
Abstract
The revitalisation of wild vegetables by consuming and cultivating them in home gardens is one of the strategies that agriculturalists have recently identified to combat food insecurity. However, data on mineral uptake with maturity are not available for a larger majority of wild vegetables. S. nigrum, one of the popular wild vegetables consumed in the Eastern Cape Province of South Africa, was cultivated both on the field and glasshouse to gain a better understanding of the uptake of some macrominerals as the plant matures. Five treatments (control; 100 kg N/ha; 8.13 t manure/ha; 100 kg N/ha + 8.13 t manure/ha and 50 kg N/ha + 4.07 t manure/ha) were set up in a Randomised Complete Block Design in both trials. The data were subjected to analysis of variance using MINITAB statistical software package. K (%) ranged between 3.11–6.98 in the glasshouse and 3.88–6.98 on the field; Ca (%) between 1.23–3.72 in the glasshouse and 1.39–3.98 on the field; Mg (%) between 0.31–0.93 in the glasshouse and 0.48–0.82 on the field; P (mg/kg) between 0.23–0.80 in the glasshouse and 0.29–0.77 on the field and Na (mg/kg) ranged between 749–3070 in the glasshouse and 187–3070 on the field. The application of 50 kg N/ha + 4.07 t manure/ha increased the uptake of a majority of the minerals. K, P and Na decreased with maturity while Ca increased and Mg varied. Although the current results indicate that for K, P and Na nutritional interventions, the leaves of S. nigrum are best harvested during the early phase of growth and the final phase for Ca, the concentration of these minerals has the potential to supply sufficient quantities of the minerals in human physiology at all stages of the plant's growth. This wild vegetable is therefore recommended for both consumption and cultivation.
Introduction
Wild vegetables have recently received a renewed interest in their growth patterns, nutrient compositions, medicinal properties as well as antioxidant activities. Among other reasons, this has been necessitated by the ever increasing incidences of hunger, malnutrition and disease prevalence especially among the poor and marginalised communities in the developing world.
Although Solanum nigrum is a well-known nutraceutical plant, this wild vegetable is also highly endowed with nutritive substances which are essential in human physiology. A majority of authors have reported on the nutritional composition of the mature Solanum species. For example, Edmonds and Chweya (Citation1997), Akubugwo et al. (Citation2007), Sarma and Sarma (Citation2011) as well as Gbile and Adesina (Citation1988) studied the nutritional composition of S. nigrum at maturity and found it to be favourable. Sarma and Sarma (Citation2011) further studied various phytochemical compositions of S. nigrum while Arulmozhi et al. (Citation2010) concentrated on the antioxidant and antihyperlipidemic properties of this plant.
Data on the uptake of macrominerals in S. nigrum at various stages of maturity are inadequate. Fortuin and Omta (Citation1980) analysed leaf and berry yield of S. nigrum from the 8th to the 20th week after transplanting. However, these previous authors did not analyse the nutritional composition. In the present study, the growth and physiological response of S. nigrum to fertilisers at various stages of maturity were initially analysed and found to be positive (Bvenura & Afolayan Citation2013, Citation2014).
Plants generally respond differently to soil amendments. For example, Singh and Singh (Citation1939) reported that some members of the Amaranthaceae family readily absorb K and some N while P is uniformly absorbed. However, in their study, it was noted that maximum absorption was in the pre-flowering stage. In studies conducted on other species, Flyman and Afolayan (Citation2008) investigated the effect of plant maturity on the mineral composition of Momordica balsamina as well as Vigna unguiculata. Atta et al. (Citation2010) studied mineral concentration in Hibiscus Sabdariffa at different stages of growth. These authors were able to determine the peak stage of harvest of each wild vegetable based on mineral requirements.
The need for food and the supply of macrominerals in human physiology at all stages of life cannot be overemphasised. In order to combat the problem of hunger and malnutrition, Faber and Wenhold (Citation2007) suggested the cultivation of wild vegetables in home gardens as one of the strategies that can be implemented. Based on its popularity among the Eastern Cape rural populace, S. nigrum was selected and targeted for domestication trials. The effect of organic and/or inorganic fertilisers on macromineral (Ca, K, Mg, Na, P) uptake per growth stage were investigated with the aim to determine the peak time of harvest of each mineral and the best fertiliser amendment.
Materials and methods
Experimental site
The trials were conducted in the glasshouse and field at the University of Fort Hare, South Africa, located at 32° 47′ S and 26° 50′ E and 535 m above sea level. The area is within a semi-arid ecological zone with an average annual rainfall of approximately 575 mm in summer; mean daily temperatures of 22.5°C during the day and 18.8°C at night while during the winter the temperature is about 13.6°C during the day and less than 10.3°C at night (Marais & Brutsch Citation1994). According to the Soil Classification Working Group, Soil and Irrigation Research Institute (South Africa) (Citation1991), the soils are deep alluvial of the Oakleaf form, belong to the Jozini series and are texturally sandy loam. The properties of the soil used for this experiment are shown in .
Table 1. The chemical properties of the experimental soil (upper 0–30 cm depth) and organic fertilizer.
Experimental material
Ripe and mature S. nigrum berries were harvested between the 3 April 2012 and 26 April 2012 from the wild in Alice. The seeds were separated from the pulp, washed in distilled water and dried at room temperature on the laboratory bench for 2 h and kept in sealed bottles until further use. In both trials, the extracted seeds were planted in cavity trays in the glasshouse and later transplanted when they were about 6 weeks old and with at least 6 true leaves and about 10–15 cm tall. For the glasshouse trial, the seedlings were transplanted into prepared polythene bags containing 5 kg of 2 mm sieved soil. The soil used in the glasshouse was obtained from the field where the field trial was conducted so that the properties of the soils for the two experiments would be almost similar. The organic fertiliser (goat manure) used in this experiment was obtained from the University of Fort Hare animal farm while the inorganic fertilisers (NPK [2:3:4] and limestone ammonium nitrate [LAN]) were purchased from a local fertiliser dealer. The properties of the organic fertiliser used for this experiment are shown in .
Treatments
The experiments were laid out in a Randomised Complete Block Design with five treatments and five replicates. The treatments were: control (T1); 100 kg N/ha (T2); 8.13 t manure/ha (T3); 100 kg N/ha + 8.13 t manure/ha (T4) and 50 kg N/ha + 4.07 t manure/ha (T5). Nitrogen was supplied in the form of NPK and LAN fertilisers. Organic fertiliser (goat manure) and NPK were applied at transplanting and LAN fertiliser applied 4 weeks after transplanting. These fertilisers were applied in the top 5–7 cm of soil depth by mixing with a spade in plots measuring 3 m × 2 m on the field. In the glasshouse, each treatment had 5 replicates, but each replicate had 10 experimental units to ensure a sufficient number of plant samples for the trial period since the destructive method of data collection was used. Each replicate consisted of one S. nigrum plant in a polythene bag containing 5 kg of soil.
Data collection
The third youngest fully expanded leaves (Jones & Case Citation1990) were collected from the shoots by uprooting the whole plant; washed in distilled water to remove sediments and other impurities before drying the samples in a dust free, forced-draft oven at 40°C to a constant weight. The samples were then ground to a powder using a mortar and pestle and passed through a 2-mm sieve. The samples were kept in plastic val containers and stored in a refrigerator at 4°C till when needed. The first data were collected on the day of transplanting (week 0 on Figures of Results) followed by 3 weeks after transplanting, after which data were collected on a weekly basis. The experiment was terminated in the 9th week for the glasshouse experiment and 12th week on the field when all the berries on the plant were mature and ripe.
Mineral analysis
About 0.5 g of finely ground vegetable samples was placed in dry, clean digestion tubes and 5 ml digestion mixture comprising 1 part HCIO4 + 2 parts HNO3 added. This mixture was digested at 230°C on a digestion block for 70 min, allowed to cool down and made up to 100 ml volume with distilled water. The concentration of Ca, K, Mg, Na and P was then determined using the Inductively Coupled Plasma – Optical Emission Spectrometer (ICP OES; AGRILASA Citation2008).
Statistical analysis
Data of the macromineral concentrations of various treatments were subjected to statistical analysis using MNITAB Release 12 statistical software package. A one way analysis of variance was used to compare the means of various macromineral concentrations among the treatments and a two way analysis of variance used to determine the interaction between plant age (weeks after transplanting) and treatment on macromineral accumulation in the plant. Means were segregated using Duncan's multiple range test. The means were treated as significantly different at p < 0.05.
Results and discussion
Calcium (Ca)
On the field, Ca increased exponentially between the time of transplanting and the 3rd week and varied until the 11th week and exponentially increased again in the 12th week (). The treatment means differed significantly (p < 0.05) and ranged between 1.39 and 3.98% at the time of transplanting and 12th week respectively. The means for the duration of the trial were highest in T1 (2.54%) and lowest in T2 (2.37%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Ca. Regression analysis with Ca as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 58.2% indicating that plant age had a fairly significant effect on Ca. The glasshouse treatment means ranged between 1.23 and 3.72% in the 4th and 9th weeks respectively (). The means for the duration of the trial were lower than those from the field and were highest in T2 (2.02%) and lowest in T5 (1.57%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Ca. Regression analysis with Ca as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 12.7% indicating that plant age had a minimum effect on Ca.
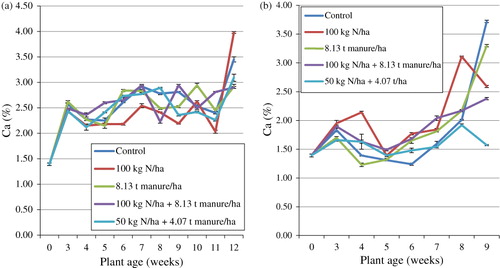
The results of Ca concentration are in line with the observation of Khader and Rama (Citation2003). However, they were at variance with what was reported by Flyman and Afolayan (Citation2008) as well as Mahala et al. (Citation2012). Ca increased from the early stages to the final stages of growth in the present study. This might be due to the immobile nature of the mineral and its failure to translocate from older parts of the plant to the growing and flowering parts as is the tendency of minerals in plants (Loneragen Citation1968). The accumulation of Ca in more mature plants than the younger plants was also reported by Loneragen and Snowball (Citation1969). The results of the current study showed that the leaves of S. nigrum from both the field and glasshouse studies accumulate sufficient Ca that could supplement human diet according to the New Zealand and Australian standards (NHMRC Citation2005). This study further reveals that the best time to harvest S. nigrum leaves for Ca requirements is in the final stages of the plant's growth life cycle. Comparing values reported by Odhav et al. (Citation2007) for wild uncultivated S. nigrum and the values reported in the present trial, the cultivated species clearly possesses a higher nutritional concentration than the wild uncultivated species.
Magnesium (Mg)
Mg increased from the time of transplanting to the 4th week after which it decreased and increased again in the 10th week and declined between the 11th and 12th weeks (). The least and highest values of Mg (0.48 and 0.82%) were respectively recorded in the 12th and 10th weeks. Treatment means were significantly different (p < 0.05) throughout the trial and the means for the duration of the trial were highest in T1 (0.72%) and least in T2 (0.66%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Mg. Regression analysis with Mg as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 53.10% indicating that plant age had a fairly significant effect on Mg. In the glasshouse, the concentration of Mg varied between 0.31 and 0.93% in the 7th and 4th weeks respectively and decreased from the time of transplanting to the 9th week (). The treatment means differed significantly throughout the trial and the means for the duration of the trial were highest in T2 (0.69%) and lowest in T1 (0.49%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Mg. Regression analysis with Mg as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 77.6% indicating that plant age had a significant effect on Mg. In comparison with the field trial, the glasshouse means were lower than those from the field trial.
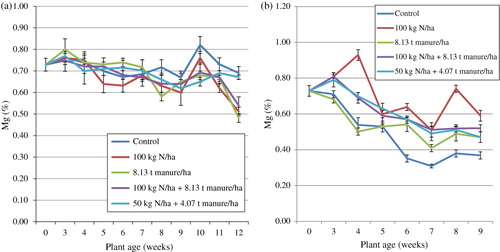
Results of the present study show appreciable amounts of Mg in both the field and glasshouse treatments. This is in line with the work of Flyman and Afolayan (Citation2008). They reported a range of 0.38 and 0.90% in V. unguiculata and 0.35 and 1.45 (%) in M. balsamina leaves. Also, in India, Khader and Rama (Citation2003) reported very high values of Mg between 22.33 and 80.33% in 6 wild vegetable species. Comparing the soil concentration of Mg () and the results of the current study, it is clear that minimal amounts of Mg were absorbed. Some authors argue that Mg is subject to leaching in considerable amounts or may be mined from the soil through harvesting of plant products (Romheld & Kirkby Citation2007; Gransee & Fuhrs Citation2013). While it has been reported that organic matter increases Mg content of soils and considerably prevents leaching of the mineral, this was not particularly true for S. nigrum in the present trial. Soil type, availability of other nutrients, rainfall as well as timing and crop species are some of the factors that influence the actual availability of Mg for plant absorption (Gransee & Fuhrs Citation2013). It is therefore inferable that any of these factors could have contributed to the low uptake of the mineral in comparison with soil and organic matter content of the present study. Notwithstanding this, Mg in the leaves of S. nigrum observed in the present study at all stages of growth has the potential to supply human diet with recommended daily intake values according to the New Zealand and Australian standards (NHMRC Citation2005).
Phosphorus (P)
Phosphorus accumulation decreased as S. nigrum matured and was highest in the 5th week and lowest in the 12th week (). The treatment means differed significantly (p < 0.05) throughout the trial and the means for the duration of the trial were highest in T2 (0.62%) and lowest in T4 (0.56%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on P. Regression analysis with P as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 81.3% indicating that plant age had a very significant effect on P. The glasshouse treatment means differed significantly (p < 0.05) and showed a decreasing trend from the time of transplanting to the 9th week (). The means for the duration of the trial were highest in T5 (0.63%) and lowest in T1 (0.40%). These were lower than those recorded on the field with the exception of T5. The treatment means ranged between 0.23 and 0.80% in the 7th and 3rd weeks respectively. Statistical analysis showed an interaction between plant age and the fertiliser treatment on P. Regression analysis with P as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R 2) of 92.6% indicating that plant age had a very significant effect on P.
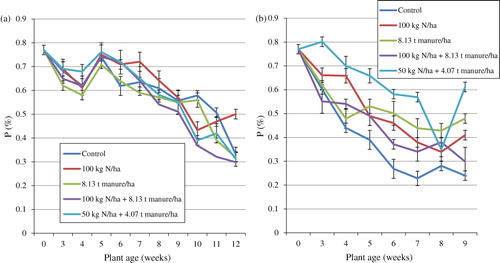
Concentrations of P in the leaves of S. nigrum observed in the present study are almost similar to those reported by Flyman and Afolayan (Citation2008). They reported a range between 0.19 and 0.51% in V. unguiculata and 0.15 and 0.53 (%) in M. balsamina leaves while in the present study, the range was between 0.23 and 0.80%. The findings of this study are also in agreement with work by Khader and Rama (Citation2003) who reported a decrease in P concentration in some wild vegetables with advancing age. Although total amount of P in the soil may be high, it is often immobile and up to 80% is available in organic form and this is not readily available to the plant for uptake (Schachtman et al. Citation1998). This may be responsible for the mineral's low concentrations in S. nigrum. In addition, P is known to be most abundant in actively growing tissues of young plants and this possibly explains the high concentration of the element in the first stages of growth. The results of this study indicate that harvesting the plant at its early growth stages would be suitable for P requirements. Also, these results further indicate that cultivated S. nigrum has the ability to supply the daily recommended concentrations of P at all stages of its growth according to the New Zealand and Australian standards for mineral intake (NHMRC Citation2005).
Potassium (K)
Potassium decreased on the field from the time of transplanting to the final week of observation (). The treatment means significantly differed (p < 0.05) and ranged between 3.88 and 6.98% between the 12th and 4th weeks respectively. The means for the duration of the trial were highest in T5 (5.25%) and lowest in T3 (4.97%); however, the mineral was at its peak in the 4th week in T2 (6.98%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on K. Regression analysis with K as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R 2) of 63.6% indicating that plant age had a significant effect on K. In the glasshouse, K showed a variable trend from the time of transplanting to the final week of observation but was at its peak in the final week of observation in T5 (). Treatment means differed significantly throughout the trial and the means for the duration of the trial were highest in T5 (6.20%) and least in T1 (4.08%). Statistical analysis showed an interaction between plant age and the fertiliser treatment on K. Regression analysis with K as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 83.6% indicating that plant age had a significantly strong effect on K. Comparing the field and glasshouse trials, higher values were recorded in T1–T3 on the field whereas higher values were recorded in T3 and T4 in the glasshouse.
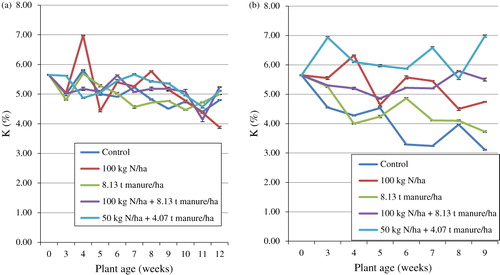
The results of the present study are comparable with those of Flyman and Afolayan (Citation2008). In their report, K concentration decreased with maturity in V. unguiculata while in M. balsamina, they increased up to the third week and started to decrease. The values obtained by these authors are however lower than what is reported in the current study. Because of the decrease in K content with plant maturity on the field, it would be ideal to harvest S. nigrum in its early stages of growth and 50 kg N/ha + 4.07 t manure/ha would be the most favourable fertiliser if the mineral of interest is K. Potassium is relatively immobile in the soil and unavailable in dry soil conditions especially when moisture is low; but available in soils with a high clay content and a pH between 6 and 7 (Potassium uptake Citation1998). The results of the current study further indicate that there is more than sufficient K in cultivated S. nigrum to supply human daily requirements according to the New Zealand and Australian standards (NHMRC Citation2005). The current results also indicate that harvesting the leaves of the plant at any age for consumption will potentially ensure an adequate supply of the mineral to the body per day.
Sodium (Na)
The concentration of Na in S. nigrum cultivated on the field exponentially decreased between the time of transplanting and the 3rd week and varied but was lowest in the 12th week (). The treatment means differed significantly (p < 0.05) and ranged between 187 and 1534.5 mg/kg in the 12th week and time of transplanting, respectively. Means for the duration of the trial were highest in T5 (721.7 mg/kg) and least in T4 (635.3 mg/kg). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Na. Regression analysis with Na as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 68.8% indicating that plant age had a significant effect on Na. In the glasshouse, Na decreased and the highest value was recorded at the time of transplanting and the lowest in the ninth week (). The means for the duration of the trial were highest in T4 (1287.2 mg/kg) and lowest in T3 (1071.2 mg/kg). Statistical analysis showed an interaction between plant age and the fertiliser treatment on Na. Regression analysis with Na as the dependable variable and time (plant age) as the regressor showed a coefficient of determination (R2) of 34.6% indicating that plant age had a minimal effect on Na. The glasshouse treatment shows an increase between 51 and 66%.
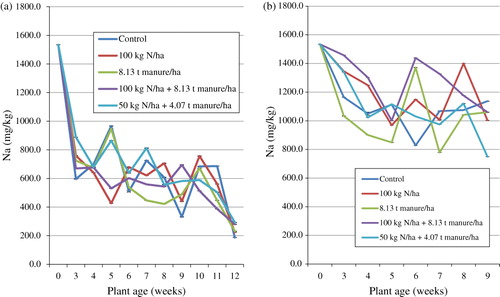
The trend for the accumulation of Na in S. nigrum leaves in the present study is in contrast with the trend reported for V. unguiculata leaves (Flyman & Afolayan Citation2008). These authors reported that Na concentration in V. unguiculata increased up to 4 weeks after transplanting, after which it started to decrease. Also, the current results from the field study were lower than those reported by Mahala et al. (Citation2012) who observed an increase in Na concentration with plant maturity. It has been observed that the extent to which Na is taken up by plants varies with species and is influenced by other minerals present in the soil especially K and N. Due to the chemical similarity between K and Na, competition for common absorption sites in the roots may lead most plant species to readily absorb K instead of Na even though the concentration of Na may be high in the soil (Smith et al. Citation1980). Observation from the present study shows that S. nigrum readily absorbed K as compared to Na. The K:Na ratio in the glasshouse ranged between 43.91 and 59.62 while on the field it was between 72.74 and 80.75, indicating the natrophobic nature of the plant. The findings of this work indicate that Na accumulation in the leaves of S. nigrum may not be sufficient at some stages to take care of recommended daily intake of Na according to the New Zealand and Australian standards for mineral intake (NHMRC Citation2005). However, harvesting the leaves during the early stages of growth may be ideal for Na requirements since Na content is at its peak during this time.
From this investigation, it has been revealed that S. nigrum is highly endowed with macrominerals and can be cultivated and incorporated into the diets of poor rural communities whose diets are mainly starch based, both for nutritional purposes and for diet fortification. The best fertiliser to apply and best time to harvest the plant leaves for food is best determined based on specific nutritional interventions. For example, K, P and Na seemingly decrease in concentration with maturity while Ca increases with maturity and Mg decreases and increases at a later phase. Therefore, S. nigrum is conclusively best harvested at the early stages of growth for K, P and Na and to harness Ca, the wild vegetable is best harvested at the late stages of growth while for Mg; both the early and late phases of growth ensure maximum concentrations. It was also observed that there was a significant interaction between the plant age and treatment on the accumulation of all the minerals. Plant age therefore had an effect on macromineral uptake in S. nigrum. These results further indicate that on the field, 100 kg N/ha increased P uptake and 50 kg N/ha + 4.07 t manure/ha increased the uptake of K and Na while Mg and Ca were highest in the control. In the glasshouse, 100 kg N/ha increased the uptake of Ca and Mg while 50 kg N/ha + 4.07 t manure/ha increased the uptake of K and P and 100 kg N/ha + 8.13 t manure/ha increased the uptake of Na. Therefore, the application of 50 kg N/ha + 4.07 t manure/ha increased the uptake of the majority of minerals in both the field and glasshouse. In general, the nutrient values recorded indicate the ability of the wild vegetable to supply the molarity of recommended daily mineral intakes of both the macro and microminerals at all stages of the plant's growth.
Funding
The authors are grateful to the Govan Mbeki Research and Development Center, University of Fort Hare, South Africa for funding this research.
Additional information
Funding
References
- AGRILASA. 2008. Method 6.1.2: wet ashing. In: Palic C, editor. Plant and feed analysis handbook. Pretoria, (South Africa): Agri Laboratory Association of Southern Africa; Unpublished manuscript.
- Akubugwo IE, Obasi AN, Ginika SC. 2007. Nutritional potential of the leaves and seeds of black nightshade-Solanum nigrum L. Var virginicum from Afikpo-Nigeria. Pak. J. Nutr. 6:232–326. 10.3923/pjn.2007.323.326
- Arulmozhi V, Krishnaveni M, Karthishwaran K, Dhamodharan G, Mirunalini S. 2010. Antioxidant and antihyperlipidemic effect of Solanum nigrum fruit extract on the experimental model against chronic ethanol toxicity. Pharmacogn Mag. 6:42–50. 10.4103/0973-1296.59965
- Atta S, Diallo AB, Bakasso Y, Sarr B, Saadou M, Glew RH. 2010. Micro-element contents in Roselle (Hibiscus sabdariffa) at different growth stages. Afr J Food Agric Nutr Dev. 10:2615–2628.
- Bvenura C, Afolayan AJ. 2013. Growth and physiological response to organic and/or inorganic fertilisers of wild Solanum nigrum L. cultivated under field conditions in Eastern Cape Province, South Africa. Acta Agric Scand Sect B Soil Plant Sci. 63:683–693.
- Bvenura C, Afolayan AJ. 2014. Growth and physiological response of Solanum nigrum L. to organic and/or inorganic fertilisers. J Appl Bot Food Qual. 87:168–174.
- Edmonds JM, Chweya JA. 1997. Black Nightshade. Solanum nigrum L. and related species. Promoting the conservation and use of underutilised and neglected crops. 15. Rome: Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute.
- Faber M, Wenhold F. 2007. Nutrition in contemporary South Africa. Water SA. 33:393–400.
- Flyman MV, Afolayan AJ. 2008. Effect of plant maturity on the mineral content of the leaves of Momordica balsamina L. and Vigna unguiculata subsp. J Food Qual. 31:661–671. 10.1111/j.1745-4557.2008.00218.x
- Fortuin FTJM, Omta SWP. 1980. Growth analysis and shade experiment with Solanum nigrum L., the black nightshade, a leaf and fruit vegetable in West Java. Neth J Agr Sci. 28:199–210.
- Gbile ZO, Adesina SK. 1988. Nigerian Solanum species of economic importance. Ann Missouri Bot Gard. 75:862–865. 10.2307/2399374
- Gransee A, Fuhrs H. 2013. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil. 368:5–21. 10.1007/s11104-012-1567-y
- Jones Jr JB, Case VW. 1990. Sampling, handling and analyzing plant tissue samples. In Westerman RL, editor. Soil testing and plant analysis. 3rd ed. Madison (WI): Soil Science Society of America Inc; p. 390–420.
- Khader V, Rama S. 2003. Effect of maturity on macromineral content of selected leafy vegetables. Asia Pac J Clin Nutr. 12:45–49.
- Loneragen JF. 1968. Nutrient requirements of plants. Nature. 220:1307–1308. 10.1038/2201307a0
- Loneragen JF, Snowball K. 1969. Calcium requirements of plants. Aust J Agr Res. 20:465–478. 10.1071/AR9690465
- Mahala AG, Amasiab SO, Monera A, Elsadig A. 2012. Effect of plant age on DM yield and nutritive value of some leguminous plants (Cyamopsis tetragonoloba, Lablab purpureus and Clitoria (Clitoria ternatea). Inter Res J Agri Sci S Sci. 2:502–508.
- Marais JN, Brutsch MO. 1994. The Ehlers system of assessing the suitability of temperature regime of a region for crop production. Paper presented at 1994 SASHS Congress, Nelspruit, South Africa.
- NHMRC. 2005. Nutrient reference values for Australia and New Zealand: Including recommended dietary intakes. Canberra: Australia.
- Odhav B, Beekrum S, Akula U, Baijnath H. 2007. Preliminary assessment of nutritional value of traditional leafy vegetables in KwaZulu-Natal, South Africa. J Food Comp Anal. 20:430–435. 10.1016/j.jfca.2006.04.015
- 1998. Potassium uptake and availability. Better Crops. 82:14–15.
- Romheld V, Kirkby EA. 2007. Magnesium functions in crop nutrition and yield. Proceedings of the Conference of the International Fertilizer Society; 2007 Dec 7; Cambridge: York-International Fertlizer Society.
- Sarma H, Sarma A. 2011. Solanum nigrum L., a nutraceutical enriched herb or invasive weed? International Conference on Environmental and BioScience, IPCBEE. Singapore: IACSIT Press.
- Schachtman DP, Reid RJ, Ayling SM. 1998. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 116:447–453. 10.1104/pp.116.2.447
- Singh BN, Singh LB. 1939. Relative absorption of nutrients by weeds of arable land. Soil Sci. 47:227–235. 10.1097/00010694-193903000-00007
- Smith GS, Middleton KR, Edmonds AS. 1980. Sodium nutrition of pasture plants.1. translocation of sodium and potassium in relation to transpiration rates. New Phytologist. 84:603–612. 10.1111/j.1469-8137.1980.tb04774.x
- Soil Classification Working Group, Soil and Irrigation Research Institute (South Africa). 1991. Soil classification: a taxonomic system for South Africa. A report on a research project conducted under the auspices of the Soil and Irrigation Research Institute. 2nd ed. Pretoria: Department of Agricultural Development.