducted in Poland have revealed yield losses caused by Colorado potato beetle amounting to 20% (Wójtowicz & Jörg, Citation2006) or 40% (Sidorczyk, Citation1965). Thus the beetle is also a serious pest in potato production in Poland (Przybysz et al., Citation1996; Pawińska & Mrówczynski, Citation2000; Węgorek et al., Citation2001). Overwintered adults emerge from the soil in the spring and search for food plants. Eggs are deposited on foliage in late spring. Larvae emerge and pass through four instars that feed on potato foliage. Mature larvae burrow into the soil and pupate. Approximately two weeks later, adults of the new generation emerge and begin feeding on the maturing potato plants. When ready to diapause, beetles either enter the soil within potato fields or move out in search of peripheral overwintering sites (Węgorek, Citation1959). The number of beetle generations depends on temperature and amounts to four in the warmest areas where it is found, where the development cycle takes 30 days, to one full or even a partial generation in colder regions. Thus in Poland and in Germany, where the mean annual air temperature is about 8 and 9°C, respectively, there is one full generation of Colorado potato beetle per year, whilst in Italy there are two and three generations in regions where the mean annual air temperature exceeds 14 and 16°C, respectively (Ferrari et al., Citation1993).
The close connection between air temperature and the number of pest generations (Anon, Citation1994) means that there is an urgent need to define the impact of predicted climate warming on the development of Colorado potato beetle. This has already been studied in Germany, Lithuania, Czech Republic and Austria (Šmatas et al., Citation2008; Wittchen & Freier, Citation2008; Kocmánková et al., Citation2010). The present study examined the influence of predicted temperature increases on the development of Colorado potato beetle in the Wielkopolska region of Poland, with the overall aim of determining the potential response of the pest to climate change and allowing future management policy to be formulated accordingly.
Methods
The study was carried out using two simulation models: SimLep 1 and NumoGen 1. SimLep 1 was originally developed in the former German Democratic Republic based on the literature data and the results of experiments conducted in the field and in the laboratory. The model was validated in the periods 1975–1981 and 1983–1985 (Kurth & Rossberg, Citation1986). According to the actual development, SimLep 1 presents the following results: date of first occurrence of the hibernating beetles, date of first occurrence of egg clusters, abundance of egg clusters, abundance of young larvae, abundance of old larvae and date of first occurrence of young adult beetles (Jörg et al., Citation2007). Detailed information on the sources used in construction of the SimLep 1 algorithms can be found in Kurth et al. (Citation1984). However, due to the fact that in Polish climate conditions the life cycle of Colorado potato beetle simulated by the SimLep 1 model began 3–4 weeks earlier than the observed life cycle, we abandoned the direct use of this model to predict the development of the pest. Instead, we used the results generated by SimLep 1 to estimate the effect of increasing temperature on the onset of the beetle egg-laying period, based on simulations using meteorological data recorded between 2002 and 2011 and virtual data obtained after raising the actual temperature by 1–6°C. The following relationship was obtained:
where y=acceleration of the onset of egg-laying (days), and x=temperature increase (1–6°C).
This equation was then incorporated into the model NumoGen 1, where it was used to determine the egg-laying date of the pest in conditions where the recorded temperature had increased by 1–6°C. The procedure for determining the date of egg-laying at elevated temperatures was performed after inputting data of the observed date of occurrence of this developmental stage in ambient thermal conditions. NumoGen 1 simulates the development of Colorado potato beetle from the occurrence of egg clusters to the beginning of the winter diapause and helps in estimation of the effect of temperature on the number of beetle generations. The model was developed for the purposes of the present study based on scientific reports describing the influence of temperature on successive stages of Colorado potato beetle, and publications on the effects of photoperiod on the pest diapause. Data presented by Larczenko (Citation1958) were used to simulate the development of the pest from egg to adult form, and data presented by Alyokhina and Ferro (Citation1999) were applied to estimate the dates of egg-laying by female beetles of the second or third generation. The beginning of winter diapause was determined on the basis of data reported by Tauber et al. (Citation1988), who found that all females reared at a photoperiod between 10:14 and 14:10 (L:D) entered diapause. This information was used to determine the end of the simulation period, with NumoGen 1 setting the date of beetle diapause based on day length in Winna Góra, Poland. The information about day length is available on the internet (http://calendar.zoznam.sk/sunset-pl.php). The beginning of the simulation was determined by the date of egg-laying on monitored potato crops or the date calculated by the NumoGen 1 model using the SimLep1 results. The first option was used for calculations based on meteorological data recorded in Winna Góra and the second when the temperature was increased by 1–6°C. Sample data are presented in . presents the parameters used in NumoGen 1. The NumoGen 1 model was validated in two ways, both of which concerned the development of the first generation of the beetle. One approach involved comparing the results generated by NumoGen 1 against the results of field observations in 2009–2011 (), while the other approach involved comparing the results produced by the NumoGen 1 and SimLep 1 models (). This method of validation required the models to be synchronised, which was achieved as follows. Simulations using the model SimLep1 were carried out first, and the date of egg-laying determined by that model was then used as input to NumoGen 1. The simulations were conducted using actual data recorded at Winna Góra in the period 2002–2011 and values obtained after transformation of the recorded data to reflect a temperature increase of 1–6°C.
Table I. Sample data set (year-month-day).
Table II. Parameters used in NumoGen 1 model.
Table III. Comparison of NumoGen 1 model results with field observations.
Table IV. Comparison of simulation results generated by NumoGen 1 and SimLep 1 on the basis of meteorological data registered at Winna Góra.
Results
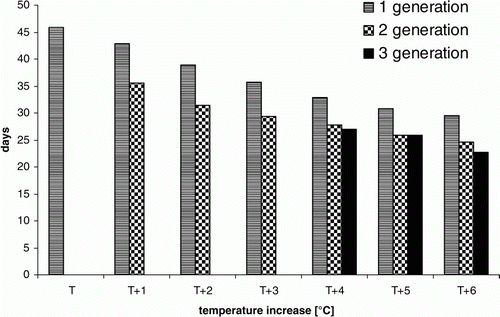
Comparison of results of simulations based on real data and the data obtained after raising the recorded temperature by 1–6°C confirmed the stimulating effect of temperature on the rate of beetle development. On the basis of real data, the number of days required for the development of the Colorado potato beetle first generation from egg to adult amounted on average to 46 days (). Raising the temperature by 1°C resulted in a shortening of the period by 3 days, to 43 days. Subsequent modifications involving an increase in temperature of 2, 3 and 4°C gave a total generation period of 39, 36 and 33 days, respectively, i.e., a decrease of 3–4 days per 1°C increase. Further increasing the temperature by 5 and 6°C gave a smaller (1 day/°C) decrease in the generation period, to 31 and 30 days, respectively. The relationship between temperature increase and development of the first generation of the beetle is expressed using the following equation:
Figure 1. Influence of temperature increase on the number of days needed to complete the first, second and third generations of Colorado potato beetle. T1–T6 refer to temperature increase of 1–6°C. The number of days expressed on the y-axis was achieved using the following equations: for first generation y=17.7572171454938 + exp(3.35092368334087+(−0.152325086952484)*x); x=temperature increase (1–6°C); for second generation y=21.07043775382 + exp(2.92595137352898+(−0.269914662094477)*x); x=temperature increase (1–6°C); for third generation y= − 497.148817944526 + exp(6.27873052566205+(−0.00411496277771)*x); x=temperature increase (4–6°C).
where y=number of days needed for the development of the first generation of the Colorado potato beetle and x=temperature increase (1–6°C).
Simulations performed on the basis of real data collected in the period 2002–2011 showed that the date of appearance of adult forms of the first generation ranged from 11 July to 5 August (). The observed dates of appearance of first-generation adult beetles in the period 1–28 July were used in calculations conducted after raising the temperature by 1°C. With an increase in temperature of 2 and 3°C, the dates of appearance of first-generation adults were 23 June–20 July and 17 June–12 July, respectively. The corresponding dates produced by simulations performed on data obtained after a real temperature increase of 4, 5 and 6°C were 13 June–6 July, 8 June–1 July and 6–28 June, respectively.
Table V. Date of appearance of first, second and third generation of Colorado potato beetle adults resulted in simulations using NumoGen 1 model.
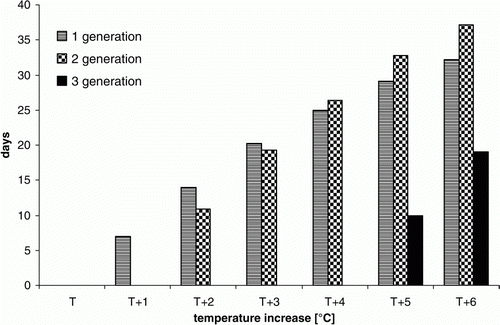
Thus the simulations showed that the average date of Colorado potato beetle adult appearance based on data recorded at Winna Góra was 21 July. Raising the temperature by 1°C resulted in an acceleration of the Colorado potato beetle first generation appearance by 7 days, while a simulated increase in temperature of 2 and 3°C gave an acceleration of 14 and 20 days. When the temperature was increased further by 4, 5 and 6 °C, the acceleration in appearance of first-generation adults amounted to 25, 29 and 32 days.
The relationship between temperature increase and acceleration of the first-generation adult appearance is expressed using the following equation:
where y=acceleration of the first-generation adult appearance (days) and x=temperature increase (1–6°C).
The calculations also revealed a close relationship between temperature and development of the second generation of Colorado potato beetle. The number of days required for the development of the second generation, estimated on the basis of simulations of a recorded temperature increase of 1°C, amounted to 36 days (). Raising the temperature by 2 and 3°C shortened the period until the development of the second beetle generation to 32 and 29 days, respectively. A temperature increase of 4, 5 and 6°C shortened the period required by a further 1–2 days, to 28, 26 and 25 days, respectively. The relationship between temperature increase and development of the second beetle generation is expressed using the following the equation:
where y=number of days needed for the development of the second generation of the Colorado potato beetle and x=temperature increase (1–6°C).
Raising the temperature also affected the date of appearance of the second-generation adults of Colorado potato beetle (). The results of calculations conducted on the basis of data obtained after a temperature increase of 1°C showed that the second-generation adults appeared in the period 10–24 August. When the temperature was increased by 2 and 3°C, the dates ranged from 1 to 25 August and 28 July to 16 August, respectively. As an effect of a 4, 5 and 6°C increase in temperature, the corresponding dates were 19 July–9 August, 12 July–4 August and 7–29 July, respectively. The average data of appearance date of the second-generation adults achieved in calculations when the temperature was increased by 1°C was 18 August. Raising the temperature by 2 and 3°C accelerated the appearance of Colorado potato beetle second-generation adults by 11 and 19 days, respectively (). As a result of data transformation consisting of a temperature elevation of 4, 5 and 6°C, the period for appearance of the second-generation adults was shortened to 26, 33 and 37 days, respectively.
Figure 2. Relationship between temperature increase and acceleration (expressed in days) of the appearance of adults of the first, second and third generation of Colorado potato beetle. T1–T6 refer to temperature increase of 1–6°C. The number of days expressed on the y-axis was achieved using the following equations: for first generation y= − 0.438095*x**2 + 8.05*x–0.240476; x=temperature increase (1–6°C); for second generation y= − 0.723214*x**2 + 12.45774*x+ − 11.5500; x=temperature increase (2–6°C); for third generation y= − 0.5*x**2 + 14.5*x−50.0; x=temperature increase (4–6°C).
The relationship between temperature increase and acceleration of the second-generation adult appearance is expressed using the following equation:
where y=acceleration of the second-generation adult appearance (days) and x=temperature increase (2–6°C).
The simulations also showed a relationship between temperature and development of a third generation of Colorado potato beetle. Calculations using the real and virtual data obtained after a temperature increase of 1–3°C showed that development of a third generation of the beetle was not possible but with an increase in temperature of at least 4°C, the simulations generated dates for appearance of third-generation adults. The average number of days required for the development of the third generation of the Colorado potato beetle after a temperature increase of 4, 5 and 6°C was 27, 26 and 23 days, respectively ().
The relationship between temperature increase and development of the third pest generation was as follows:
where y=number of days needed for the development of the third generation of the Colorado potato beetle and x=temperature increase (4–6°C).
Simulations performed with a 4, 5 and 6°C increase in mean temperature showed that the period of the third-generation beetle appearance ranged from 21 to 24 August, 17 to 24 August and 12 to 23 August, respectively (). The average date of appearance of the third generation adults following an increase in temperature of 4, 5 and 6°C was 23, 15 and 7 August, respectively. This means that the acceleration of appearance of the third-generation adult as a consequence of a temperature increase of 5 and 6°C in comparison with that achieved after a temperature increase by 4°C amounted to 10 and 19 days, respectively (). The relationship between temperature increase and acceleration of third-generation adult appearance was:
where y=acceleration of the third-generation adult appearance (days) and x=temperature increase (4–6°C).
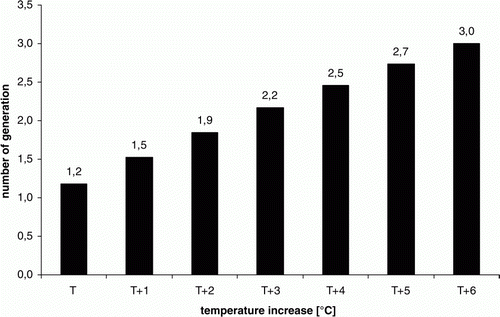
The analyses conducted using the model NumoGen 1 also allowed the impact of anticipated changes in climate conditions on the number of generations of the beetle to be estimated. Nine out of ten simulations performed on the basis of real data showed the appearance of only one generation of the beetle (). Following a simulated increase in temperature of 1°C, 60% of the results indicated the possibility of two generations of Colorado potato beetle. The remaining data enabled two generations as a result of increasing the temperature by 2°C. Third-generation appearance was conditioned by increasing the temperature recorded in 2002, 2003, 2006 and 2011 by 4°C. Similar results were achieved when the temperature recorded in the years 2005, 2007 and 2010 was increased by 5°C. However, the simulations using the meteorological data recorded in 2004 and 2009 generated the appearance of three pest generations only after raising the temperature by 6°C. The results of simulations allowed the relationship between temperature increase and the number of Colorado potato beetle generations to be determined as:
Table VI. Comparison of mean temperature in April–August with results of simulation using the NumoGen 1 model.
where y=number of Colorado potato beetle generations and x=temperature increase (1–6°C).
Equation Equation8 is demonstrated graphically in .
Figure 3. Influence of temperature increase on the number of Colorado potato beetle generations achieved in simulations using NumoGen 1. T1–T6 refer to temperature increase of 1–6°C. Numbers 1.2–3.0 refer to the number of Colorado potato beetle generations resulted in calculation using the following equation: y= − 0.008333*x*2 + 0.353571*x+1.17619, where y=number of Colorado potato beetle generations and x=temperature increase (1–6°C).
Discussion
Our computer simulations indicated a growing threat to potato from Colorado potato beetle. An increase of 1–2°C in actual temperature recorded at Winna Góra (52.2N; 17.4E) in recent years would create opportunities for the development of two full generations of the insect in the Wielkopolska region of Poland. An increase in temperature of at least 4°C could result in three generations. The results obtained corresponded well with the actual development of Colorado potato beetle recorded in regions characterised by a higher temperature than that in the Wielkopolska region. For example, in the regions of Campania in Italy and in Caucasus, where the mean temperature recorded in the period April–August amounts to 21 and 20.6°C, respectively, three generations of Colorado potato beetle occur. Two full generations and one partial generation of the pest also occur in North Carolina, where the mean temperature in the period April–August is 19.5°C. Our results dealing with the influence of temperature increase on the number of Colorado potato beetle generations confirm the findings by Kocmánková et al. (Citation2010) that Colorado potato beetle usually develops one generation in the Czech Republic but during particularly warm seasons lowland areas can experience two generations. The simulations performed by Kocmánková et al. (Citation2010) using Climex software tool predicted an increase in the number of Colorado potato beetle generations from one to even three for 2050 as a result of expected climate change in the Czech Republic. That study also confirmed the influence of temperature increase on widening the area occupied by Colorado potato beetle. An expansion of the area occupied by this pest was also reported by Baker et al. (Citation2000). It is known from previous research that temperature influences the speed of insect development time (Howe, Citation1960; Cammell & Knight, Citation1992), and the results from our study confirmed this for the case of development time of Colorado potato beetle, which decreased with increasing temperature. Similar results for the first generation of Colorado potato beetle were reported by Wittchen and Freier (Citation2008), who used the SimLep 1 model to study the effect of climate change on the population dynamics of the pest. Their results showed that a temperature increase of 2 and 3.7°C decreased the developmental time from egg to appearance of adults by 7 and 15 days, respectively. However, relating the development of Colorado potato beetle solely to temperature is a major simplification of reality. According to Harrington et al. (Citation2001), temperature does not act in isolation to influence pest status and interactions with other variables such as rainfall, humidity, irradiance and carbon dioxide concentrations should be taken into account. It is also known that insect developmental rate is a nonlinear function of temperature (Logan et al., Citation1976). A linear approximation for this function could result in underestimated development at low temperatures and overestimated development at high temperatures (Bergant et al., Citation2005). In addition, growth models of potato estimating the influence of climate change on plant ontogenesis would improve the credibility of the simulation results. However, despite the simplifications involved, the present study provides valuable information on the response of Colorado potato beetle to a temperature increase and according to some researchers, the effect of temperature on insects largely overwhelms the effects of other environmental factors (Bale et al., Citation2002). Faster developmental time and more generations of Colorado potato beetle as a result of expected temperature increase, as shown in this study, could lead to larger populations and consequently more damage to host plants (Bergant et al., Citation2005). However, such a scenario would only be possible in a situation of continuous availability of food for the insect. According to Haverkort (Citation2008), elongation of the vegetation season in the northern hemisphere will create conditions for earlier potato crops and for obtaining higher yields. A similar view was presented by Davies et al. (Citation1996), Peiris et al. (Citation1996) and Carter et al. (Citation1996), who forecast an increase in potato yields as a result of climate warming in England, Scotland and Finland, respectively. For Poland, an increase in the length of the vegetation period of between 30 and 40 days is projected for nearly all of the country as a result of temperature increase (Szwed et al., Citation2010). Kedziora (Citation1995) predicted an even greater extension of the vegetation period in Poland, by 51–125 days. The longer growing season would lead to increased pest populations and associated insecticide use. Therefore one possible impact of Colorado potato beetle response to climate change would be increased cost of potato production, which would increase the market price of potatoes in the long term. Moreover, repeated exposure to pesticide is the primary driver in the development of pest resistance, and resistance can develop rapidly in pests that have many generations per year, with multiple generations exposed to pesticide. Such a situation is expected to be common for Colorado potato beetle when the temperature increases. These findings indicate a need for sustained investment in integrated pest management and in breeding for potatoes with pest resistance, in addition to increased emphasis on breeding and selection for heat tolerance.
References
- Alyokhin , A. V. and Ferro , D. N. 1999 . Modifications in dispersal and oviposition of Bt-resistant and Bt-susceptible Colorado potato beetles as a result of exposure to Bacillus thuringiensis subsp. tenebrionis Cry3A toxin . Entomologia Experimentalis et Applicata , 90 : 93 – 101 . doi: 10.1046/j.1570-7458.1999.00426.x
- Amiri , A. , Talebi , A. A. , Zamani , A. A. & Kamali , K. 2010 . Effect of temperature on demographic parameters of the hawthorn red midget moth, Phyllonorycter corylifoliella, on apple . Accessed 1 February 2010, available at: http://www.insectscience.org/10.134/i1536-2442-10-134.pdf
- Anon . 1994 . Leptinotarsa decemlineata s 193–199 in Kwarantannowe Agrofagi Europy ( Warszawa 1069 ss: Inspektorat Kwarantanny Roślin) [Quarantine Pests for Europe (Warszawa 1069 ss:Inspectorate of Plants Quarantine)] .
- Baker , R. H. A. , Sansford , C. E. , Jarvis , C. H. , Cannon , R. J. C. , Macleod , A. and Walters , K. F. A. 2000 . The role of climatic mapping in predicting the potential geographical distribution of non-indigenous pests under current and future climates . Climates , 82 : 57 – 71 .
- Bale , J. S. , Masters , G. J. , Hodkinson , I. D. , Awmack , C. , Bezemer , T. M. , Brown , V. K. , Butterfield , J. , Buse , A. , Coulson , J. C. , Farrar , J. , Good , J. E. G. , Harrington , R. , Hartley , S. , Jones , T. H. , Lindroth , R. L. , Press , M. C. , Symrnioudis , I. , Watt , A. D. and Whittaker , J. B. 2002 . Herbivory in global climate change research: Direct effects of rising temperatures on insect herbivores . Global Change Biology , 8 : 1 – 16 . doi: 10.1046/j.1365-2486.2002.00451.x
- Bergant , K. , Trdan , S. , Znidarcic , D. , Crepinsek , Z. and Kajfez-Bogataj , L. K. 2005 . Impact of climate change on developmental dynamics of Thrips tabaci (Thysanoptera: hripidae): Can it be quantified? . Environmental Entomology , 34 : 755 – 766 . doi: 10.1603/0046-225X-34.4.755
- Cammell , M. and Knight , J. 1992 . Effects of climatic change on the population dynamics of crop pests . Advances in Ecological Research , 22 : 117 – 162 .
- Carter , T. R. , Saarikko , R. A. and Niemi , K. J. 1996 . Assessing the risks and uncertainties of regional crop potential under a changing climate in Finland . Agricultural and Food Science in Finland , 3 : 329 – 349 .
- Chen , S. , Fleischer , S. J. , Tobin , P. C. and Saunders , M. C. 2011 . Projecting insect voltinism under high and low greenhouse gas emission conditions . Environmental Entomology , 40 : 505 – 515 . doi: 10.1603/EN10099
- Collier , R. H. , Finch , S. , Phelps , K. and Thompson , A. R. 1991 . Possible impact of global warming on cabbage root fly (Delia radicum) activity in UK . Annals of Applied Biology , 118 : 261 – 271 . doi: 10.1111/j.1744-7348.1991.tb05627.x
- Cranshaw , W. S. and Radcliffe , E. B. 1980 . Effect of defoliation on yield of potatoes . Journal of Economic Entomology , 73 : 131 – 134 .
- Davies , A. , Jenkins , T. , Pike , A. , Shaq , J. , Carson , I. , Pollock , C. J. and Parry , M. I. 1996 . Modelling the predicted geographic and economic response of UK cropping systems to climate change scenarios: The case of potatoes . Aspects of Applied Biology , 45 : 63 – 69 .
- Deutsch , C. A. , Tewksbury , J. J. , Huey , R. B. , Sheldon , K. S. , Ghalambor , C. K. , Haark , D. C. and Martin , P. R. 2008 . Impacts of climate warming on terrestrial ectotherms across latitude . Proceedings of the National Academy of Sciences USA , 105 : 6668 – 6672 . doi: 10.1073/pnas.0709472105
- Ferrari , R. , Burgio , G. and Maini , S. 1993 . Comparison between preparations of Bacillus thuringiensis Berliner and insecticides, against Leptinotarsa decemlineata (Say) (Col. Chrysomelidae) in potato field . Bollettino dell'lstituto di Entomologia della Universita degli Studi di Bologna , 48 : 19 – 26 .
- Ferro , D. N. , Logan , J. A. , Voss , R. H. and Elkinton , J. S. 1985 . Colorado potato beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates . Environmental Entomology , 14 : 343 – 348 .
- Ferro , D. N. , Morzuch , B. J. and Margolies , D. 1983 . Crop loss assessment of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potatoes in western Massachusetts . Journal of Economic Entomology , 76 : 349 – 356 .
- Hare , D. J. 1980 . Impact of defoliation by the Colorado potato beetle on potato yields . Journal of Economic Entomology , 73 : 369 – 373 .
- Harrington , R. , Fleming , R. A. and Woiwod , P. 2001 . Climate change impacts on insect management and conservation in temperate regions: Can they be predicted? . Agricultural and Forest Entomology , 3 : 233 – 240 . doi: 10.1046/j.1461-9555.2001.00120.x
- Haverkort , A. 2008 . The influence of climate change on potato development, growth and biotic and abiotic factors . 17th Triennial Conference of the European Association for Potato Research, Abstract of Papers and Posters , July 6–10 , Brasov , Romania , 7 – 9
- Howe , R. 1960 . The effects of temperature and humidity on the rate of development and the mortality of Tribolium confusum Duval (Coleoptera. Tenebrionidae) . Annals of Applied Biology , 48 : 363 – 376 . doi: 10.1111/j.1744-7348.1960.tb03532.x
- Inouye , D. W. 2008 . Effects of climate change on phenology, frost damage and floral abundance of montane wildflowers . Ecology , 89 : 353 – 362 . doi: 10.1890/06-2128.1
- IPCC (Intergovernmental Panel on Climate Change) . 2007 . Summary for policymakers . In S. Solomon , D. Qin , M. Manning , Z. Chen , M. Marquis , K. B. Averyt , M. Tignor and H. L. Miller Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change Cambridge , , UK, and New York, NY : Cambridge University Press , pp. 1 – 18 . Accessed 20 August 2012, available at: http://ipcc-wg1.ucar.edu/wg1/Report/AR4WG1_Print_SPM.pdf
- Jörg , E. , Racca , P. , Preiß , U. , Butturini , A. , Schmiedl , J. and Wójtowicz , A. 2007 . Control of Colorado potato beetle with the SIMLEP decision support system . EPPO Bulletin , 37 : 353 – 358 . doi: 10.1111/j.1365-2338.2007.01137.x
- Kedziora , A. 1995 . Prognoza zmian klimatycznych [Climatic Change Prognosis; in Polish] . In S. Kozłowski Prognoza ostrzegawcza zmian srodowiskowych warunków życia człowieka w Polsce na poczatku XXI wieku [A Cautionary Prognosis Regarding Changes in the Environmental Living Conditions of Humans in Poland at the Beginning of the 21st Century; in Polish] , Zeszyty Naukowe Komitetu “Człowiek i środowisko”, 10, Oficyna Wydawnicza Instytutu Ekologii PAN , pp. 97 – 132 .
- Kocmánková , E. , Trnka , M. , Eitzinger , J. , Formayer , H. , Dubrovsk , M. , Semerádová , D. , Žalud , Z. , Juroch , J. and Možny , M. 2010 . Estimating the impact of climate change on the occurrence of selected pests in the Central European region . Climate Research , 44 : 95 – 105 . doi: 10.3354/cr00905
- Kurth , H. and Rossberg , D. 1986 . Kartoffelkäferprognose und der Grundlage eines imulationsmodells . Archiv für Phytopathologie und Pflanzenschutz , 22 : 65 – 77 . doi: 10.1080/03235408609436006
- Kurth , H. , Rossberg , D. and Köppen , H. 1984 . Ein Simulationsmodell der Populationsdynamik des Kartoffelkäfers. Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Konzeptionund ökologische Grunglagen von SimLep . Beiträge zur Entomologie , 34 : 159 – 166 . (in German)
- Łarczenko , K. 1958 . Długość rozwoju stonki ziemniaczanej w zależności od temperatur . Rocznik Nauk Rolniczych 78 seria A zeszyt , 1 : 27 – 42 . (in Polish)
- Linden , P. & Mitchell , J. F. B. 2009 . Climate change and its impacts: Summary of research and results from the ENSEMBLES project . FitzRoy Road , Exeter EX1 3PB , UK : Met Office Hadley Centre , p. 160 .
- Logan , J. A. , Wollkind , D. J. , Hoyt , S. C. and Tanigoshi , L. K. 1976 . An analytical model for description of temperature dependent rate phenomena in arthropods . Environmental Entomology , 5 : 1133 – 1140 .
- Mezeyová , I. , Šiška , B. & Mezey , J. 2008 . Potential infection pressure of codling moth (Laspeyresia = Cydia pomonella) as influenced by changing climate in the conditions of Slovak Republic . Proceedings 43rd Croatian and 3rd International Symposium on Agriculture , February 18–21 , Opatija , Croatia , 885 – 888 .
- Pawińska , M. and Mrówczyński , M. 2000 . Występowanie i zwalczanie stonki ziemniaczanej (Leptinotarsa decemlineata Say) w latach 1978–1999 . Progress in Plant Protection/Postęppy w Ochronie Roslin , 40 : 291 – 299 . (in Polish)
- Peiris , D. R. , Crawford , J. W. , Grashoff , C. , Jefferies , R. A. , Porter , J. R. and Marshall , B. 1996 . A simulation study of crop growth and development under climate change . Agricultural and Forest Meteorology , 79 : 271 – 287 . doi: 10.1016/0168-1923(95)02286-4
- Petzoldt , C. & Seaman , A. 2006 . Climate Change Effects on Insects and Pathogens. Climate Change and Agriculture: Promoting Practical and Profitable Responses Geneva : New York State Agricultural Extension Station III . pp. 1 – 16 .
- Porter , J. H. , Parry , M. L. and Carter , T. R. 1991 . The potential effects of climatic change on agricultural insect pests . Agricultural and Forest Meteorology , 57 : 221 – 240 . doi: 10.1016/0168-1923(91)90088-8
- Przybysz , E. , Pawińska , M. and Węgorek , P. 1996 . Monitoring odporności stonki ziemniaczanej na niektóre stosowane w Polsce insektycydy . Progress in Plant Protection/Postępy w Ochronie Roślin , 36 : 338 – 342 . (in Polish)
- Sidorczyk , S. 1965 . Przyczynek do badań nad metodyką obliczania strat powodowanych przez stonkę ziemniaczaną (Leptinotarsa decemlineata Say) . Biuletyn IOR nr , 30 : 13 – 25 . (in Polish)
- Šmatas , R. , Semaškienė , R. and Lazauskas , S. 2008 . The impact of climate conditions on the occurrence of the Colorado potato beetle (Leptinotarsa decemlineata) . Žemdirbystė = Agriculture , 95 : 235 – 241 .
- Szwed , M. , Karg , G. , Pińskwar , I. , Radziejewski , M. , Graczyk , D. , Kędziora , A. & Kundzewicz , Z. W. 2010 . Climate change and its effect on agriculture. Water resources and human health sectors in Poland . Accessed 20 August 2012, available at: http://www.nat-hazards-earth-syst-ci.net/10/1725/2010/nhess-10-1725-2010.pdf
- Tauber , M. , Tauber , C. , Obrycki , J. , Gollands , B. and Wright , R. 1988 . Voltinism and the induction of aestival diapause in the colorado potato beetle Leptinotarsa decemlineata (Coleoptera: chrysomelidae) . Annals of the Entomological Society of America , 81 : 748 – 754 .
- Ungerer , M. J. , Ayres , M. P. and Lombardero , M. J. 1999 . Climate and the northern distribution limits of Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae) . Journal of Biogeography , 26 : 1133 – 1145 . doi: 10.1046/j.1365-2699.1999.00363.x
- Węgorek , W. 1959 . The Colorado potato beetle (Leptinotarsa decemlineata Say) . Prace Naukowe Instytutu Ochrrony Roślin 1 , 1 – 105 ( in Polish ).
- Węgorek , P. , Mrówczyński , M. & Wachowiak , H. 2001 . Current status of resistance in Colorado potato beetle (Leptinotarsa decemlineata Say) in Poland ( Rothamsted: Materialy Konferencji Resistance 2001 Meeting the Callange 46 ss ).
- Wittchen , U. and Freier , B. 2008 . Nutzung von Datenreihen und simulationsmodellen zur Abschätzung des Einflusses des Klimawandels auf die Befallsentwicklung von Schadorganismen in landwirtschaftlichen Pflanzenbeständen Nachrichtenbl . Nachrichtenblatt des Deutschen Pflanzenschutzdienstes , 60 : 157 – 162 . (in German)
- Wójtowicz , A. & Jörg , E. 2006 . Ocena wiarygodności i przydatności komputerowej symulacji rozwoju populacji stonki ziemniaczanej (Leptinotarsa decemlineata) prowadzonej za pomocą systemu SimLep 3 . Biuletyn IHAR 242 , 217 – 224 ( in Polish ).