Abstract
Cloudberry (Rubus chamaemorus) is a dioecious perennial shrub growing on peat land. The berries are a combined fruit with a color ranging from yellow to orange and bright red. Berries are highly valued in the Nordic countries and berries with orange to red color are preferred. Cultivation and breeding has started, and four Norwegian varieties are on the market. These clones were selected on the basis of good shoot production (both sexes), good pollinators (males), and big berries (females). Recently, berry quality has been introduced in breeding programs as a trait for selection. In this study we present the variation in levels of individual anthocyanins and hydroxycinnamic derivates between different clones of cloudberry. Both levels of anthocyanins and hydroxycinnamic derivates varied considerably between the different clones. The most abundant anthocyanin was cyanidin 3-glucoside, followed by cyanidin 3-rutinoside. One single clone (AN1372) from Andøya in Northern Norway had the highest levels of all the analyzed anthocyanins. Among the two analyzed hydroxycinnamic acids, p-coumaric acid presented the highest ratio in all analyzed berries. Differences between different years were relatively high, but not consistent. Levels of caffeic acid did not vary much between clones. The results indicate a genetic basis for the variation between clones, but since numbers of samples were few and sampling only were done during two growth seasons, further analysis will be needed to examine the variation and genetic basis of anthocyanin composition of cloudberry.
Introduction
Cloudberries (Rubus chamaemorus L.) are regarded as a delicacy in the Nordic countries. Berries are consumed fresh or in different products as jams, juices, yoghurts, liquors, and sweets. Ripe berries are combined fruits consisting of several drupes, each with one stony seed. The color range from yellow to orange and red, and the berry flesh is soft, high in vitamin C (Walker et al., Citation2006) and different polyphenols (Martinussen et al., Citation2010; McDougall et al., Citation2011; Uleberg et al., 2011). The phenolic composition of cloudberry is dominated by ellagitannins and flavonols with varying but low amounts of anthocyanins (Kähkönen et al., Citation2001, Citation2012; Määttä-Riihinen et al., Citation2004; Heinonen, Citation2007).
Anthocyanins, responsible for the desirable red/orange coloring of cloudberries, are water-soluble glycosides and acylglycosides of anthocyanidins, and belong to the flavonoid class of polyphenols (Clifford, Citation2000). Cloudberry anthocyanins are all cyanidin derivates acilated with glucose, rutinose, sophorose, and glucosylrutinose. Cyanidin derivatives, generally found as cyanidin 3-O-glucoside, are the major anthocyanins in many berry species as black elderberry, black chokeberry, blackberry, red raspberry, and lingonberry (Määttä-Riihinen et al., Citation2004).
Häkkinen et al. (Citation1999) analyzed phenolic compounds in many berry species, among them cloudberry. In cloudberry ellagic acid is the main phenolic compound and contributes up to 75–80% of the total phenolics. The rest of the phenolic profile constitute of hydroxycinnamic acids (p-coumaric acid, caffeic acid, and ferulic acid) (Määttä-Riihinen et al., Citation2004), flavonols (quercetin, kaempferol, and myricetin) and other hydroxybenzoic acids (p-Hydroxybenzoic acid and gallic acid). Of these, p-coumaric acid (8.5%) and gallic acid (6%) had the highest levels in the study of Häkkinen et al. (Citation1999).
Numerous studies have suggested that secondary plant polyphenolics have positive effects on human health. They have been found to have antiallergenic (Yamaura et al, Citation2011), anti-inflammatory (Wang et al., Citation1999), antiviral (Krawitz et al., Citation2011), antiproliferative (Zikri et al., Citation2009), antioxidative activity (Rice-Evans et al., Citation1996; Robards et al., Citation1999), and anticarcinogenic effects (Wang & Stoner, Citation2008). High levels of polyphenols have, therefore, become valuable traits in the breeding of high quality fruits (Diamanti et al., Citation2011) and are now incorporated in several breeding programs; e.g. strawberry (Capocasa et al., Citation2008), raspberry (Khanizadeh et al., Citation2009), blueberries (Lee et al., Citation2004), and cloudberry (Uleberg et al., Citation2011).
It has been shown that clones that produce red berries have a higher content of total anthocyanins (Martinussen et al., Citation2010). In a previous publication Uleberg et al. (Citation2011) presented an evaluation of a clone collection of cloudberry for the characters total anthocyanins, total phenols, and total carotenoids. Here we present levels of individual anthocyanins and hydroxycinnamic acids for the same clone collection.
The aim of this work has been to investigate the variation in individual anthocyanins and hydroxycinnamic acids in the genotypes and to evaluate the potential of targeting individual compounds in the breeding program for cloudberry.
Material and methods
Plant material
The evaluations were performed at Bioforsk Nord Holt in Tromsø (latitude 69°65′, altitude 30 m above sea level) and have been previously described by Uleberg et al. (Citation2011). In brief, the different clones were planted on peat in open benches outdoors, each one in separated squares. Every clone was represented in two replicate squares at random positions in the bench. Bench 1 had square sizes of 1 m2, while bench 2 had square sizes of 0.8 m2. In both benches each square had 16 plants. Plants were established in natural Sphagnum peat (H2-H4, dry substance ca 60 kg/m3, pH 3.0–4.0) from Fyrkløver. Complete fertilizer (14-6-16) was mixed with the peat (300 g per 150 l peat) at establishment. Subsequent years there were no fertilization. The plants have been irrigated if needed. Bench 1 consisted of 12 clones (3 male and 9 female) collected in Northern Norway (11 in Nordland County and one in Finnmark County) (). Bench 2 consisted of clones (1 male and 11 female) collected from four different Norwegian counties; two in Northern Norway (Finnmark and Nordland), and two in Southern Norway (Aust Agder and Hedmark) (). In addition, one clone was collected in England and one at Svalbard. Further details can be found in Uleberg et al. (Citation2011).
Table I. Origin and sex of the evaluated cloudberry clones.
Sample preparation
Berries were harvested when they reached the right softness and could easily be removed from the sepals. Berry color will vary from yellow to orange and red dependent on the clone. Frozen berry samples (−80°C), between 5 and 10 individual berries in a 15 ml tube, were weighted and freeze dried in VirTis BenchTop 2K freeze drier (SP Scientific, USA) for 3 days at 7 mTorr pressure. Dry samples were weighted again and grinded. Approximately 100 mg of powder was mixed with 5 ml extraction solvent (methanol [Sigma-Aldrich Chemie GmbH, Germany]: water = 80:20; v/v).
Individual anthocyanin determination
Before analysis of the anthocyanins, the extracts were diluted with 1% trifluoroacetic acid (TFA, Sigma, Germany) in water by using 1:1 (v:v) ratio. Samples were diluted with acidified water in order to maintain the symmetry of chromatographic peaks. Prior to the injection the samples were filtered through 45 µm Millipore HPLC filter. All analyses were made in duplicates. The analytical method applied have been previously described by Trošt et al. (Citation2008), slightly modified for the purpose of these analyses. Separation and quantification of cloudberry anthocyanins: cyanidin 3-sophoroside (Cy 3-Sop), cyanidin 3-(2G-glucosylrutinoside) (Cy 3-GluRut), cyanidin 3-glucoside (Cy 3-Glu) and cyanidin 3-rutinoside (Cy 3-Rut) were performed using gradient high performance liquid chromatography with the diode array detection (DAD) at 520 nm. Analyses were made with Waters Alliance chromatographic system (Waters Corporation, USA). Detection was made using 2998 Photodiode Array detector (Waters Corporation, USA). Individual anthocyanins were separated on Nova-Pak Column (C 18, 3.9×150 mm; Waters, Milford, MA, USA), quantified as cyanidin 3-glucoside (k=53,173, R2 = 99.94%, DL = 0.01 mg/l, QL= 0.3 mg/l) and identified by standard retention time and previously described data (Määttä-Riihinen et al., Citation2004). The samples were stable for at least 48 hours. In order to determine method repeatability the same sample was injected six times. The relative standard deviations of observed peak areas for individual anthocyanins were: Cy 3-Sop, 4.2%; Cy 3-GluRut, 5.0%; Cy 3-Glu, 5.0%; and Cy 3-Rut, 2.9%. Analyses were performed at room temperature. Injection volume was 20 µl. For anthocyanin separation gradient of mobile phases was used. Mobile phase A was composed from water. Mobile phase B was composed from acetonitrile and water 60:40 (v/v). Both mobile phases were acidified with 0.2 vol% TFA (Sigma, Germany). The gradient of mobile phase B changed from 10% to 25% in 40 min. In the next minute the percentage of mobile phase B increased from 25% to 100%. Afterwards gradient was steady for 4 minutes. In the end equilibration to initial concentration was established. The flow rate through the gradient of 0.7 ml/min was used. Average relative standard deviation for the sum of anthocyanins was 10.5%.
Hydroxycinnamic acid determination
Hydroxycinnamic acids were determined in the same run as anthocyanins using the same chromatographic system and conditions. Hydroxycinnamic acids were quantified as p-coumaric acid (k=177,168, R2 = 100%, DL = 0.01 mg/l, QL = 0.25 mg/l) and identified by standard retention time and previously described data (Määttä-Riihinen et al., Citation2004). In order to determine method repeatability the same sample was injected six times. The relative standard deviations of observed peak areas for individual anthocyanins were: p-coumaric acid, 0.6%; caffeic acid, 1.0). Average relative standard deviation for the sum of hydroxycinnamic acids was 8.6%.
Statistical method
Statistical analyses were performed using the General Linear Model of Minitab 15 in a model that included clone, year, and replicate. Bench 1 and Bench 2 were analyzed separately.
Results
In Bench 1 there was a statistical significant effect of clone on the levels of Cy 3-Sop (p=0.012) and in Bench 2 there was an effect of replicate on the levels of Cy 3-Glu (p=0.005), Cy 3-Rut (p=0.013) and Sum Anthocyanins (p=0.014). No other significant effects were found and in general there was large variation both between and within clones and year ( and ). There was a difference between the benches, as Bench 1 had the highest contents for all analyzed compounds except for caffeic acid that had equal levels in both benches. The clones in the benches had different ages and there was no genetic links between the benches, thus the background for the observed differences could not be established.
Figure 1. Individual anthocyanins and hydroxycinnamic acid derivates analyzed in cloudberries expressed as mg/100 g FW of berries. Results are means of samples from year 2006 and 2007 from Bench 1.
Abbreviations: cyanidin 3-sophoroside (Cy 3-Sop), cyanidin 3-(2G-glucosylrutinoside) (Cy 3-GluRut), cyanidin 3-glucoside (Cy 3-Glu), cyanidin 3-rutinoside (Cy 3-Rut) and hydroxy cinnamic acids (HC).
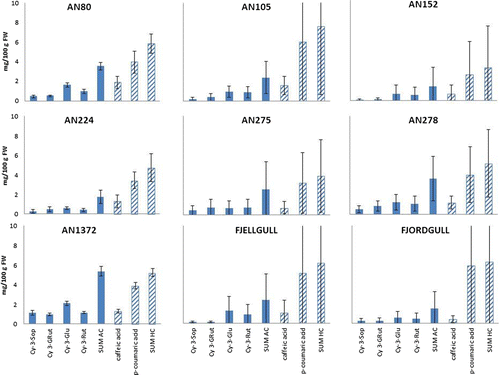
Figure 2. Individual anthocyanins and hydroxycinnamic acid derivates analyzed in cloudberries expressed as mg/100 g FW of berries. Results are means of samples from year 2006 and 2007 from Bench 2.
Abbreviations: cyanidin 3-sophoroside (Cy 3-Sop), cyanidin 3-(2G-glucosylrutinoside) (Cy 3-GluRut), cyanidin 3-glucoside (Cy 3-Glu), cyanidin 3-rutinoside (Cy 3-Rut) and hydroxycinnamic acids (HC).
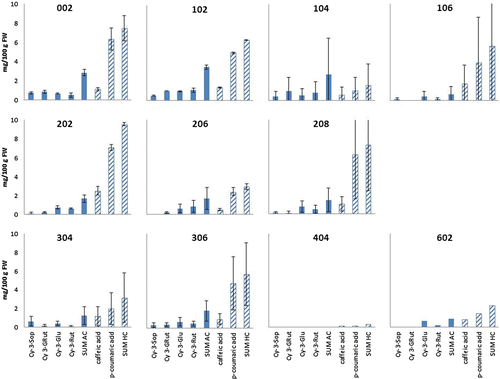
In this study the highest sum of individual anthocyanins were present in AN1372 with 5.37 mg/100 g FW of berries while the lowest levels of 0.62 mg/100 g FW were found in berries of clone 106 ( and ). The most abundant pigment presenting red anthocyanin color was cyanidin 3-glucoside, followed by cyanidin 3-rutinoside. Clone AN1372 had highest levels of all clones for all the analyzed single anthocyanins, but the only significant effect was found for levels of Cy 3-Sop, where AN1372 had significant higher levels than the other clones in the bench.
Among the two analyzed hydroxycinnamic acids, p-coumaric acid presented the highest ratio ( and ). Differences between different years were relatively high, but not consistent. Highest levels of hydroxycinnamic acids were present in clone 202, with 9.5 mg/100 g FW. Other clones with hydroxycinnamic acid concentrations above 7 mg/100 g FW were AN105 and clones 002 and 208 (). Levels of caffeic acid varied from 0.2 mg/100 g FW in clone 404 to 2.5 mg/100 g FW in clone 106. There were no differences in caffeic acid content between the two clone collections with a mean value of 1.1 mg/100 g FW for both benches.
All analyzed compounds were positively correlated, with correlations ranging from 0.126 to 0.781. Correlation between sum of anthocyanins and sum of hydroxycinnamic acids were 0.42.
Discussion
In this study we could not establish statistically significant differences in contents of individual anthocyanins and hydroxycinnamic derivates in the investigated clones. Unfortunately, there were quite few available samples for analysis. About 8 out of 20 included clones are represented with two samples both harvesting years, while 9 clones were sampled only one year. Thus it was difficult to distinguish the effect of year from the genetic differences. The average temperature during fruit ripening (June, July, and August) was 10.8°C first harvesting year (2006) and 11.8°C the second year (2007). Previous studies have shown effects of temperature on the levels of anthocyanins (Martinussen et al., Citation2010; McDougall et al., Citation2011).
Bench 1 was established in 1998 while Bench 2 was established in 2003. In addition, there were no genetic links between the benches and, therefore, not possible to test for differences between the benches. Bench 1 did, however, show higher levels of all analyzed contents. The differences were especially evident for the sum of anthocyanins, where the average value was about 65% higher in Bench 1 () than in Bench 2 (). For the sum of hydroxycinnamic acids the difference was small, the average value in Bench 1 was about 6% higher than in Bench 2. Jaakola et al. (Citation2012) found that the main factors affecting chemical composition of cloudberry were the climatic conditions, while Martinussen et al. (Citation2010) and Uleberg et al. (Citation2011) have shown clonal differences in anthocyanin contents, but not in phenolic contents. Several studies on bilberries indicate that northern plants produce berries with higher anthocyanin concentrations than southern plants (Martinelli et al., Citation1986; Lätti et al., Citation2008, Citation2010, Åkerström et al., Citation2010). The higher levels of anthocyanins in Bench 1 could be explained by the fact that the clones were all collected from Northern Norway, while the clones in Bench 2 are from a much broader geographical region (). In previous publication (Uleberg et al., Citation2011) total anthocyanins was analyzed from the same samples as used in the present study for analysis of single anthocyanins. Fjellgull had the highest levels of total anthocyanins followed by clone 104 and clone AN1372 and AN278. These results are in contradiction to the present study were clone AN1372 had the highest levels of all analyzed anthocyanins. The amounts of total anthocyanins in AN1372 reported in Uleberg et al. (Citation2011) were exactly the same as the sum of the four anthocyanins presented here. Fjellgull, 104 and AN278 had much higher levels in the previous analyses on total anthocyanins and the sum of the four anthocyanins that was analyzed in the present study represented only 35%, 42%, and 65% of the total anthocyanins in Fjellgull, 104 and AN278, respectively. This might indicate that these four single anthocyanins represent the main anthocyanin contents of AN1372 while other anthocyanins are higher represented in some of the other clones. However, these four anthocyanins have been shown to be the most abundant in cloudberry (Määttä-Riihinen et al., Citation2004).
The tested clones varied in content of p-coumaric acid from 0.16 to 7.08 mg/100 g FW of berries. Määttä-Riihinen et al. (Citation2004) reported the content of p-coumaric acid to be 43 mg/kg fresh weight of berries which is 60% of the level found in clone 202. This variation is consistent with the findings of Häkkinen (Citation2000), who found relatively large differences in contents of hydroxycinnamic acids between cultivars of strawberry (p-coumaric acid) and blueberry (caffeic acid).
Our results show that the different clones have berries with considerable variation in the content of anthocyanins and hydroxycinnamic acid derivates, but numbers of samples are relatively few and the sampling has only been done during two growth seasons. Further analysis will be needed to examine the variation and genetic basis of anthocyanin composition of cloudberry.
References
- Åkerström , A. , Jaakola , L. , Bång , U. and Jäderlund , A. 2010 . Effects of latitude-related factors and geographical origin on anthocyanidin concentrations in fruits of Vaccinium myrtillus L. (Bilberries) . Journal of Agricultural and Food Chemistry , 58 : 11939 – 11945 . doi: 10.1021/jf102407n
- Capocasa , F. , Scalzo , J. , Mezzetti , B. and Battino , M. 2008 . Volum Combining quality and antioxidant attributes in the strawberry: The role of genotype . Food Chemistry , 111 : 872 – 878 . doi: 10.1016/j.foodchem.2008.04.068
- Clifford , M. N. 2000 . Anthocyanins – Nature, occurrence and dietary burden . Journal of the Science of Food and Agriculture , 80 : 1063 – 1072 . doi: 10.1002/(SICI)1097-0010(20000515)80:7%3C1063::AID-JSFA605%3E3.0.CO;2-Q
- Diamanti , J. , Battino , M. & Mezzetti , B. 2011 . Breeding for Fruit Nutritional and Nutraceutical Quality . In M. A. Jenks and P. J. Bebeli Breeding for Fruit Quality Hoboken , NJ : John Wiley & Sons, Inc , doi: 10.1002/9780470959350 .
- Häkkinen , S. 2000 . Flavonols and phenolic acids in berries and berry products . Doctoral thesis , Kuopio University Publications D. Medical Sciences 221. ISBN 951–781–801–7 .
- Häkkinen , S. , Heinonen , M. , Kärenlampi , S. , Mykkänen , H. , Ruuskanen , J. and Törrönen , R. 1999 . Screening of selected flavonoids and phenolic acids in 19 berries . Food Research International , 32 : 345 – 353 . doi: 10.1016/S0963-9969(99)00095-2
- Heinonen , M. 2007 . Antioxidant activity and antimicrobial effect of berry phenolics – A Finnish perspecitive . Molecular Nutrition and Food Research , 51 : 684 – 691 . doi: 10.1002/mnfr.200700006
- Jaakola , M. , Korpelainen , V , Hoppula , K. and Virtanen , V. 2012 . Chemical composition of ripe fruits of Rubus chamaemorus L. grown in different habitats . Journal of the Science of Food and Agriculture , 92 : 1324 – 1330 . doi: 10.1002/jsfa.4705
- Kähkönen , M. P. , Hopia , A. I. and Heinonen , M. 2001 . Berry phenolics and their antioxidant activity . Journal of Agricultural and Food Chemistry , 49 : 4076 – 4082 . doi: 10.1021/jf010152t
- Kähkönen , M. , Kylli , P. , Ollilainen , V. , Salminen , J.-P. and Heinonen , M. 2012 . Antioxidant activity of isolated ellagitannins from red raspberries and cloudberries . Journal of Agricultural and Food Chemistry , 60 : 1167 – 1174 . doi: 10.1021/jf203431g
- Khanizadeh , S. , Rekika , D. , Ehsani-Moghaddam , B. , Tsao , R. , Yang , R. , Charles , M. T. , Sullivan , J. A. , Gauthier , L. , Gosselin , A. , Potel , A. M. , Reynaud , G. and Thomas , E. 2009 . Horticultural characteristics and chemical composition of advanced raspberry lines from Quebec and Ontario . LWT – Food Science and Technology , 42 : 893 – 898 . doi: 10.1016/j.lwt.2008.08.016
- Krawitz , C. , Mraheil , M. A. , Stein , M. , Imirzalioglu , C. , Domann , E. , Pleschka , S. and Hain , T. 2011 . Inhibitory activity of a standardized elderberry liquid extract against clinically-relevant human respiratory bacterial pathogens and influenza A and B viruses . BMC Complementary and Alternative Medicine , 11 : 16 doi: 10.1186/1472-6882-11-16
- Lätti , A. K. , Riihinen , K. R. and Kainulainen , P. S. 2008 . Analysis of anthocyanin variation in wild populations of bilberry (Vaccinium myrtillus L.) in Finland . Journal of Agricultural and Food Chemistry , 56 : 190 – 196 . doi: 10.1021/jf072857m
- Lätti , A. K. , Jaakola , L. , Riihinen , K. R. and Kainulainen , P. S. 2010 . Anthocyanin and flavonol variation in bog bilberries (Vaccinium uliginosum L.) . Journal of Agricultural and Food Chemistry , 58 : 427 – 433 . doi: 10.1021/jf903033m
- Lee , J. , Finn , C. E. and Wrolstad , R. E. 2004 . Anthocyanin pigment and total phenolic content of three vaccinium species native to the Pacific Northwest of North America . HortScience , 39 : 959 – 964 .
- Määttä-Riihinen , K. R. , Kamal-Eldin , A. and Törrönen , A. R. 2004 . Identification and quantification of phenolic compounds in berries of Fragaria and Rubus species (Family Rosaceae) . Journal of Acricultural and Food Chemistry , 52 : 6178 – 6187 . doi: 10.1021/jf049450r
- Martinelli , E. M. , Baj , A. and Bombardelli , E. 1986 . Computer-aided evaluation of liquid-chromatographic profiles for anthocyanins in Vaccinium myrtillus fruits . Analytica Chimica Acta , 191 : 275 – 281 . doi: 10.1016/S0003-2670(00)86314-X
- Martinussen , I. , Uleberg , E. , McDougall , G. , Stewart , D. and Junttila , O. 2010 . Development and quality of cloudberry (Rubus chamaemorus L.) as affected by female parent, male parent and temperatur . Journal of Berry Research , 1 : 91 – 101 .
- McDougall , G. , Martinussen , I. , Junttila , O. , Verrall , S. and Stewart , D. 2011 . Assessing the influence of genotype and temperature on polyphenol composition in cloudberry (Rubus chamaemorus L.) using a novel mass spectrometric Me . Journal of Agricultural and Food Chemistry , 59 : 10860 – 10868 . doi: 10.1021/jf202083b
- Rice-Evans , C. A. , Miller , N. J. and Paganga , G. 1996 . Structure-antioxidant activity relationships of flavonoids and phenolic acids . Free Radical Biology and Medicine , 20 : 933 – 956 . doi: 10.1016/0891-5849(95)02227-9
- Robards , K. , Prenzler , P. D. , Tucker , G. , Swatsitang , P. and Glover , W. 1999 . Phenolic compounds and their role in oxidative processes in fruits . Food Chemistry , 66 : 401 – 436 . doi: 10.1016/S0308-8146(99)00093-X
- Trošt , K. , Golc-Wondra , A. , Prošek , M. and Milivojevicì , L. 2008 . Anthocyanin degradation of blueberry – Aronia nectar in glass compared with carton during storage . Journal of Food Science , 73 : 405 – 411 .
- Uleberg , E. , Trost , K. , Stavang , J. E. , Røthe , G. and Martinussen , I. 2011 . Evaluation of cloudberry (Rubus chamaemorus L.) clones for selection of high-quality varieties . Acta Agriculturae Scandinavica Section B – Soil and Plant Science , 61 : 18 – 26 . doi: 10.1080/09064710.2011.599330
- Walker , P. G. , Gordon , S. L. , Brennan , R. M. and Hancock , R. D. 2006 . A high-throughput monolithic HPLC method for rapid vitamin C phenotyping of berry fruit . Phytochemical Analysis , 17 : 284 – 290 . doi: 10.1002/pca.916
- Wang , H. , Nair , M. G. , Strasburg , G. M. , Chang , Y. C. , Booren , A. M. , Gray , J. I. and DeWitt , D. L. 1999 . Antioxidant and antiinflammatory activities of anthocyanins and their aglycon, cyanidin, from tart cherries . Journal of Natural Products , 62 : 294 – 296 . doi: 10.1021/np980501m
- Wang , L. S. and Stoner , G. D. 2008 . Anthocyanins and their role in cancer prevention . Cancer Letters , 269 : 281 – 290 . doi: 10.1016/j.canlet.2008.05.020
- Yamaura , K. , Shimada , M. and Ueno , K. 2011 . Anthocyanins from bilberry (Vaccinium myrtillus L.) alleviate pruritus in a mouse model of chronic allergic contact dermatitis . Pharmacognosy Research , 3 : 173 – 177 . doi: 10.4103/0974-8490.85002
- Zikri , N. N. , Riedl , K. M. , Wang , L. S. , Lechner , J. F. , Schwartz , S. J. and Stoner , G. D. 2009 . Black raspberry components inhibit proliferation, induce apoptosis and modulate gene expression in rat esophageal epithelial cells . Nutrition and Cancer , 61 : 816 – 826 . doi: 10.1080/01635580903285148